Remote-Sensing Hydraulic Characterization of Channel Habitat Units in a Tropical Montane River: Bladen River, Belize


Department of Geography, University of Alabama, Box 870322, Tuscaloosa, AL 35487-0322, USA
*
Author to whom correspondence should be addressed.
Remote Sens. 2017, 9(12), 1295; https://doi.org/10.3390/rs9121295
Submission received: 9 November 2017
/
Revised: 30 November 2017
/
Accepted: 8 December 2017
/
Published: 12 December 2017
(This article belongs to the Special Issue Application of Remote Sensing in Hydrological Modeling and Watershed Management)
Abstract
:The physical characteristics of river systems exert significant control on the habitat for aquatic species, including the distribution of in-stream channel habitat units. Most previous studies on channel habitat units have focused on midlatitude rivers, which differ in several substantive ways from tropical rivers. Field delineation of channel habitat units is especially challenging in tropical rivers, many of which are remote and difficult to access. Here, we developed an approach for delineating channel habitat units based on a combination of field measurements, remote sensing, and hydraulic modeling, and applied it to a 4.1-km segment of the Bladen River in southern Belize. We found that the most prevalent channel habitat unit on the study segment was runs, followed by pools and riffles. Average spacing of channel habitat units was up to twice as high on the study segment than the typical values reported for midlatitude rivers, possibly because of high erosion rates in the tropical environment. The approach developed here can be applied to other rivers to build understanding of the controls on and spatial distribution of channel habitat units on tropical rivers and to support river management and conservation goals.
1. Introduction
Physical hydraulic and geomorphic characteristics of river systems serve as important controls on the habitat for aquatic species. Characterization of these physical habitat variables is important for describing and comparing different river systems, examining changes in habitat resulting from natural or human-caused disturbances, and developing and implementing conservation and restoration goals. To aid in these objectives, numerous channel habitat unit classification systems have been developed. Many of these systems are based on hydraulic and geomorphic variables. For example, Frissell et al. [1] developed a hierarchical classification for stream habitats, with channel habitat units classified on the basis of reach class, bed topography, water surface slope, morphogenetic structure or process, substrate, and bank configuration. Hawkins et al. [2] developed a channel habitat unit classification system based on a division between fast water (which is further subdivided into rough and smooth) and slow water (which is further subdivided into scour pools and dammed pools). Montgomery and Buffington [3] classified mountain channels based on typical bed material, bedform pattern, dominant roughness elements, dominant sediment sources, sediment storage elements, typical confinement, and typical pool spacing. Thomson et al. [4] used the River Styles framework to classify channel habitat units according to planform, channel geometry, and textural controls. A common feature of hydraulic and geomorphic classifications is that they are based primarily on physical variables such as slope, substrate, and width-to-depth ratio.
Other channel habitat unit classification approaches relate species assemblages to physical habitat characteristics. For example, Buffagni et al. [5] used macroinvertebrate species assemblages to identify channel habitat units, resulting in classes of margin with macrophytes, margin without macrophytes, backwater, run-riffle, and macrophytes in the flow, based on flow velocity, depth, Froude number, granulometric composition and substratum roughness, and cover. Newson and Newson [6] proposed using a “habitat hydraulics” approach to channel habitat unit classification, based on flow types and physical biotopes analogous to mesohabitats and functional habitats in ecology. In these ecological approaches, the species assemblages are classified first, and the physical variables that control their spatial patterns are determined afterwards.
Regardless of whether channel habitat units are classified using geomorphic or ecological approaches, the role of classification in meeting conservation goals is similar. Many aquatic species have specific physical habitat requirements, which may differ for different stages of their life cycles. A diversity of channel habitat units is therefore typically recognized as being conducive to healthy aquatic ecosystems. For example, Crispin et al. [7] found that restoration of large woody debris resulted in a fivefold increase in summer habitat for coho salmon and a sixfold increase in winter habitat through the creation of additional pool-and-riffle channel habitat units. Quinn and Peterson [8] also found that wild juvenile coho salmon survival rates were strongly correlated with complexity of channel habitat units. Angermeier and Karr [9] found that all fish feeding guilds other than aquatic insectivores were most concentrated in deep pools in streams in Panama. Willis et al. [10] analyzed fish assemblages in a Venezuelan river and found that species density was negatively associated with flow velocity and positively associated with habitat complexity. Montaña and Winemiller [11] found that cichlid species partitioned habitat at the local scale, based on features such as leaf litter, sand banks, rocky shoals, and woody debris. Delineation of channel habitat units is an important preliminary step in assessing the baseline condition of a river system and in setting goals for conservation and restoration.
Despite the importance of being able to delineate channel habitat units, the process is not always straightforward. Channel habitat units are often delineated by visual assessment during a field visit, which is problematic for two reasons. First, visual assessments can be subjective, even for well-defined channel habitat units such as pools, riffles, and runs. Roper and Scarnecchia [12] determined that there was significant variability in how trained observers classified channel habitat units, especially when there was a large number of habitat unit types to choose from, although the variability decreased with a higher level of training. To alleviate the problem of observer subjectivity, some researchers have proposed ecohydraulic channel habitat unit classifications, such as those based on velocity/depth ratio, Froude number, and slope [13]. Such ecohydraulic approaches make for more objective and robust classifications.
A more fundamental limitation of the surveying-based approach to channel habitat unit delineation is that it requires costly and time-consuming fieldwork. This limitation can be addressed through the use of remote sensing and hydraulic modeling. With some field data available for validation, it is possible to use remotely-sensed imagery to estimate some hydraulic variables needed for ecohydraulic characterization of channel habitat units (i.e., depth). Remotely-sensed data can in turn be used to initialize hydraulic models that can simulate additional variables needed for classification (i.e., velocity). A combination of minimal field measurements, remote sensing, and hydraulic modeling can be used to characterize channel habitat units for longer segments of river than would be feasible to survey manually, or to delineate channel habitat units for remote regions that are difficult to access.
Tropical rivers are especially rich targets for remotely-sensed channel habitat unit classifications, because they are often relatively difficult to access and lack extensive observed data. In part because of the difficulty of fieldwork on tropical rivers, these systems are generally under-studied in comparison with midlatitude rivers, for which the vast majority of channel habitat unit classification systems were developed. Although both midlatitude and tropical rivers are highly variable, there are some common characteristics of tropical rivers that make them different enough that ideas regarding channel habitat units on midlatitude rivers may not necessarily apply [14]. For example, tropical climates are the wettest in the world, so tropical rivers have high rates of discharge per unit area. Despite overall high rates of precipitation and discharge, many tropical climates are also highly variable in their hydroclimatic regime, with distinct wet and dry seasons, and often large interannual variability resulting from oceanic-atmospheric phenomena such as the El Niño-Southern Oscillation. These pronounced differences in precipitation between wet and dry periods result in large variations in discharge. During high flows, sediment transport capacity is large, resulting in significant bedload sediment transport. Sediment supply is likely to be large, because the warm temperatures and abundant moisture mean that the tropics have the highest weathering rates in the world. Decomposition rates are also high, so large woody debris does not persist in channels for as long as in midlatitude rivers, although the supply of new large woody debris is large because of the dense vegetation. Except at the highest elevations, tropical river basins were never glaciated and can therefore generally be expected to be better-graded than many higher-latitude rivers that are still adjusting to the retreat of Pleistocene ice sheets. In general—because of the high rates of discharge and sediment transport, lack of stabilized large woody debris, and history of no glaciation—tropical rivers could reasonably be expected to adjust their channels more rapidly to changes in external drivers, compared to many midlatitude rivers.
In this paper, we develop a method for delineating channel habitat units from remotely-sensed imagery and hydraulic modeling, and apply the method to a tropical montane river. We also examine the spatial patterns of delineated channel habitat units on the study river and compare them to those that would be expected based on studies of midlatitude rivers. The method described here is novel, and this study is also among the first that focuses on channel habitat units of tropical rivers.
2. Materials and Methods
2.1. Study Area
The study area is the Bladen River in southern Belize, which joins with the Swasey Branch to form the Monkey River approximately 16 km upstream from the Caribbean Sea at Monkey River Town (Figure 1a and Table 1). The total drainage area of the Monkey River is 1291 km2, and the basin relief is 1011 m. The Bladen River originates in the Maya Mountains, a highly eroded mountain range resulting from tectonic uplift and granitic intrusion during the Triassic Period. During the Cretaceous period, the region was inundated by seawater and fossiliferous limestones were deposited. The Monkey River Basin is dominated by intrusive igneous and volcanic lithology to the north and limestone to the south. Land cover in the basin is dominated by evergreen broadleaf forest (89%), with some cropland (7%, mostly banana and citrus groves in the lower basin), savannah (2%), and mixed forest (1%). The climate of the region is tropical, with mean annual temperature of 25 °C and mean annual precipitation of 406 cm.
Within the Bladen River, our study site was located at the Belize Foundation for Research and Environmental Education (BFREE) field station and in the adjacent Bladen Nature Reserve, approximately 37 km inland (Figure 1b). This particular site was selected because the Bladen is considered one of the most biodiverse and ecologically intact areas in Mesoamerica [15]. Because of its sheltered location west of the karst hills, the Bladen region is protected from hurricane landfalls, which allows for undisturbed mature forest. The Bladen is connected to other protected areas in Belize, Guatemala, and Mexico, forming one of the largest remaining blocks of intact forest in Central America. In addition to the numerous terrestrial species of concern that find habitat in the basin, the Bladen River itself provides quality habitat for many aquatic species, including Astyanax aeneus (banded tetra), Vieja maculicauda (blackbelt cichlid), Cichlasoma salvini (yellow-belly cichlid), Thorichthys meeki (firemouth cichlid), Archocentrus spilurum (blue-eye cichlid), and Lontra longicaudis (Neotropical river otter). Nevertheless, aquatic species in the Bladen River are potentially imperiled by expanding agriculture downstream of the protected area, which could affect the water quality for migratory species traveling between the Bladen and the Caribbean, and by the encroachment of invasive Oreochromis spp. (African tilapia). The relatively undisturbed nature of the Bladen Nature Reserve made it an ideal site to study channel habitat units, and a better understanding of the spatial distribution of habitat units can assist reserve managers in targeting priority areas for conservation and restoration. An additional reason for selecting the Bladen Nature Reserve study site was the resources made available by BFREE (field access, logistical support, high-resolution aerial imagery).
2.2. Data Sources
The overall goal of this project was to characterize channel habitat units for the Bladen River. For an approximately 700-m reach immediately upstream of the BFREE field station (“Reach 1”), we made field measurements of bed elevation, water depth, and flow velocity for a related project that modeled the physical controls on cichlid nesting sites in the Bladen River (Figure 2). For the same project, we made additional measurements of depth and velocity (but not bed elevation) for a discontinuous 600-m reach further upstream (“Reach 2”). We considered this limited amount of field data collection to be insufficient for the purpose of delineating channel habitat units that are representative of the entire Bladen River. We did, however, also have access to a high-resolution (10-cm) aerial image of a 4.1-km segment of the Bladen River within the Bladen Nature Reserve, which includes Reach 1 and Reach 2 as well as an addition 2.7 km of river upstream of the BFREE field station that we were unable to survey (“upstream segment”). The objective was to use a combination of field measurements, remote sensing, and hydraulic modeling to characterize channel habitat units for the entire 4.1 km of the Bladen River covered in the aerial image.
Figure 3 shows the overall process of measuring, calculating, and estimating the variables needed for characterizing channel habitat units. The main variables needed for channel habitat unit classification are depth and velocity, for which we had field measurements in Reach 1 and Reach 2 (Table 2). To extend the classification to the upstream segment where we had no field measurements, we could use a hydraulic model to estimate depth and velocity, but such models require input data on the bed topography. We had field-measured bed elevations for Reach 1, but not for Reach 2 or the upstream segment. We could, however, use remote sensing to estimate the water depth from the aerial image for the entire 4.1-km segment. We could then calculate the bed elevation by subtracting the estimated depth from the water-surface elevation. We did not have direct measurements of water-surface elevation, but it was straightforward to calculate for Reach 1, where we did have measurements of bed elevation and water depth, and the water-surface elevation is simply the sum of the two. We assumed that the bed slope and water-surface slope were the same (a common assumption in many hydraulic models) and that the slope was constant throughout the 4.1-km segment (a reasonable assumption given the lack of any discontinuities or structural controls that would result in changing slopes). We used this constant slope to extend the water-surface elevation to Reach 2 and the upstream segment, and subtracted the remotely-sensed depths from the estimated water-surface elevation to calculate the bed elevation. The resulting topographic surface could then be used as input to a hydraulic model. Details on the field methods, remote-sensing techniques, and hydraulic model are given below.
2.3. Field Methods
Fieldwork was carried out in May 2016, during the dry season when flows are low in the Bladen River. In Reach 1 and Reach 2, we set up transects across the channel every 20 m and made depth and velocity measurements every 2 m across each transect, for a total of 761 measurements. Depth was measured with a stadia rod, and velocity with a Marsh-McBirney current meter (Figure 4a). In Reach 1 (but not Reach 2), we also measured the bed elevation using a construction-grade laser level, for a total of 575 measurements (Figure 4b). Because the laser level measures relative rather than absolute elevation and no surveyed benchmark was available nearby, we measured elevations relative to an arbitrary location near the upstream end of Reach 1 rather than a defined datum. Relative elevations were sufficient for our purpose of initializing a hydraulic model with the bed topography.
2.4. Remote Sensing
Multiple methods exist to estimate water depth from remotely-sensed imagery, including techniques optimized for both coastal and riverine environments [16]. The data source we used to estimate depth for the upstream segment, where field measurements of depth were unavailable, was a high-resolution (10-cm) aerial image of the Bladen Nature Reserve, acquired and made available to us by Jamie Rotenberg of the University of North Carolina-Wilmington. We applied the Hydraulically Assisted Bathymetry 2 (HAB-2) model, an algorithm designed to estimate water depth from remotely-sensed imagery based on the Beer-Lambert law of exponential absorption of light in water columns with minimal scattering [17]. HAB-2 works best in clear water, and the turbidity of the Bladen River during the dry season, when both the field data and the aerial imagery were acquired, is very low. The accuracy of the HAB-2 depth estimates are also affected by other factors such as shading by trees. However, because the Bladen River has a relatively wide channel, such impacts affected only a small fraction of the total channel area. The high resolution of the BFREE imagery ensures that cross-sectional variations in depth can be adequately captured using the HAB-2 model, which is important for our purpose of using depth to estimate bed topography.
The HAB-2 model is based on the equation:
where D is the depth of a given pixel, DN is the digital number for that pixel, DN0 is the digital number for the dry riverbed, and β is a diffuse attenuation coefficient. First, we performed an unsupervised classification of the entire image to mask out non-water pixels. We used unsupervised classification because it was the simplest approach to automatically extracting the water pixels, which were clearly identifiable visually in the image. We defined the DN0 value as the digital number of the river pixel with the highest digital number, which translates to a very shallow depth and a pixel covered entirely by sand (when sand is the brightest substrate on the riverbed, as it is in this case). β is a function of the light-extinction depth commonly measured with a Secchi disk in deep water, but in a shallow, clear river such as the Bladen, the light-extinction depth cannot be measured directly. Instead, we used an iterative process to determine β in which we calculated water depth using HAB-2 with different integer values of β, until the calculated water depths best matched the field-measured water depths from Reach 1 and Reach 2 (Figure 5a and Table 3). Note that the date of acquisition for the field data and aerial imagery was not the same, so it is not expected that the observed and modeled depth data would match exactly even if the model’s performance were perfect. Nevertheless, both the field data and the aerial imagery were acquired during the dry season during low flow. Based on comparison of field-measured wetted width with the wetted width measured from the aerial imagery, the discharge when the aerial imagery was acquired and when we did fieldwork was similar, which provides support for using the HAB-2 depths and field-measured water-surface elevations to estimate the bed topography in Reach 2 and the upstream segment.
D = ln(DN/DN0) − β
2.5. Hydraulic Modeling
In order to estimate flow velocity, one of the ecohydraulic variables needed to delineate channel habitat units, for the upstream segment, we used the International Rivers Interface Cooperative (iRIC) hydraulic modeling software suite [18]. iRIC takes input elevation data in several formats, including point data from field surveys. We used the field-measured bed elevation points as the input elevation data for Reach 1. For Reach 2 and the upstream segment, we used the water-surface elevation and constant water-surface slope calculated from the bed elevation of Reach 1 to extend the water-surface elevation upstream and subtracted the HAB-2 depth values to calculate the bed elevation, then converted the gridded elevation to points for use in iRIC. We used iRIC’s curvilinear orthogonal interpolation function to create a model grid, which preferentially interpolates in the longitudinal rather than cross-sectional direction. Our input elevation data were mapped to this grid.
iRIC contains a number of solvers. The solver we used was Flow and Sediment Transport with Morphological Evolution of Channels (FaSTMECH), a two-dimensional quasi-steady hydraulic model. In addition to the topographic input, FaSTMECH requires upstream and downstream boundary conditions. For the upstream boundary condition, we used steady flow input of the discharge measured in the field. Although this discharge was measured in Reach 1, near the downstream end of the modeled river segment, there are no major tributaries entering the Bladen River throughout the entire segment, so it is assumed that the discharge at the upstream end is similar. For the downstream boundary condition, we used the stage at the downstream end, calculated as the measured bed elevation plus measured depth, relative to the arbitrarily defined reference datum used in the topographic surveying. We ran FaSTMECH using these boundary conditions and produced outputs of water depth and flow velocity for the entire segment. Because we used the low-flow boundary conditions at the time of our field data collection and aerial image acquisition, the hydraulic results should only be considered valid during the dry season. We used the hydraulic modeling results to delineate channel habitat units based on the ratio between velocity and depth. We defined the units using thresholds developed by Jowett [13], where a ratio of greater than 3.20 indicates a riffle, a ratio between 1.24 and 3.20 indicates a run, and a ratio of less than 1.24 indicates a pool. Because this definition of channel habitat units is based on ratios between velocity and depth, the classification should be robust across ranges of different discharge. We performed some simple spatial statistics—including calculation of the total length, percent of segment length, and average spacing of each unit—to analyze the spatial distribution of channel habitat units across the modeled segment of the Bladen River.
3. Results
Figure 2b shows the HAB-2 water depths for the Bladen River, which reach a maximum depth of 4 m and are significantly correlated with observed depths in Reach 1 and Reach 2 (Figure 5 and Table 3). Figure 2c shows the water-surface elevation, which was estimated by extending a constant water-surface slope upstream from the measured water-surface elevation of Reach 1. The HAB-2 depths (Figure 2b) were subtracted from the estimated water-surface elevation (Figure 2c) to create the bed elevation used as input to the FaSTMECH model (Figure 2d).
Figure 6 shows the FaSTMECH modeling results. The modeled water depths (Figure 6a) are substantially higher than the observed values in Reach 1 and Reach 2 (Figure 5b), and the correlation between the two is lower than between the HAB-2 estimated depths and the measured values, although the two are significantly correlated (Table 3). The modeled flow velocities (Figure 6b) are also substantially higher than the observed values. The distribution of velocities is clustered around relatively small values (<2 m s−1), but there are several instances of high velocity (>3 m s−1) that show up in both the observations and the model output (Figure 5c). These high-velocity points are associated with rapids exiting a deep pool near the upstream end of Reach 1. Overall, the correlation between observed and modeled flow velocity is significant, and the R2 is higher for FaSTMECH-simulated velocity than for FaSTMECH-simulated depth, but is not as high as for HAB-2 depths. This finding indicates that the hydraulic modeling component introduces more uncertainty into the overall channel habitat unit delineation process than does the remote sensing. Both depths estimated from HAB-2 and depths and velocities modeled by FaSTMECH are positively biased, meaning depth and velocity are both overestimated by the modeling procedures (Figure 5). The most likely explanation for this systematic bias is that discharge was higher during the period of aerial imagery acquisition than during the field data collection period, resulting in higher estimates of both depth and velocity.
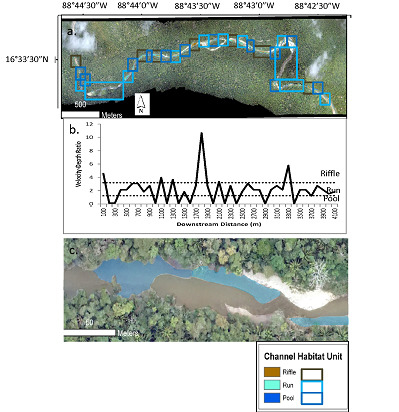
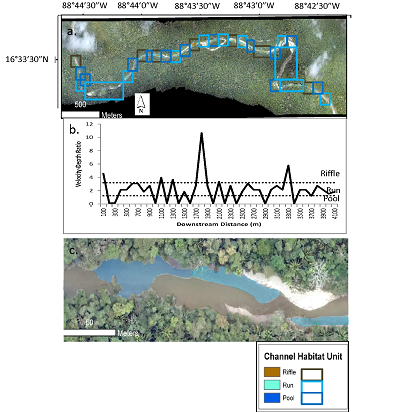
We used the modeled depth and velocity to classify the entire study segment into three types of channel habitat unit: pools, riffles, and runs. Although there are more types of channel habitat unit, we selected these three because (1) they are the most commonly identified channel habitat units that are known to provide a range of different habitat conditions of importance to aquatic species [1,2,3,4]; and (2) these channel habitat units can be classified on the basis of depth and velocity alone, without the need for additional data, such as substrate conditions, that are not available for the entire study segment. Figure 7a shows the delineated channel habitat units, based on applying on the Jowett [13] velocity-depth thresholds to the FaSTMECH modeling results, for the entire Bladen River study segment, while Figure 7c shows a detailed close-up of a particular reach. Figure 7b shows average velocity-depth ratios for 100-m bins of the channel from upstream to downstream, along with the thresholds for the channel habitat units. In general, the most frequent shifts between channel habitat units are concentrated in the middle segment, which is where the transition from the confined montane to alluvial floodplain channel occurs.
To validate the channel habitat unit classification, we also applied the Jowett [13] velocity-depth ratios to 186 points in Reach 1 and Reach 2 for which we had field measurements of velocity and depth, and compared these points to the channel habitat units delineated for those locations using the HAB-2/FaSTMECH procedure. Overall, 83% of points were classified accurately, with accuracies ranging from 81% for runs to 88% for riffles (Table 4). This finding indicates that, despite the previously noted systematic biases in the HAB-2 depths and FaSTMECH-simulated depth and velocities, the classification procedure results in reasonably accurate channel habitat units.
According to the channel habitat unit classification, the unit that covered the greatest length in the study segment was runs, which made up nearly half (47%) of the total segment length (Table 5). The dominance of runs may be explained by the intermediate values of velocity-depth ratio used to define a run, which makes it essentially a catchall channel habitat unit for any portion of the channel that is not a pool or riffle. The second most prevalent habitat unit was pools, which made up 32% of the total segment length. As expected, pools were most commonly found on the outside of bends, and also in some areas along one side of the channel with a riffle on the other side (Figure 7c. The least prevalent channel habitat unit was riffles, which made up just 21% of the total segment length. The low prevalence of riffles may be explained in part by the igneous lithology of the study segment, in which quartz-rich parent material weathers predominantly to sand. The sand is deposited relatively evenly in the channel to create runs, rather than the concentrated deposition needed to create riffles, which would be expected for coarser bed material that requires a higher competence, such as gravels [19].
Table 4 also shows the average spacing between each delineated channel habitat unit. The spacing ranges from 284 m for pools to 550 m for runs. This indicates that, although runs covered more of the segment length than any other channel habitat unit, they were predominantly found in several very long expanses that were far apart from one another (Figure 7a). Pools and riffles, in contrast, while covering less of the total segment length, tended to alternate more frequently with other channel habitat units than did runs. The commonly reported value for pool and riffle spacing is five to seven bankfull channel widths [20]. Based on our field-measured average bankfull width value for Reach 1 of ~35 m, the expected pool and riffle spacing for the Bladen River study segment would be only 175 to 245 m. Our delineated channel habitat units all had greater average spacing than this expected value, which suggests the potential for greater overall amplitude of channel habitat units in the study segment.
4. Discussion
This study involved the development of an innovative approach to delineating channel habitat units using a combination of fieldwork, remote sensing, and hydraulic modeling. Channel habitat units have often been delineated from aerial imagery, but usually this is done using supervised classification. For example, Marcus et al. [21] classified in-stream habitat using supervised classification of high-resolution hyperspectral imagery. An advantage of the approach presented here is that it is based on hydraulic variables (depth and velocity), rather than purely on the spectral signatures generated by different channel habitat units. This incorporation of hydraulic variables can only be accomplished if hydraulic modeling is combined with remote sensing. For example, Legleiter and Goodchild [22] incorporated hydraulic modeling into the remote sensing process with a combination of supervised classification and fuzzy clustering to delineate channel habitat units. Such combined approaches, however, have rarely been applied, especially on tropical rivers.
A limitation of optical remote sensing to delineate channel habitat units is that the algorithms do not work in turbid water, or where canopy cover obstructs the channel. The Bladen River presented an ideal case study for optical remote sensing of channel habitat units, because its water is very clear during the dry season and it is wide enough that the channel is never completely obstructed by vegetation. Such ideal conditions, however, are not common in tropical rivers, which often experience high turbidity and have dense riparian vegetation. In such cases, alternative sensors such as blue-green LiDAR may be preferable for mapping channel habitat units. For example, McKean et al. [23] used a narrow-beam aquatic-terrestrial LiDAR to map and calculate the volume of pools. Because of the great expense of LiDAR sensors and the need to have the imagery flown, however, approaches based on optical imagery still fulfill a necessary function.
In this study, both the HAB-2 estimates of depth and the FaSTMECH-simulated depths and velocities exhibited a positive bias. The likely cause is the fact that the dates of aerial image acquisition and field data collection were not the same. If the discharge was higher when the aerial imagery was flown compared to when we collected the field data, this would result in overestimates of depth in the HAB-2 model. This positive bias in depth would then propagate to the depths and velocities simulated by FaSTMECH. Future research on initializing hydraulic models with remotely-sensed depth estimates should endeavor to collect field data concurrently with image acquisition, to avoid such cases of systematic bias. The bias problem could also be alleviated with observed discharge data from gaging stations, which would allow for more precise calibration of the β diffuse attenuation coefficient in the HAB-2 model. These corrective steps were not possible on the Bladen River because it lacks gaging stations, so the systematic bias in the depth and velocity measurements remains a fundamental limitation of the study. It should be noted, however, that the ratio between velocity and depth is more important than their absolute values in delineating channel habitat units, and the comparison of the observed and modeled ratios indicates that the classification procedure was largely successful in accurately distinguishing channel habitat units in Reach 1 and Reach 2.
The results of this study indicate that the greatest source of uncertainty in the approach taken here is the hydraulic modeling. The depth estimates from the HAB-2 model match reasonably well to observations, but the FaSTMECH-modeled depth and velocity did not match observations as well. A number of limitations of this study contribute to FaSTMECH’s modeling performance. In particular, the estimation of water-surface elevation in the upstream segment and the estimation of the upstream discharge boundary condition using downstream discharge are likely to be the main sources of error in the hydraulic modeling. These issues could be remedied by a minimal amount of additional field data collection at the upstream end of the study segment, including measurement of water-surface elevation and discharge to serve as upstream boundary conditions. In particular, both issues would be addressed if a stream gage were available to record stage and discharge at the upstream end of the study segment. Availability of gage data would also allow the low-flow conditions during the dry season simulated here to be extended to high flows during the rainy season, to allow for analysis of change over time in channel habitat units. The Bladen River is currently ungaged, which is common in remote tropical rivers. Future research should consider using aerial photography specifically acquired for processing through Structure-from-Motion, so that water level could be mapped directly from the imagery [24].
This study found some evidence that the spacing of channel habitat units on this segment of the Bladen River is larger than the typical values reported for midlatitude rivers. This difference could potentially suggest an overall amplified wavelength of channel habitat units, which would be consistent with the high erosion rates resulting from high discharge per unit area and high weathering rates in tropical rivers. Many other empirical relationships developed to predict characteristics of rivers, primarily based on midlatitude rivers, have been found inadequate when applied to tropical rivers. For example, Baker [25] found that the sediment-size sinuosity relationships developed for temperate rivers do not apply to tropical rivers. As noted by Latrubesse et al. [26], the empirical equations used to relate channel morphology and sediment load to predict channel patterns often fail to predict tropical river planforms. Because this study examined only a short segment of a single river system, there is not enough information to conclude that tropical rivers generally have larger spacing of channel habitat units than is typical for midlatitude rivers, but future research should investigate this possibility by examining longer river segments in a diversity of tropical environments.
Moreover, many tropical rivers experience significant flow variability between the dry and wet seasons, which could result in channel habitat units being more dynamic than in many midlatitude rivers. For example, Rayner et al. [27] found that small tropical rivers that lack extensive floodplain storage experience a reduction in habitat heterogeneity during the wet season, when bed sediments are mobilized and aquatic vegetation is scoured. To address the question of whether channel habitat units in tropical rivers are more dynamic seasonally or change over time more rapidly than those in midlatitude rivers, additional research is needed to delineate changes in channel habitat units over time.
In addition to what it reveals about fundamental fluvial processes, the delineation of channel habitat units has important implications for river management. Physical habitat assessments are widely used for targeting priority areas for conservation, designing stream restoration projects, and setting environmental instream flow standards [28]. Managing rivers for conservation objectives is especially critical in tropical rivers, given their high levels of aquatic biodiversity relative to higher-latitude rivers [29]. Native aquatic species in the Bladen River, including several cichlid species, have been found to be sensitive to physical habitat variables [30,31]. Given the high aquatic biodiversity of tropical rivers—and threats to those rivers from climate change, land-use change, dam construction, and invasive species—it is imperative to build more comprehensive understanding of the controls on and spatial distribution of channel habitat units in tropical rivers. In particular, species richness is expected to be higher in heterogeneous habitat types, such as sequences of pools and riffles, rather than in relatively uniform stretches of runs.
5. Conclusions
This study used a novel approach combining field measurements, remote sensing, and hydraulic modeling to delineate channel habitat units for a 4.1-km segment of the Bladen River in southern Belize. The approach taken here generally performed well, although a significant amount of uncertainty was contributed by the lack of field-measured upstream boundary conditions for the hydraulic model. Results indicate that the most common channel habitat unit on this segment of the Bladen River was runs, followed by pools and riffles. The results also indicate that the spacing of channel habitat units is possibly larger than the typical expected values for midlatitude rivers, although additional research is needed to verify if this is true of tropical rivers more generally. Given the exceptional aquatic biodiversity of tropical rivers and the threats posed to that diversity, gaining a better understanding of the controls and spatial patterns of channel habitat units on these rivers is critical. Because of the difficulty of extensive field surveys in remote tropical regions, approaches that incorporate remote sensing and hydraulic modeling offer a promising alternative that can contribute to the understanding of channel habitat units on tropical rivers.
Acknowledgments
Funding was provided by the University of Alabama Research Grants Committee (to S.P.), University of Alabama graduate student funding sources (to E.B.), American Association of Geographers Water Resources Specialty Group (to E.B.), and the Edward H. Moseley Endowed Scholarship (to E.B.). The Belize Foundation for Research and Environmental Education (BFREE) provided field access and logistical support. Jamie Rotenberg of the University of North Carolina—Wilmington provided the aerial imagery. The Belize Fisheries Department and Ya’axche Conservation Trust provided access to the Bladen Nature Reserve. Melito Bustamente and Melvin Choc assisted in the field.
Author Contributions
S.P. and E.B. developed the research design; E.B. and S.P. collected field data; S.P. did the remote sensing analysis and hydraulic modeling; S.P. and E.B. analyzed the data; S.P. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Frissell, C.A.; Liss, W.J.; Warren, C.E.; Hurley, M.D. A hierarchical framework for stream habitat classification: Viewing streams in a watershed context. Environ. Manag. 1986, 10, 199–214. [Google Scholar] [CrossRef]
- Hawkins, C.P.; Kershner, J.L.; Bisson, P.A.; Bryant, M.D.; Decker, L.M.; Gregory, S.V.; McCullough, D.A.; Overton, C.K.; Reeves, G.H.; Steedman, R.J.; et al. A hierarchical approach to classifying stream habitat features. Fisheries 1993, 18, 3–12. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Buffington, J.M. Channel-reach morphology in mountain drainage basins. Geol. Soc. Am. Bull. 1997, 109, 596–611. [Google Scholar] [CrossRef]
- Thomson, J.R.; Taylor, M.P.; Fryirs, K.A.; Brierley, G.J. A geomorphological framework for river characterization and habitat assessment. Aquat. Conserv. 2001, 11, 373–389. [Google Scholar] [CrossRef]
- Buffagni, A.; Crosa, G.A.; Harper, D.M.; Kemp, J.L. Using macroinvertebrate species assemblages to identify river channel habitat units: An application of the functional habitats concept to a large, unpolluted Italian river (River Ticino, northern Italy). Hydrobiologia 2000, 435, 213–225. [Google Scholar] [CrossRef]
- Newson, M.D.; Newson, C.L. Geomorphology, ecology, and river channel habitat: Mesoscale approaches to basin-scale challenges. Prog. Phys. Geog. 2000, 24, 195–217. [Google Scholar] [CrossRef]
- Crispin, V.; House, R.; Roberts, D. Changes in instream habitat, large woody debris, and salmon habitat after the restructuring of a coastal Oregon stream. N. Am. J. Fish Manag. 1993, 13, 96–102. [Google Scholar] [CrossRef]
- Quinn, T.P.; Peterson, N.P. The influence of habitat complexity and fish size on over-winter survival and growth of individually marked juvenile coho salmon (Oncorhynchus kisutch) in Big Beef Creek, Washington. Can. J. Fish. Aquat. Sci. 1996, 53, 1555–1564. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Karr, J.R. Fish communities along environmental gradients in a system of tropical streams. Environ. Biol. Fish. 1983, 9, 117–135. [Google Scholar] [CrossRef]
- Willis, S.C.; Winemiller, K.O.; Lopez-Fernandez, H. Habitat structural complexity and morphological diversity of fish assemblages in a Neotropical floodplain river. Oecologia 2005, 142, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Montaña, C.G.; Winemiller, K.O. Local-scale habitat influences morphological diversity of species assemblages of cichlid fishes in a tropical floodplain river. Ecol. Freshw. Fish 2010, 19, 216–227. [Google Scholar] [CrossRef]
- Roper, B.B.; Scarnecchia, D.L. Observer variability in classifying habitat types in stream surveys. N. Am. J. Fish. Manag. 1995, 15, 49–53. [Google Scholar] [CrossRef]
- Jowett, I.G. A method for objectively identifying pool, run, and riffle habitats from physical measurements. N. Z. J. Mar. Fresh. 1993, 27, 241–248. [Google Scholar] [CrossRef]
- Wohl, E. Downstream hydraulic geometry along a tropical mountain river. In The Río Chagres, Panama; Harmon, R.S., Ed.; Springer: Dordrecht, The Netherlands, 2005; pp. 169–188. [Google Scholar]
- Iremonger, S.; Sayre, R. A Rapid Ecological Assessment of the Bladen Nature Reserve, Belize; The Nature Conservancy: Arlington, VA, USA, 1994. [Google Scholar]
- Stumpf, R.P.; Holderied, K.; Sinclair, M. Determination of water depth with high-resolution satellite imagery over variable bottom types. Limnol. Oceanogr. 2003, 48, 547–556. [Google Scholar] [CrossRef]
- Fonstad, M.A.; Marcus, W.A. Remote sensing of stream depths with hydraulically assisted bathymetry (HAB) models. Geomorphology 2005, 72, 320–399. [Google Scholar] [CrossRef]
- Nelson, J.M.; Shimizu, Y.; Abe, T.; Asahi, K.; Gamou, M.; Inoue, T.; Iwasaki, T.; Kakinuma, T.; Kawamura, S.; Kimura, I.; et al. The international river interface cooperative: Public domain flow and morphodynamics software for education and applications. Adv. Water Resour. 2016, 93, 62–74. [Google Scholar] [CrossRef]
- Gibbins, C.N.; Vericat, D.; Batalla, R.J. Relations between invertebrate drift and flow velocity in sand-bed and riffle habitats and the limits imposed by substrate stability and benthic density. J. N. Am. Benthol. Soc. 2010, 29, 945–958. [Google Scholar] [CrossRef]
- Leopold, L.B.; Wolman, M.G.; Miller, J.P. Fluvial Processes in Geomorphology; Dover Publications: New York, NY, USA, 1964. [Google Scholar]
- Marcus, W.A.; Legleiter, C.J.; Aspinall, R.J.; Boardman, J.W.; Crabtree, R.L. High spatial resolution hyperspectral mapping of in-stream habitats, depths, and woody debris in mountain streams. Geomorphology 2003, 55, 363–380. [Google Scholar] [CrossRef]
- Legleiter, C.J.; Goodchild, M.F. Alternative representations of in-stream habitat: Classification using remote sensing, hydraulic modeling, and fuzzy logic. Int. J. Geogr. Inf. Sci. 2005, 19, 29–50. [Google Scholar] [CrossRef]
- McKean, J.; Nagel, D.; Tonina, D.; Bailey, P.; Wright, C.W.; Bohn, C.; Nayegandhi, A. Remote sensing of channels and riparian zones with a narrow-beam aquatic-terrestrial LIDAR. Remote Sens. 2009, 1, 1065–1093. [Google Scholar] [CrossRef]
- Fonstad, M.A.; Dietrich, J.T.; Courville, B.C.; Jensen, J.L.; Carbonneau, P.E. Topographic structure from motion: A new development in photogrammetric measurement. Earth Surf. Process. Landf. 2013, 38, 421–430. [Google Scholar] [CrossRef]
- Baker, V.R. Adjustment of fluvial systems to climate and source terrain in tropical and subtropical environments. Fluv. Sedimentol. 1977, 5, 211–230. [Google Scholar]
- Latrubesse, E.M.; Stevaux, J.C.; Sinha, R. Tropical Rivers. Geomorphology 2005, 70, 187–206. [Google Scholar] [CrossRef]
- Rayner, T.S.; Pusey, B.J.; Pearson, R.G. Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, North-East Queensland: Towards a new conceptual framework for understanding fish-habitat dynamics in small tropical rivers. Mar. Freshw. Res. 2008, 59, 97–116. [Google Scholar] [CrossRef]
- Maddock, I. The importance of physical habitat assessment for evaluating river health. Freshw. Biol. 1999, 41, 373–391. [Google Scholar] [CrossRef]
- Allan, J.D.; Flecker, A.S. Biodiversity conservation in running waters. BioScience 1993, 43, 32–43. [Google Scholar] [CrossRef]
- Esselman, P.C.; Freeman, M.C.; Pringle, C.M. Fish-assemblage variation between geologically defined regions and across a longitudinal gradient in the Monkey River Basin, Belize. J. N. Am. Benthol. Soc. 2006, 25, 142–156. [Google Scholar] [CrossRef]
- Cochran-Biederman, J.L.; Winemiller, K.O. Relationships among habitat, ecomorphology and diets of cichlids in the Bladen River, Belize. Environ. Biol. Fish. 2010, 88, 143–452. [Google Scholar] [CrossRef]
Figure 1.
Location of (a) the Bladen River in Belize; (b) the study reach on the Bladen River.

Figure 2.
(a) Locations of Reach 1 and Reach 2 with field survey points, and the upstream segment, on high-resolution aerial image; (b) water depth, estimated from HAB-2; (c) water-surface elevation, estimated by extending a constant water-surface slope upstream from Reach 1; (d) bed elevation, calculated by subtracting HAB-2 water depths from estimated water-surface elevation, and interpolated to grid using iRIC’s curvilinear orthogonal function; (e) zoomed-in view of Reach 1 and Reach 2; (f) zoomed-in view of HAB-2 water depth for Reach 1 and Reach 2; (g) zoomed-in view of water-surface elevation for Reach 1 and Reach 2; (h) zoomed-in view of bed elevation for Reach 1 and Reach 2.
Figure 2.
(a) Locations of Reach 1 and Reach 2 with field survey points, and the upstream segment, on high-resolution aerial image; (b) water depth, estimated from HAB-2; (c) water-surface elevation, estimated by extending a constant water-surface slope upstream from Reach 1; (d) bed elevation, calculated by subtracting HAB-2 water depths from estimated water-surface elevation, and interpolated to grid using iRIC’s curvilinear orthogonal function; (e) zoomed-in view of Reach 1 and Reach 2; (f) zoomed-in view of HAB-2 water depth for Reach 1 and Reach 2; (g) zoomed-in view of water-surface elevation for Reach 1 and Reach 2; (h) zoomed-in view of bed elevation for Reach 1 and Reach 2.

Figure 3.
Sources of data and the measurement, calculation, or estimation method used.

Figure 4.
Field measurements of (a) water depth using a stadia rod and flow velocity using a Marsh-McBirney current meter; (b) bed elevation using a construction-grade laser level.
Figure 4.
Field measurements of (a) water depth using a stadia rod and flow velocity using a Marsh-McBirney current meter; (b) bed elevation using a construction-grade laser level.

Figure 5.
(a) Measured (stadia rod) and modeled (HAB-2) water depths for Reach 1 and Reach 2; (b) measured (stadia rod) and modeled (FaSTMECH) water depths for Reach 1 and Reach 2; (c) measured (current meter) and modeled (FaSTMECH) flow velocities for Reach 1 and Reach 2 (two outliers excluded); (d) measured (current meter) and modeled (FaSTMECH) flow velocities for Reach 1 and Reach 2 (all points included).
Figure 5.
(a) Measured (stadia rod) and modeled (HAB-2) water depths for Reach 1 and Reach 2; (b) measured (stadia rod) and modeled (FaSTMECH) water depths for Reach 1 and Reach 2; (c) measured (current meter) and modeled (FaSTMECH) flow velocities for Reach 1 and Reach 2 (two outliers excluded); (d) measured (current meter) and modeled (FaSTMECH) flow velocities for Reach 1 and Reach 2 (all points included).

Figure 6.
FaSTMECH-simulated (a) water depth; and (b) flow velocity.

Figure 7.
(a) Delineated channel habitat units for entire Bladen River study segment, based on simulated velocity-depth ratios for 100-m bins of the channel; (b) average velocity-depth ratios and channel habitat units from upstream to downstream; (c) detail for a particular reach. The black box in (a) delineates the reach shown in (c).
Figure 7.
(a) Delineated channel habitat units for entire Bladen River study segment, based on simulated velocity-depth ratios for 100-m bins of the channel; (b) average velocity-depth ratios and channel habitat units from upstream to downstream; (c) detail for a particular reach. The black box in (a) delineates the reach shown in (c).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the Monkey River Basin, Belize.
| Characteristic | Value |
|---|---|
| Drainage area | 1291 km2 |
| Basin relief | 1011 m |
| Lithology | 49% Late Carboniferous-Permian volcanic material, 33% Quaternary alluvial material, 17% Triassic limestone |
| Physiography | 37% Maya Mountains igneous/metamorphic complex, 31% Monkey River floodplain, 14% moderate-gradient volcanic slopes |
| Soils | 37% Leptosol-Cambisol, 31% Fluvisol-Cambisol-Vertisol, 14% Litosol-Cambisol |
| Land cover | 89% evergreen broadleaf forest, 7% cropland, 2% woody savannah, 1% mixed forest, 1% savannah |
| Mean annual temperature | 25 °C |
| Mean annual precipitation | 406 cm |
Table 2.
Data sources used for different reaches and variables.
| Reach | Variable | Source |
|---|---|---|
| Reach 1 | Depth | Measured (stadia rod) |
| Velocity | Measured (current meter) | |
| Bed elevation | Measured (laser level) | |
| Water-surface elevation | Calculated (bed elevation plus depth) | |
| Water-surface slope | Calculated (from measured bed elevation) | |
| Reach 2 | Depth | Measured (stadia rod) |
| Velocity | Measured (current meter) | |
| Bed elevation | Calculated (water-surface elevation minus depth) | |
| Water-surface elevation | Estimated (extended upstream from Reach 1) | |
| Water-surface slope | Assumed constant | |
| Upstream | Depth | Estimated (HAB-2) |
| Velocity | Modeled (FaSTMECH) | |
| Bed elevation | Calculated (water-surface elevation minus depth) | |
| Water-surface elevation | Estimated (extended upstream from Reach 1) | |
| Water-surface slope | Assumed constant |
Table 3.
Validation statistics for HAB-2 depth estimates and FaSTMECH depth and velocity estimates.
| Model | Variable | R2 | p | Intercept | Coefficient |
|---|---|---|---|---|---|
| HAB-2 | Depth | 0.17 | <0.01 | 1.57 | 0.43 |
| FaSTMECH | Depth | 0.04 | <0.01 | 1.49 | 0.29 |
| Velocity | 0.10 | <0.01 | 1.10 | 0.40 |
Table 4.
Accuracy of HAB-2 channel habitat unit classification compared to observed channel habitat units, determined by applying Jowett [13] velocity-depth thresholds for 186 points in Reach 1 and Reach 2 for which field measurements of velocity and depth were available.
Table 4.
Accuracy of HAB-2 channel habitat unit classification compared to observed channel habitat units, determined by applying Jowett [13] velocity-depth thresholds for 186 points in Reach 1 and Reach 2 for which field measurements of velocity and depth were available.
| Predicted Riffle | Predicted Run | Predicted Pool | Overall Prediction Accuracy | |
|---|---|---|---|---|
| Observed riffle | 42 | 5 | 1 | 0.88 |
| Observed run | 11 | 82 | 8 | 0.81 |
| Observed pool | 2 | 4 | 30 | 0.83 |
| Total | 56 | 91 | 39 | 0.83 |
Table 5.
Total length, percentage of segment length, and average spacing for delineated channel habitat units.
Table 5.
Total length, percentage of segment length, and average spacing for delineated channel habitat units.
| Unit | Total Length (m) | Percentage of Segment Length (%) | Average Spacing (m) |
|---|---|---|---|
| Riffle | 863 | 21 | 518 |
| Run | 1942 | 47 | 550 |
| Pool | 1295 | 32 | 284 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Praskievicz, S.; Buege, E. Remote-Sensing Hydraulic Characterization of Channel Habitat Units in a Tropical Montane River: Bladen River, Belize. Remote Sens. 2017, 9, 1295. https://doi.org/10.3390/rs9121295
AMA Style
Praskievicz S, Buege E. Remote-Sensing Hydraulic Characterization of Channel Habitat Units in a Tropical Montane River: Bladen River, Belize. Remote Sensing. 2017; 9(12):1295. https://doi.org/10.3390/rs9121295
Chicago/Turabian StylePraskievicz, Sarah, and Emily Buege. 2017. "Remote-Sensing Hydraulic Characterization of Channel Habitat Units in a Tropical Montane River: Bladen River, Belize" Remote Sensing 9, no. 12: 1295. https://doi.org/10.3390/rs9121295
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.