
Response of Land Surface Phenology to Variation in Tree Cover during Green-Up and Senescence Periods in the Semi-Arid Savanna of Southern Africa


Abstract
:
1. Introduction
2. Materials and Methods
- -
- determining land surface phenological metric from leaf area index (LAI) time series data via a Gaussian model approach;
- -
- determining the influence of percentage tree cover on various land surface phenological metrics in two ways: (i) assessing whether significant differences exist between three tree cover classes for each land surface phenological metric; and (ii) assessing the strength of the relationship between each land surface phenological metric and percentage tree cover; and
- -
- assessing the influence of rainfall on land surface phenology by Pearson’s correlation between mean rainfall data with the land surface phenological metric.
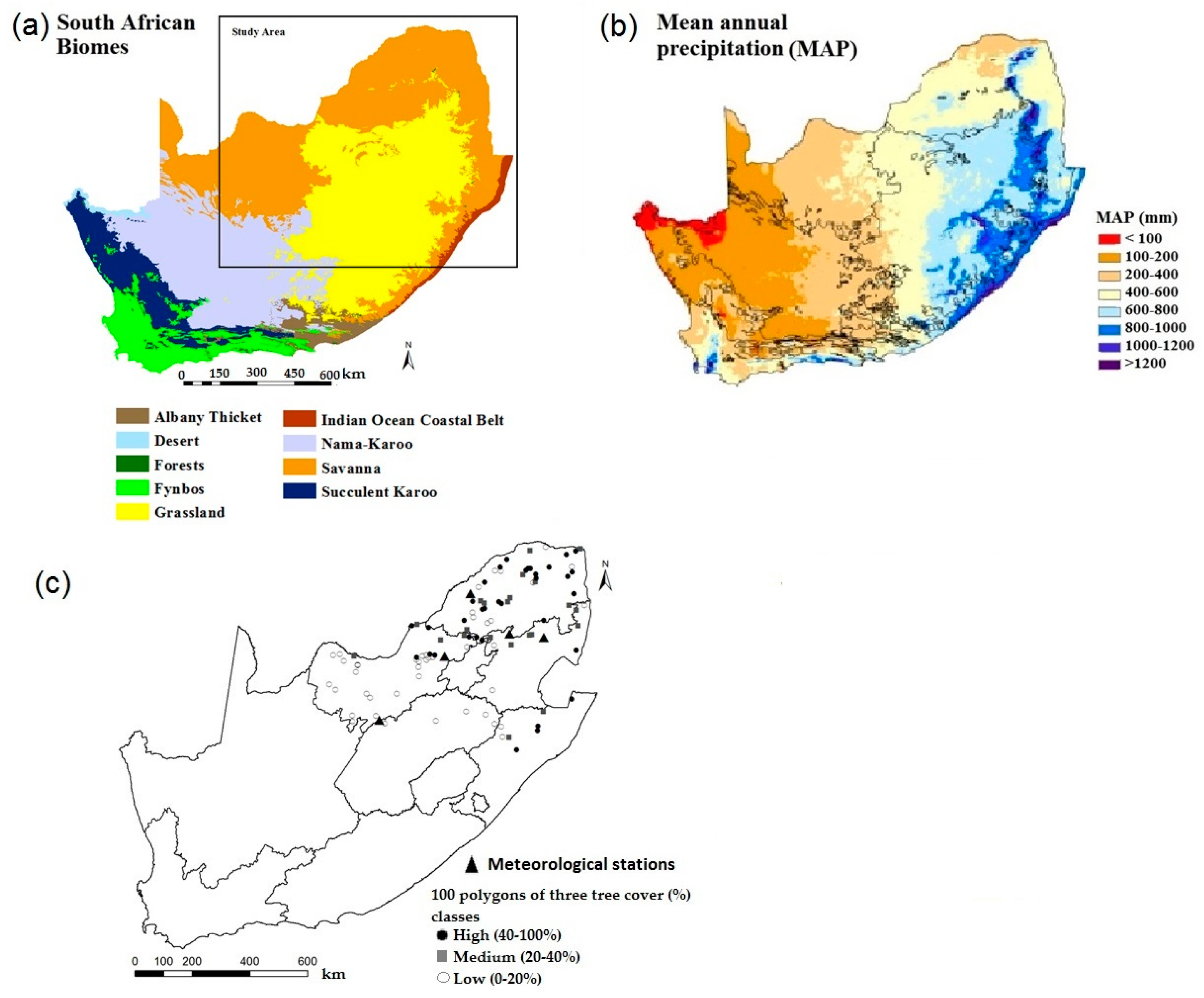
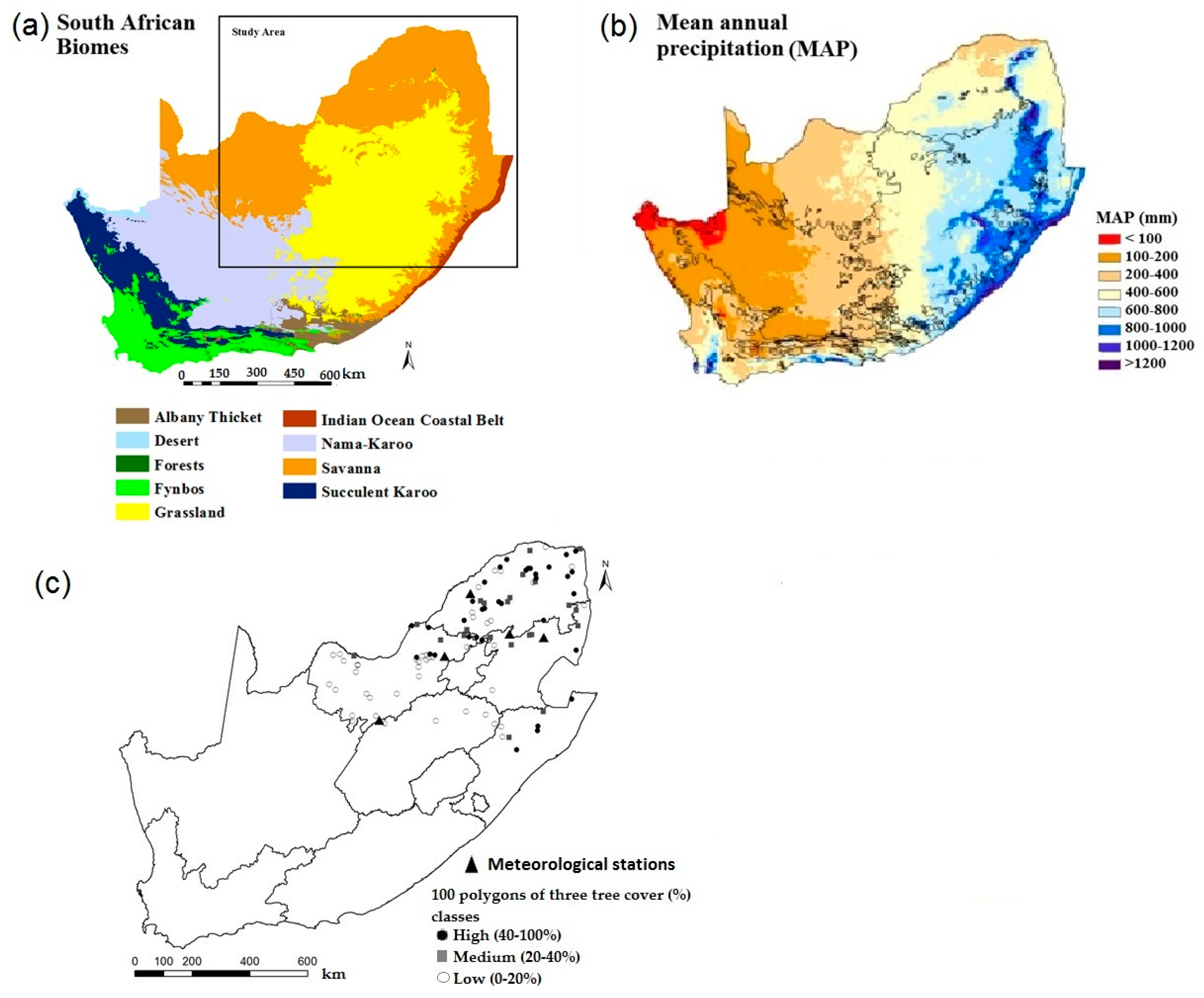
2.1. Study Area
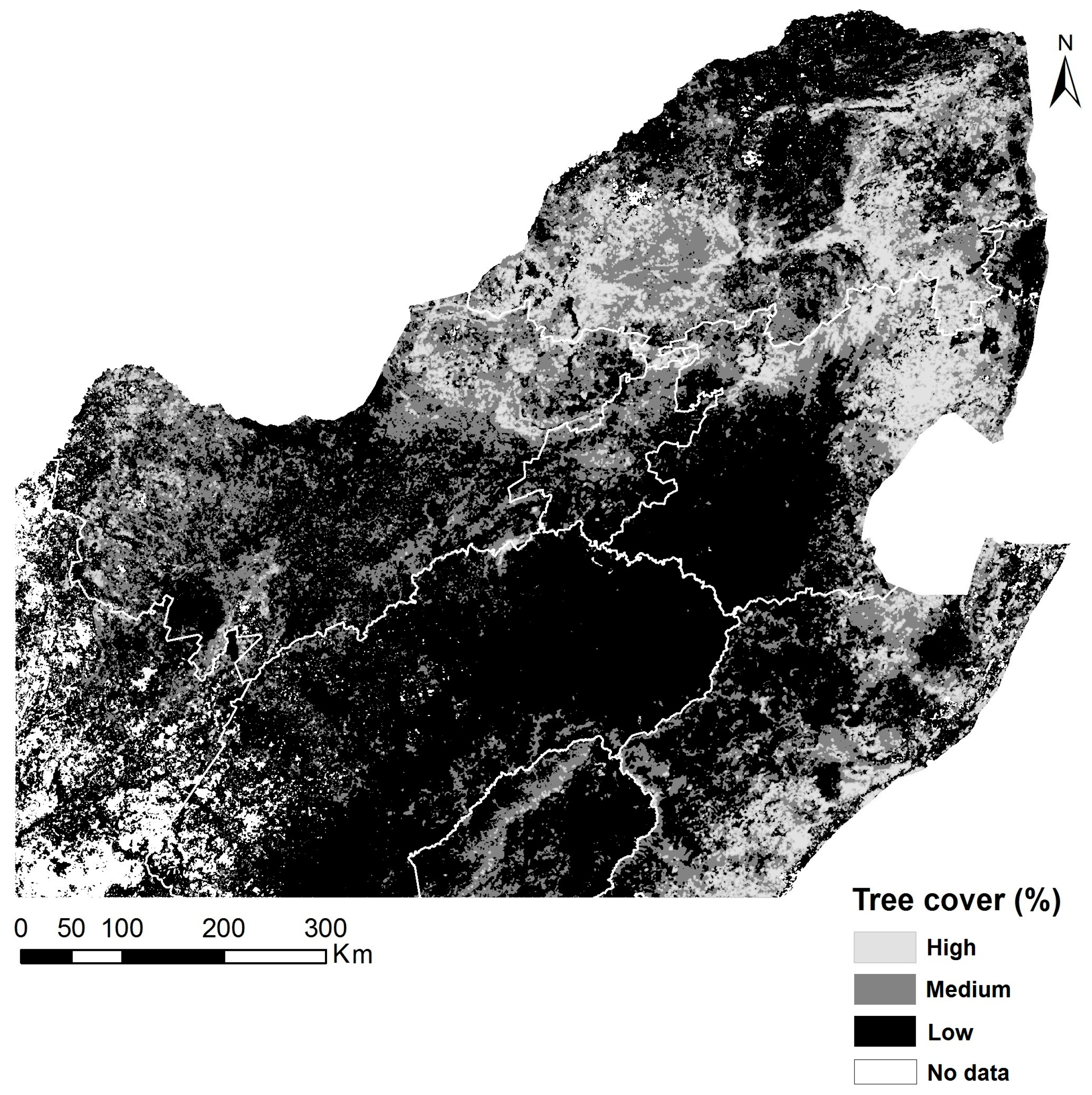
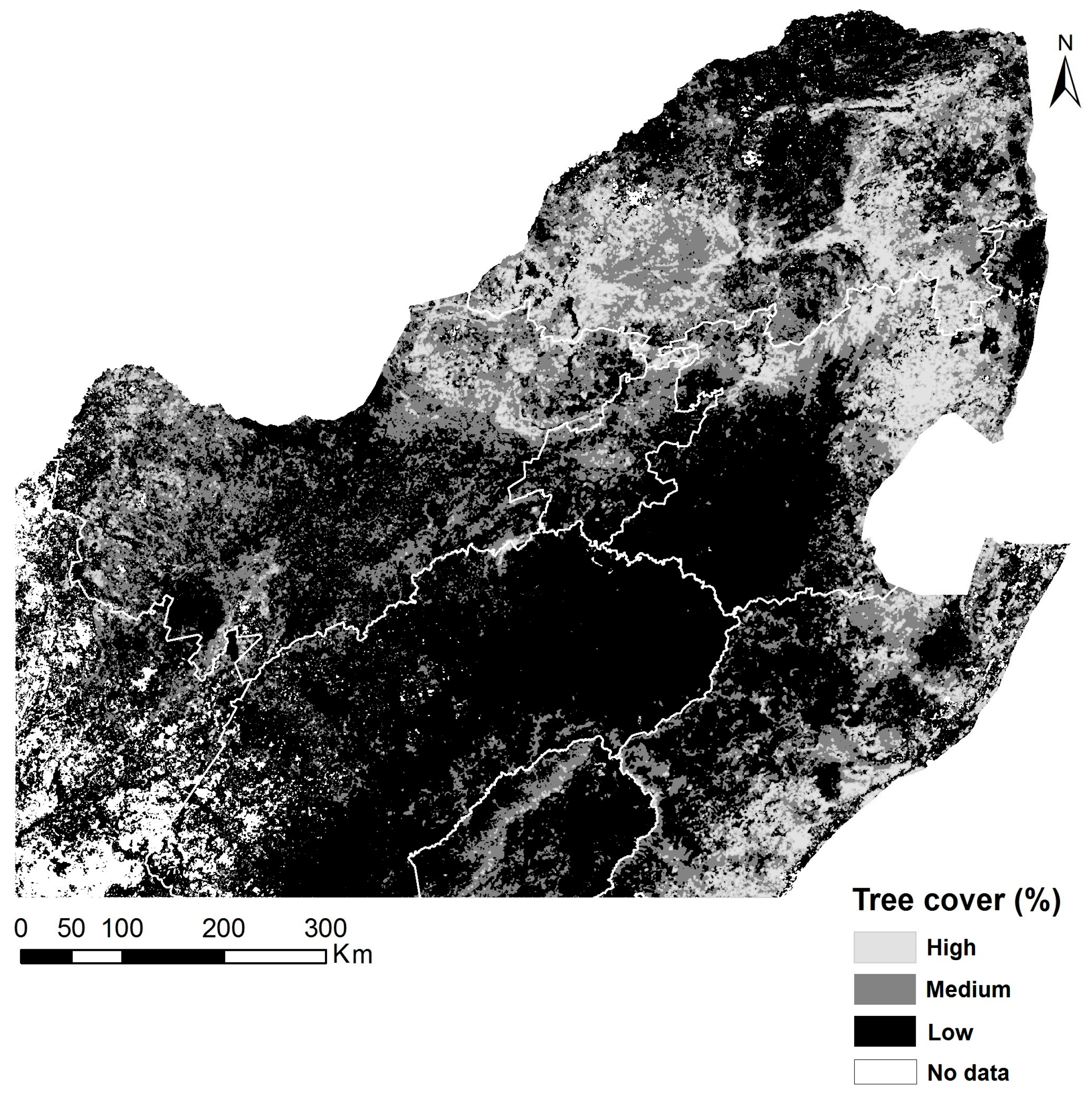
2.2. Tree Cover Percentage Estimation
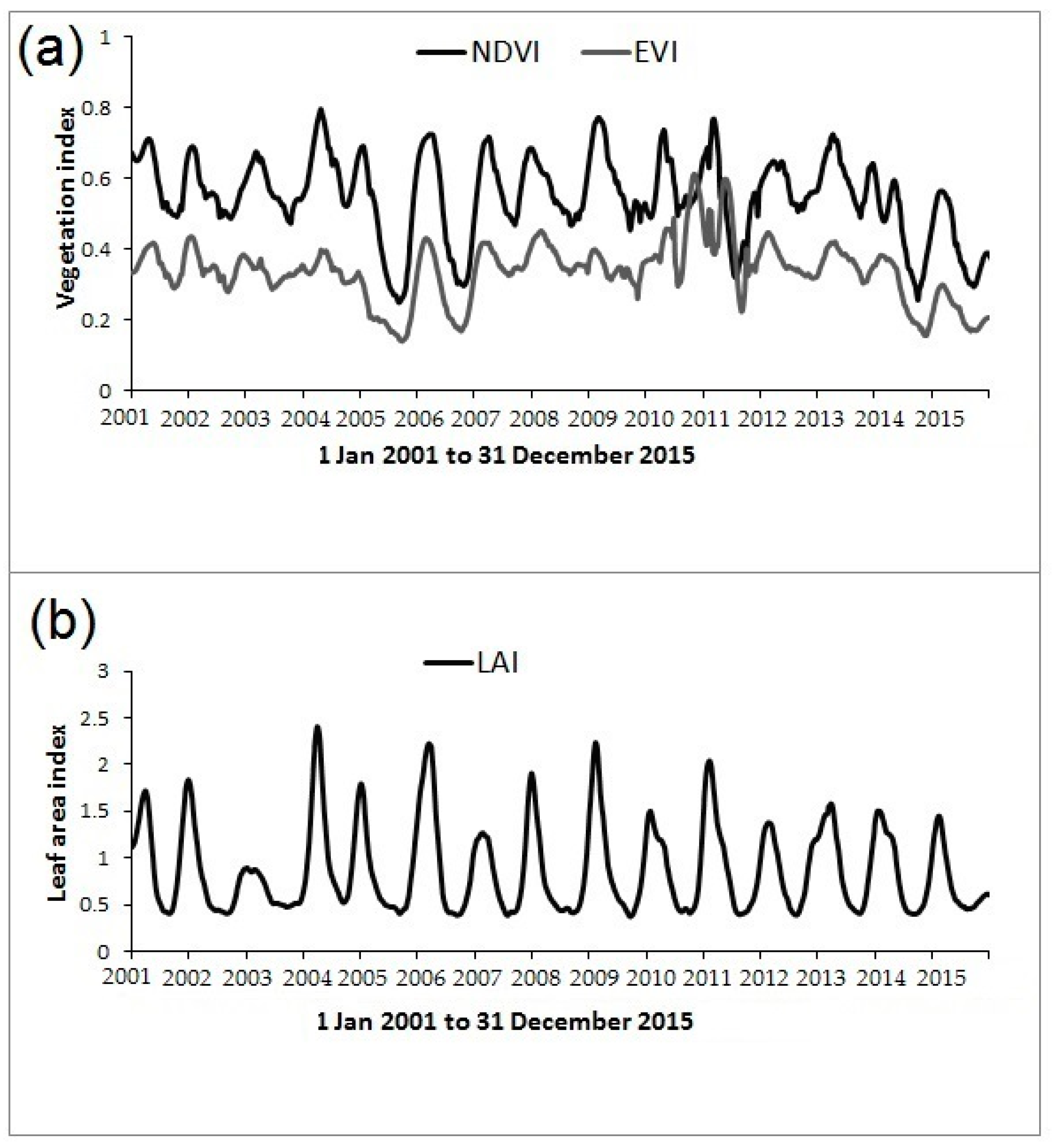
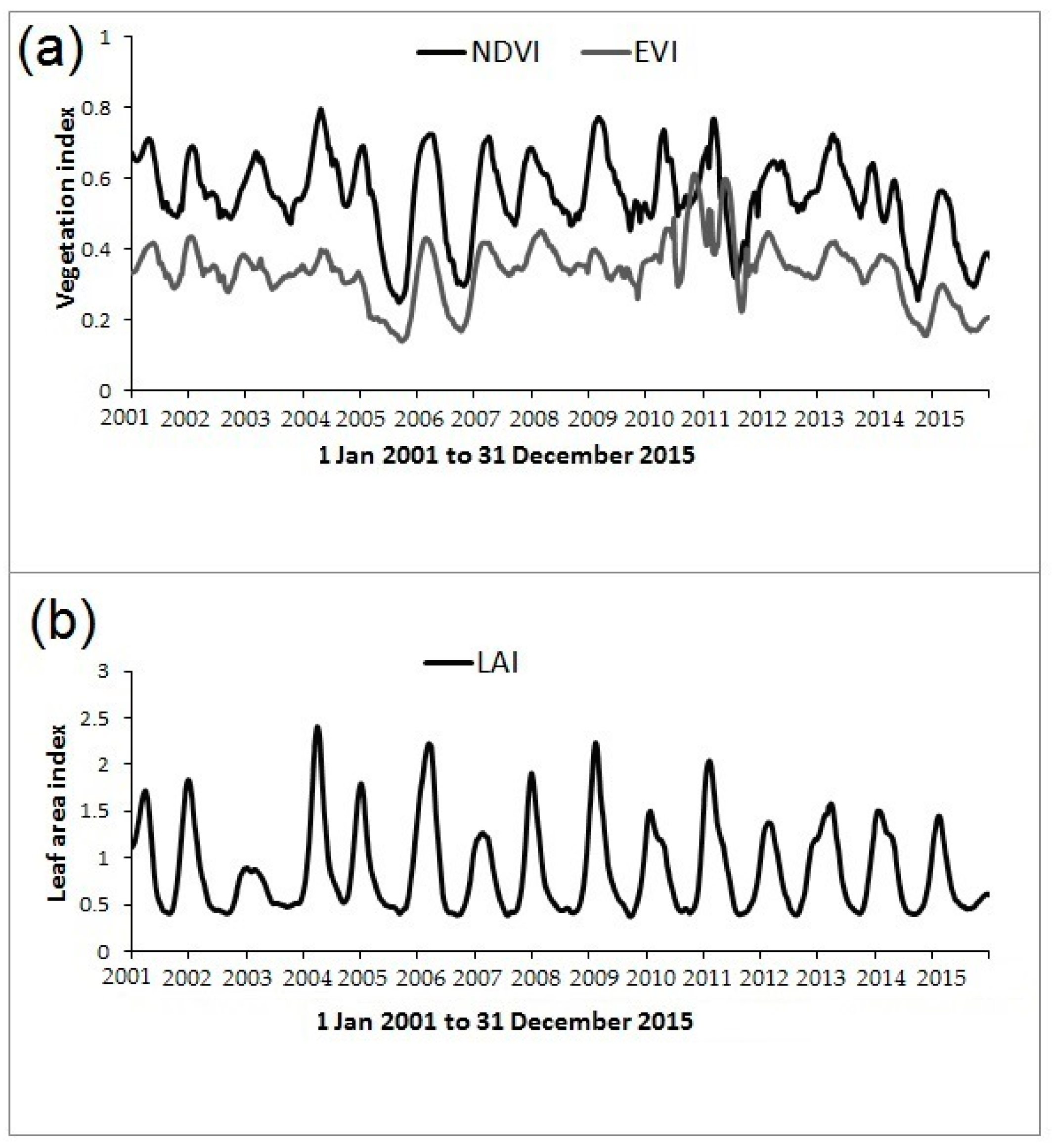
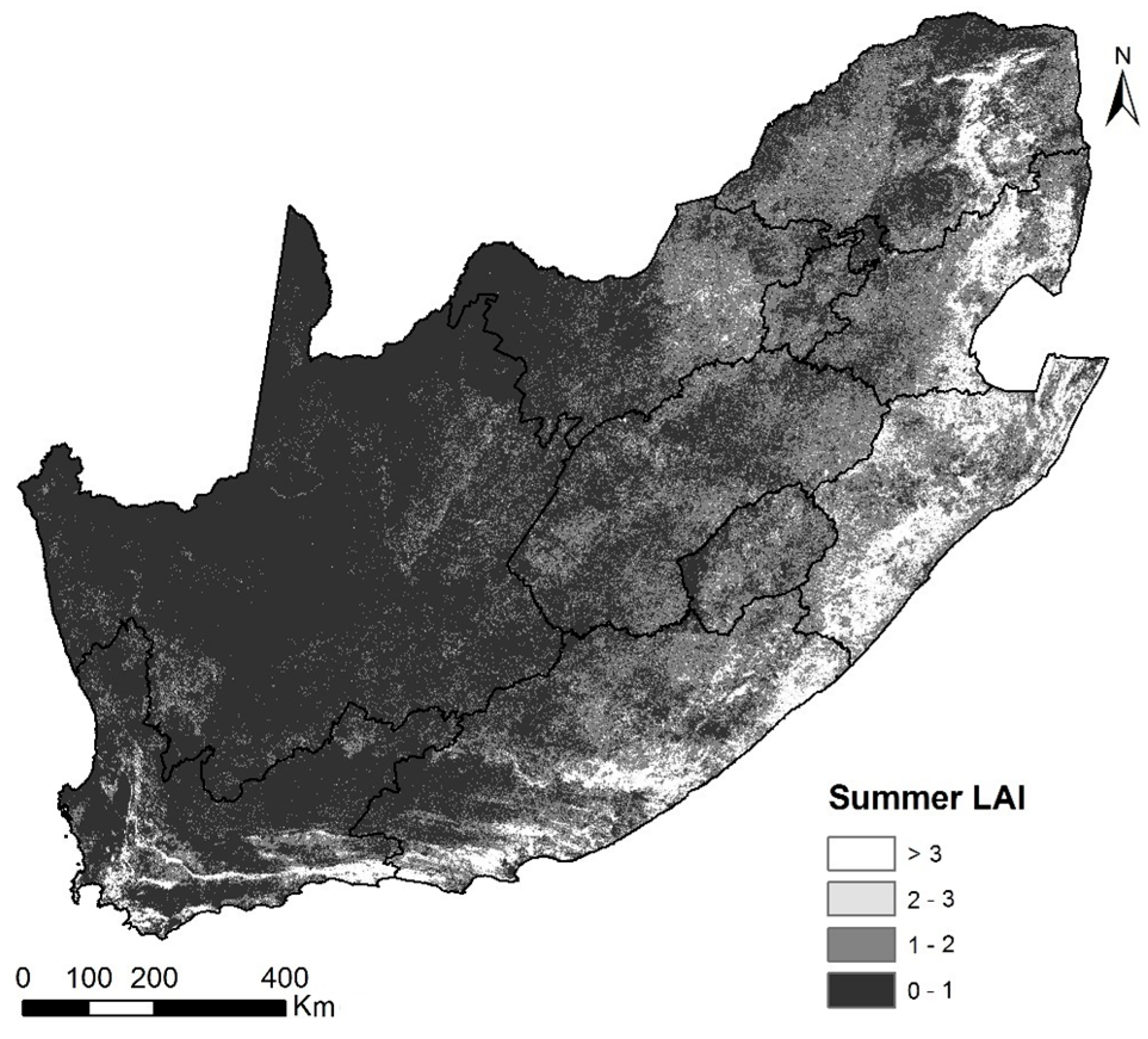
2.3. MODIS Imagery and Leaf Area Index (LAI) Time Series
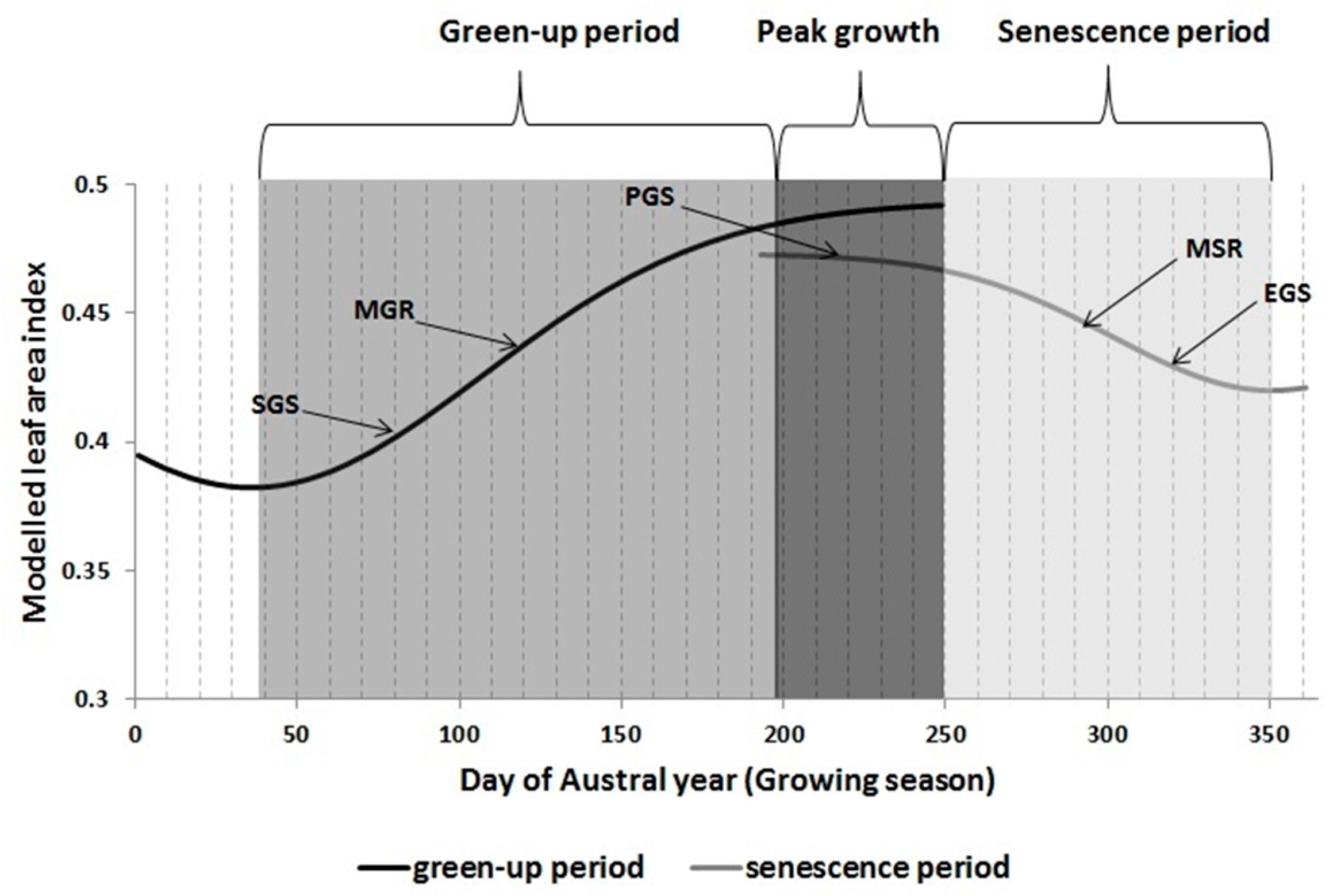
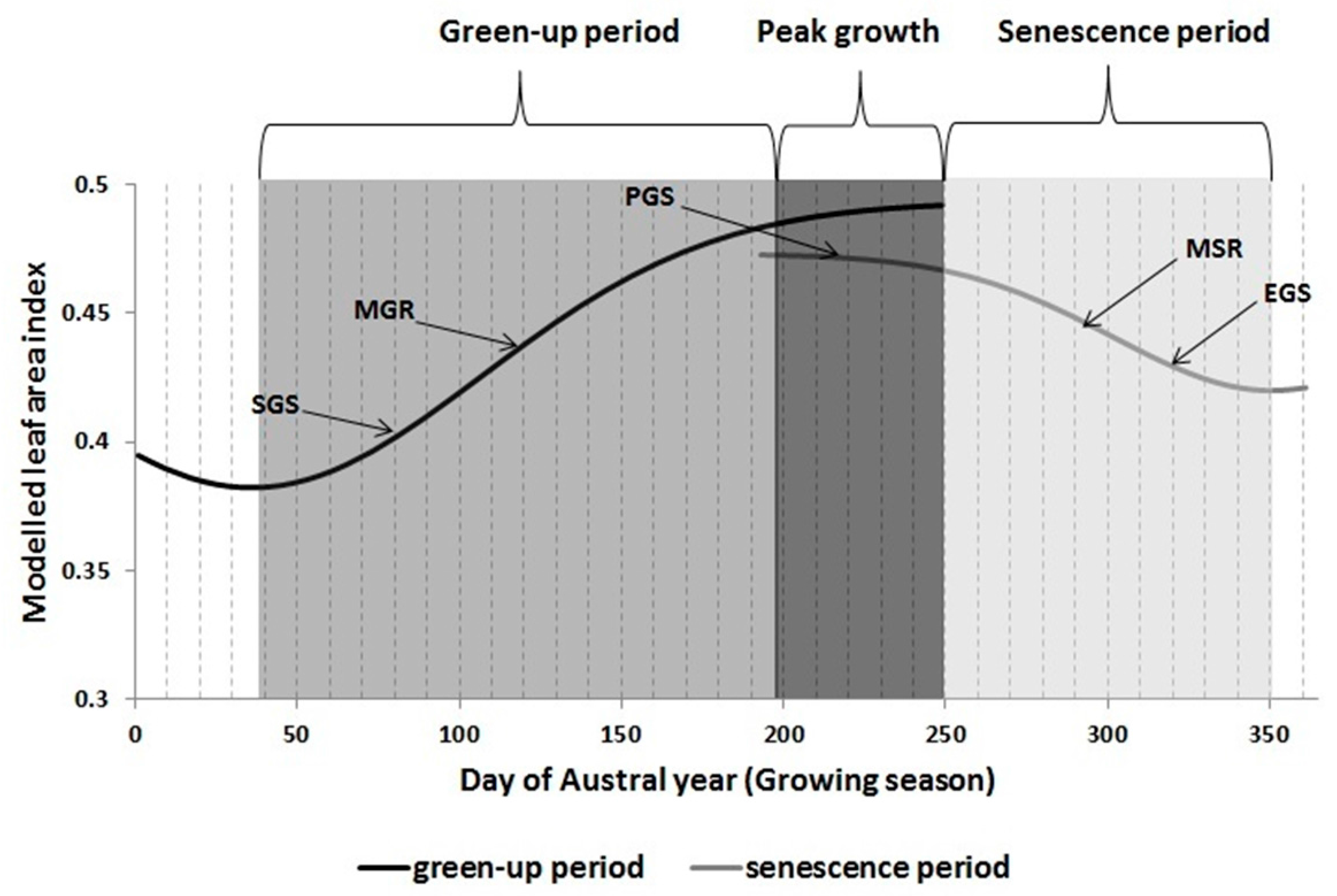
2.4. Determining Land Surface Phenological Metrics from LAI Time Series
- start of growing season (SGS)
- end of growing season (EGS)
- peak or maximum growth (PGS)
- maximum growing rate (MGR) i.e., the inflexion point on the growing phase
- maximum senescing rate (MSR) i.e., inflexion point on the senescing phase
2.5. Data Analysis
3. Results
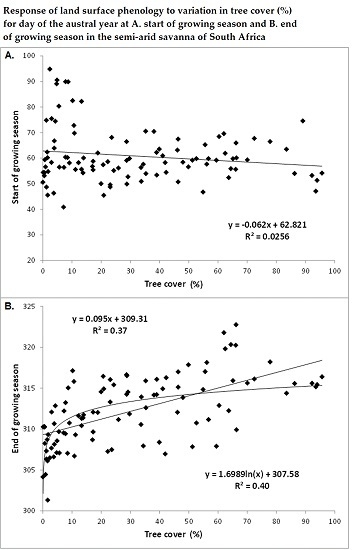
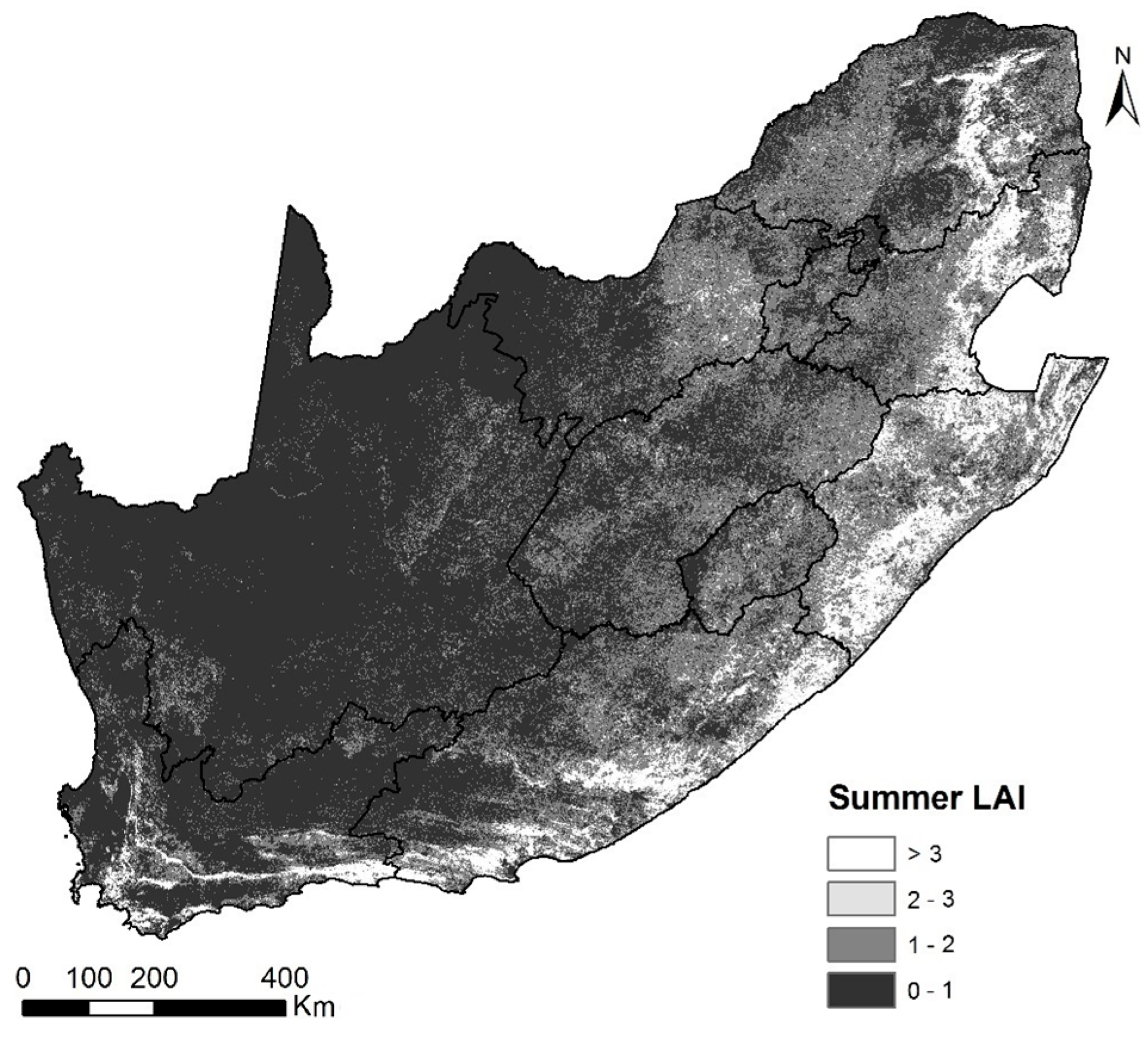
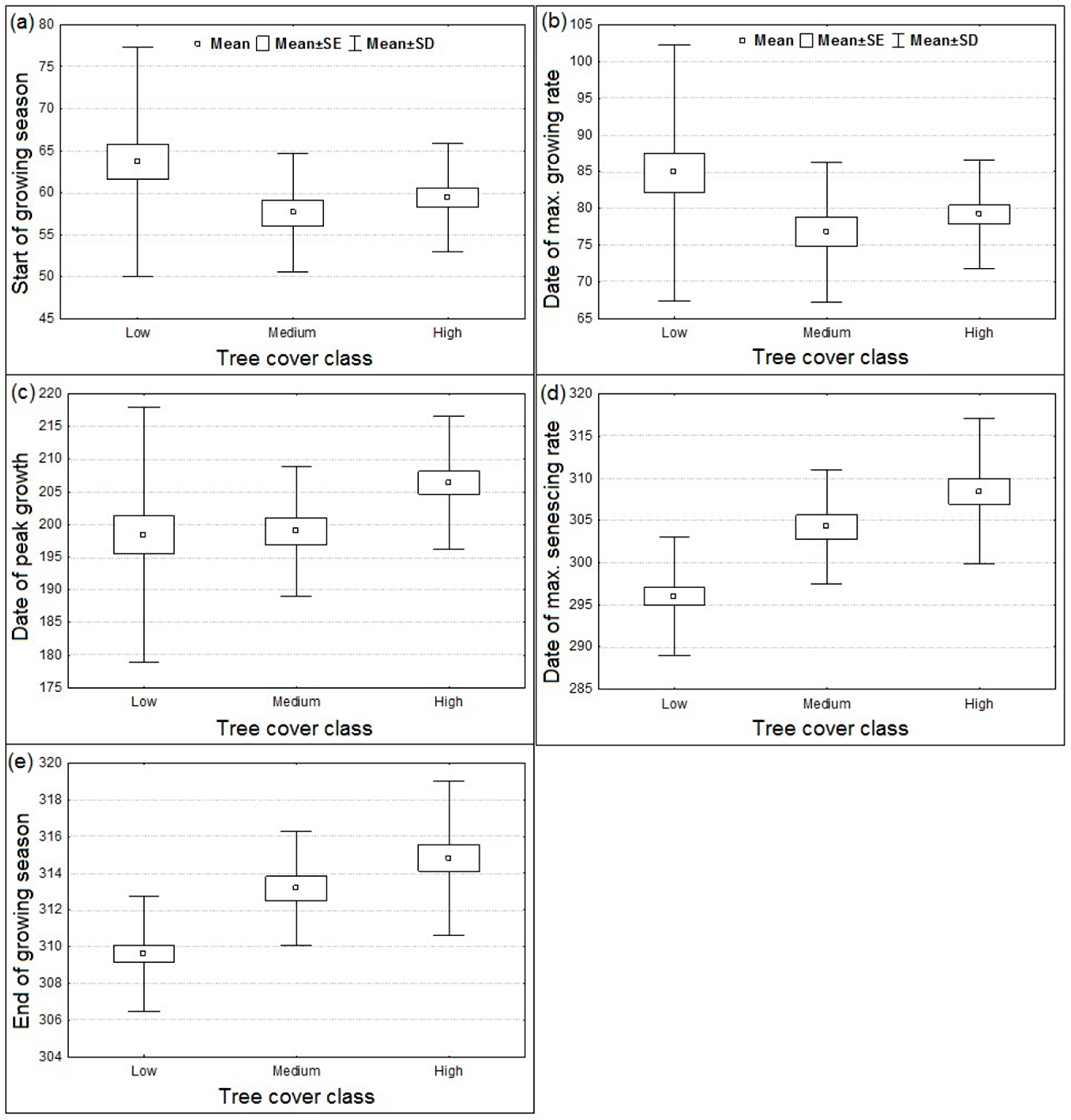
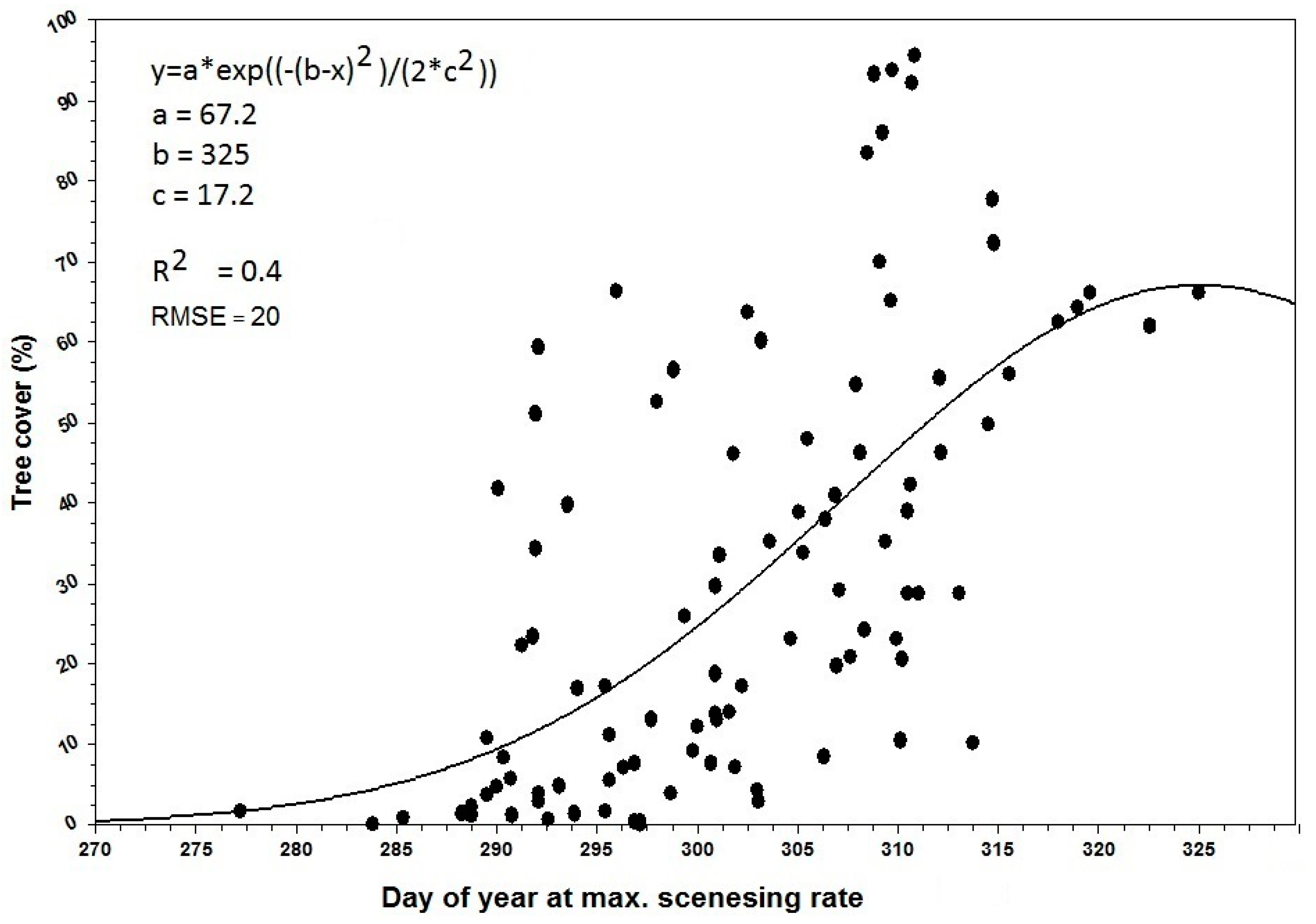
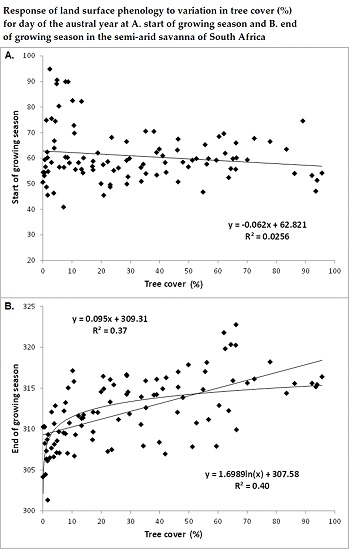
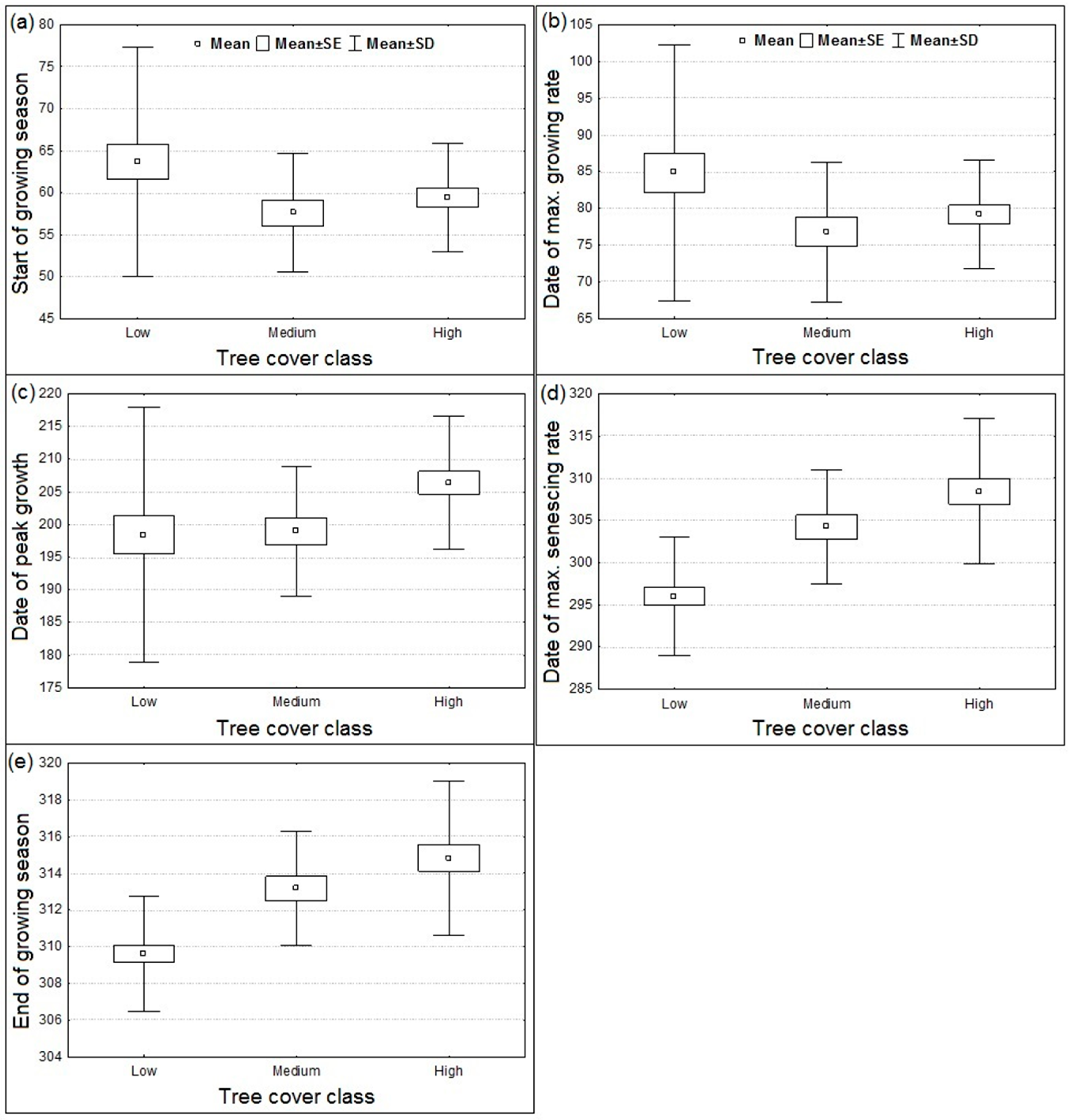
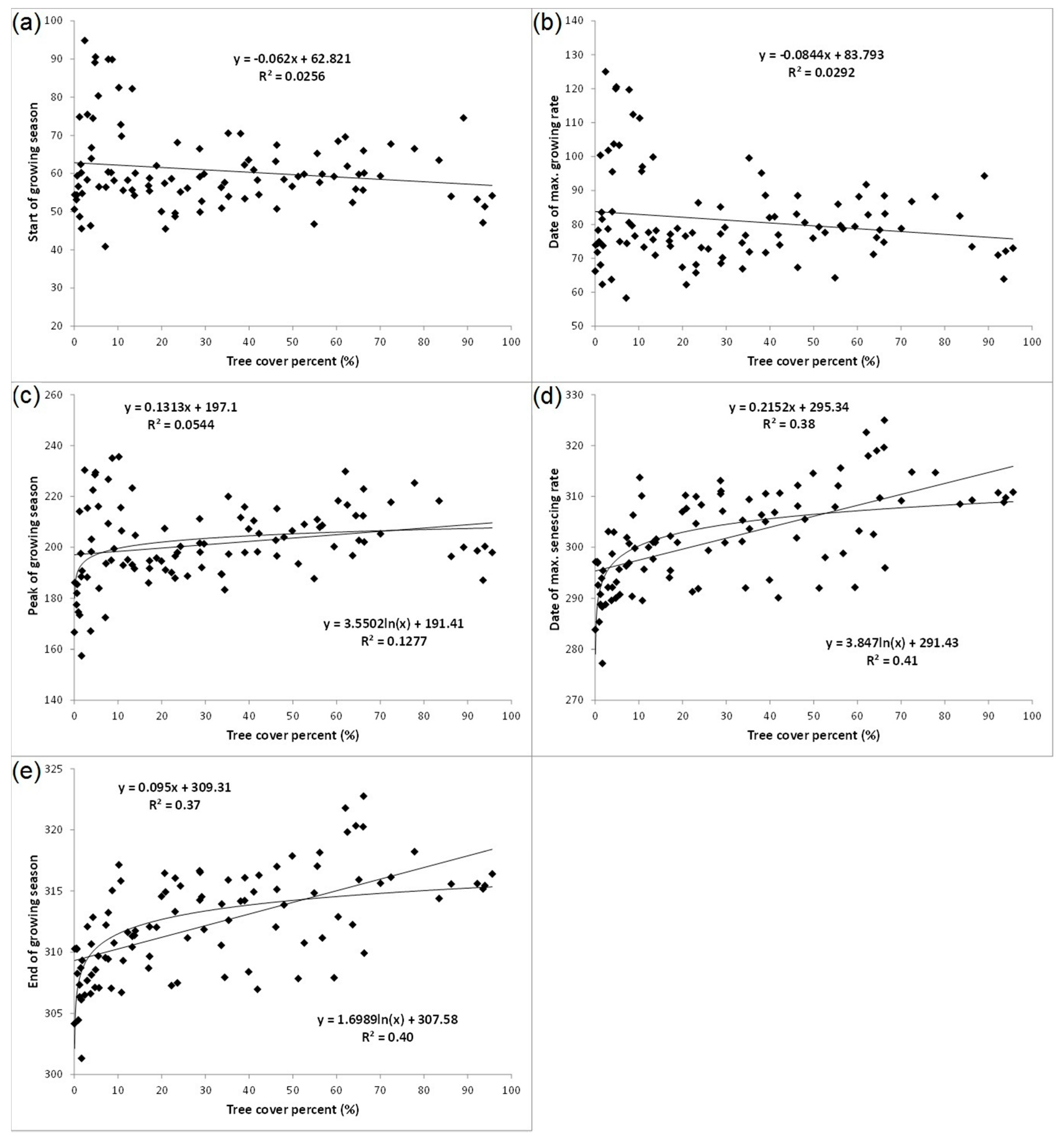
3.1. Spatial Patterns of Land Surface Phenological Metrics and Differences between Tree Cover (%) Classes
- -
- The vegetated landscape starts to green-up (day of the year at SGS) at about August 31 ± 2 days (95% CI).
- -
- The vegetated landscape reaches peak production (PGS) at about January 20 ± 3 days (95% CI).
- -
- The growing season ends (EGS) at about May 11 ± 1 day (95% CI).
3.2. Differences in Land Surface Phenological Metrics between Tree Cover Classes
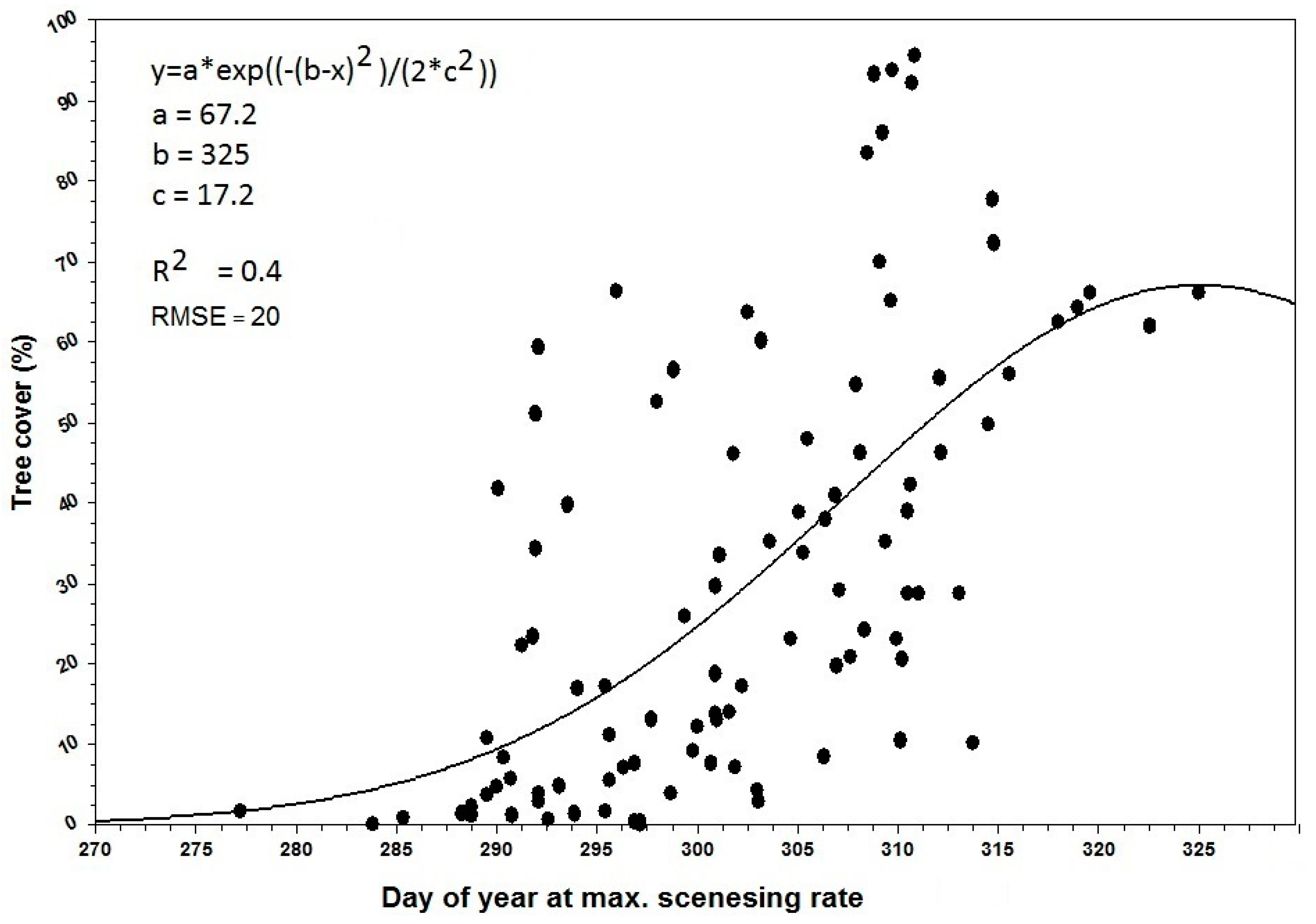
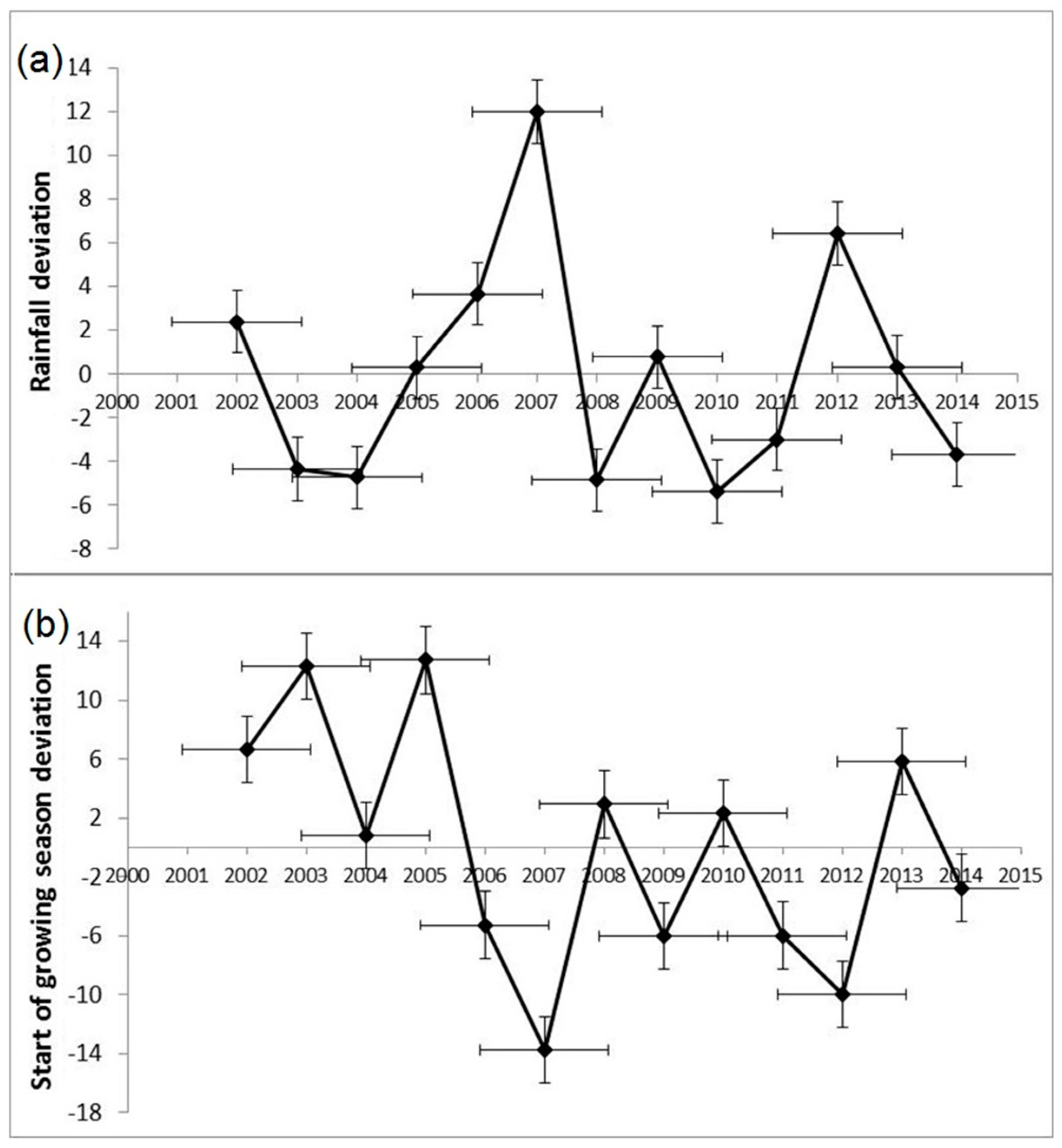
3.3. Inter-Annual Variability in Land Surface Phenology
3.4. Understanding Impact of Precipitation on Land Surface Phenology
4. Discussion
5. Conclusions
- Areas with tree cover greater than 20% showed an earlier day of the year at SGS or MGR in comparison to areas with tree cover of less than 20%, although the difference was statistically not significant at p < 0.05 due to the wide variation in day of the year at SGS or MGR among the low tree cover polygons located across the study region.
- There was a significant positive trend in the phenological metrics computed from the senescing phase, e.g., day of the year at MSR or EGS increases with increasing tree cover.
- Phenological metrics computed from the growing phase of the annual LAI time series showed higher inter-annual variability in comparison to the senescing phase metrics.
- Rainfall data for the region explained inter-annual variations in land surface phenological metrics during the green-up period of the vegetation, but not for the senescence period.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guan, K.; Wood, E.F.; Medvigy, D.; Kimball, J.; Pan, M.; Caylor, K.K.; Sheffield, J.; Xu, X.; Jones, M.O. Terrestrial hydrological controls on land surface phenology of African savannas and woodlands. J. Geophys. Res. Biogeosci. 2014, 119, 1652–1669. [Google Scholar] [CrossRef]
- Tan, B.; Morisette, J.T.; Wolfe, R.E.; Gao, F.; Ederer, G.A.; Nightingale, J.; Pedelty, J.A. An Enhanced TIMESAT Algorithm for Estimating Vegetation Phenology Metrics from MODIS Data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2011, 4, 361–371. [Google Scholar] [CrossRef]
- Suepa, T.; Qi, J.; Lawawirojwong, S.; Messina, J.P. Understanding spatio-temporal variation of vegetation phenology and rainfall seasonality in the monsoon Southeast Asia. Environ. Res. 2016, 147, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Spencer, H.; Weiblen, G.; Flick, B. Phenology of Ficus variegata in a seasonal wet tropical forest at Cape Tribulation, Australia. J. Biogeogr. 1996, 23, 467–475. [Google Scholar] [CrossRef]
- Damstra, K.S.J.; Richardson, S.; Reeler, B. Synchronized fruiting between trees of Ficus thonningii in seasonally dry habitats. J. Biogeogr. 1996, 23, 495–500. [Google Scholar] [CrossRef]
- Archibald, S.; Scholes, R.J. Leaf green-up in a semi-arid African savanna -separating tree and grass responses to environmental cues. J. Veg. Sci. 2007, 18, 583–594. [Google Scholar]
- White, M.A.; Thornton, P.E.; Running, S.W. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Jin, C.; Xiao, X.; Merbold, L.; Arneth, A.; Veenendaal, E.; Kutsch, W.L. Phenology and gross primary production of two dominant savanna woodland ecosystems in Southern Africa. Remote Sens. Environ. 2013, 135, 189–201. [Google Scholar] [CrossRef]
- Wessels, K.; Steenkamp, K.; von Maltitz, G.; Archibald, S. Remotely sensed vegetation phenology for describing and predicting the biomes of South Africa. Appl. Veg. Sci. 2011, 14, 49–66. [Google Scholar] [CrossRef]
- Scholes, R.J. Convex Relationships in Ecosystems Containing Mixtures of Trees and Grass. Environ. Resour. Econ. 2003, 26, 559–574. [Google Scholar] [CrossRef]
- Wakeling, J.L.; Cramer, M.D.; Bond, W.J. The savanna-grassland ‘treeline’: Why don’t savanna trees occur in upland grasslands? J. Ecol. 2012, 100, 381–391. [Google Scholar] [CrossRef]
- Ernst, W.H.O.; Veenendaal, E.M.; Kebakile, M.M. Possibilities for Dispersal in Annual and Perennial Grasses in a Savanna in Botswana. Vegetatio 1992, 102, 1–11. [Google Scholar] [CrossRef]
- Ramoelo, A.; Skidmore, A.K.; Cho, M.A.; Schlerf, M.; Mathieu, R.; Heitkönig, I.M.A. Regional estimation of savanna grass nitrogen using the red-edge band of the spaceborne RapidEye sensor. Int. J. Appl. Earth Obs. 2012, 19, 151–162. [Google Scholar] [CrossRef]
- Tomlinson, K.W.; Poorter, L.; Bongers, F.; Borghetti, F.; Jacobs, L.; Frank, V.L. Relative growth rate variation of evergreen and deciduous savanna tree species is driven by different traits. Ann. Bot. 2014, 114, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, R.; Naidoo, L.; Cho, M.A.; Leblon, B.; Main, R.; Wessels, K.; Asner, G.P.; Buckley, J.; Van Aardt, J.; Erasmus, B.F.N.; et al. Toward structural assessment of semi-arid African savannahs and woodlands: The potential of multitemporal polarimetric RADARSAT-2 fine beam images. Remote Sens. Environ. 2013, 138, 215–231. [Google Scholar] [CrossRef]
- Cho, M.A.; Debba, P.; Mathieu, R.; Naidoo, L.; Van Aardt, J.; Asner, G.P. Improving discrimination of savanna tree species through a multiple-endmember spectral angle mapper approach: Canopy-level analysis. IEEE Trans. Geosci. Remote 2010, 48, 4133–4142. [Google Scholar] [CrossRef]
- Chidumayo, E. Climate and phenology of savanna vegetation in southern Africa. J. Veg. Sci. 2001, 12, 347–354. [Google Scholar] [CrossRef]
- Shackleton, C. The main aim of this study was therefore to determine the influence of varying tree cover on the phenology of mixed remotely sensed pixel otherwise known as land surface phenology. Glob. Ecol. Biogeogr. 1999, 8, 125–136. [Google Scholar] [CrossRef]
- Sankaran, M.H.N.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; Ardo, J.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Whitecross, M.A.; Witkowski, E.T.F.; Archibald, S. Savanna tree-grass interactions: A phenological investigation of green-up in relation to water availability over three seasons. S. Afr. J. Bot. 2017, 108, 29–40. [Google Scholar] [CrossRef]
- Jolly, W.M.; Running, S.W. Effects of precipitation and soil water potential on drought deciduous phenology in the Kalahari. Glob. Chang. Biol. 2004, 10, 303–308. [Google Scholar] [CrossRef]
- Fuller, D.O. Canopy phenology of some mopane and miombo woodlands in eastern Zambia. Glob. Ecol. Biogeogr. 1999, 8, 199–209. [Google Scholar] [CrossRef]
- Verbesselt, J.; Somers, B.; Lhermitte, J.; van Aardt, J.; Jonckheere, I.; Coppin, P. Fire risk assessment in savanna ecosystems with multi-temporal satellite data. Commun. Agric. Appl. Biol. Sci. 2005, 70, 23–26. [Google Scholar] [PubMed]
- Verbesselt, J.; Jönsson, P.; Lhermitte, J.; van Aardt, J.; Jonckheere, I.; Coppin, P. Evaluating satellite and climate data-derived indices as fire risk indicators in savanna ecosystems. IEEE Trans. Geosci. Remote Sens. 2006, 44, 1622–1632. [Google Scholar] [CrossRef]
- Bajocco, S.; Dragoz, E.; Gitas, I.; Smiraglia, D.; Salvati, L.; Ricotta, C. Mapping Forest Fuels through Vegetation Phenology: The Role of Coarse-Resolution Satellite Time-Series. PLoS ONE 2015, 10, e0119811. [Google Scholar] [CrossRef] [PubMed]
- Staver, A.C.; Archibald, S.; Levin, S. Tree cover in sub-Saharan Africa: rainfall and fire constrain forest and savanna as alternative states. Ecology 2011, 92, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; South African Biodiversity Institute: Pretoria, South Africa, 2006; Volume 19. [Google Scholar]
- Duhl, T.R.; Guenther, A.; Helmig, D. Estimating urban vegetation cover fraction using Google Earth® images. J. Land Use Sci. 2012, 7, 311–329. [Google Scholar] [CrossRef]
- Ludwig, A.; Meyer, H.; Nauss, T. Automatic classification of Google Earth images for a larger scale monitoring of bush encroachment in South Africa. Int. J. Appl. Earth Obs. 2016, 50, 89–94. [Google Scholar] [CrossRef]
- Pan, Z.; Huang, J.; Zhou, Q.; Wang, L.; Cheng, Y.; Zhang, H.; Blackburn, G.A.; Yan, J.; Liu, J. Mapping crop phenology using NDVI time-series derived from HJ-1 A/B data. Int. J. Appl. Earth Obs. 2015, 34, 188–197. [Google Scholar] [CrossRef]
- Wu, C.; Peng, D.; Soudani, K.; Siebicke, L.; Gough, C.M.; Arain, M.A.; Bohrer, G.; Lafleur, P.M.; Peichl, M.; Gonsamo, A.; et al. Land surface phenology derived from normalized difference vegetation index (NDVI) at global FLUXNET sites. Agric. For. Meteorol. 2017, 233, 171–182. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Cho, M.A.; Ramoelo, A.; Mathieu, R. Estimation of leaf area index (LAI) of South Africa from MODIS imager by inversion of PROSAIL radiative transfer model. In Proceedings of the 2014 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Quebec City, QC, Canada, 13–18 July 2014; pp. 2590–2593. [Google Scholar]
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Bradley, B.A.; Jacob, R.W.; Hermance, J.F.; Mustard, J.F. A curve fitting procedure to derive inter-annual phenologies from time series of noisy satellite NDVI data. Remote Sens. Environ. 2007, 106, 137–145. [Google Scholar] [CrossRef]
- Groten, S.M.E.; Ocatre, R. Monitoring the length of the growing season with NOAA. Int. J. Remote Sens. 2002, 23, 2797–2815. [Google Scholar] [CrossRef]
- Scholes, R.J. Woodlands of South Africa. In Indigenous Forest and Woodlands in South Africa: Policy People and Practices; Lawes, M.J., Eeley, H.A.C., Shackleton, C.M., Geach, B.G.S., Eds.; University of KwaZulu-Natal Press: Pietermaritzburg, South Africa, 2004. [Google Scholar]
- Guan, K.; Medvigy, D.; Wood, E.F.; Caylor, K.K.; Li, S.; Jeong, S.J. Deriving Vegetation Phenological Time and Trajectory Information Over Africa Using SEVIRI Daily LAI. IEEE Trans. Geosci. Remote Sens. 2014, 52, 1113–1130. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Effects of climate on the growth of exotic and indigenous trees in central Zambia. J. Biogeogr. 2005, 32, 111–120. [Google Scholar] [CrossRef]
- Schulze, R.E. South African Atlas of Climatology and Agrohydrology; WRC Report 1489/06; Water Research Commission: Pretoria, South Africa, 2007. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Austral Year | Green-Up Period | Senescence Period (in Reverse Order) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| July–August | July–September | July–October | July–November | July–December | June–May | June–April | June–March | June–February | June–January | |
| 2001–2002 | 15.3 | 21.6 | 24.4 | 28.5 | 32.5 | |||||
| 2002–2003 | 8.0 | 10.1 | 27.2 | 30.9 | 43.4 | 14.1 | 10.4 | 18.7 | 23.2 | 37.0 |
| 2003–2004 | 0.3 | 3.3 | 10.8 | 22.3 | 26.5 | 2.4 | 12.6 | 36.3 | 45.0 | 62.6 |
| 2004–2005 | 1.8 | 2.9 | 2.2 | 8.0 | 35.6 | 4.6 | 17.5 | 26.3 | 26.7 | 42.0 |
| 2005–2006 | 7.6 | 8.0 | 12.4 | 30.8 | 34.2 | 11.7 | 49.7 | 37.3 | 49.7 | 74.2 |
| 2006–2007 | 15.1 | 11.3 | 15.1 | 28.0 | 41.7 | 7.3 | 22.4 | 22.2 | 23.3 | 29.9 |
| 2007–2008 | 8.1 | 19.7 | 48.0 | 62.4 | 69.3 | 24.6 | 26.5 | 37.0 | 37.7 | 61.5 |
| 2008–2009 | 3.6 | 2.8 | 22.2 | 36.1 | 45.1 | 12.1 | 18.8 | 38.1 | 32.1 | 77.7 |
| 2009–2010 | 8.9 | 8.4 | 20.9 | 36.3 | 49.1 | 9.8 | 43.2 | 48.5 | 50.7 | 65.1 |
| 2010–2011 | 3.0 | 2.3 | 13.2 | 28.9 | 42.3 | 3.1 | 23.6 | 26.0 | 26.1 | 46.6 |
| 2011–2012 | 4.8 | 4.7 | 17.8 | 27.3 | 47.2 | 2.8 | 7.7 | 20.2 | 21.3 | 43.1 |
| 2012–2013 | 0.9 | 14.1 | 26.6 | 36.2 | 54.3 | 2.9 | 22.9 | 28.0 | 32.1 | 47.9 |
| 2013–2014 | 2.4 | 8.0 | 17.8 | 26.2 | 46.2 | 2.0 | 11.5 | 44.6 | 39.4 | 61.5 |
| 2014–2015 | 5.7 | 4.0 | 12.5 | 22.0 | 43.3 | 5.2 | 12.8 | 31.0 | 36.8 | 48.0 |
| 2015–2016 | 2.6 | 7.0 | 9.7 | 12.9 | 23.4 | |||||
| Tree Cover Class | Mean ± 95% CI | Standard Deviation | Coefficient of Variation (%) |
|---|---|---|---|
| a. Day of year at start of growing season | |||
| Low | 64 ± 4.1 | 14 | 22 |
| Medium | 58 ± 2.6 | 7 | 12 |
| High | 59 ± 3.3 | 6 | 10 |
| Mean | 60 ± 2.1 | 9 | 15 |
| b. Day of year at maximum growth rate | |||
| Low | 85 ± 5.2 | 17 | 20 |
| Medium | 77 ± 3.6 | 9 | 12 |
| High | 79 ± 3.9 | 7 | 9 |
| Mean | 80 ± 2.7 | 11 | 14 |
| c. Day of year at peak growth | |||
| Low | 198 ± 5.5 | 20 | 10 |
| Medium | 199 ± 3.9 | 10 | 5 |
| High | 206 ± 4.9 | 10 | 5 |
| Mean | 201 ± 2.7 | 13 | 7 |
| d. Day of year at maximum senescing rate | |||
| Low | 296 ± 2.2 | 7 | 2 |
| Medium | 304 ± 2.9 | 7 | 2 |
| High | 308 ± 3.5 | 9 | 3 |
| Mean | 303 ± 1.7 | 8 | 3 |
| e. Day of year at end of growing season | |||
| Low | 310 ± 1.0 | 3 | 1 |
| Medium | 313 ± 1.3 | 3 | 1 |
| High | 315 ± 1.7 | 4 | 1 |
| Mean | 313 ± 0.8 | 3 | 1 |
| Phenological Metrics | t-Value for Differences in Means | p for Differences in Means | F-Ratio for Differences in Variances | p for Differences Variances |
|---|---|---|---|---|
| a. Day of year at start of growing season | ||||
| Low vs. Medium | 1.972 | 0.053 | 3.718 | 0.002 |
| Low vs. High | 1.695 | 0.094 | 4.438 | 0.001 |
| Medium vs. High | −0.991 | 0.326 | 1.194 | 0.634 |
| b. Day of the year at maximum growth rate | ||||
| Low vs. Medium | 2.012 | 0.048 | 3.376 | 0.004 |
| Low vs. High | 1.774 | 0.080 | 5.531 | 0.001 |
| Medium vs. High | −1.044 | 0.301 | 1.638 | 0.199 |
| c. Day of the year at peak growth | ||||
| Low vs. Medium | −0.122 | 0.904 | 3.918 | 0.001 |
| Low vs. High | −2.171 | 0.033 | 3.666 | 0.001 |
| Medium vs. High | −2.711 | 0.009 | 1.068 | 0.892 |
| d. Day of the year at maximum senescence rate | ||||
| Low vs. Medium | −4.495 | 0.001 | 1.071 | 0.890 |
| Low vs. High | −6.944 | 0.001 | 1.492 | 0.220 |
| Medium vs. High | −1.928 | 0.059 | 1.598 | 0.263 |
| e. Day of the year at end of growing season | ||||
| Low vs. Medium | −4.324 | 0.001 | 1.030 | 0.972 |
| Low vs. High | −6.261 | 0.001 | 1.787 | 0.074 |
| Medium vs. High | −1.605 | 0.114 | 1.841 | 0.145 |
| A. Green-Up Period | July–August | July–September | July–October | July–November | July–December |
| a. Start of growing season | −0.27 | −0.57 * | −0.55 | −0.50 | −0.82 * |
| b. Date of max. growth rate | −0.28 | −0.53 | −0.50 | −0.46 | −0.80 * |
| c. Date of peak production | −0.43 | −0.62 * | −0.75 * | −0.71 * | −0.83 * |
| B. Senescence Period | June–May | June–April | June–March | June–February | June–January |
| d. Date of max. senescing rate | −0.41 | 0.15 | 0.10 | 0.12 | −0.02 |
| e. End of growing season | −0.35 | 0.12 | 0.09 | 0.09 | −0.02 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, M.A.; Ramoelo, A.; Dziba, L. Response of Land Surface Phenology to Variation in Tree Cover during Green-Up and Senescence Periods in the Semi-Arid Savanna of Southern Africa. Remote Sens. 2017, 9, 689. https://doi.org/10.3390/rs9070689
Cho MA, Ramoelo A, Dziba L. Response of Land Surface Phenology to Variation in Tree Cover during Green-Up and Senescence Periods in the Semi-Arid Savanna of Southern Africa. Remote Sensing. 2017; 9(7):689. https://doi.org/10.3390/rs9070689
Chicago/Turabian StyleCho, Moses A., Abel Ramoelo, and Luthando Dziba. 2017. "Response of Land Surface Phenology to Variation in Tree Cover during Green-Up and Senescence Periods in the Semi-Arid Savanna of Southern Africa" Remote Sensing 9, no. 7: 689. https://doi.org/10.3390/rs9070689