Effects of Resveratrol Supplementation on Bone Growth in Young Rats and Microarchitecture and Remodeling in Ageing Rats


 and
and
Abstract
:1. Introduction
2. Experimental Section
2.1. Animal Trials and Specimen Collection
2.2. Bone Volume and Micro-Architecture
2.3. Histomorphometric Analysis of Metaphysis
2.4. Quantitative RT-PCR (qRT-PCR) Analysis of Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| CycA | GAGCTGTTTGCAGACAAAGTTC | CCTGGCACATGAATCCTGG |
| Osterix | GCTTTTCTGTGGCAAGAGGTTC | CTGATGTTTCTCAAGTGGTCG |
| Osteocalcin | AAGCCTTCATGTCCAAGCAGG | AGGCGGTGTTGAAGCCATACT |
| C/EBPα | TCGCCATGCCGGGAGAACTCTAAC | CTGGAGGTGGCTGCTCATCGGGG |
| Sirt 1 | AGA AACAATTCCTCCACCTGA | GCTTTGGTGGTTCTGAAAGG |
| FABP4 | GGAATTCGATGAAATCACCCC | TGGTCGACTTTCCATCCCACT |
2.5. Measurement of Collagen Type I Cross-Linked C-Telopeptide (CTX-1) and Alkaline Phosphotase (ALP) in Serum
2.6. Statistical Analysis
3. Results
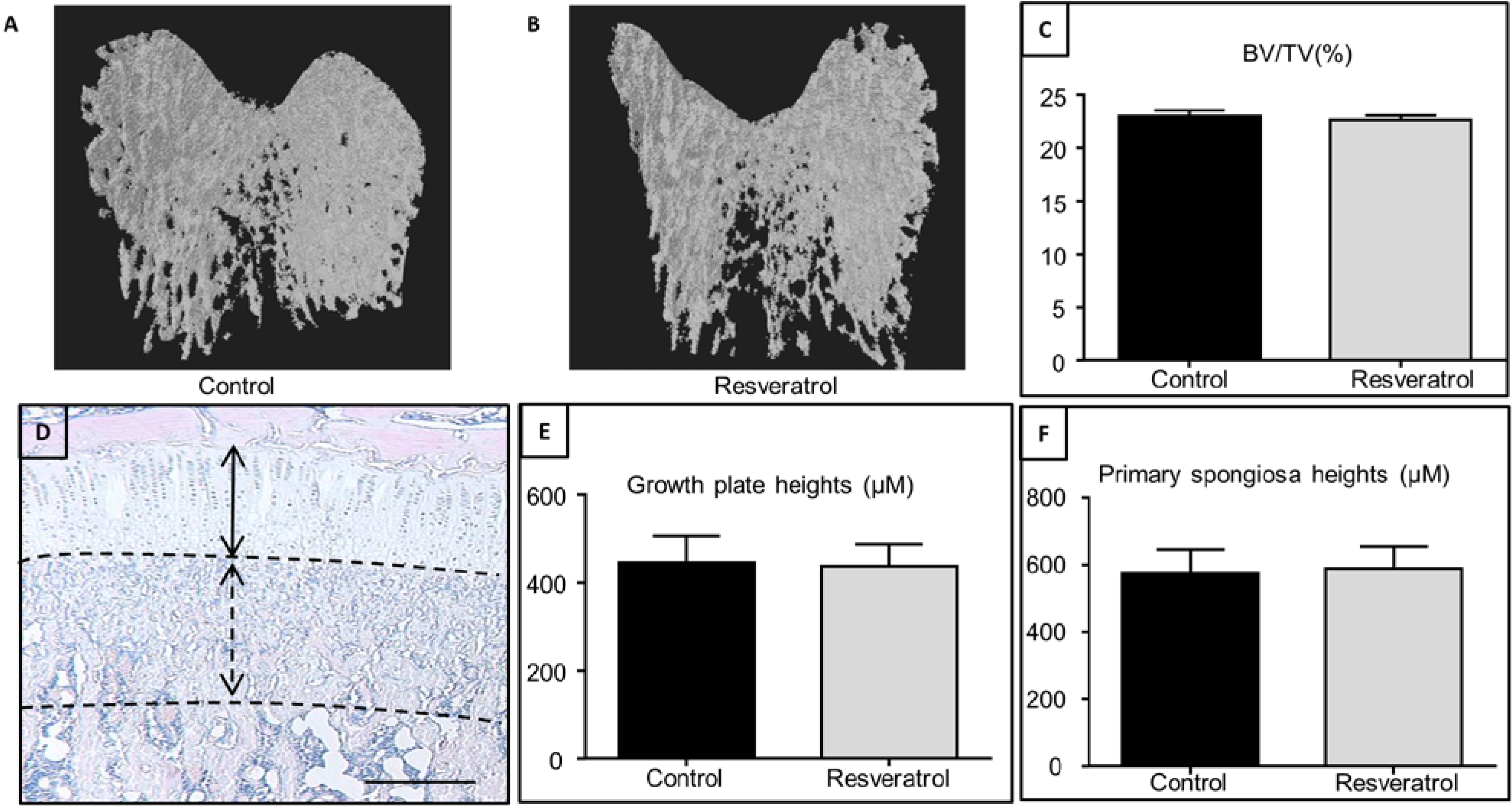
3.1. Effects of Resveratrol Supplementation in Young Animals

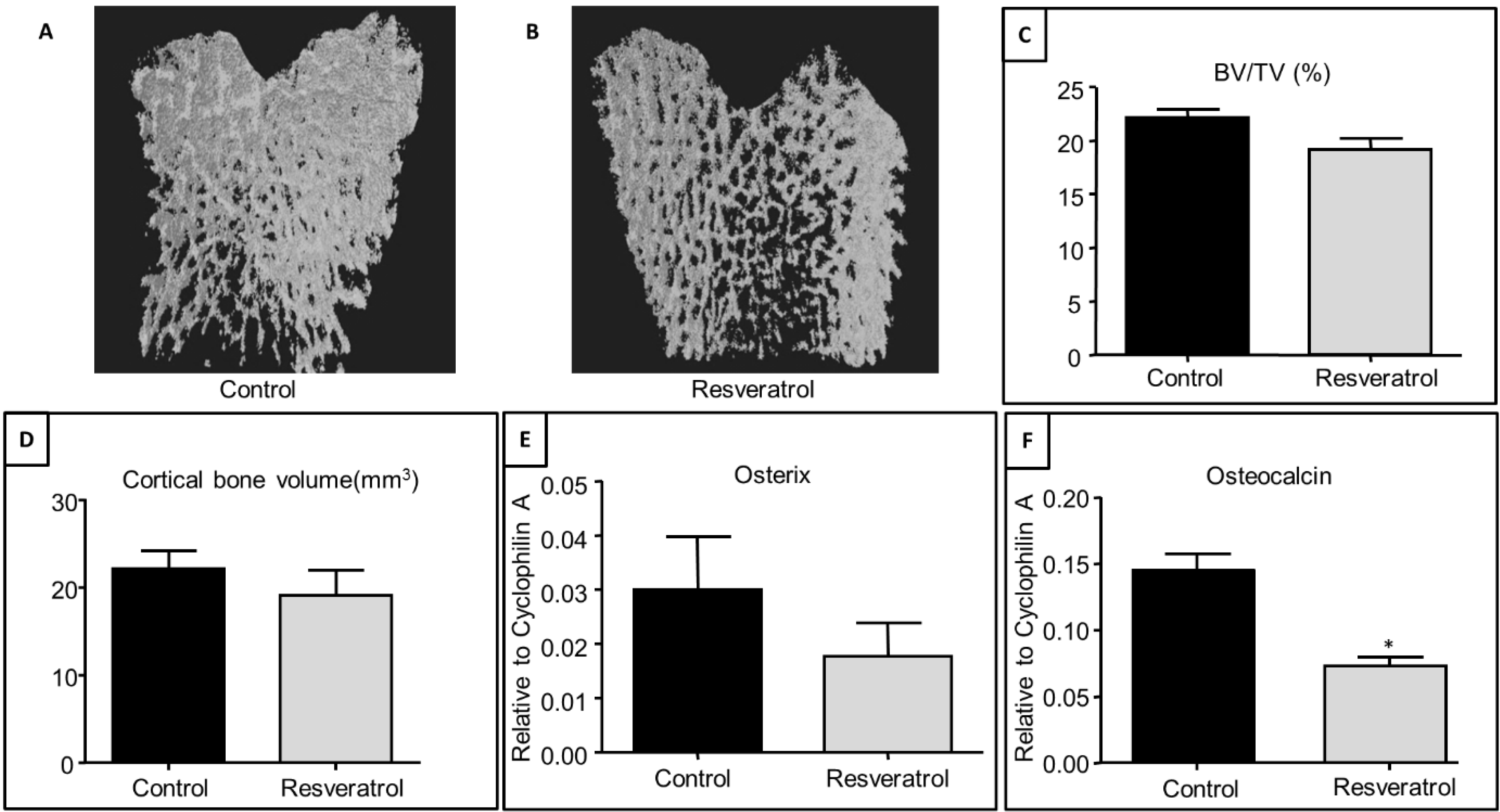
3.2. Effects of Resveratrol Supplementation on Bone Structure and Osteogenesis Regulatory Genes in Ageing Rats

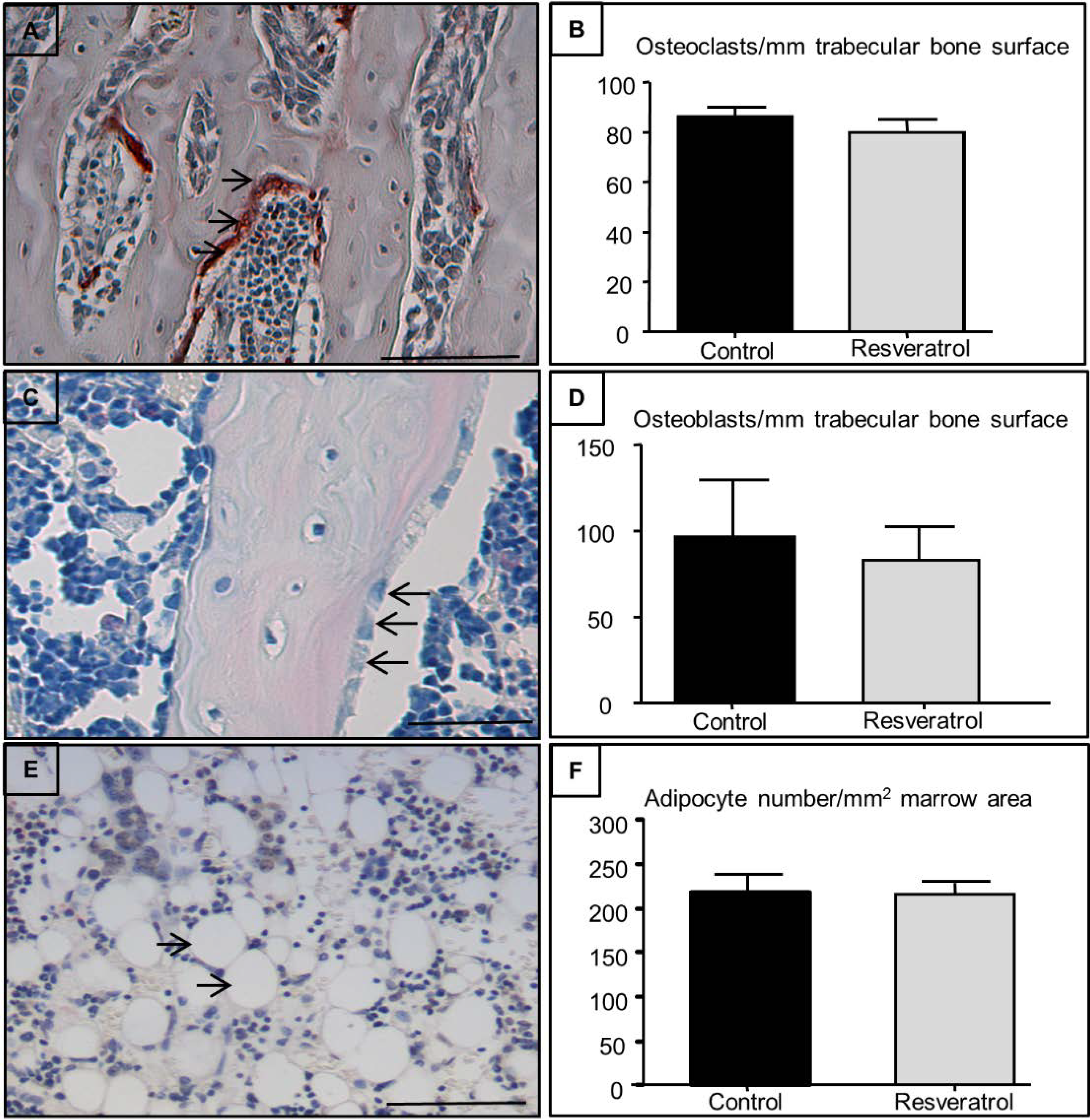
3.3. Effects on Osteoclast, Osteoblast, and Adipocyte Densities in the Secondary Spongiosa of Ageing Rats

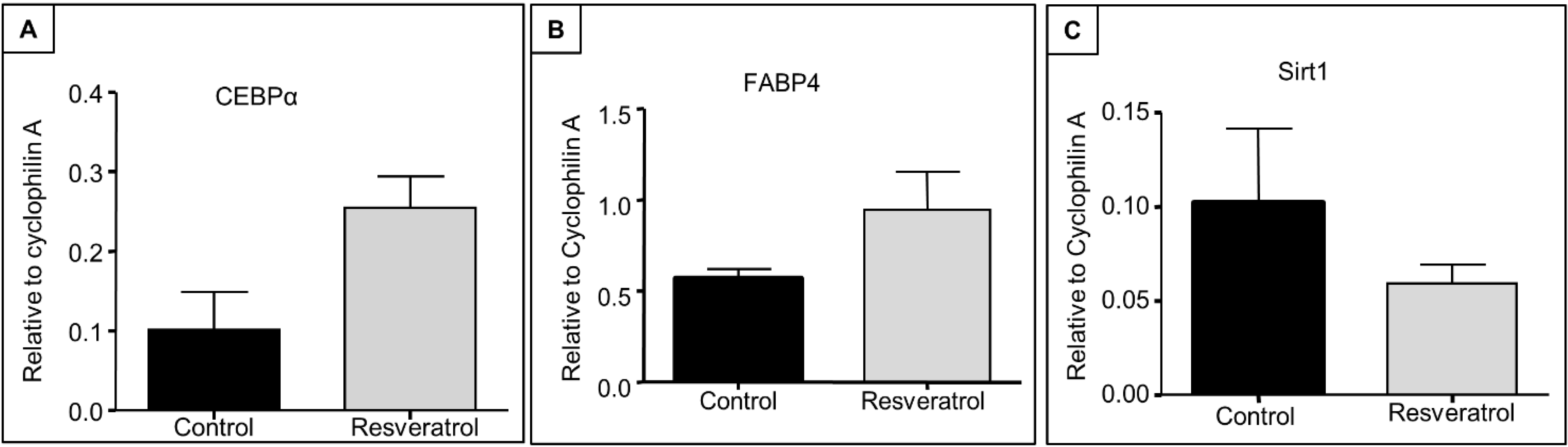
3.4. Expression of Adipogenesis Regulatory Genes and Sirt 1 in Early Ageing Rats

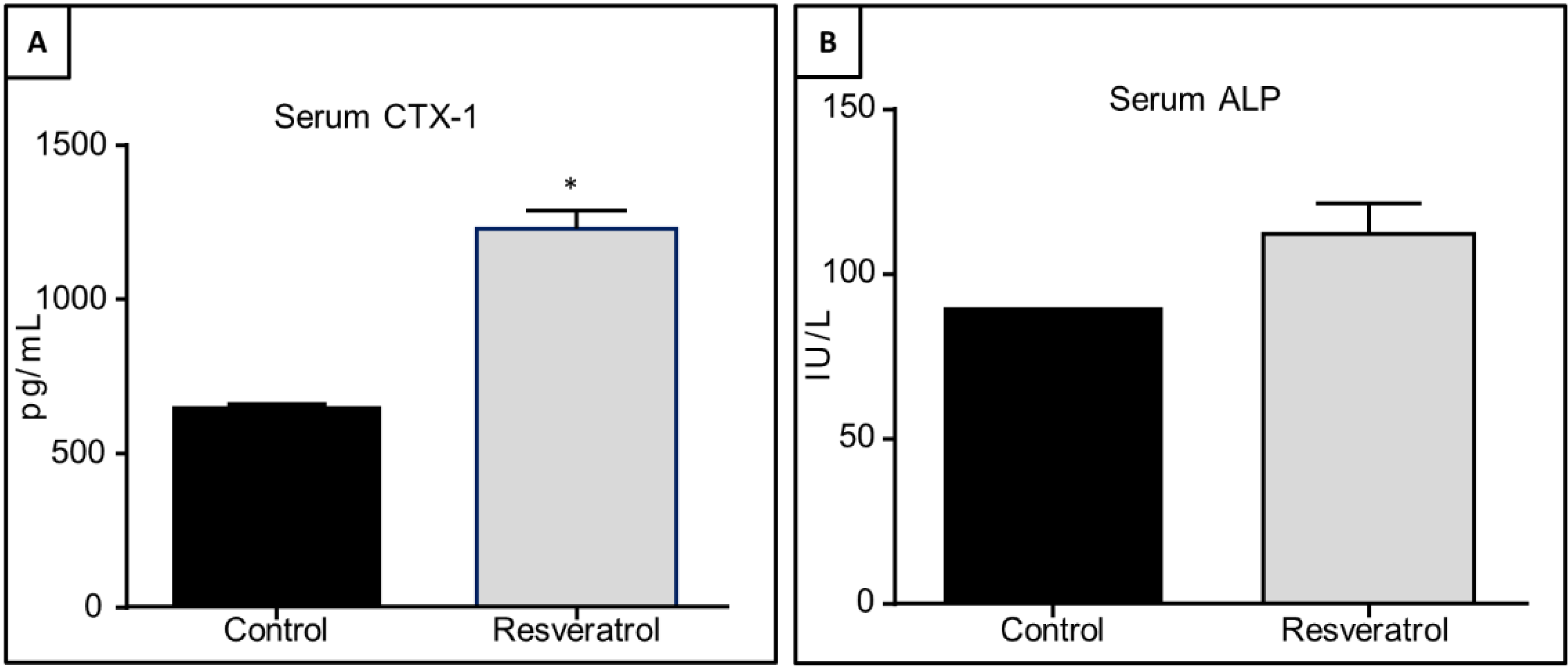
3.5. Effects of Resveratrol Supplementation in Serum Levels of CTX-1 and ALP in Ageing Rats

4. Discussion
4.1. Effects of Resveratrol Supplementation on Peak Bone Mass
4.2. Potential Effects of Resveratrol on Ageing-Induced Bone Loss
4.3. Effects of Resveratrol Supplementation on Gene Expression in the Bone
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kanis, J.A.; Brazier, J.E.; Stevenson, M.; Calvert, N.W.; Lloyd Jones, M. Treatment of established osteoporosis: A systematic review and cost-utility analysis. Health Technol. Assess. 2002, 6, 1–146. [Google Scholar] [CrossRef] [PubMed]
- Johnell, O. The socioeconomic burden of fractures: Today and in the 21st century. Am. J. Med. 1997, 103, 20–26. [Google Scholar] [CrossRef]
- Khosla, S.; Bilezikian, J.P.; Dempster, D.W.; Lewiecki, E.M.; Miller, P.D.; Neer, R.M.; Recker, R.R.; Shane, E.; Shoback, D.; Potts, J.T. Benefits and risks of bisphosphonate therapy for osteoporosis. J. Clin. Endocrinol. Metab. 2012, 97, 2272–2282. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Crockett, J.C. Osteoporosis—A current view of pharmacological prevention and treatment. Drug Des. Dev. Ther. 2006, 7, 435–448. [Google Scholar]
- WHO Study Group. Assessment of fracture risk and its application to screening for postmenopausal osteoporosis. World Health Organ. Tech. Rep. Ser. 1994, 843, 1–129. [Google Scholar]
- Cummings, S.R.; Black, D.M.; Nevitt, M.C.; Browner, W.; Cauley, J.; Ensrud, K.; Genant, H.K.; Palermo, L.; Scott, J.; Vogt, T.M. Bone density at various sites for prediction of hip fractures. The Study of Osteoporotic Fractures Research Group. Lancet 1993, 341, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.; Johnell, O.; Wedel, H. Meta-analysis of how well measures of bone mineral density predict occurrence of osteoporotic fractures. BMJ 1996, 312, 1254–1259. [Google Scholar] [CrossRef] [PubMed]
- Lonzer, M.D.; Imrie, R.; Rogers, D.; Worley, D.; Licata, A.; Secic, M. Effects of heredity, age, weight, puberty, activity, and calcium intake on bone mineral density in children. Clin. Pediatr. (Phila) 1996, 35, 185–189. [Google Scholar] [CrossRef]
- Riggs, B.L.; O’Fallon, W.M.; Muhs, J.; O’Connor, M.K.; Kumar, R.; Melton, L.J. Long-term effects of calcium supplementation on serum parathyroid hormone level, bone turnover, and bone loss in elderly women. J. Bone Miner. Res. 1998, 13, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Department of Health. Nutrition and Bone Health: With Particular Reference to Calcium and Vitamin D. Report of the Subgroup on Bone Health, Working Group on the Nutritional Status of the Population of the Committee on Medical Aspects of Food and Nutrition Policy; The Stationery Office: London, England, 1998. [Google Scholar]
- Dai, Z.; Li, Y.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol enhances proliferation an osteoblastic differentiation in human mesenchymal stem cells via ER-dependent ERK1/2 activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Baolin, L.; Inami, Y.; Tanaka, H.; Inagaki, N.; Iinuma, M.; Nagai, H. Resveratrol inhibits the release of mediators from bone marrow-derived mouse mast cells in vitro. Planta Med. 2004, 70, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dudley, J.I.; Das, D.K. Dose-dependency of resveratrol in providing health benefits. Dose Response 2010, 8, 478–500. [Google Scholar] [CrossRef] [PubMed]
- Cecchinato, V.; Chiaramonte, R.; Nizzardo, M.; Cristofaro, B.; Basile, A.; Sherbet, G.V.; Comi, P. Resveratrol-induced apoptosis in human T-cell acute lymphoblastic leukaemia MOLT-4 cells. Biochem. Pharmacol. 2007, 74, 1568–1574. [Google Scholar] [CrossRef] [PubMed]
- Boissy, P.; Andersen, T.L.; Abdallah, B.M.; Kassem, M.; Plesner, T.; Delaissé, J.M. Resveratrol Inhibits myeloma cell growth, prevents osteoclast formation, and promotes osteoblast differentiation. Cancer Res. 2005, 65, 9943–9952. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, K.; Ikeda, K.; Kawai, Y.; Yamori, Y. Resveratrol stimulates the proliferation and differentiation of osteoblastic MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 1998, 253, 859–863. [Google Scholar] [PubMed]
- Ulsperger, E.; Hamilton, G.; Raderer, M.; Baumgartner, G.; Hejna, M.; Hoffmann, O.; Mallinger, R. Resveratrol pretreatment desensitizes AHTO-7 human osteoblasts to growth stimulation in response to carcinoma cell supernatants. Int. J. Oncol. 1999, 15, 955–959. [Google Scholar] [PubMed]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado De Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-γ. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Lomb, D.J.; Laurent, G.; Haigis, M.C. Sirtuins regulate key aspects of lipid metabolism. Biochim. Biophys. Acta (BBA)—Proteins Proteomics 2010, 1804, 1652–1657. [Google Scholar] [CrossRef]
- Rayalam, S.; Yang, J.Y.; Ambati, S.; della-Fera, M.A.; Baile, C.A. Resveratrol induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Phytother. Res. 2008, 22, 1367–1371. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.; Rayalam, S.; della-Fera, M.A.; Baile, C.A. Phytochemicals and adipogenesis. BioFactors 2010, 36, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Tseng, P.-C.; Hou, S.M.; Chen, R.J.; Peng, H.W.; Hsieh, C.F.; Kuo, M.L.; Yen, M.L. Resveratrol promotes osteogenesis of human mesenchymal stem cells by up-regulating RUNX2 gene expression via SIRT1/FOXO3A axis. J. Bone Miner. Res. 2011, 26, 2552–2563. [Google Scholar] [CrossRef] [PubMed]
- Bäckesjö, C.-M.; Li, Y.; Lindgren, U.; Haldosén, L.A. Activation of Sirt1 decreases adipocyte formation during osteoblast differentiation of mesenchymal stem cells. J. Bone Miner. Res. 2006, 21, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Momken, I.; Stevens, L.; Bergouignan, A.; Desplanches, D.; Rudwill, F.; Chery, I.; Zahariev, A.; Zahn, S.; Stein, T.P.; Sebedio, J.L.; et al. Resveratrol prevents the wasting disorders of mechanical unloading by acting as a physical exercise mimetic in the rat. FASEB J. 2011, 25, 3646–3660. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Peshkin, L.; Price, N.L.; Labinskyy, N.; Swindell, W.R.; Kamara, D.; Minor, R.K.; Perez, E.; et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.P.; Li, W.X.; Yu, B.; Huang, J.; Sun, J.; Huo, J.S.; Liu, C.X. Effects of trans-resveratrol from Polygonum cuspidatum on bone loss using the ovariectomized rat model. J. Med. Food 2005, 8, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Habauzit, V.; Horcajada, M.-N. Phenolic phytochemicals and bone. Phytochem. Rev. 2008, 7, 313–344. [Google Scholar] [CrossRef]
- Habold, C.; Momken, I.; Ouadi, A.; Bekaert, V.; Brasse, D. Effect of prior treatment with resveratrol on density and structure of rat long bones under tail-suspension. J. Bone Miner. Metab. 2011, 29, 15–22. [Google Scholar] [CrossRef]
- Juhasz, B.; Mukherjee, S.; Das, D.K. Hormetic response of resveratrol against cardioprotection. Exp. Clin. Cardiol. 2010, 15, 134–138. [Google Scholar]
- Delucchi, F.; Berni, R.; Frati, C.; Cavalli, S.; Graiani, G.; Sala, R.; Chaponnier, C.; Gabbiani, G.; Calani, L.; del Rio, D.; et al. Resveratrol treatment reduces cardiac progenitor cell dysfunction and prevents morpho-functional ventricular remodeling in type-1 diabetic rats. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Juan, M.E.; Vinardell, M.P.; Planas, J.M. The daily oral administration of high doses of trans-resveratrol to rats for 28 days is not harmful. J. Nutr. 2002, 132, 257–260. [Google Scholar] [PubMed]
- Fan, C.; Cool, J.C.; Scherer, M.A.; Foster, B.K.; Shandala, T.; Tapp, H.; Xian, C.J. Damaging effects of chronic low-dose methotrexate usage on primary bone formation in young rats and potential protective effects of folinic acid supplementary treatment. Bone 2009, 44, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Xian, C.J.; Cool, J.C.; Pyragius, T.; Foster, B.K. Damage and recovery of the bone growth mechanism in young rats following 5-fluorouracil acute chemotherapy. J. Cell Biochem. 2006, 99, 1688–1704. [Google Scholar] [CrossRef] [PubMed]
- Eisman, J.A.; Kelly, P.J.; Morrison, N.A.; Pocock, N.A.; Yeoman, R.; Birmingham, J.; Sambrook, P.N. Peak bone mass and osteoporosis prevention. Osteoporos. Int. 1993, 3 (Suppl. 1), 56–60. [Google Scholar]
- Jilka, R.L. The relevance of mouse models for investigating age-related bone loss in humans. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Shayan, P.; Busch, F.; Aldinger, C.; Buhrmann, C.; Lueders, C.; Mobasheri, A. Resveratrol mediated modulation of Sirt-1/Runx2 promotes osteogenic differentiation of mesenchymal stem cells: Potential role of Runx2 deacetylation. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- De Wilde, A.; Maria Rassi, C.; Cournot, G.; Colin, C.; Lacroix, H.C.; Chaumaz, G.; Coxam, V.; Bennetau-Pelissero, C.; Pointillart, A.; Lieberherr, M. Dietary isoflavones act on bone marrow osteoprogenitor cells and stimulate ovary development before influencing bone mass in pre-pubertal piglets. J. Cell. Physiol. 2007, 212, 51–59. [Google Scholar]
- Turner, R.T.; Evans, G.L.; Zhang, M.; Maran, A.; Sibonga, J.D. Is resveratrol an estrogen agonist in growing rats? Endocrinology 1999, 140, 50–54. [Google Scholar] [PubMed]
- Draper, C.R.; Edel, M.J.; Dick, I.M.; Randall, A.G.; Martin, G.B.; Prince, R.L. Phytoestrogens reduce bone loss and bone resorption in oophorectomized rats. J. Nutr. 1997, 127, 1795–1799. [Google Scholar] [PubMed]
- Picherit, C.; Chanteranne, B.; Bennetau-Pelissero, C.; Davicco, M.J.; Lebecque, P.; Barlet, J.P.; Coxam, V. Dose-dependent bone-sparing effects of dietary isoflavones in the ovariectomised rat. Br. J. Nutr. 2001, 85, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Ernst, E. Alternative and complementary therapies for the menopause. Maturitas 2010, 66, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Xian, C.J.; Cool, J.C.; Scherer, M.A.; Fan, C.; Foster, B.K. Folinic acid attenuates methotrexate chemotherapy-induced damages on bone growth mechanisms and pools of bone marrow stromal cells. J. Cell. Physiol. 2008, 214, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Tresguerres, I.F.; Tamimi, F.; Eimar, H.; Barralet, J.; Torres, J.; Blanco, L.; Tresguerres, J.A.F. Resveratrol as anti-aging therapy for age-related bone loss. Rejuvenation Res. 2014, 17, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, A.; Bertelli, A.A.; Gozzini, A.; Giovannini, L. Plasma and tissue resveratrol concentrations and pharmacological activity. Drugs Under Exp. Clin. Res. 1998, 24, 133–138. [Google Scholar]
- El-Mohsen, M.; Bayele, H.; Kuhnle, G.; Gibson, G.; Debnam, E.; Kaila Srai, S.; Rice-Evans, C.; Spencer, J.P. Distribution of [3H]trans-resveratrol in rat tissues following oral administration. Br. J. Nutr. 2006, 96. [Google Scholar] [CrossRef]
- Wenzel, E.; Somoza, V. Metabolism and bioavailability of trans-resveratrol. Mol. Nutr. Food Res. 2005, 49, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Durbin, S.M.; Jackson, J.R.; Ryan, M.J.; Gigliotti, J.C.; Always, S.E.; Tou, J.C. Resveratrol supplementation preserves long bone mass, microstructure, and strength in hindlimb-suspended old male rats. J. Bone Miner. Metab. 2014, 32, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Sehmisch, S.; Hammer, F.; Christoffel, J.; Seidlova-Wuttke, D.; Tezval, M.; Wuttke, W.; Stuermer, K.M.; Stuermer, E.K. Comparison of the phytohormones genistein, resveratrol and 8-prenylnaringenin as agents for preventing osteoporosis. Planta Med. 2008, 74, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, A.A.; Giovannini, L.; Stradi, R.; Bertelli, A.; Tillement, J.P. Plasma, urine and tissue levels of trans- and cis-resveratrol (3,4′,5-trihydroxystilbene) after short-term or prolonged administration of red wine to rats. Int. J. Tissue React. 1996, 18, 67–71. [Google Scholar] [PubMed]
- Yu, C.; Shin, Y.G.; Chow, A.; Li, Y.; Kosmeder, J.W.; Lee, Y.S.; Hirschelman, W.H.; Pezzuto, J.M.; Mehta, R.G.; van Breemen, R.B. Human, rat, and mouse metabolism of resveratrol. Pharm. Res. 2002, 19, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Juan, M.E.; Lamuela-Raventós, R.M.; de la Torre-Boronat, M.C.; Planas, J.M. Determination of trans-resveratrol in plasma by HPLC. Anal. Chem. 1999, 71, 747–750. [Google Scholar] [CrossRef] [PubMed]
- Marier, J.-F.; Vachon, P.; Gritsas, A.; Zhang, J.; Moreau, J.P.; Ducharme, M.P. Metabolism and disposition of resveratrol in rats: Extent of absorption, glucuronidation, and enterohepatic recirculation eidenced by a linked-rat model. J. Pharmacol. Exp. Ther. 2002, 302, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xu, Y.; Liu, W. Tissue distribution and excretion of resveratrol in rat after oral administration of Polygonum cuspidatum extract (PCE). Phytomedicine 2008, 15, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.; Steward, W.P.; Gescher, A.J.; Brown, K. Resveratrol in human cancer chemoprevention—Choosing the “right” dose. Mol. Nutr. Food Res. 2012, 56, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical trials of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 161–169. [Google Scholar]
- Rotches-Ribalta, M.; Andres-Lacueva, C.; Estruch, R.; Escribano, E.; Urpi-Sarda, M. Pharmacokinetics of resveratrol metabolic profile in healthy humans after moderate consumption of red wine and grape extract tablets. Pharmacol. Res. 2012, 66, 375–382. [Google Scholar] [CrossRef]
- Beresford, J.N.; Bennett, J.H.; Devlin, C.; Leboy, P.S.; Owen, M.E. Evidence for an inverse relationship between the differentiation of adipocytic and osteogenic cells in rat marrow stromal cell cultures. J. Cell Sci. 1992, 102, 341–351. [Google Scholar] [PubMed]
- Pang, W.J.; Sun, S.D.; Bai, L.; Yang, Y.J.; Yang, G.S. Effects of resveratrol on pig primary preadipocytes proliferation, differentiation and transcription expression of Sirt1 gene. Sheng Wu Gong Cheng Xue Bao 2006, 22, 850–855. (In Chinese) [Google Scholar]
- Fischer-Posovszky, P.; Kukulus, V.; Tews, D.; Unterkircher, T.; Debatin, K.M.; Fulda, S.; Wabitsch, M. Resveratrol regulates human adipocyte number and function in a Sirt1-dependent manner. Am. J. Clin. Nutr. 2010, 92, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.-H.; Jang, H.J.; Chae, H.S.; Oh, Y.C.; Choi, J.G.; Lee, Y.S.; Kim, J.H.; Kim, Y.; Kim, Y.C.; Sohn, D.H.; Park, H.; Kwon, D.Y. Anti-inflammatory mechanisms of resveratrol in activated HMC-1 cells: Pivotal roles of NF-[kappa]B and MAPK. Pharmacol. Res. 2009, 59, 330–337. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, A.M.C.; Shandala, T.; Nguyen, L.; Muhlhausler, B.S.; Chen, K.-M.; Howe, P.R.; Xian, C.J. Effects of Resveratrol Supplementation on Bone Growth in Young Rats and Microarchitecture and Remodeling in Ageing Rats. Nutrients 2014, 6, 5871-5887. https://doi.org/10.3390/nu6125871
Lee AMC, Shandala T, Nguyen L, Muhlhausler BS, Chen K-M, Howe PR, Xian CJ. Effects of Resveratrol Supplementation on Bone Growth in Young Rats and Microarchitecture and Remodeling in Ageing Rats. Nutrients. 2014; 6(12):5871-5887. https://doi.org/10.3390/nu6125871
Chicago/Turabian StyleLee, Alice M. C., Tetyana Shandala, Long Nguyen, Beverly S. Muhlhausler, Ke-Ming Chen, Peter R. Howe, and Cory J. Xian. 2014. "Effects of Resveratrol Supplementation on Bone Growth in Young Rats and Microarchitecture and Remodeling in Ageing Rats" Nutrients 6, no. 12: 5871-5887. https://doi.org/10.3390/nu6125871