l-Leucine Supplementation Worsens the Adiposity of Already Obese Rats by Promoting a Hypothalamic Pattern of Gene Expression that Favors Fat Accumulation


Abstract
:1. Introduction
2. Experimental Section
2.1. Animals
2.2. Experimental Design
2.3. Relative Gene Expression (qPCR)
2.4. Statistical Analysis
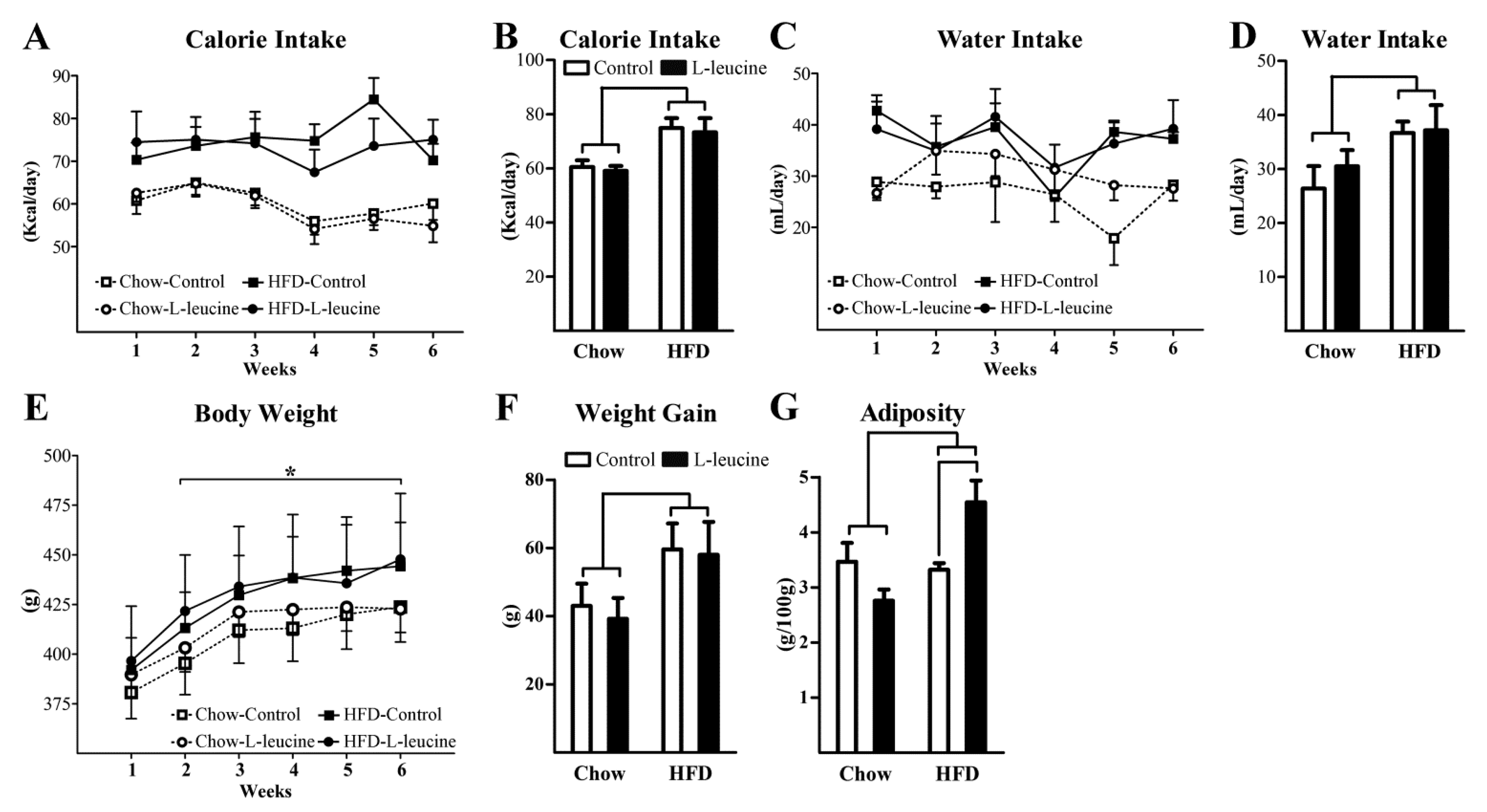
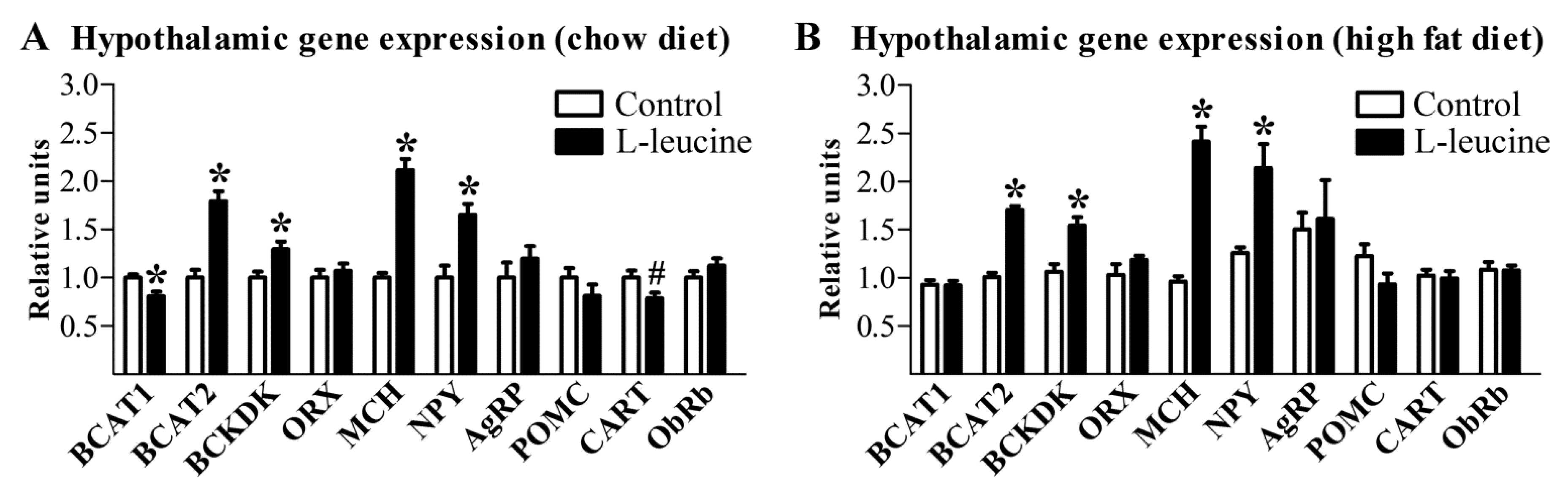
3. Results

{kind=link}
{kind=link}
| Gene of Interest | Accession No. | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Product Size (bp) |
|---|---|---|---|---|
| β-2M | Rn00560865_m1 | * | * | * |
| β-Actin | NM_031144.3 | AGCCTGGATGGCTACGTACA | CCTCTGAACCCTAAGGCCAA | 90 |
| GAPDH | NM_017008.4 | CCGTTCAGGTCTGGGATGAC | GGGCAGCCCAGAACATCAT | 76 |
| HPRT | NM_012583.2 | TTTGCTGACCTGCTGGATTAC | ACTTTTATGTCCCCCGTTGA | 125 |
| TFRC | RN01474695_m1 | * | * | * |
| RPL-37A | RN02114291_s1 | * | * | * |
| BCAT1 | NM_017253.2 | CGGCCGGACCTCAACATGAAAAGA | CCTTGTCAAACTCCGGCAGAGTGG | 70 |
| BCAT2 | NM_022400.1 | TCCCTGGCGTCTATGTGTGCCC | TGGAGGTCTGCCGCCTTGAAGT | 63 |
| BCKDK | NM_019244.2 | AGAGCTGGCCAGGGAACGCT | TGAGGCGGACTGAGGGCTTCT | 92 |
| ORX | NM_013179.2 | GCGGCCTCAGACTCCT | AGGGAGAGGCAATCCGGAGAG | 70 |
| MCH | M29712.1 | ATGCTGGCCTTTTCTTTGTTT | CTTCTACGTTCCTGATGGACTT | 70 |
| NPY | NM_012614.2 | CCGCCCGCCATGATGCTAGGTA | CCCTCAGCCAGAATGCCCAA | 88 |
| AgRP | NM_033650.1 | GCAGAGGTGCTAGATCCACAGAA | AGGACTCGTGCAGCCTTACAC | 70 |
| POMC | NM_139326.2 | ATAGACGTGTGGAGCTGGTGC | GCAAGCCAGCAGGTTGCT | 75 |
| CART | NM_017110.1 | CCGAGCCCTGGACATCTACT | CCGCCTTGGCAGCTCCTT | 64 |
| ObRb | NM_012596.1 | ACTTAATTTCCAAAAGCCTGAAACA | CCAGAAGAAGAGGACCAAATATCAC | 83 |

4. Discussion

5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Zhang, Y.; Guo, K.; LeBlanc, R.E.; Loh, D.; Schwartz, G.J.; Yu, Y.H. Increasing dietary leucine intake reduces diet-induced obesity and improves glucose and cholesterol metabolism in mice via multimechanisms. Diabetes 2007, 56, 1647–1654. [Google Scholar] [CrossRef]
- Freudenberg, A.; Petzke, K.J.; Klaus, S. Comparison of high-protein diets and leucine supplementation in the prevention of metabolic syndrome and related disorders in mice. J. Nutr. Biochem. 2012, 23, 1524–1530. [Google Scholar] [CrossRef]
- Macotela, Y.; Emanuelli, B.; Bång, A.M.; Espinoza, D.O.; Boucher, J.; Beebe, K.; Gall, W.; Kahn, C.R. Dietary leucine—An environmental modifier of insulin resistance acting on multiple levels of metabolism. PLoS One 2011, 6, e21187. [Google Scholar]
- Binder, E.; Bermudez-Silva, F.J.; Andre, C.; Elie, M.; Romero-Zerbo, S.Y.; Leste-Lasserre, T.; Belluomo, L.; Duchampt, A.; Clark, S.; Aubert, A.; et al. Leucine supplementation protects from insulin resistance by regulating adiposity levels. PLoS One 2013, 8, e74705. [Google Scholar] [CrossRef]
- Li, H.; Xu, M.; Lee, J.; He, C.; Xie, Z. Leucine supplementation increases SIRT1 expression and prevents mitochondrial dysfunction and metabolic disorders in high fat diet-induced obese mice. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1234–E1244. [Google Scholar] [CrossRef]
- Layman, D.K.; Walker, D.A. Potential importance of leucine in treatment of obesity and the metabolic syndrome. J. Nutr. 2006, 136, 319S–323S. [Google Scholar]
- Donato, J., Jr.; Pedrosa, R.G.; de Araujo, J.A., Jr.; Pires, I.S.; Tirapegui, J. Effects of leucine and phenylalanine supplementation during intermittent periods of food restriction and refeeding in adult rats. Life Sci. 2007, 81, 31–39. [Google Scholar] [CrossRef]
- Donato, J., Jr.; Pedrosa, R.G.; Cruzat, V.F.; Pires, I.S.; Tirapegui, J. Effects of leucine supplementation on the body composition and protein status of rats submitted to food restriction. Nutrition 2006, 22, 520–527. [Google Scholar] [CrossRef]
- Torres-Leal, F.; Fonseca-Alaniz, M.; Teodoro, G.; de Capitani, M.; Vianna, D.; Pantaleao, L.; Matos-Neto, E.; Rogero, M.; Donato, J.; Tirapegui, J. Leucine supplementation improves adiponectin and total cholesterol concentrations despite the lack of changes in adiposity or glucose homeostasis in rats previously exposed to a high-fat diet. Nutr. Metab. 2011, 8, 1–62. [Google Scholar] [CrossRef]
- Zampieri, T.T.; Pedroso, J.A.B.; Furigo, I.C.; Tirapegui, J.; Donato, J., Jr. Oral leucine supplementation is sensed by the brain but neither reduces food intake nor induces an anorectic pattern of gene expression in the hypothalamus. PLoS One 2013, 8, e84094. [Google Scholar]
- Lynch, C.J.; Hutson, S.M.; Patson, B.J.; Vaval, A.; Vary, T.C. Tissue-specific effects of chronic dietary leucine and norleucine supplementation on protein synthesis in rats. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E824–E835. [Google Scholar]
- Reiter, A.K.; Anthony, T.G.; Anthony, J.C.; Jefferson, L.S.; Kimball, S.R. The mtor signaling pathway mediates control of ribosomal protein mrna translation in rat liver. Int. J. Biochem. Cell Biol. 2004, 36, 2169–2179. [Google Scholar] [CrossRef]
- Cota, D.; Proulx, K.; Smith, K.A.; Kozma, S.C.; Thomas, G.; Woods, S.C.; Seeley, R.J. Hypothalamic mtor signaling regulates food intake. Science 2006, 312, 927–930. [Google Scholar] [CrossRef]
- Sweatt, A.J.; Wood, M.; Suryawan, A.; Wallin, R.; Willingham, M.C.; Hutson, S.M. Branched-chain amino acid catabolism: Unique segregation of pathway enzymes in organ systems and peripheral nerves. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E64–E76. [Google Scholar]
- Guo, K.; Yu, Y.H.; Hou, J.; Zhang, Y. Chronic leucine supplementation improves glycemic control in etiologically distinct mouse models of obesity and diabetes mellitus. Nutr. Metab. 2010, 7, 1–57. [Google Scholar] [CrossRef]
- Ropelle, E.R.; Pauli, J.R.; Fernandes, M.F.A.; Rocco, S.A.; Marin, R.M.; Morari, J.; Souza, K.K.; Dias, M.M.; Gomes-Marcondes, M.C.; Gontijo, J.A.R. A central role for neuronal AMP-activated protein kinase (AMPK) and mammalian target of rapamycin (mTOR) in high-protein diet-induced weight loss. Diabetes 2008, 57, 594–605. [Google Scholar] [CrossRef]
- Koch, C.; Göddeke, S.; Krüger, M.; Tups, A. Effect of central and peripheral leucine on energy metabolism in the djungarian hamster (Phodopus sungorus). J. Comp. Physiol. 2013, 183, 261–268. [Google Scholar]
- Eller, L.K.; Saha, D.C.; Shearer, J.; Reimer, R.A. Dietary leucine improves whole-body insulin sensitivity independent of body fat in diet-induced obese sprague-dawley rats. J. Nutr. Biochem. 2013, 24, 1285–1294. [Google Scholar] [CrossRef]
- Zeanandin, G.; Balage, M.; Schneider, S.; Dupont, J.; Hébuterne, X.; Mothe-Satney, I.; Dardevet, D. Differential effect of long-term leucine supplementation on skeletal muscle and adipose tissue in old rats: An insulin signaling pathway approach. Age 2012, 34, 371–387. [Google Scholar] [CrossRef]
- Lynch, C.J.; Patson, B.J.; Anthony, J.; Vaval, A.; Jefferson, L.S.; Vary, T.C. Leucine is a direct-acting nutrient signal that regulates protein synthesis in adipose tissue. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E503–E513. [Google Scholar]
- Polak, P.; Cybulski, N.; Feige, J.N.; Auwerx, J.; Rüegg, M.A.; Hall, M.N. Adipose-specific knockout of raptor results in lean mice with enhanced mitochondrial respiration. Cell Metab. 2008, 8, 399–410. [Google Scholar] [CrossRef]
- Chakrabarti, P.; English, T.; Shi, J.; Smas, C.M.; Kandror, K.V. Mammalian target of rapamycin complex 1 suppresses lipolysis, stimulates lipogenesis, and promotes fat storage. Diabetes 2010, 59, 775–781. [Google Scholar] [CrossRef]
- Zemel, M.B.; Bruckbauer, A. Effects of a leucine and pyridoxine-containing nutraceutical on fat oxidation, and oxidative and inflammatory stress in overweight and obese subjects. Nutrients 2012, 4, 529–541. [Google Scholar] [CrossRef]
- Bruckbauer, A.; Zemel, M. Effects of dairy consumption on SIRT1 and mitochondrial biogenesis in adipocytes and muscle cells. Nutr. Metab. 2011, 8, 1–91. [Google Scholar] [CrossRef]
- Bruckbauer, A.; Zemel, M.; Thorpe, T.; Akula, M.; Stuckey, A.; Osborne, D.; Martin, E.; Kennel, S.; Wall, J. Synergistic effects of leucine and resveratrol on insulin sensitivity and fat metabolism in adipocytes and mice. Nutr. Metab. 2012, 9, 1–77. [Google Scholar] [CrossRef] [Green Version]
- Bruckbauer, A.; Zemel, M.B. Synergistic effects of polyphenols and methylxanthines with leucine on AMPK/Sirtuin-mediated metabolism in muscle cells and adipocytes. PLoS One 2014, 9, e89166. [Google Scholar] [CrossRef]
- Paxton, R.; Harris, R.A. Regulation of branched-chain α-ketoacid dehydrogenase kinase. Arch. Biochem. Biophys. 1984, 231, 48–57. [Google Scholar] [CrossRef]
- Harris, R.A.; Joshi, M.; Jeoung, N.H. Mechanisms responsible for regulation of branched-chain amino acid catabolism. Biochem. Biophys. Res. Commun. 2004, 313, 391–396. [Google Scholar] [CrossRef]
- Myers, M.G., Jr.; Olson, D.P. Central nervous system control of metabolism. Nature 2012, 491, 357–363. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zampieri, T.T.; Torres-Leal, F.L.; Campaña, A.B.; Lima, F.B.; Donato, J., Jr. l-Leucine Supplementation Worsens the Adiposity of Already Obese Rats by Promoting a Hypothalamic Pattern of Gene Expression that Favors Fat Accumulation. Nutrients 2014, 6, 1364-1373. https://doi.org/10.3390/nu6041364
Zampieri TT, Torres-Leal FL, Campaña AB, Lima FB, Donato J Jr. l-Leucine Supplementation Worsens the Adiposity of Already Obese Rats by Promoting a Hypothalamic Pattern of Gene Expression that Favors Fat Accumulation. Nutrients. 2014; 6(4):1364-1373. https://doi.org/10.3390/nu6041364
Chicago/Turabian StyleZampieri, Thais T., Francisco L. Torres-Leal, Amanda B. Campaña, Fabio B. Lima, and Jose Donato, Jr. 2014. "l-Leucine Supplementation Worsens the Adiposity of Already Obese Rats by Promoting a Hypothalamic Pattern of Gene Expression that Favors Fat Accumulation" Nutrients 6, no. 4: 1364-1373. https://doi.org/10.3390/nu6041364