Hypolipidemic and Antioxidative Effects of Aqueous Enzymatic Extract from Rice Bran in Rats Fed a High-Fat and -Cholesterol Diet

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Composition of AEERB
2.3. Antioxidant Potential of AEERB in Vitro
2.4. Animals and Treatment

{kind=link}
{kind=link}
{kind=link}
| Normal Diet | % | Hyperlipidemic Diet | % |
|---|---|---|---|
| Protein | 20 | Normal diet | 78.8 |
| Fat | 4.5 | Lard oil | 10 |
| Carbohydrates | 52 | Yolk powder | 10 |
| Fiber | 4.3 | Cholesterol | 1.0 |
| Moisture | 9.1 | Bile salt | 0.2 |
| Ash | 8.0 | ||
| Calcium | 1.3 | ||
| Phosphorus | 0.8 |
2.5. Antioxidant Activity in Vivo
2.6. Metabolic Parameters Analysis
2.7. Hepatic HMG-CoA Reductase Activity Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characterization of AEERB
| Composition | AEERB | RB |
|---|---|---|
| Protein (%) | 26.10 ± 0.56 a | 13.43 ± 0.54 b |
| Crude fat (%) | 10.56 ± 0.3 b | 18.84 ± 0.09 a |
| Reducing sugar (%) | 8.43 ± 0.3 a | 1.47 ± 0.2 b |
| TPC (mg GAE/g) | 16.87 ± 0.006 a | 2.9 ± 0.1 b |
| TFC (mg RE/g) | 4.15 ± 0.07 a | 0.79 ± 0.06 b |
| γ-Oryzanol (mg/g) | 3.49 ± 0.1 a | 1.27 ± 0.08 b |
| Vitamin E (mg/kg) | ||
| α-Tocopherol | 5.81 ± 0.28 a | 5.40 ± 0.71 a |
| β-Tocopherol | 0.85 ± 0.17 a | 0.40 ± 0.5 b |
| γ-Tocopherol | 1.19 ± 0.20 a | 1.30 ± 0.21 a |
| α-Tocotrienol | 4.42 ± 0.31 a | 1.7 ± 0.24 b |
| γ-Tocotrienol | 10.80 ± 0.42 b | 14.2 ± 0.40 a |
| δ-Tocotrienol | 1.30 ± 0.20 a | 1.20 ± 0.36 a |
3.2. Antioxidant Potential in Vitro
| Sample | Ferric Reducing Ability (mmol FeSO4·g−1) | DPPH• Scavenging Activity (IC50) (μg·mL−1) | ABTS•+ Scavenging Activity (IC50) (μg·mL−1) | LPI (%) |
|---|---|---|---|---|
| AEERB | 3.15 ± 0.1 c | 98.87 ± 18.3 a | 103.83 ± 3.0 a | 22.72 ± 0.7 c |
| AA | 120.83 ± 0.6 a | 23.58 ± 5.5 b | 17.95 ± 0.6 b | 27.11 ± 0.3 b |
| BHT | 82 ± 0.9 b | 91.61 ± 6.3 a | 6.19 ± 0.5 c | 48.93 ± 0.8 a |
3.3. Effect on Body and Organ Weights
| Growth Parameters | ND | HD | AEERB |
|---|---|---|---|
| Body weight gain (g/rat/day) | 2.01 ± 0.88 b | 3.54 ± 1.15 a | 2.04 ± 1.11 b |
| Food intake (g/day/rat) | 24.02 ± 2.3 a | 18.19 ± 2.0 c | 19.75 ± 2.5 b |
| Liver (g/100 g body weight) | 3.22 ± 0.76 b | 3.89 ± 0.36 a | 3.54 ± 0.37 ab |
| Epididymal adipose (g) | 4.46 ± 1.33 b | 6.83 ± 2.53 a | 4.26 ± 0.95 b |
| Perirenal adipose tissue (g) | 0.57 ± 0.37 a | 1.00 ± 0.51 a | 0.76 ± 0.27 a |
| Mesenteric (g) | 2.06 ± 0.46 b | 3.05 ± 0.70 a | 2.36 ± 0.91 ab |
3.4. Antioxidant Activity in Vivo
| Group | TAOC (U/mL) | SOD (U/mL) | GSH-Px (U) | CAT (U/mL) | MDA (nmol/mL) | Protein Carbonyl (nmol/mg protein) |
|---|---|---|---|---|---|---|
| ND | 15.42 ± 1.62 a | 293.05 ± 20.09 a | 719.02 ± 87.75 a | 11.85 ± 2.49 a | 6.08 ± 1.87 a | 0.0518 ± 0.013 b |
| HD | 10.34 ± 1.75 b | 87.85 ± 22.36 c | 600.98 ± 173.00 a | 8.72 ± 3.26 a | 8.00 ± 1.44 a | 0.0787 ± 0.016 a |
| AEERB | 14.13 ± 0.85 a | 161.63 ± 26.57 b | 688.78 ± 124.28 a | 9.23 ± 3.44 a | 7.34 ± 1.87 a | 0.0596 ± 0.022 ab |
| Group | TAOC (U/mg protein) | SOD (U/mg protein) | GSH-Px (U) | CAT (U/mg protein) | MDA (nmol/mg protein) | Protein Carbonyl (nmol/mg protein) |
|---|---|---|---|---|---|---|
| Live tissue | ||||||
| ND | 1.83 ± 0.45 a | 109.17 ± 16.39 a | 117.34 ± 14.11 a | 45.21 ± 12.48 a | 1.03 ± 0.14 b | 1.78 ± 0.18 c |
| HD | 1.06 ± 0.25 b | 65.21 ± 10.60 b | 97.85 ± 17.23 b | 22.85 ± 6.77 b | 1.19 ± 0.12 a | 3.46 ± 0.50 a |
| AEERB | 1.40 ± 0.25 b | 97.63 ± 5.65 ab | 111.74 ± 11.01 ab | 41.81 ± 4.95 a | 1.08 ± 0.16 ab | 2.77 ± 0.77 b |
| Brain tissue | ||||||
| ND | 1.09 ± 0.24 a | 55.20 ± 23.80 a | 66.33 ± 17.06 a | 11.83 ± 3.17 a | 3.52 ± 0.90 b | 2.28 ± 0.89 b |
| HD | 0.91 ± 0.13 a | 46.45 ± 18.09 a | 50.75 ± 27.42 a | 5.32 ± 1.85 b | 6.56 ± 1.35 a | 3.67 ± 0.53 a |
| AEERB | 1.04 ± 0.25 a | 52.24 ± 19.68 a | 63.13 ± 12.47 a | 6.79 ± 1.68 b | 5.52 ± 1.19 a | 3.29 ± 1.06 a |
3.5. Serum Lipid Analysis
| Serum Lipid Parameters | ND | HD | AEERB |
|---|---|---|---|
| TC (mmol/L) | 1.52 ± 0.08 c | 2.65 ± 0.17 a | 2.00 ± 0.12 b |
| TG (mmol/L) | 0.50 ± 0.22 b | 0.85 ± 0.16 a | 0.63 ± 0.04 b |
| HDL-C (mmol/L) | 0.87 ± 0.06 a | 0.58 ± 0.06 c | 0.77 ± 0.09 b |
| LDL-C (mmol/L) | 0.51 ± 0.12 c | 0.98 ± 0.11 a | 0.73 ± 0.10 b |
| ApoA (g/L) | 0.039 ± 0.01 a | 0.022 ± 0.01 b | 0.027 ± 0.01 b |
| ApoB (g/L) | 0.026 ± 0.01 b | 0.039 ± 0.01 a | 0.033 ± 0.01 ab |
| Lp (a) (mg/L) | 1.37 ± 0.23 b | 1.88 ± 0.23 a | 1.54 ± 0.14 b |
| AI1 | 0.75 ± 0.15 c | 3.59 ± 0.48 a | 1.65 ± 0.43 b |
| AI2 | 0.58 ± 0.16 c | 1.71 ± 0.27 a | 0.97 ± 0.23 b |
3.6. Liver Lipid Analysis
| Group | Total Lipids (%) | TC (μmol/g) | Free Cholesterol (μmol/g) | Cholesterol Ester (μmol/g) | TG (μmol/g) |
|---|---|---|---|---|---|
| ND | 33.46 ± 11.51 b | 67.33 ± 7.46 b | 28.54 ± 1.63 b | 38.79 ± 7.90 b | 58.23 ± 6.04 b |
| HD | 55.46 ± 26.20 a | 94.21 ± 2.83 a | 37.60 ± 6.74 a | 56.61 ± 6.73 a | 71.15 ± 5.5 a |
| AEERB | 15.47 ± 6.61 b | 70.01 ± 8.49 b | 33.68 ± 2.69 a | 36.32 ± 9.76 b | 41.40 ± 8.38 c |
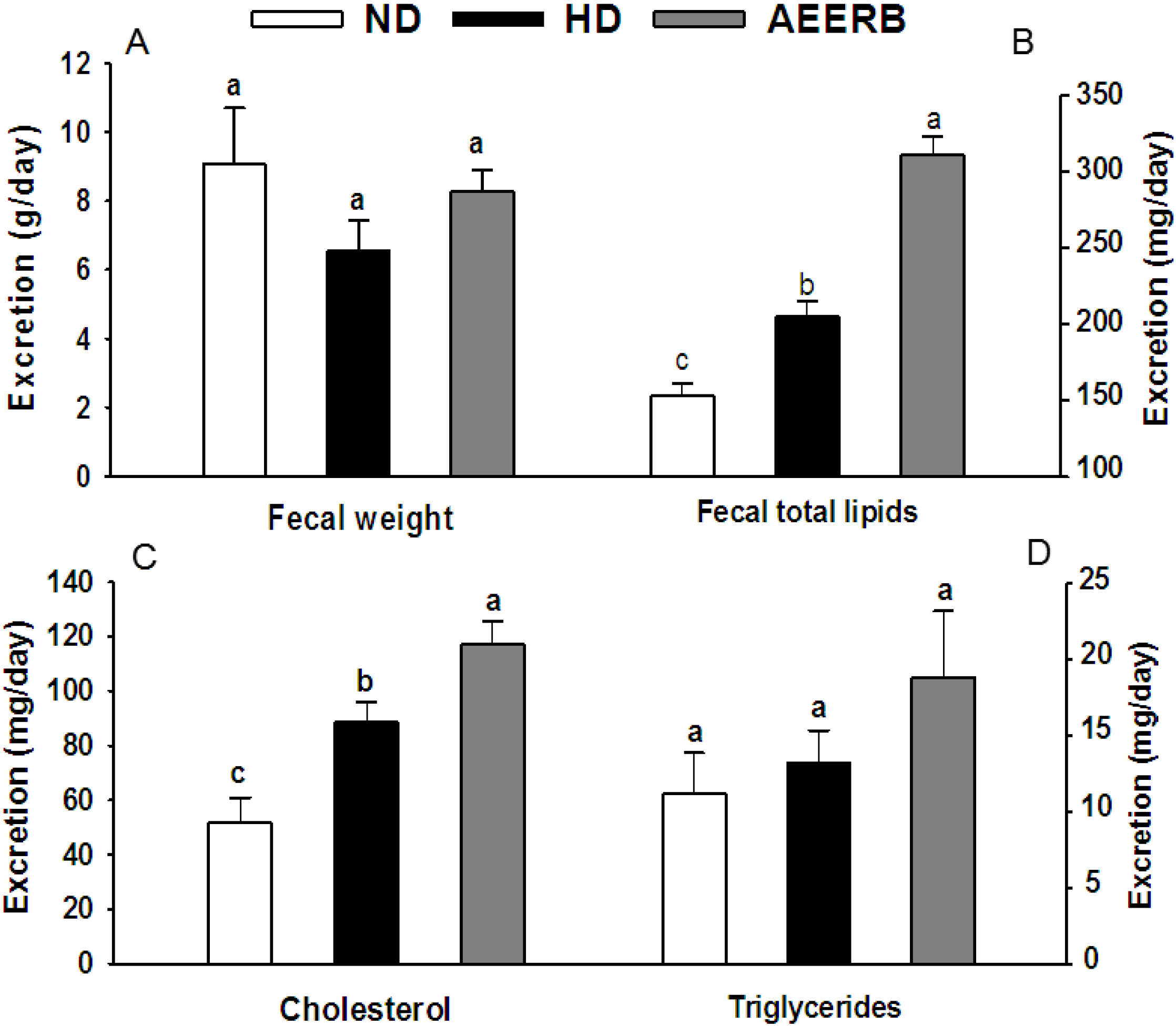
3.7. Fecal Lipid Analysis

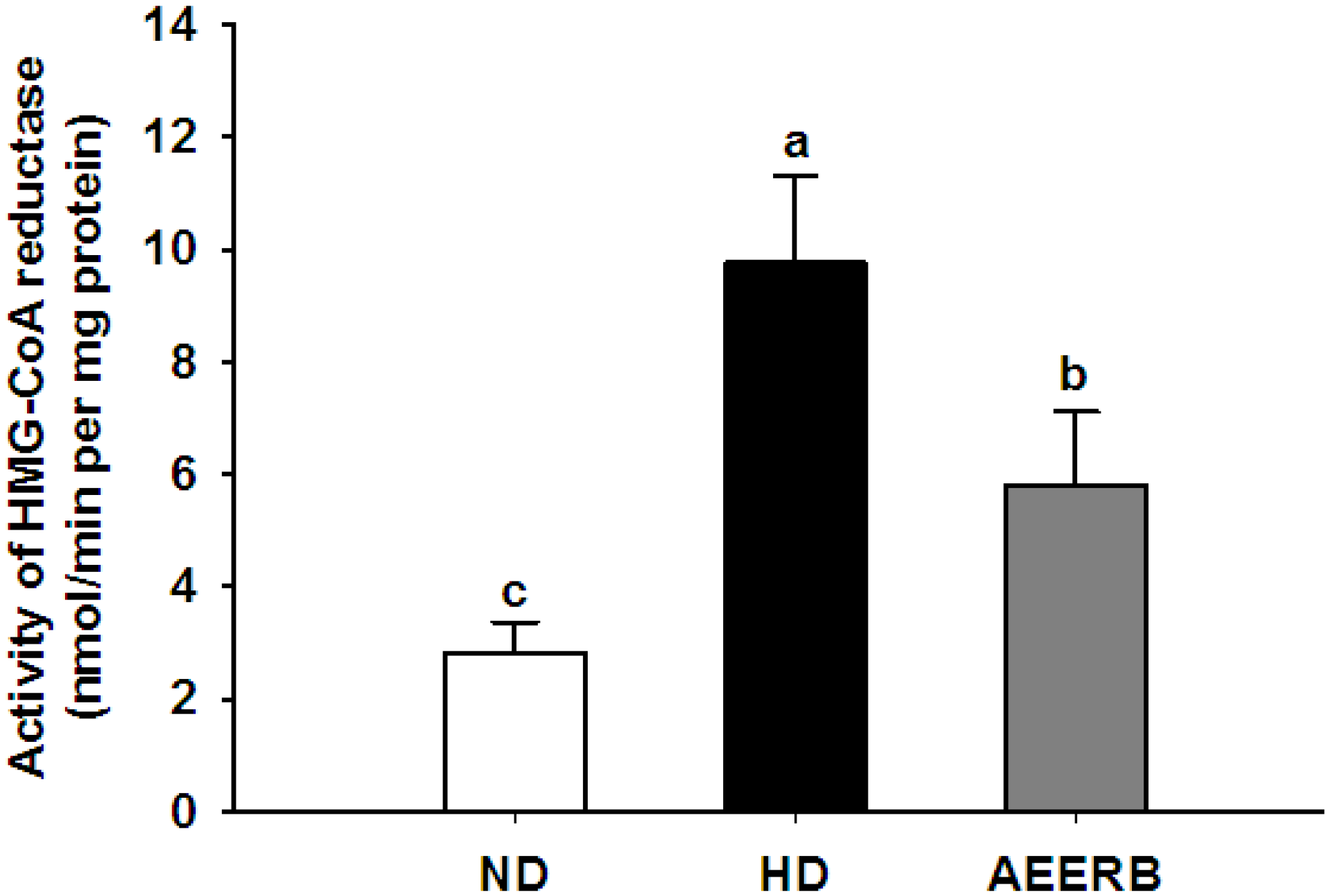
3.8. Hepatic HMG-CoA Reductase Activity

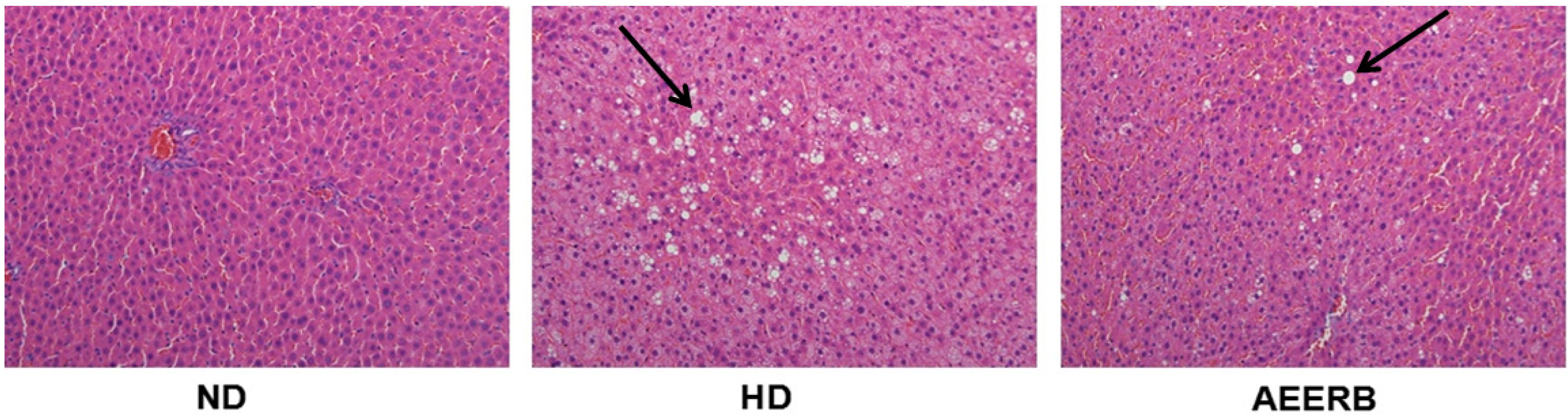
3.9. Pathological Examination

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Roberts, R.; Stewart, A.F.; Well, G.A.; Williams, K.A.; Kavaslar, N.; Mcpherson, R. Identifying genes for coronary artery disease: An idea whose time has come. Can. J. Cardiol. 2007, 23, 7A–15A. [Google Scholar] [CrossRef] [PubMed]
- Ausman, L.M.; Rong, N.; Nicolosi, R.J. Hypocholesterolemic effect of physically refined rice bran oil: Studies of cholesterol metabolism and early atherosclerosis in hypercholesterolemic hamsters. J. Nutr. Biochem. 2005, 16, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Revilla, E.; Maria, C.S.; Miramontes, E.; Bautista, J.; García-Martínez, A.; Cremades, O.; Parrado, J. Nutraceutical composition, antioxidant activity and hypocholesterolemic effect of a water-soluble enzymatic extract from rice bran. Food Res. Int. 2009, 42, 387–393. [Google Scholar] [CrossRef]
- Münzel, T.; Sinning, C.; Post, F.; Warnholtz, A.; Schulz, E. Pathophysiology, diagnosis and prognostic implications of endothelial dysfunction. Ann. Med. 2008, 40, 180–196. [Google Scholar]
- Ha, T.-Y.; Han, S.; Kim, S.-R.; Kim, I.-H.; Lee, H.-Y.; Kim, H.-K. Bioactive components in rice bran oil improve lipid profiles in rats fed a high-cholesterol diet. Nutr. Res. 2005, 25, 597–606. [Google Scholar] [CrossRef]
- Jariwalla, R.J. Rice-bran products: Phytonutrients with potential applications in preventive and clinical medicine. Drug. Exp. Clin. Res. 2001, 27, 17–26. [Google Scholar]
- Most, M.M.; Tulley, R.; Morales, S.; Lefevre, M. Rice bran oil, not fiber, lowers cholesterol in humans. Am. J. Clin. Nutr. 2005, 81, 64–68. [Google Scholar] [PubMed]
- Parrado, J.; Miramontes, E.; Jover, M.; Márquez, J.C.; Mejias, M.; Collantes de Terán, L.; Bautista, J. Prevention of brain protein and lipid oxidation elicited by a water soluble oryzanol enzymatic extract derived from rice bran. Eur. J. Nutr. 2003, 42, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Justo, M.L.; Rodriguez-Rodriguez, R.; Claro, C.M.; de Sotomayor, M.A.; Parrado, J.; Herrera, M.D. Water-soluble rice bran enzymatic extract attenuates dyslipidemia, hypertension and insulin resistance in obese Zucker rats. Eur. J. Nutr. 2013, 52, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Gaddi, A. Rice bran oil and gamma-oryzanol in the treatment of hyperlipoproteinaemias and other conditions. Phytother. Res. 2001, 15, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.A.; Nicolosi, R.J.; Woolfrey, B.; Kritchevsky, D. Rice bran oil and oryzanol reduce plasma lipid and lipoprotein cholesterol concentrations and aortic cholesterol ester accumulation to a greater extent than ferulic acid in hypercholesterolemic hamsters. J. Nutr. Biochem. 2007, 18, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Chopra, R.; Sambaiah, K. Effects of rice bran oil enriched with n-3 PUFA on liver and serum lipids in rats. Lipids 2009, 44, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Rein, D.; Schäfer, A.; Monnard, I.; Gremaud, G.; Lambelet, P.; Bertoli, C. Similar cholesterol-lowering properties of rice bran oil, with varied gamma-oryzanol, in mildly hypercholesterolemic men. Eur. J. Nutr. 2005, 44, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Davi, G.; Santilli, F.; Patrono, C. Nutraceuticals in diabetes and metabolic syndrome. Cardiovasc. Ther. 2010, 28, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Abete, I.; Goyenechea, E.; Zulet, M.A.; Martínez, J.A. Obesity and metabolic syndrome: Potential benefit from specific nutritional components. Nutr. Metab. Cardiovasc. 2011, 21, B1–B15. [Google Scholar] [CrossRef]
- Sun, A.M.; Yu, G.P.; Wang, Y.X.; Bian, J.; Fu, R.; Liu, M.J.; Xu, M.X. Aqueous enzymatic method for preparing the mixed extract with antioxidant activity from rice bran. Adv. Mater. Res. 2013, 781, 792–798. [Google Scholar] [CrossRef]
- Somogyi, M. Notes on sugar determination. J. Boil. Chem. 1952, 195, 19–23. [Google Scholar]
- Sungsopha, J.; Moongngarm, A.; Kanesakoo, R. Application of germination and enzymatic treatment to improve the concentration of bioactive compounds and antioxidant activity of rice bran. Austr. J. Basic Appl. Sci. 2009, 3, 3653–3661. [Google Scholar]
- Liu, J.; Wang, C.; Wang, Z.; Zhang, C.; Lu, S.; Liu, J. The antioxidant and free-radical scavenging activities of extract and fractions from corn silk (Zea mays L.) and related flavone glycosides. Food Chem. 2011, 126, 261–269. [Google Scholar]
- Park, J.W.; Jang, E.Y.; Kim, J.; Yi, B.; Kim, M.J.; Park, K.W.; Lee, J. Effects of visible light irradiation on the oxidative stability in rice bran. J. Cereal Sci. 2013, 58, 178–181. [Google Scholar] [CrossRef]
- Yu, L.; Haley, S.; Perret, J.; Harris, M.; Wilson, J.; Qian, M. Free radical scavenging properties of wheat extracts. J. Agric. Food Chem. 2002, 50, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Carlos, A.G.; Grace, G.; Mercedes, B.M.; Patricio, H.; Victor, C.G. Correlation of tocopherol, tocotrienol, γ-oryzanol and total polyphenol content in rice bran with different antioxidant capacity assays. Food Chem. 2007, 102, 1228–1232. [Google Scholar] [CrossRef]
- Kong, B.H.; Xiong, Y.L. Antioxidant activity of zein hydrolysates in a liposome system and the possible mode of action. J. Agric. Food Chem. 2006, 54, 6059–6068. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yao, H.Y. Antioxidant activities of barley seeds extracts. Food Chem. 2007, 102, 732–737. [Google Scholar] [CrossRef]
- He, L.; Zhang, X.; Xu, H.; Xu, C.; Yuan, F.; Knez, Ž.; Novak, Z.; Gao, Y.X. Subcritical water extraction of phenolic compounds from pomegranate (Punica granatum L.) seed residues and investigation into their antioxidant activities with HPLC-ABTS•+ assay. Food Bioprod. Process. 2012, 90, 215–223. [Google Scholar]
- Siddhuraju, P.; Manian, S. The antioxidant activity and free radical-scavenging capacity of dietary phenolic extracts from horse gram (Macrotyloma uniflorum (Lam.) Verdc.) seeds. Food Chem. 2007, 105, 950–958. [Google Scholar]
- Pyo, Y.H.; Seong, K.S. Hypolipidemic effects of Monascus-fermented soybean extracts in rats fed a high-fat and -cholesterol diet. J. Agric. Food Chem. 2009, 57, 8617–8622. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Tong, L.T.; Zhong, K.; Liu, L.; Guo, L.; Cao, L.; Zhou, S. Oat oil lowers the plasma and liver cholesterol concentrations by promoting the excretion of faecal lipids in hypercholesterolemic rats. Food Chem. 2014, 142, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Anila, L.; Vijayalakshmi, N.R. Antioxidant action of flavonoids from Mangifera indica and Emblica officinalis in hypercholesterolemic rats. Food Chem. 2003, 83, 569–574. [Google Scholar] [CrossRef]
- Zou, Y.P.; Lu, Y.H.; Wei, D.Z. Hypocholesterolemic effects of a flavonoid-rich extract of Hypericum perforatum L. in rats fed a cholesterol-rich diet. J. Agric. Food Chem. 2005, 53, 2462–2466. [Google Scholar]
- Gris, E.F.; Mattivi, F.; Ferreira, E.A.; Vrhovsek, U.; Filho, D.W.; Pedrosa, R.C.; Bordignon-Luiz, M.T. Stilbenes and tyrosol as target compounds in the assessment of antioxidant and hypolipidemic activity of Vitis vinifera red wines from southern Brazil. J. Agric. Food Chem. 2011, 59, 7954–7961. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.P.; Lu, Y.H.; Wei, D.Z. Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro. J. Agric. Food Chem. 2004, 52, 5032–5039. [Google Scholar] [CrossRef]
- Martinez, L.O.; Jacquet, S.; Terce, F.; Collet, X.; Perret, B.; Barbaras, R. New insight on the molecular mechanisms of highdensity lipoprotein cellular interaction. Cell. Mol. Life Sci. 2004, 61, 2343–2360. [Google Scholar] [CrossRef] [PubMed]
- Sugano, M.; Tsuji, E. Rice bran oil and cholesterol metabolism. J. Nutr. 1997, 127, 521S–524S. [Google Scholar] [PubMed]
- Temel, R.E.; Hou, L.; Rudel, L.L.; Shelness, G.S. ACAT2 stimulates cholesteryl ester secretion in apo B-containing lipoproteins. J. Lipid Res. 2007, 48, 1618–1627. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, J.H.; Xu, T.; Nie, M.H.; Yang, H.K. Hypocholesterolemic effect of rice protein is due to regulating hepatic cholesterol metabolism in adult rats. Gene 2013, 512, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Minhajuddin, M.; Beg, Z.H.; Iqbal, J. Hypolipidemic and antioxidant properties of tocotrienol rich fraction isolated from rice bran oil in experimentally induced hyperlipidemic rats. Food Chem. Toxicol. 2005, 43, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Minhajuddin, M.; Beg, Z.H. Suppression of 7,12-dimethylbenz[α]anthracene-induced carcinogenesis and hypercholesterolaemia in rats by tocotrienol-rich fraction isolated from rice bran oil. Eur. J. Cancer Prev. 2003, 12, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Sami, S.A.; Salser, W.A.; Khan, F.A. Dose dependent suppression of serum cholesterol by tocotrienol-rich fraction (TRF25) of rice bran in hypercholesterolemic humans. Atherosclerosis 2002, 161, 199–207. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.-X.; Li, Y.; Sun, A.-M.; Wang, F.-J.; Yu, G.-P. Hypolipidemic and Antioxidative Effects of Aqueous Enzymatic Extract from Rice Bran in Rats Fed a High-Fat and -Cholesterol Diet. Nutrients 2014, 6, 3696-3710. https://doi.org/10.3390/nu6093696
Wang Y-X, Li Y, Sun A-M, Wang F-J, Yu G-P. Hypolipidemic and Antioxidative Effects of Aqueous Enzymatic Extract from Rice Bran in Rats Fed a High-Fat and -Cholesterol Diet. Nutrients. 2014; 6(9):3696-3710. https://doi.org/10.3390/nu6093696
Chicago/Turabian StyleWang, Yu-Xin, Yang Li, An-Min Sun, Feng-Jiao Wang, and Guo-Ping Yu. 2014. "Hypolipidemic and Antioxidative Effects of Aqueous Enzymatic Extract from Rice Bran in Rats Fed a High-Fat and -Cholesterol Diet" Nutrients 6, no. 9: 3696-3710. https://doi.org/10.3390/nu6093696