Effects of Oils Rich in Linoleic and α-Linolenic Acids on Fatty Acid Profile and Gene Expression in Goat Meat

Abstract
:1. Introduction
2. Experimental Section
2.1. Animal Welfare
2.1.1. Animals, Diets, and Management
2.1.2. Chemical Analyses

{kind=link}
{kind=link}
{kind=link}
| Diets | LLNA | MLNA | HLNA |
|---|---|---|---|
| Ingredients (% of DM) | |||
| Oil palm frond silage | 30.00 | 30.00 | 30.00 |
| Corn, grain | 17.00 | 17.00 | 17.00 |
| Soybean meal | 13.30 | 13.30 | 13.30 |
| Palm kernel cake | 25.11 | 25.11 | 25.11 |
| Rice bran | 8.18 | 8.18 | 8.18 |
| Flaxseed oil | 0.00 | 0.40 | 1.30 |
| Palm kernel oil | 1.10 | 1.00 | 0.10 |
| Sunflower oil | 2.30 | 2.00 | 2.00 |
| Mineral premix | 0.50 | 0.50 | 0.50 |
| Vitamin premix | 0.50 | 0.50 | 0.50 |
| Ammonium chloride | 1.00 | 1.00 | 1.00 |
| Limestone | 1.00 | 1.00 | 1.00 |
| Chemical Composition | |||
| ME (Mcal/Kg) 1 | 2.51 | 2.51 | 2.51 |
| CP% | 13.00 | 13.00 | 13.00 |
| EE% | 7.00 | 7.00 | 7.00 |
| NDF% | 48.90 | 48.90 | 48.90 |
| ADF% | 33.00 | 33.00 | 33.00 |
| Ca% | 0.68 | 0.68 | 0.68 |
| P% | 0.36 | 0.36 | 0.36 |
| Fatty Acid Composition (% of Total Identified Fatty Acids) | |||
| C10:0, Capric | 0.90 | 0.90 | 0.53 |
| C12:0, Lauric | 7.04 | 7.12 | 3.53 |
| C14:0, Myristic | 3.04 | 3.73 | 1.79 |
| C16:0, Palmitic | 16.14 | 15.36 | 15.65 |
| C16:1, Palmitoleic | 0.22 | 0.24 | 0.21 |
| C17:0, Margaric | 0.29 | 0.28 | 0.29 |
| C18:0, Stearic | 5.85 | 5.88 | 5.68 |
| C18:1 n-9, Oleic | 27.40 | 27.62 | 27.78 |
| C18:2 n-6, Linoleic | 35.68 | 32.40 | 30.92 |
| C18:3 n-3, α-Linolenic | 3.44 | 6.47 | 13.63 |
2.1.3. Measurement of FA
2.1.4. Tissue Collection and RNA Extraction and Purification and Real-Time Polymerase Chain Reaction (PCR)
| Target Group | Sequence 5′—3′ | Length, nt | Reference | |
|---|---|---|---|---|
| β-actin | F | CGC CAT GGA TGA TGA TAT TGC3 | 123 | [23] |
| R | AAG CGG CCT TGC ACA T3 | |||
| PPARα | F | TGC CAA GAT CTG AAA AAG CA | 101 | [33] |
| R | CCT CTT GGC CAG AGA CTT GA | |||
| PPARγ | F | CTT GCT GTG GGG ATG TCT C | 121 | [33] |
| R | GGT CAG CAG ACT CTG GGT TC | |||
| SCD | F | CCC AGC TGT CAG AGA AAA GG | ||
| R | GAT GAA GCA CAA CAG CAG GA | 115 | [33] |
2.2. Data Analysis
3. Results and Discussion
3.1. Nutrient and Fatty Acid Composition of Experimental Diets
3.2. Fatty Acid Composition of ST Muscle
3.2.1. n-3 and n-6 Polyunsaturated Fatty Acids
3.2.2. Triene and Diene Biohydrogenation Products
| ST | LLNA | MLNA | HLNA | SE | p-Value |
|---|---|---|---|---|---|
| C10:0, Capric | 0.35 | 0.33 | 0.32 | 0.02 | 0.75 |
| C12:0, Lauruic | 3.55 | 3.31 | 3.15 | 0.22 | 0.53 |
| C14:0, Myristic | 3.53 | 2.69 | 1.39 | 0.23 | 0.04 |
| C14:1, Myristoleic | 0.36 | 0.58 | 0.29 | 0.10 | 0.05 |
| C15:0, Pentadecanoic | 0.44 | 0.59 | 0.59 | 0.08 | 0.55 |
| C15:1, Pentadecenoic | 0.41 | 0.36 | 0.40 | 0.10 | 0.16 |
| C16:0, Palmitic | 21.55 | 21.63 | 20.31 | 1.35 | 0.14 |
| C16:1 n-7, Plamitoleic | 1.64 | 1.94 | 1.53 | 0.14 | 0.46 |
| C17:0, Margaric | 1.00 | 1.16 | 0.92 | 0.23 | 0.29 |
| C17:1, Margaroleic | 1.09 | 0.75 | 0.70 | 0.10 | 0.51 |
| C18:0, Stearic | 12.00 | 12.44 | 11.54 | 0.68 | 0.06 |
| C18:1 n-9, Oleic | 34.66 | 36.16 | 40.77 | 1.76 | 0.03 |
| C18:1 t-11 Vaccenic | 1.99 | 1.33 | 0.95 | 0.17 | 0.04 |
| C18:2n-6, Linoleic | 10.92 | 9.99 | 9.47 | 0.33 | 0.02 |
| CLA c-9 t-11 | 1.22 | 0.77 | 0.50 | 0.12 | 0.001 |
| CLA c-12 t-10 | 0.53 | 0.46 | 0.42 | 0.06 | 0.07 |
| C18:3n-3, α-Linolenic | 0.54 | 0.87 | 2.00 | 0.11 | 0.001 |
| CLNA c-9,t-11,c-15 | 0.28 | 0.33 | 0.41 | 0.03 | 0.03 |
| CLNA c-9,t-11,t-15 | 0.03 | 0.05 | 0.07 | 0.01 | 0.18 |
| C20:4 n-6, Arachidonic | 3.51 | 3.31 | 2.73 | 0.17 | 0.02 |
| C20:5 n-3, Eicosapentaenoic | 0.28 | 0.67 | 0.74 | 0.07 | 0.005 |
| C22:5n-3, Docosapentaenoic | 0.05 | 0.04 | 0.07 | 0.01 | 0.05 |
| C22:6n-3, Docosahexaenoic | 0.38 | 0.63 | 1.21 | 0.08 | 0.04 |
| SFA | 42.42 | 42.14 | 38.22 | 1.63 | 0.04 |
| MUFA | 40.15 | 41.12 | 44.64 | 1.80 | 0.02 |
| PUFAn-3 | 1.24 | 2.21 | 4.03 | 0.20 | 0.001 |
| PUFAn-6 | 14.43 | 12.88 | 12.20 | 9.04 | 0.01 |
| Total PUFA | 15.68 | 15.09 | 16.23 | 0.90 | 0.06 |
| Total trans FA | 1.99 | 1.33 | 0.95 | 0.17 | 0.01 |
| Total CLA | 1.75 | 1.23 | 0.92 | 0.14 | 0.001 |
| Total CLNA | 0.31 | 0.38 | 0.48 | 0.03 | 0.04 |
| n-6:n-3 FAR | 11.67 | 6.25 | 3.07 | 0.42 | 0.03 |
| PUFA:SFA Ratio | 0.37 | 0.37 | 0.43 | 0.02 | 0.57 |
3.2.3. Monounsaturated Fatty Acids (MUFA)
3.2.4. Saturated Fatty Acids (SFA)
3.2.5. Nutritional Quality of Meat Fatty Acids
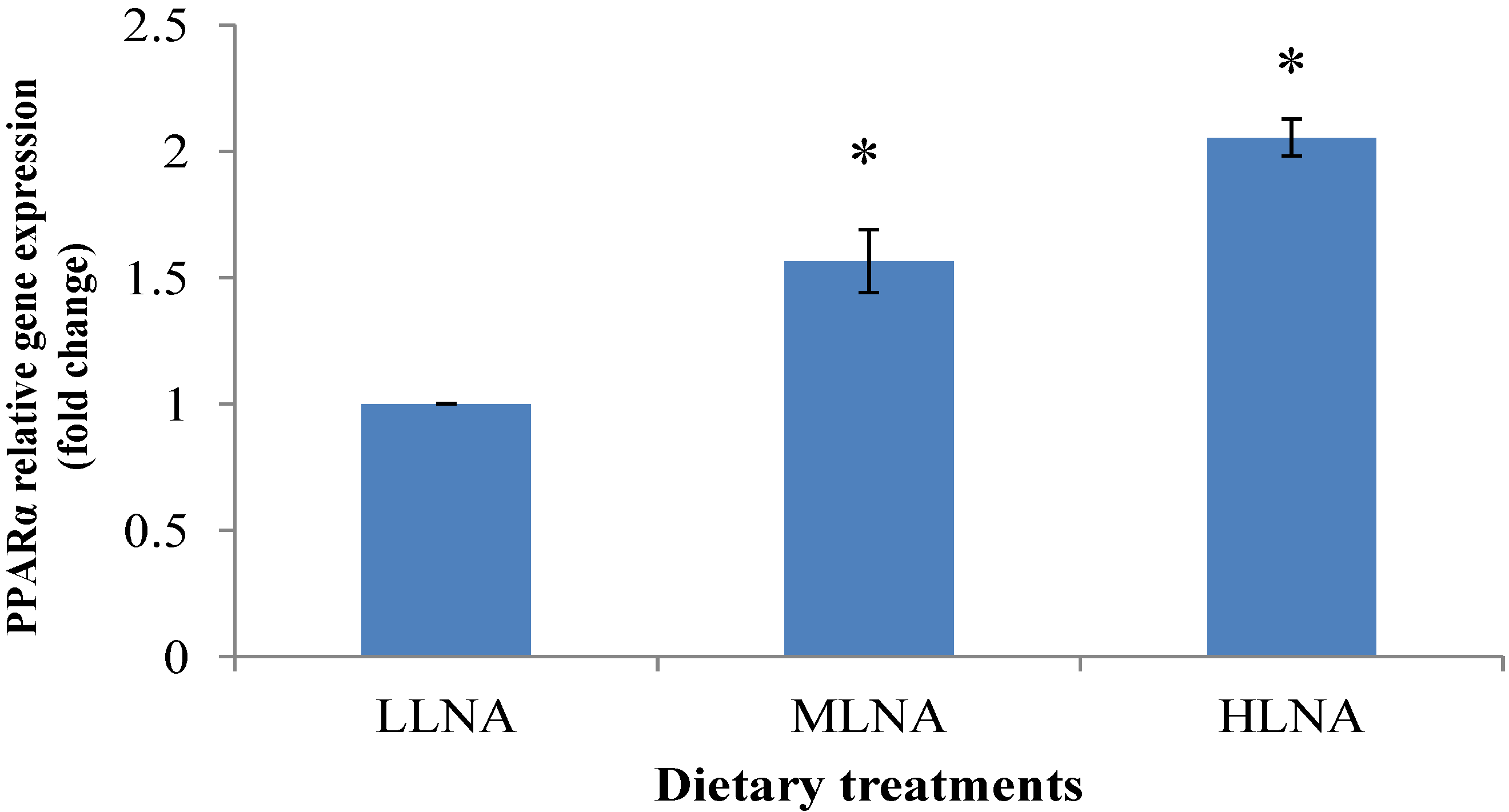
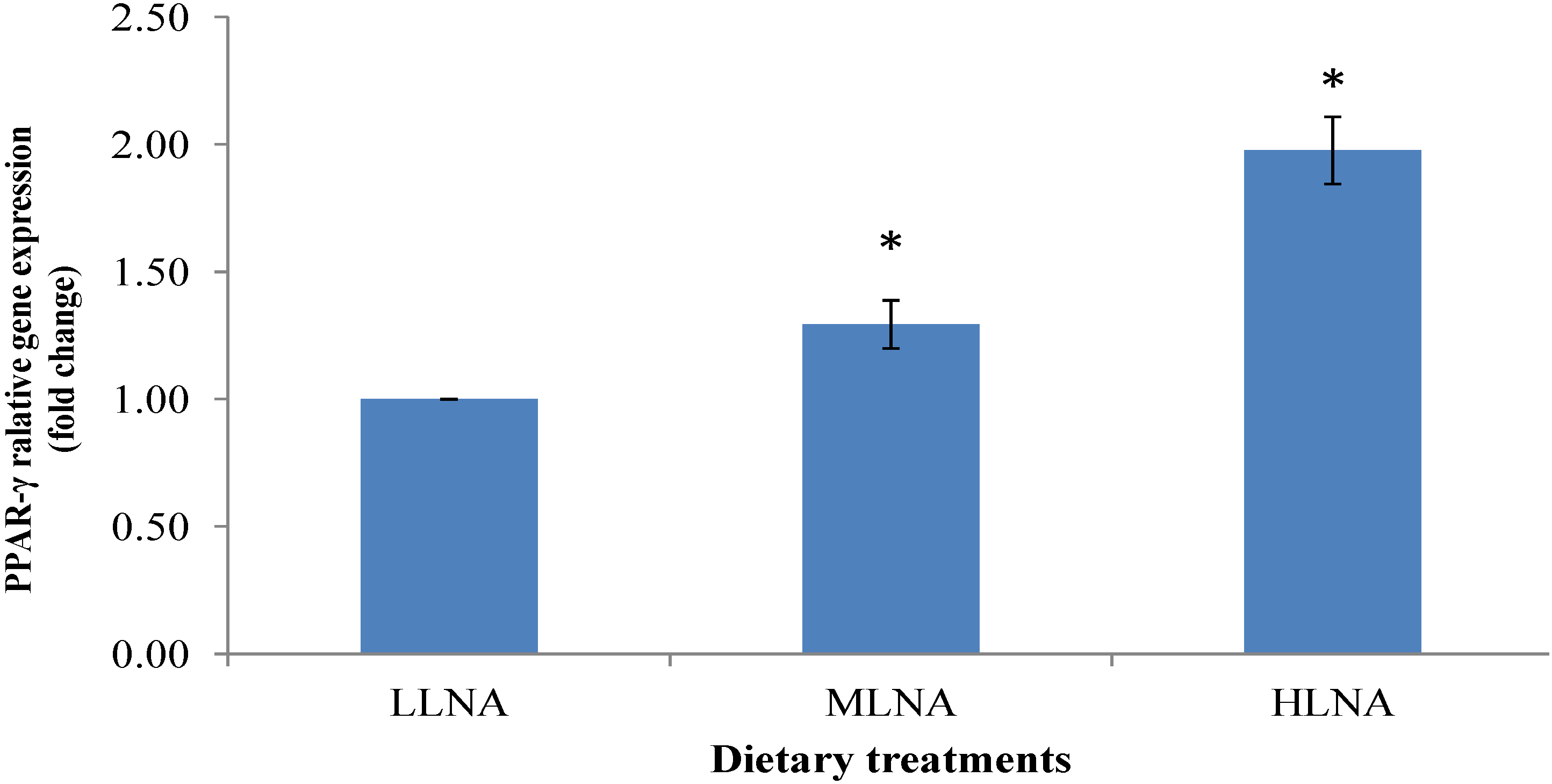
3.3. Effect of Dietary n-6:n-3 FAR on mRNA Expression of PPARα, PPARγ and SCD in ST Muscle

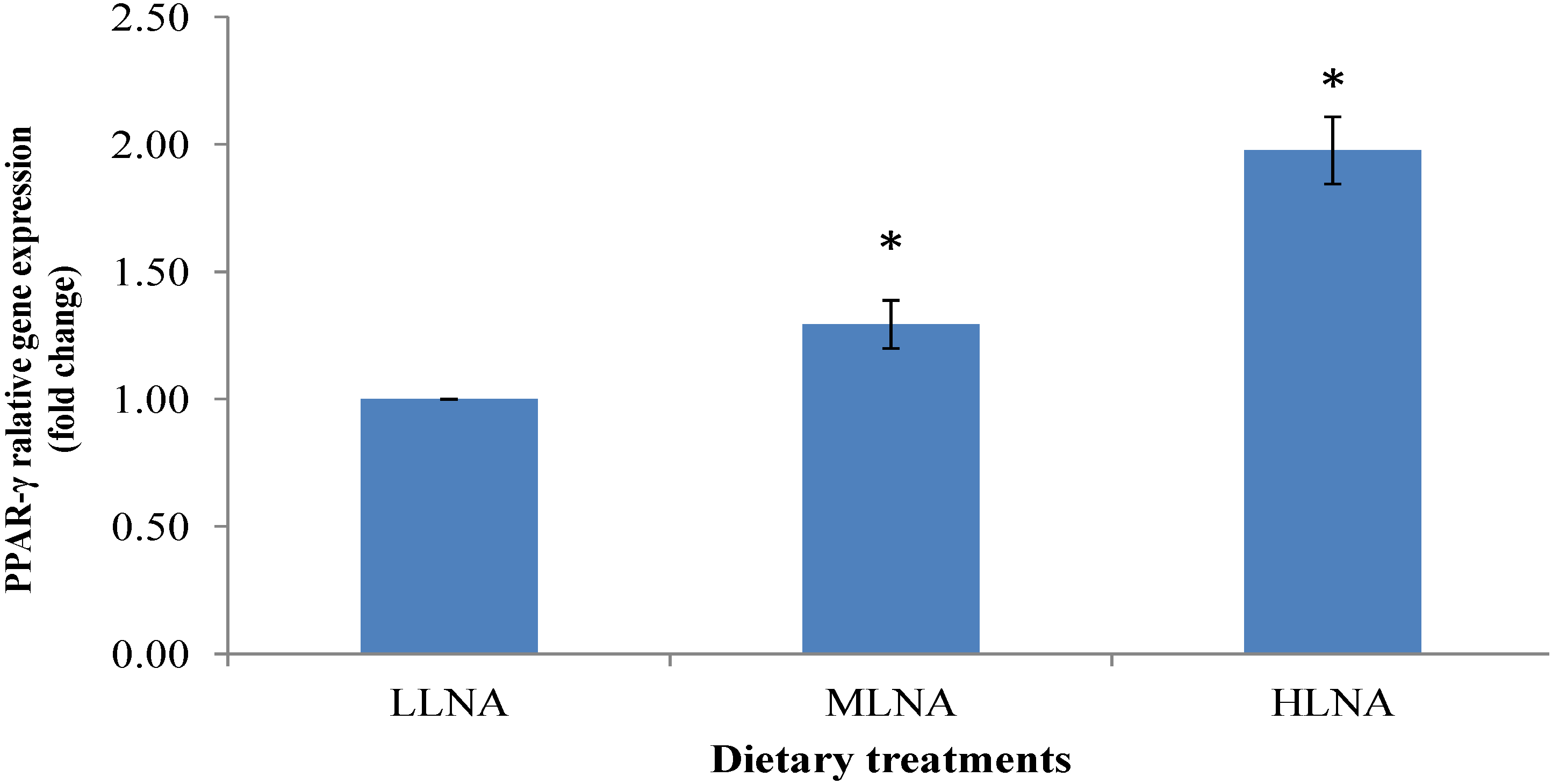
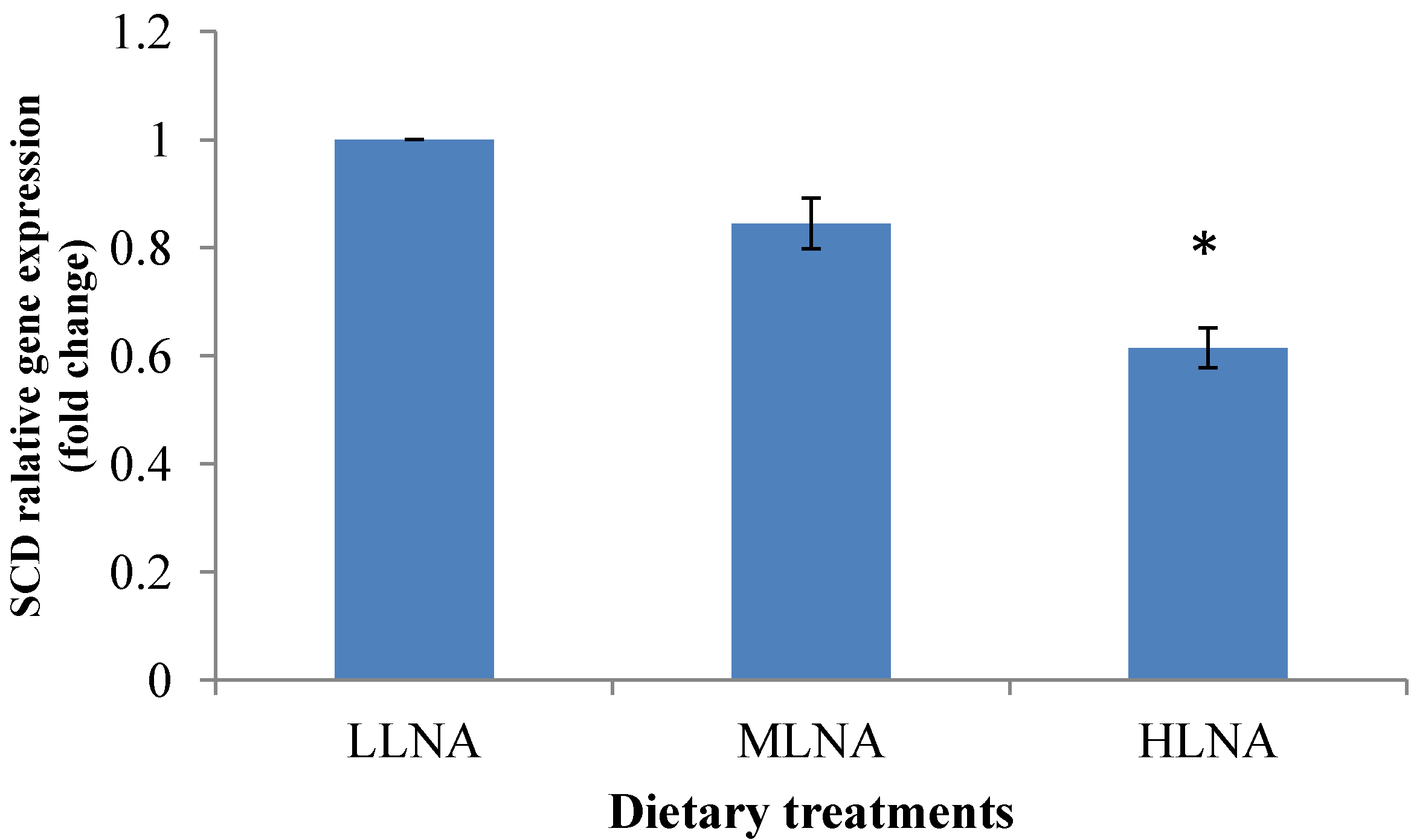
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ng, T.K.W. Towards improved fat intake and nutrition for Malaysians. Malays. J. Nutr. 1995, 1, 21–30. [Google Scholar]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Meta-analysis of prospective cohort studies evaluating the association of saturated fat with cardiovascular disease. Am. J. Clin. Nutr. 2010, 91, 535–546. [Google Scholar]
- Wahle, K.W.J.; Heys, S.D.; Rotondo, D. Conjugated linoleic acids: Are they beneficial or detrimental to health? Prog. Lipid Res. 2004, 43, 553–587. [Google Scholar]
- Cutrignelli, M.I.; Calabrò, S.; Bovera, F.; Tudisco, R.; D’Urso, S.; Marchiello, M.; Piccolo, V.; Infascelli, F. Effects of two protein sources and energy level of diet on the performance of young Marchigiana bulls. 2. Meat quality. Ital. J. Anim. Sci. 2010, 7, 271–286. [Google Scholar]
- De la Vega, F.; Guzmán, J.L.; Delgado-Pertíñez, M.; Zarazaga, L.A.; Argüello, A. Fatty acid composition of muscle and adipose tissues of organic and conventional Blanca Andaluza suckling kids. Span. J. Agric. Res. 2013, 11, 770–779. [Google Scholar]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar]
- Simopoulos, A.P.; Leaf, A.; Salem, N., Jr. Essentiality of and recommended dietary intakes for omega-6 and omega-3 fatty acids. Ann. Nutr. Metab. 1999, 43, 127–130. [Google Scholar]
- Givens, D.I.; Kliem, K.E.; Gibbs, R.A. The role of meat as a source of n-3 polyunsaturated fatty acids in the human diet. Meat Sci. 2006, 74, 209–218. [Google Scholar]
- Wood, J.D.; Enser, M. Factors Influencing Fatty Acids in Meat and the Role of Antioxidants in Improving Meat Quality. Br. J. Nutr. 1997, 78, S49–S60. [Google Scholar]
- Deckelbaum, R.J.; Torrejon, C. The omega-3 fatty acid nutritional landscape: Health benefits and sources. J. Nutr. 2012, 142, 587S–591S. [Google Scholar]
- Givens, D.I.; Gibbs, R.A. Current intakes of EPA and DHA in European populations and the potential of animal-derived foods to increase them. Proc. Nutr. Soc. 2008, 67, 273–280. [Google Scholar]
- Tudisco, R.; Cutrignelli, M.I.; Calabrò, S.; Piccolo, G.; Bovera, F.; Guglielmelli, A.; Moniello, G.; Infascelli, F. Influence of organic systems on milk fatty acid profile and CLA in goats. Small Rumin. Res. 2010, 88, 151–155. [Google Scholar]
- Xu, J.; Gao, H.; Song, L.; Yang, W.; Chen, C.; Deng, Q.; Huang, Q.; Yang, J.; Huang, F. Flaxseed oil and α-lipoic acid combination ameliorates hepatic oxidative stress and lipid accumulation in comparison to lard. Lipids Health Dis. 2013, 12, 58. [Google Scholar]
- Legrand, P.; Schmitt, B.; Mourot, J.; Catheline, D.; Chesneau, G.; Mireaux, M.; Kerhoas, N.; Weill, P. The consumption of food products from linseed-fed animals maintains erythrocyte omega-3 fatty acids in obese humans. Lipids 2010, 45, 11–19. [Google Scholar]
- Park, Y. Conjugated linoleic acid (CLA): Good or bad trans fat? J. Food Compos. Anal. 2009, 22, S4–S12. [Google Scholar]
- Nassu, R.T.; Dugan, M.E.R.; He, M.L.; McAllister, T.A.; Aalhus, J.L.; Aldai, N.; Kramer, J.K.G. The effects of feeding flaxseed to beef cows given forage based diets on fatty acids of longissimus thoracis muscle and backfat. Meat Sci. 2011, 89, 469–477. [Google Scholar]
- Göttlicher, M.; Widmark, E.; Li, Q.; Gustafsson, J.A. Fatty acids activate a chimera of the clofibric acid-activated receptor and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 4653–4657. [Google Scholar]
- Issemann, I.; Green, S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature 1990, 347, 645–650. [Google Scholar]
- Hajjar, T.; Meng, G.; Rajion, M.; Vidyadaran, S.; Othman, F.; Farjam, A.; Li, T.; Ebrahimi, M. Omega 3 polyunsaturated fatty acid improves spatial learning and hippocampal Peroxisome Proliferator Activated Receptors (PPARα and PPARγ) gene expression in rats. BMC Neuroscience 2012, 13, 109. [Google Scholar]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar]
- Igarashi, M.; Ma, K.; Chang, L.; Bell, J.M.; Rapoport, S.I. Dietary n-3 PUFA deprivation for 15 weeks upregulates elongase and desaturase expression in rat liver but not brain. J. Lipid Res. 2007, 48, 2463–2470. [Google Scholar]
- Graugnard, D.; Piantoni, P.; Bionaz, M.; Berger, L.; Faulkner, D.; Loor, J. Adipogenic and energy metabolism gene networks in longissimus lumborum during rapid post-weaning growth in Angus and Angus × Simmental cattle fed high-starch or low-starch diets. BMC Genomics 2009, 10, 142. [Google Scholar]
- Waters, S.M.; Kelly, J.P.; O’Boyle, P.; Moloney, A.P.; Kenny, D.A. Effect of level and duration of dietary n-3 polyunsaturated fatty acid supplementation on the transcriptional regulation of Δ9-desaturase in muscle of beef cattle. J. Anim. Sci. 2009, 87, 244–252. [Google Scholar]
- Ntambi, J.M. Regulation of stearoyl-CoA desaturase by polyunsaturated fatty acids and cholesterol. J. Lipid Res. 1999, 40, 1549–1558. [Google Scholar]
- NRC. Nutrient Requirements of Small Ruminant, 6th ed.; National Academy Press: Washington, DC, USA, 2007; p. 384. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Herlick, K., Ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990; p. 1230. [Google Scholar]
- Van Soest, P.; van; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanely, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Rajion, M.A.; McLean, J.G.; Cahill, R.N. Essential fatty acids in the fetal and newborn lamb. Aust. J. Biol. Sci. 1985, 38, 33–40. [Google Scholar]
- Ebrahimi, M.; Rajion, M.A.; Goh, Y.M.; Sazili, A.Q. Impact of different inclusion levels of oil palm (Elaeis guineensis Jacq.) fronds on fatty acid profiles of goat muscles. J. Anim. Physiol. Anim. Nutr. 2012, 96, 962–969. [Google Scholar]
- Destaillats, F.; Trottier, J.P.; Galvez, J.M.G.; Angers, P. Analysis of α-Linolenic Acid Biohydrogenation Intermediates in Milk Fat with Emphasis on Conjugated Linolenic Acids. J. Dairy Sci. 2005, 88, 3231–3239. [Google Scholar]
- Kramer, J.K.; Hernandez, M.; Cruz-Hernandez, C.; Kraft, J.; Dugan, M.E. Combining results of two GC separations partly achieves determination of all cis and trans 16:1, 18:1, 18:2 and 18:3 except CLA isomers of milk fat as demonstrated using Ag-ion SPE fractionation. Lipids 2008, 43, 259–273. [Google Scholar]
- Dervishi, E.; Serrano, C.; Joy, M.; Serrano, M.; Rodellar, C.; Calvo, J.H. The effect of feeding system in the expression of genes related with fat metabolism in semitendinous muscle in sheep. Meat Sci. 2011, 89, 91–97. [Google Scholar]
- Ebrahimi, M.; Rajion, M.A.; Goh, Y.M.; Sazili, A.Q.; Schonewille, J.T. Effect of linseed oil dietary supplementation on fatty acid composition and gene expression in adipose tissue of growing goats. BioMed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Mapiye, C.; Aalhus, J.L.; Turner, T.D.; Rolland, D.C.; Basarab, J.A.; Baron, V.S.; McAllister, T.A.; Block, H.C.; Uttaro, B.; Lopez-Campos, O.; et al. Effects of feeding flaxseed or sunflower-seed in high-forage diets on beef production, quality and fatty acid composition. Meat Sci. 2013, 95, 98–109. [Google Scholar]
- Bernard, L.; Bonnet, M.; Leroux, C.; Shingfield, K.J.; Chilliard, Y. Effect of sunflower-seed oil and linseed oil on tissue lipid metabolism, gene expression, and milk fatty acid secretion in alpine goats fed maize silage-based diets. J. Dairy Sci. 2009, 92, 6083–6094. [Google Scholar]
- Khas-Erdene, Q.; Wang, J.Q.; Bu, D.P.; Wang, L.; Drackley, J.K.; Liu, Q.S.; Yang, G.; Wei, H.Y.; Zhou, L.Y. Short communication: Responses to increasing amounts of free α-linolenic acid infused into the duodenum of lactating dairy cows. J. Dairy Sci. 2010, 93, 1677–1684. [Google Scholar]
- Dewhurst, R.J.; Scollan, N.D.; Lee, M.R.F.; Ougham, H.J.; Humphreys, M.O. Forage breeding and management to increase the beneficial fatty acid content of ruminant products. Proc. Nutr. Soc. 2003, 62, 329–336. [Google Scholar]
- Scollan, N.D.; Choi, N.J.; Kurt, E.; Fisher, A.V.; Enser, M.; Wood, J.D. Manipulating the fatty acid composition of muscle and adipose tissue in beef cattle. Br. J. Nutr. 2001, 85, 115–124. [Google Scholar]
- Jerónimo, E.; Alves, S.P.; Prates, J.A.M.; Santos-Silva, J.; Bessa, R.J.B. Effect of dietary replacement of sunflower oil with linseed oil on intramuscular fatty acids of lamb meat. Meat Sci. 2009, 83, 499–505. [Google Scholar]
- Burdge, G.C.; Calder, P.C. α-Linolenic acid metabolism in adult humans: the effects of gender and age on conversion to longer-chain polyunsaturated fatty acids. Eur. J. Lipid Sci. Technol. 2005, 107, 426–439. [Google Scholar]
- Brenner, R.R. Factors influencing fatty acid chain elongation and desaturation. In The Role of Fats in Human Nutrition; Podmore, J., Ed.; Acadamic Press: San Diego, CA, USA, 1989; pp. 45–80. [Google Scholar]
- Cooper, S.L.; Sinclair, L.A.; Wilkinson, R.G.; Hallett, K.G.; Enser, M.; Wood, J.D. Manipulation of the n-3 polyunsaturated fatty acid content of muscle and adipose tissue in lambs. J. Anim. Sci. 2004, 82, 1461–1470. [Google Scholar]
- Demirel, G.; Wachira, A.M.; Sinclair, L.A.; Wilkinson, R.G.; Wood, J.D.; Enser, M. Effects of dietary n-3 polyunsaturated fatty acids, breed and dietary vitamin E on the fatty acids of lamb muscle, liver and adipose tissue. Br. J. Nutr. 2004, 91, 551–565. [Google Scholar]
- Nute, G.R.; Richardson, R.I.; Wood, J.D.; Hughes, S.I.; Wilkinson, R.G.; Cooper, S.L.; Sinclair, L.A. Effect of dietary oil source on the flavour and the color and lipid stability of lamb meat. Meat Sci. 2007, 77, 547–555. [Google Scholar]
- Sampath, H.; Ntambi, J.M. Polyunsaturated fatty acid regulation of genes of lipid metabolism. Ann. Rev. Nutr. 2005, 25, 317–340. [Google Scholar]
- Bessa, R.J.B.; Alves, S.P.; Jerónimo, E.; Alfaia, C.M.; Prates, J.A.M.; Santos-Silva, J. Effect of lipid supplements on ruminal biohydrogenation intermediates and muscle fatty acids in lambs. Eur. J. Lipid Sci. Technol. 2007, 109, 868–878. [Google Scholar]
- Noci, F.; French, P.; Monahan, F.J.; Moloney, A.P. The Fatty Acid Composition of Muscle Fat and Subcutaneous Adipose Tissue of Grazing Heifers Supplemented with Plant Oil-Enriched Concentrates. J. Anim. Sci. 2007, 85, 1062–1073. [Google Scholar]
- Vasta, V.; Mele, M.; Serra, A.; Scerra, M.; Luciano, G.; Lanza, M.; Priolo, A. Metabolic fate of fatty acids involved in ruminal biohydrogenation in sheep fed concentrate or herbage with or without tannins. J. Anim. Sci. 2009, 87, 2674–2684. [Google Scholar]
- Aharoni, Y.; Orlov, A.; Brosh, A. Effects of high-forage content and oilseed supplementation of fattening diets on conjugated linoleic acid (CLA) and trans fatty acids profiles of beef lipid fractions. Anim. Feed Sci. Technol. 2004, 117, 43–60. [Google Scholar]
- Bartoň, L.; Marounek, M.; Kudrna, V.; Bureš, D.; Zahradkova, R. Growth performance and fatty acid profiles of intramuscular and subcutaneous fat from Limousin and Charolais heifers fed extruded linseed. Meat Sci. 2007, 76, 517–523. [Google Scholar]
- Food and Nutrition Board, Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; The National Academies Press: Washington, DC, USA, 2005; Volume 423. [Google Scholar]
- Raes, K.; de Smet, S.; Demeyer, D. Effect of dietary fatty acids on incorporation of long chain polyunsaturated fatty acids and conjugated linoleic acid in lamb, beef and pork meat: A review. Anim. Feed Sci. Technol. 2004, 113, 199–221. [Google Scholar]
- Jaudszus, A.; Jahreis, G.; Schlörmann, W.; Fischer, J.; Kramer, R.; Degen, C.; Rohrer, C.; Roth, A.; Gabriel, H.; Barz, D.; et al. Vaccenic acid-mediated reduction in cytokine production is independent of c9, t11-CLA in human peripheral blood mononuclear cells. Biochim. Biophys. Acta 2012, 1821, 1316–1322. [Google Scholar]
- Sofi, F.; Buccioni, A.; Cesari, F.; Gori, A.M.; Minieri, S.; Mannini, L.; Casini, A.; Gensini, G.F.; Abbate, R.; Antongiovanni, M. Effects of a dairy product (pecorino cheese) naturally rich in cis-9, trans-11 conjugated linoleic acid on lipid, inflammatory and haemorheological variables: A dietary intervention study. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 117–124. [Google Scholar]
- Dilzer, A.; Park, Y. Implication of conjugated linoleic acid (CLA) in human health. Crit. Rev. Food Sci. Nutr. 2012, 52, 488–513. [Google Scholar]
- Pawar, A.; Jump, D.B. Unsaturated fatty acid regulation of peroxisome proliferator-activated receptor alpha activity in rat primary hepatocytes. J. Biol. Chem. 2003, 278, 35931–35939. [Google Scholar]
- Al-Hasani, H.; Joost, H.G. Nutrition-/diet-induced changes in gene expression in white adipose tissue. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 589–603. [Google Scholar]
- Ntambi, J.M.; Miyazaki, M. Regulation of stearoyl-CoA desaturases and role in metabolism. Prog. Lipid Res. 2004, 43, 91–104. [Google Scholar]
- Kim, H.J.; Miyazaki, M.; Ntambi, J.M. Dietary cholesterol opposes PUFA-mediated repression of the stearoyl-CoA desaturase-1 gene by SREBP-1 independent mechanism. J. Lipid Res. 2002, 43, 1750–1757. [Google Scholar]
- Deiuliis, J.; Shin, J.; Murphy, E.; Kronberg, S.L.; Eastridge, M.L.; Suh, Y.; Yoon, J.-T.; Lee, K. Bovine adipose triglyceride lipase is not altered and adipocyte fatty acid-binding protein is increased by dietary flaxseed. Lipids 2010, 45, 963–973. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ebrahimi, M.; Rajion, M.A.; Goh, Y.M. Effects of Oils Rich in Linoleic and α-Linolenic Acids on Fatty Acid Profile and Gene Expression in Goat Meat. Nutrients 2014, 6, 3913-3928. https://doi.org/10.3390/nu6093913
Ebrahimi M, Rajion MA, Goh YM. Effects of Oils Rich in Linoleic and α-Linolenic Acids on Fatty Acid Profile and Gene Expression in Goat Meat. Nutrients. 2014; 6(9):3913-3928. https://doi.org/10.3390/nu6093913
Chicago/Turabian StyleEbrahimi, Mahdi, Mohamed Ali Rajion, and Yong Meng Goh. 2014. "Effects of Oils Rich in Linoleic and α-Linolenic Acids on Fatty Acid Profile and Gene Expression in Goat Meat" Nutrients 6, no. 9: 3913-3928. https://doi.org/10.3390/nu6093913