Occurrence of Pre- and Post-Harvest Mycotoxins and Other Secondary Metabolites in Danish Maize Silage

Abstract
:
1. Introduction
2. Results and Discussion
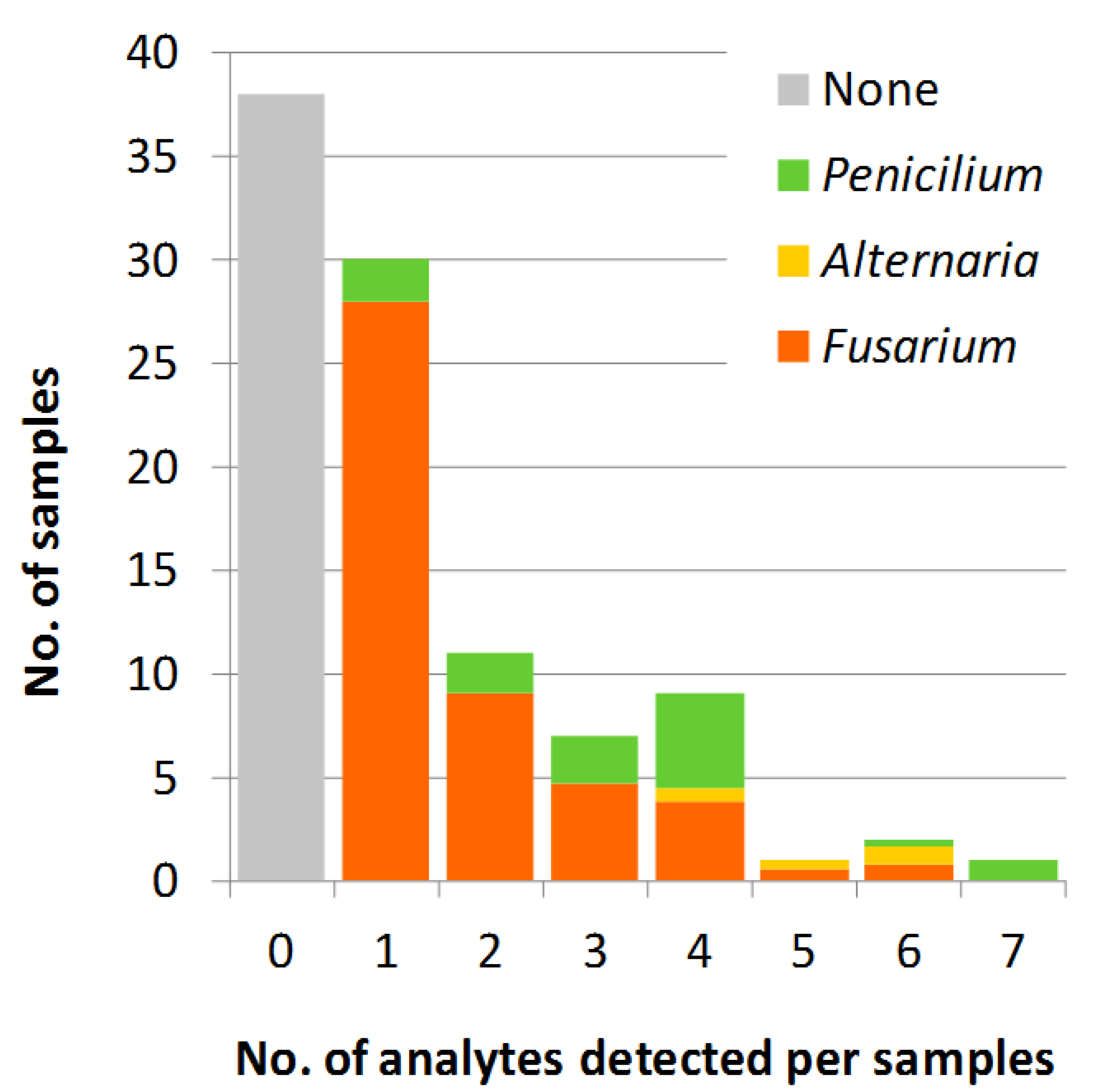
2.1. Method Performance

{kind=link}
{kind=link}
| Analyte | Abbreviation | Mean | Reproducibility | LOD (µg·kg−1) | LOQ (µg·kg−1) | |
|---|---|---|---|---|---|---|
| Recovery (%) | RSDIR (%) | |||||
| Quantitative | Alternariol | AOH | 78 | 14 | 10 | 20 |
| Alternariol momomethyl ether | AME | 79 | 10 | 6 | 12 | |
| Andrastin A | AND A | 122 | 12 | 1 | 2 | |
| Cyclopiazonic acid | CPA | 63 | 35 | 15 | 30 | |
| Deoxynivalenol | DON | 83 | 18 | 739 | 1478 | |
| Enniatin B | ENN B | 60 | 24 | 24 | 48 | |
| Fumitremorgin A | FUT A | 93 | 23 | 76 | 152 | |
| Gliotoxin | GLI | 85 | 13 | 71 | 142 | |
| Mevinolin | MEV | 68 | 27 | 25 | 50 | |
| Mycophenolic acid | MPA | 90 | 13 | 7 | 14 | |
| Nivalenol | NIV | 68 | 15 | 122 | 244 | |
| Ochratoxin A | OTA | 71 | 9 | 10 | 20 | |
| Patulin | PAT | 100 | 17 | 371 | 742 | |
| Penitrem A | PEN A | 107 | 12 | 8 | 16 | |
| Roquefortine C | ROQ C | 205 | 25 | 158 | 316 | |
| Sterigmatocystin | STE | 72 | 9 | 8 | 16 | |
| T-2 toxin | T-2 | 55 | 26 | 96 | 192 | |
| Tenuazonic acid | TEA | 37 | 20 | 121 | 242 | |
| Zearalenone | ZEA | 90 | 16 | 9 | 18 | |
| Qualitative | Altersetin | ALS | 91 | 14 | - | - |
| Citreoisocoumarin | CICO | 84 | 7 | - | - | |
| Fumigaclavine A | FUC A | 93 | 21 | - | - | |
| Fumigaclavine C | FUC C | 176 | 13 | - | - | |
| Marcfortine A | MAC A | 63 | 16 | - | - | |
| Marcfortine B | MAC B | 61 | 9 | - | - | |
| PR-toxin | PR | 56 | 32 | - | - | |
| Roquefortine A | ROQ A | 103 | 32 | - | - |
2.2. Mycotoxins in Maize and Maize Silage
| Fresh Maize (n = 17) | Ensiled Maize (n = 82) | Total (n = 99) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compound | npos | avgpos (SEM) | Max | npos | avgpos (SEM) | Max | npos | avgpos (SEM) | Max | |
| Quantitative | AME | 1 | 11 | 11 | 2 | 8(1) | 8.8 | 3 | 9(1) | 11 |
| AND A | 15 | 169(54) | 691 | 15 | 169(54) | 691 | ||||
| AOH | 2 | 18(6) | 24 | 2 | 18(6) | 24 | ||||
| DON | 2 | 2369(293) | 2,662 | 5 | 1629(365) | 2,974 | 7 | 1841(293) | 2974 | |
| ENN B | 8 | 128(40) a | 365 | 20 | 53(7) b | 152 | 28 | 75(13) | 365 | |
| MPA | 2 | 43(9) | 52 | 2 | 43(9) | 52 | ||||
| NIV | 5 | 255(37) | 351 | 11 | 266(53) | 758 | 16 | 263(38) | 758 | |
| ROQ C | 2 | 173(15) | 189 | 2 | 173(15) | 189 | ||||
| ZEA | 11 | 83(59) | 666 | 23 | 66(15) | 311 | 34 | 71(21) | 666 | |
| Qualitative | CICO | 1 | 7 | 8 | ||||||
| MAC A | 6 | 6 | ||||||||
| MAC B | 1 | 1 | ||||||||
| ROQ A | 9 | 9 | ||||||||
2.2.1. Fusarium Toxins
2.2.2. Penicillium Toxins
2.2.3. Alternaria Toxins
2.2.4. Other Fungal Metabolites
2.2.5. Multiple Mycotoxins in the Same Samples

2.2.6. Sample Origin and Storage Effects
3. Experimental Section
3.1. Sample Collection and Preparation
3.2. Extraction
3.3. Sample Analysis
3.4. Data Analysis
4. Conclusions
Supplementary Files
Supplementary File 1Abbreviations
| ALS | altersetin |
| AME | alternariol monomethyl ether |
| AND A | andrastin A |
| AOH | alternariol |
| CICO | citreoisocoumarine |
| CPA | cyclopiazonic acid |
| DAS | diacetoxyscirpenol |
| DON | deoxynivalenol |
| ENN B | enniatin B |
| FUC A | fumigaclavine A |
| FUC C | fumigaclavine C |
| FUT A | fumitremorgin A |
| GLI | gliotoxin |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| LOD | limit of detection |
| LOQ | limit of quantification |
| MAC A | marcfortine A |
| MAC B | marcfortine B |
| MEV | mevinolin |
| MPA | mycophenolic acid |
| NIV | nivalenol |
| OTA | ochratoxin A |
| PAT | patulin |
| PEN A | penitrem A |
| PR | PR toxin |
| QuEChERS | Quick, Easy, Cheap, Effective, Rugged, Safe, a multi-method developed for the analysis of pesticide residues in fruit and vegetables |
| ROQ A | roquefortine A |
| ROQ C | roquefortine C |
| RSD | relative standard deviation |
| RSDIR | the relative standard deviation under intra-laboratory reproducibility conditions |
| SDIR | standard deviation at intra-laboratory conditions |
| STE | sterigmatocystin |
| T-2 | T-2 toxin |
| TEA | tenuazonic acid |
| ZEA | zearalenone |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wilkinson, J.M.; Toivonen, M.I. World Silage: A Survey of Forage Conservation around the World; Chalcombe Publications: Lincoln, UK, 2003. [Google Scholar]
- Eastridge, M.L. Major advances in applied dairy cattle nutrition. J. Dairy Sci. 2006, 89, 1311–1323. [Google Scholar] [CrossRef]
- Cheli, F.; Campagnoli, A.; Dell’Orto, V. Fungal populations and mycotoxins in silages: From occurrence to analysis. Anim. Feed Sci. Tech. 2013, 183, 1–16. [Google Scholar] [CrossRef]
- Storm, I.M.; Sørensen, J.L.; Rasmussen, R.R.; Nielsen, K.F.; Thrane, U. Mycotoxins in silage. Stewart Posthar. Rev. 2008, 4. [Google Scholar] [CrossRef]
- Korosteleva, S.N.; Smith, T.K.; Boermans, H.J. Effects of feed naturally contaminated with Fusarium mycotoxins on metabolism and immunity of dairy cows. J. Dairy Sci. 2009, 92, 1585–1593. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef]
- Scudamore, K.A.; Livesey, C.T. Occurrence and significance of mycotoxins in forage crops and silage: A review. J. Sci. Food. Agric. 1998, 77, 1–17. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Jouany, J.-P. Mycotoxins in feeds and their fate in animals: A review. Anim. Res. 2002, 51, 81–99. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Riley, R.T. An historical overview of field disease outbreaks known or suspected to be caused by consumption of feeds contaminated with Fusarium toxins. Anim. Feed Sci. Technol. 2007, 137, 201–212. [Google Scholar] [CrossRef]
- Rasmussen, R.R.; Rasmussen, P.H.; Larsen, T.O.; Bladt, T.T.; Binderup, M.L. In vitro toxicity of fungi spoiling maize silage. Food Chem. Toxicol. 2011, 49, 31–44. [Google Scholar] [CrossRef]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Giffel, M.C.T. Occurrence of mycotoxins in maize, grass and wheat silage for dairy cattle in the Netherlands. Food Addit. Cont. Part. B 2008, 1, 41–50. [Google Scholar] [CrossRef]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Giffel, M.C.T. Occurrence of Mycotoxins in Feedstuffs of Dairy Cows and Estimation of Total Dietary Intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar] [CrossRef]
- Garon, D.; Richard, E.; Sage, L.; Bouchart, V.; Pottier, D.; Lebailly, P. Mycoflora and multimycotoxin detection in corn silage: Experimental study. J. Agric. Food Chem. 2006, 54, 3479–3484. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Jones, A.D.; Kuldau, G.A. Contamination of fresh and ensiled maize by multiple Penicillium mycotoxins. Phytopathology 2008, 98, 330–336. [Google Scholar] [CrossRef]
- Monbaliu, S.; Poucke, C.V.; Detavernier, C.; Dumoulin, F.; Velde, M.V.D.; Schoeters, E.; Dyck, S.V.; Averkieva, O.; Peteghem, C.V.; Saeger, S.D. Occurrence of mycotoxins in feed as analyzed by a multi-mycotoxin LC-MS/MS method. J. Agric. Food Chem. 2010, 58, 66–71. [Google Scholar] [CrossRef]
- Müller, H.M.; Amend, R. Formation and disappearance of mycophenolic acid, patulin, penicillic acid and PR toxin in maize silage inoculated with Penicillium roquefortii. Arch. Anim. Nutr. 1997, 50, 213–225. [Google Scholar]
- Placinta, C.M.; D’Mello, J.P.F.; Macdonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar]
- Rasmussen, R.R.; Storm, I.M.; Rasmussen, P.H.; Smedsgaard, J.; Nielsen, K.F. Multi-mycotoxin analysis of maize silage by LC-MS/MS. Anal. Bioanal. Chem. 2010, 397, 765–776. [Google Scholar] [CrossRef]
- Richard, E.; Heutte, N.; Sage, L.; Pottier, D.; Bouchart, V.; Lebailly, P.; Garon, D. Toxigenic fungi and mycotoxins in mature corn silage. Food Chem. Toxicol. 2007, 45, 2420–2425. [Google Scholar] [CrossRef]
- Schneweis, I.; Meyer, K.; Hörmansdorfer, S.; Bauer, J. Mycophenolic acid in silage. Appl. Environ. Microbiol. 2000, 66, 3639–3641. [Google Scholar] [CrossRef]
- Schollenberger, M.; Müller, H.-M.; Rüfle, M.; Suchy, S.; Plank, S.; Drochner, W. Natural occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany. Mycopathologia 2006, 161, 43–52. [Google Scholar] [CrossRef]
- Sørensen, J.L.; Nielsen, K.F.; Rasmussen, P.H.; Thrane, U. Development of a LC-MS/MS method for analysis of enniatins and beauvericin in whole fresh and ensiled maize. J. Agric. Food Chem. 2008, 56, 10439–10443. [Google Scholar] [CrossRef]
- Van Pamel, E.; Verbeken, A.; Vlaemynck, G.; de Boever, J.; Daeseleire, E. Ultrahigh-performance liquid chromatographic—Tandem mass spectrometric multimycotoxin method for quantitating 26 mycotoxins in maize silage. J. Agric. Food Chem. 2011, 59, 9747–9755. [Google Scholar] [CrossRef]
- FAO. Worldwide Regulations for Mycotoxins in Food and Feed in 2003; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- EU commission. Commission Recommendation 2006/576/EC of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding. 2006. Available online: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:229:0007:0009:EN:PDF (accessed on 19 July 2014).
- Cotty, P.J.; Jaime-Garcia, R. Influence of climate on aflatoxin producing fungi and aflatoxin contamination. Int. J. Food Microbiol. 2007, 119, 109–115. [Google Scholar] [CrossRef]
- Sørensen, J.L. Preharvest. fungi and their mycotoxins in maize. Ph.D. Thesis, Technical University of Denmark, Lyngby, Denmark, June 2009. [Google Scholar]
- Storm, I.M.; Kristensen, N.B.; Raun, B.M.; Smedsgaard, J.; Thrane, U. Dynamics in the microbiology of maize silage during whole-season storage. J. Appl. Microbiol. 2010, 109, 1017–1026. [Google Scholar] [CrossRef]
- EU Commission. Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on Undesirable Substances in Animal Feed. 2002. Available online: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2002:140:0010:0021:EN:PDF (accessed on 19 July 2014).
- EU Commission—SCF. Scientific Committee on Food: Opinion on Fusarium. Toxins—Part. 1: Deoxynivalenol. (DON). 1999. Available online: http://ec.europa.eu/food/fs/sc/scf/out44_en.pdf (accessed on 19 July 2014).
- EU Commission—SCF. Scientific Committee on Food: Opinion on Fusarium. Toxins—Part. 2: Zearalenone. (ZEA). 2000a. Available online: http://ec.europa.eu/food/fs/sc/scf/out65_en.pdf (accessed on 19 July 2014).
- EFSA. Scientific Opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar]
- Jestoi, M. Emerging fusarium-mycotoxins fusapriliferin, beauvericin, enniatins, and moniliformin: A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Mohr, A.I.; Lorenz, I.; Baum, B.; Hewicker-Trautwein, M.; Pfaffl, M.; Dzidic, A.; Meyer, H.H.D.; Bauer, J.; Meyer, K. Influence of oral application of mycophenolic acid on the clinical health status of sheep. J. Vet. Med. A 2007, 54, 76–81. [Google Scholar] [CrossRef]
- Tüller, G. Einfluss. von Roquefortin. C auf Tiergesundheid. und Lebensmittelqualität. bei Wiederkäuern. Ph.D. Thesis, Technische Universität München, München, Germany, August 2005. [Google Scholar]
- Cole, R.J.; Cox, R.H. Handbook of Toxic Fungal Metabolites; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Frisvad, J.C.; Thrane, U.; Samson, R.A.; Pitt, J.I. Important mycotoxins and the fungi which produce them. In Advances in Food Mycology; Hocking, A.D., Pit, J.I., Samson, R.A., Thrane, U., Eds.; Springer Science + Business Media Inc.: New York, NY, USA, 2006; pp. 3–31. [Google Scholar]
- Pfeiffer, E.; Eschbach, S.; Metzler, M. Alternaria toxins: DNA strand-breaking activity in mammalian cells in vitro. Mycotoxin Res. 2007, 23, 152–157. [Google Scholar] [CrossRef]
- Liu, G.T.; Qian, Y.Z.; Zhang, P.; Dong, W.H.; Qi, Y.M.; Guo, H.T. Etiological role of Alternaria alternata in human esophageal cancer. Chin. Med. J. 1992, 105, 394–400. [Google Scholar]
- Pereyra, C.M.; Alonso, V.A.; Rosa, C.A.R.; Chiacchiera, S.M.; Dalcero, A.M.; Cavaglieri, L.R. Gliotoxin natural incidence and toxigenicity of Aspergillus fumigatus isolated from corn silage and ready dairy cattle feed. World Mycotoxin J. 2008, 1, 457–462. [Google Scholar] [CrossRef]
- EU Commission—SCF. Scientific Committee on Food: Opinion on Fusarium Toxins—Part. 4: Nivalenol. 2000b. Available online: http://ec.europa.eu/food/fs/sc/scf/out74_en.pdf (accessed on 19 July 2014).
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- EFSA. Deoxynivalenol in food and feed: Occurrence and exposure. EFSA J. 2013, 11, 3379. [Google Scholar]
- Corrier, E. Mycotoxicosis: Mechanisms of immunosuppression. Vet. Immunol. Immunopathol. 1991, 30, 73–87. [Google Scholar] [CrossRef]
- EU Commission—SCF. Opinion of the Scientific Committee on Food on Fusarium Toxins Part. 6: Group Evaluation of T-2 toxin, HT-2 Toxin, Nivalenol and Deoxynivalenol. 2002. Available online: http://ec.europa.eu/food/fs/sc/scf/out123_en.pdf (accessed on 19 July 2014).
- Mansfield, M.A.; Kuldau, G.A. Microbiological and molecular determination of mycobiota in fresh and ensiled maize. Mycologia 2007, 99, 269–278. [Google Scholar] [CrossRef]
- Raun, B.M.L.; Kristensen, N.B. Prevalence of propanol fermentation in maize silage. Acta Agric. Scand. Sect. A Anim. Sci. 2010, 60, 53–59. [Google Scholar]
- Kristensen, N.B.; Sloth, K.H.; Højberg, O.; Spliid, N.H.; Jensen, C.; Thøgersen, R. Effects of microbial inoculants on corn silage fermentation, microbial contents, aerobic stability, and milk production under field conditions. J. Dairy Sci. 2010, 93, 3764–3774. [Google Scholar] [CrossRef]
- Lehotay, S.J.; de Kok, A.; Hiemstra, M.; van Bodegraven, P. Validation of a fast and easy method for the determination of residues from 229 pesticides in fruits and vegetables using gas and liquid chromatography and mass spectrometric detection. J. AOAC Int. 2005, 88, 595–614. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Storm, I.M.L.D.; Rasmussen, R.R.; Rasmussen, P.H. Occurrence of Pre- and Post-Harvest Mycotoxins and Other Secondary Metabolites in Danish Maize Silage. Toxins 2014, 6, 2256-2269. https://doi.org/10.3390/toxins6082256
Storm IMLD, Rasmussen RR, Rasmussen PH. Occurrence of Pre- and Post-Harvest Mycotoxins and Other Secondary Metabolites in Danish Maize Silage. Toxins. 2014; 6(8):2256-2269. https://doi.org/10.3390/toxins6082256
Chicago/Turabian StyleStorm, Ida M. L. Drejer, Rie Romme Rasmussen, and Peter Have Rasmussen. 2014. "Occurrence of Pre- and Post-Harvest Mycotoxins and Other Secondary Metabolites in Danish Maize Silage" Toxins 6, no. 8: 2256-2269. https://doi.org/10.3390/toxins6082256