Rodents Versus Pig Model for Assessing the Performance of Serotype Chimeric Ad5/3 Oncolytic Adenoviruses

Abstract
:1. Introduction
2. Results
2.1. Analysis of Virus-Cell Binding
2.2. Analysis of Gene Transduction
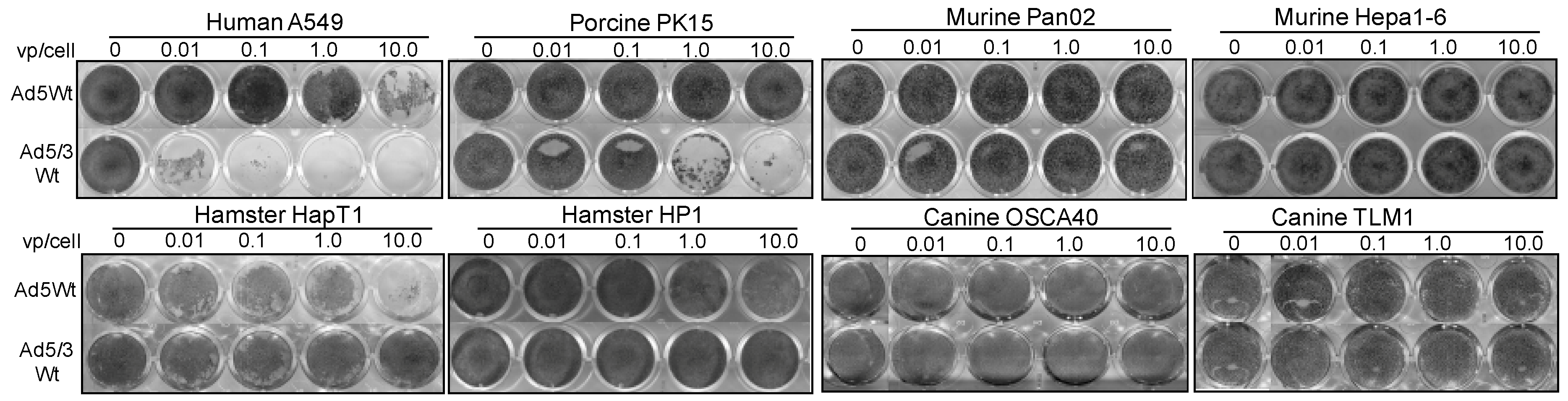
2.3. Replication-Mediated Cell Killing
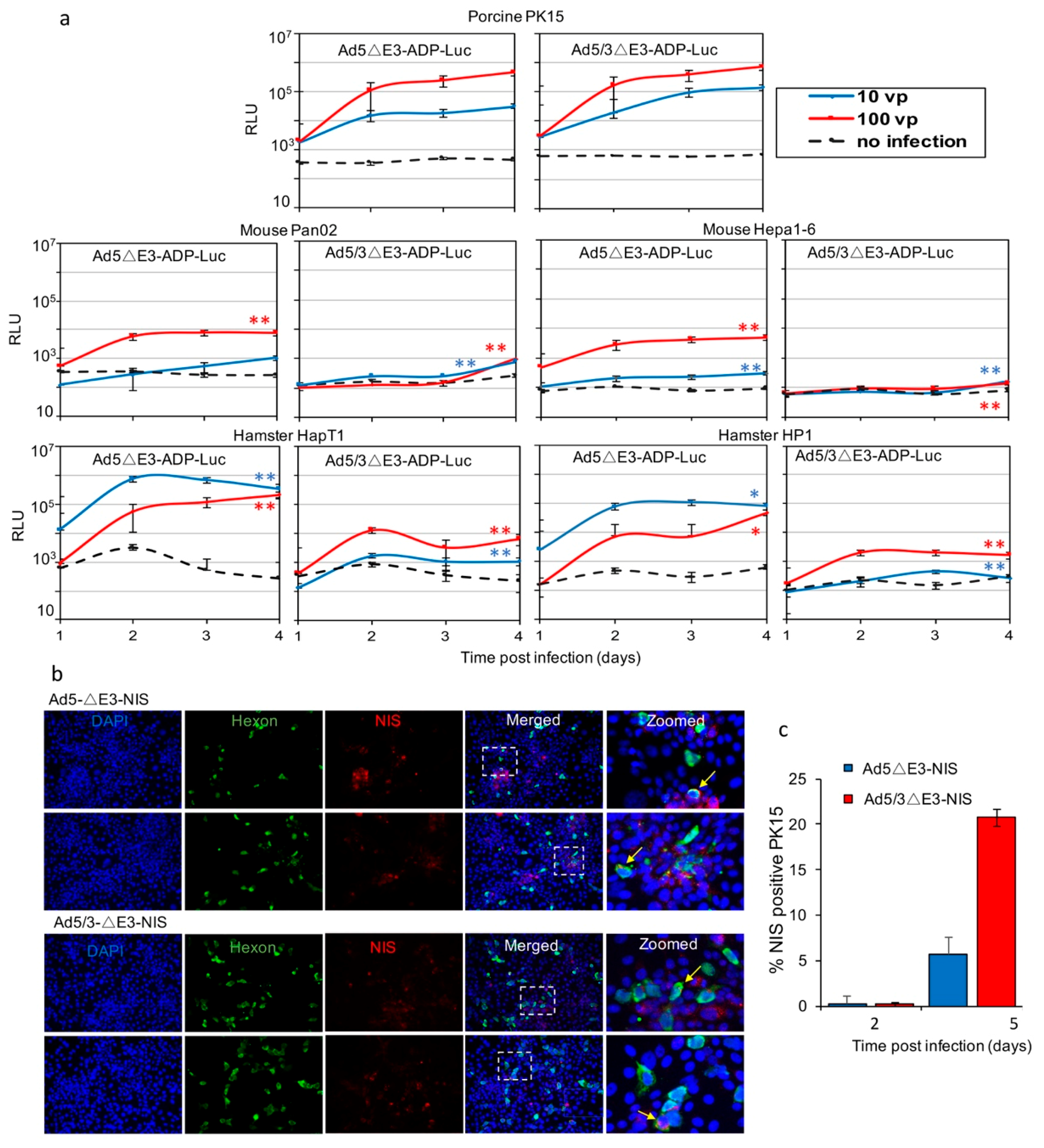
2.4. Analysis of Replication-Dependent Gene Expression
2.5. In Vivo Analysis of Ad5 and Ad5/3 Replication in Pigs
2.6. Analysis of Systemic Toxicity in Pigs after Systemic Administration of Adenoviral Vectors
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Adenoviral Vector Construction
4.3. In Vitro Analysis of Ad5 and Ad5/3 Binding Ability
4.4. In Vitro Analysis of Ad5 and Ad5/3 Gene Transduction
4.5. In Vitro Analysis of Ad5 and Ad5/3 Replication Ability or Cytocidal Effect
4.6. Cell Viability Assay
4.7. In Vitro Analysis of Ad Replication Dependent Gene Expression in Porcine Cells
4.8. In Vivo Analysis of Adenovirus Replication in Pigs
4.9. Statistical Analysis
4.10. Ethical Approval
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Russell, W.C. Adenoviruses: Update on structure and function. J. Gen. Virol. 2009, 90, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell 1993, 73, 309–319. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Roelvink, P.W.; Lizonova, A.; Lee, J.G.; Li, Y.; Bergelson, J.M.; Finberg, R.W.; Brough, D.E.; Kovesdi, I.; Wickham, T.J. The coxsackievirus-adenovirus receptor protein can function as a cellular attachment protein for adenovirus serotypes from subgroups A, C, D, E, and F. J. Virol. 1998, 72, 7909–7915. [Google Scholar] [PubMed]
- Arnberg, N. Adenovirus receptors: Implications for targeting of viral vectors. Trends Pharmacol. Sci. 2012, 33, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Gaggar, A.; Shayakhmetov, D.M.; Lieber, A. CD46 is a cellular receptor for group B adenoviruses. Nat. Med. 2003, 9, 1408–1412. [Google Scholar] [CrossRef] [PubMed]
- Marttila, M.; Persson, D.; Gustafsson, D.; Liszewski, M.K.; Atkinson, J.P.; Wadell, G.; Arnberg, N. CD46 is a cellular receptor for all species B adenoviruses except types 3 and 7. J. Virol. 2005, 79, 14429–14436. [Google Scholar] [CrossRef] [PubMed]
- Sirena, D.; Lilienfeld, B.; Eisenhut, M.; Kalin, S.; Boucke, K.; Beerli, R.R.; Vogt, L.; Ruedl, C.; Bachmann, M.F.; Greber, U.F.; et al. The Human Membrane Cofactor CD46 Is a Receptor for Species B Adenovirus Serotype 3. J. Virol. 2004, 78, 4454–4462. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Z.Y.; Liu, Y.; Persson, J.; Beyer, I.; Moller, T.; Koyuncu, D.; Drescher, M.R.; Strauss, R.; Zhang, X.B.; et al. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat. Med. 2011, 17, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Tuve, S.; Wang, H.J.; Ware, C.; Liu, Y.; Gaggar, A.; Bernt, K.; Shayakhmetov, D.; Li, Z.Y.; Strauss, R.; Stone, D.; et al. A new group B adenovirus receptor is expressed at high levels on human stem and tumor cells. J. Virol. 2006, 80, 12109–12120. [Google Scholar] [CrossRef]
- Short, J.J.; Vasu, C.; Holterman, M.J.; Curiel, D.T.; Pereboev, A. Members of adenovirus species B utilize CD80 and CD86 as cellular attachment receptors. Virus Res. 2006, 122, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Bauerschmitz, G.J.; Barker, S.D.; Hemminki, A. Adenoviral gene therapy for cancer: From vectors to targeted and replication competent agents (Review). Int. J. Oncol. 2002, 21, 1161–1174. [Google Scholar] [CrossRef] [PubMed]
- Krasnykh, V.N.; Mikheeva, G.V.; Douglas, J.T.; Curiel, D.T. Generation of recombinant adenovirus vectors with modified fibers for altering viral tropism. J. Virol. 1996, 70, 6839–6846. [Google Scholar] [PubMed]
- Davydova, J.; Le, L.P.; Gavrikova, T.; Wang, M.; Krasnykh, V.; Yamamoto, M. Infectivity-enhanced cyclooxygenase-2-based conditionally replicative adenoviruses for esophageal adenocarcinoma treatment. Cancer Res. 2004, 64, 4319–4327. [Google Scholar] [CrossRef] [PubMed]
- Guse, K.; Ranki, T.; Ala-Opas, M.; Bono, P.; Sarkioja, M.; Rajecki, M.; Kanerva, A.; Hakkarainen, T.; Hemminki, A. Treatment of metastatic renal cancer with capsid-modified oncolytic adenoviruses. Mol. Cancer Ther. 2007, 6, 2728–2736. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, A.; Mikheeva, G.V.; Krasnykh, V.; Coolidge, C.J.; Lam, J.T.; Mahasreshti, P.J.; Barker, S.D.; Straughn, M.; Barnes, M.N.; Alvarez, R.D.; et al. Targeting adenovirus to the serotype 3 receptor increases gene transfer efficiency to ovarian cancer cells. Clin. Cancer Res. 2002, 8, 275–280. [Google Scholar] [PubMed]
- Kangasniemi, L.; Kiviluoto, T.; Kanerva, A.; Raki, M.; Ranki, T.; Sarkioja, M.; Wu, H.J.; Marini, F.; Hockerstedt, K.; Isoniemi, H.; et al. Infectivity-enhanced Adenoviruses deliver efficacy in clinical samples and orthotopic models of disseminated gastric cancer. Clin. Cancer Res. 2006, 12, 3137–3144. [Google Scholar] [CrossRef]
- Rivera, A.A.; Davydova, J.; Schierer, S.; Wang, M.; Krasnykh, V.; Yamamoto, M.; Curiel, D.T.; Nettelbeck, D.M. Combining high selectivity of replication with fiber chimerism for effective adenoviral oncolysis of CAR-negative melanoma cells. Gene Ther. 2004, 11, 1694–1702. [Google Scholar] [CrossRef]
- Volk, A.L.; Rivera, A.A.; Kanerva, A.; Bauerschmitz, G.; Dmitriev, I.; Nettelbeck, D.M.; Curiel, D.T. Enhanced adenovirus infection of melanoma cells by fiber-modification—Incorporation of RGD peptide or Ad5/3 chimerism. Cancer Biol. Ther. 2003, 2, 511–515. [Google Scholar] [CrossRef]
- Zheng, S.; Ulasov, I.V.; Han, Y.; Tyler, M.A.; Zhu, Z.B.; Lesniak, M.S. Fiber-knob modifications enhance adenoviral tropism and gene transfer in malignant glioma. J. Gene Med. 2007, 9, 151–160. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H.; Yumul, R.; Gao, W.; Gambotto, A.; Morita, T.; Baker, A.; Shayakhmetov, D.; Lieber, A. Transduction of liver metastases after intravenous injection of Ad5/35 or Ad35 vectors with and without factor X-binding protein pretreatment. Hum. Gene Ther. 2009, 20, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonero, R.; Salazar, R.; Duran, I.; Osman-Garcia, I.; Paz-Ares, L.; Bozada, J.M.; Boni, V.; Blanc, C.; Seymour, L.; Beadle, J.; et al. Phase 1 study of intravenous administration of the chimeric adenovirus enadenotucirev in patients undergoing primary tumor resection. J. Immunother. Cancer 2017, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Jogler, C.; Hoffmann, D.; Theegarten, D.; Grunwald, T.; Uberla, K.; Wildner, O. Replication properties of human adenovirus in vivo and in cultures of primary cells from different animal species. J. Virol. 2006, 80, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Ternovoi, V.V.; Le, L.P.; Belousova, N.; Smith, B.F.; Siegal, G.P.; Curiel, D.T. Productive replication of human adenovirus type 5 in canine cells. J. Virol. 2005, 79, 1308–1311. [Google Scholar] [CrossRef] [PubMed]
- Toth, K.; Spencer, J.F.; Tollefson, A.E.; Kuppuswamy, M.; Doronin, K.; Lichtenstein, D.L.; La Regina, M.C.; Prince, G.A.; Wold, W.S. Cotton rat tumor model for the evaluation of oncolytic adenoviruses. Hum. Gene Ther. 2005, 16, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Wold, W.S.; Toth, K. Chapter three—Syrian hamster as an animal model to study oncolytic adenoviruses and to evaluate the efficacy of antiviral compounds. Adv. Cancer Res. 2012, 115, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Ying, B.; Toth, K.; Spencer, J.F.; Meyer, J.; Tollefson, A.E.; Patra, D.; Dhar, D.; Shashkova, E.V.; Kuppuswamy, M.; Doronin, K.; et al. INGN 007, an oncolytic adenovirus vector, replicates in Syrian hamsters but not mice: Comparison of biodistribution studies. Cancer Gene Ther. 2009, 16, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Blair, G.E.; Dixon, S.C.; Griffiths, S.A.; Blair Zajdel, M.E. Restricted replication of human adenovirus type 5 in mouse cell lines. Virus Res. 1989, 14, 339–346. [Google Scholar] [CrossRef]
- Sonabend, A.M.; Ulasov, I.V.; Han, Y.; Rolle, C.E.; Nandi, S.; Cao, D.; Tyler, M.A.; Lesniak, M.S. Biodistribution of an oncolytic adenovirus after intracranial injection in permissive animals: A comparative study of Syrian hamsters and cotton rats. Cancer Gene Ther. 2009, 16, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.C.; Morrison, B.J.; Mannan, P.; Abu-Asab, M.S.; Wildner, O.; Miles, B.K.; Yim, K.C.; Ramanan, V.; Prince, G.A.; Morris, J.C. Immunocompetent syngeneic cotton rat tumor models for the assessment of replication-competent oncolytic adenovirus. Virology 2007, 369, 131–142. [Google Scholar] [CrossRef] [PubMed]
- LaRocca, C.J.; Han, J.; Gavrikova, T.; Armstrong, L.; Oliveira, A.R.; Shanley, R.; Vickers, S.M.; Yamamoto, M.; Davydova, J. Oncolytic adenovirus expressing interferon alpha in a syngeneic Syrian hamster model for the treatment of pancreatic cancer. Surgery 2015, 157, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Salzwedel, A.O.; Han, J.; LaRocca, C.J.; Shanley, R.; Yamamoto, M.; Davydova, J. Combination of interferon-expressing oncolytic adenovirus with chemotherapy and radiation is highly synergistic in hamster model of pancreatic cancer. Oncotarget 2018, 9, 18041–18052. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.A.; Spencer, J.F.; Wold, W.S. Use of the Syrian hamster as an animal model for oncolytic adenovirus vectors. Methods Mol. Med. 2007, 130, 169–183. [Google Scholar] [PubMed]
- Dhar, D.; Spencer, J.F.; Toth, K.; Wold, W.S. Pre-existing immunity and passive immunity to adenovirus 5 prevents toxicity caused by an oncolytic adenovirus vector in the Syrian hamster model. Mol. Ther. 2009, 17, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, D.L.; Spencer, J.F.; Doronin, K.; Patra, D.; Meyer, J.M.; Shashkova, E.V.; Kuppuswamy, M.; Dhar, D.; Thomas, M.A.; Tollefson, A.E.; et al. An acute toxicology study with INGN 007, an oncolytic adenovirus vector, in mice and permissive Syrian hamsters; comparisons with wild-type Ad5 and a replication-defective adenovirus vector. Cancer Gene Ther. 2009, 16, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.F.; Sagartz, J.E.; Wold, W.S.; Toth, K. New pancreatic carcinoma model for studying oncolytic adenoviruses in the permissive Syrian hamster. Cancer Gene Ther. 2009, 16, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Ryan, M.J.; Estep, J.E.; Miniard, B.M.; Rudge, T.L.; Peggins, J.O.; Broadt, T.L.; Wang, M.; Preuss, M.A.; Siegal, G.P.; et al. A new generation of serotype chimeric infectivity-enhanced conditionally replicative adenovirals: The safety profile of ad5/3-Delta24 in advance of a phase I clinical trial in ovarian cancer patients. Hum. Gene Ther. 2011, 22, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Siurala, M.; Bramante, S.; Vassilev, L.; Hirvinen, M.; Parviainen, S.; Tahtinen, S.; Guse, K.; Cerullo, V.; Kanerva, A.; Kipar, A.; et al. Oncolytic adenovirus and doxorubicin-based chemotherapy results in synergistic antitumor activity against soft-tissue sarcoma. Int. J. Cancer 2015, 136, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Bosch, P.; Fouletier-Dilling, C.; Olmsted-Davis, E.A.; Davis, A.R.; Stice, S.L. Efficient adenoviral-mediated gene delivery into porcine mesenchymal stem cells. Mol. Reprod. Dev. 2006, 73, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Griesche, N.; Zikos, D.; Witkowski, P.; Nitsche, A.; Ellerbrok, H.; Spiller, O.B.; Pauli, G.; Biere, B. Growth characteristics of human adenoviruses on porcine cell lines. Virology 2008, 373, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Lam, E.; Ramke, M.; Groos, S.; Warnecke, G.; Heim, A. A differentiated porcine bronchial epithelial cell culture model for studying human adenovirus tropism and virulence. J. Virol. Methods 2011, 178, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, R.E.; Horvath, C.; Snyder, E.A.; Patrick, J.; Collins, N.; Evans, E.; MacDonald, J.S. Porcine toxicology studies of SCH 58500, an adenoviral vector for the p53 gene. Toxicol. Sci. 2002, 65, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Muhlhauser, J.; Jones, M.; Yamada, I.; Cirielli, C.; Lemarchand, P.; Gloe, T.R.; Bewig, B.; Signoretti, S.; Crystal, R.G.; Capogrossi, M.C. Safety and efficacy of in vivo gene transfer into the porcine heart with replication-deficient, recombinant adenovirus vectors. Gene Ther. 1996, 3, 145–153. [Google Scholar] [PubMed]
- Torres, J.M.; Alonso, C.; Ortega, A.; Mittal, S.; Graham, F.; Enjuanes, L. Tropism of human adenovirus type 5-based vectors in swine and their ability to protect against transmissible gastroenteritis coronavirus. J. Virol. 1996, 70, 3770–3780. [Google Scholar] [PubMed]
- Wesley, R.D.; Tang, M.; Lager, K.M. Protection of weaned pigs by vaccination with human adenovirus 5 recombinant viruses expressing the hemagglutinin and the nucleoprotein of H3N2 swine influenza virus. Vaccine 2004, 22, 3427–3434. [Google Scholar] [CrossRef] [PubMed]
- Davydova, J.; Gavrikova, T.; Brown, E.J.; Luo, X.H.; Curiel, D.T.; Vickers, S.M.; Yamamoto, M. In vivo bioimaging tracks conditionally replicative adenoviral replication and provides an early indication of viral antitumor efficacy. Cancer Sci. 2010, 101, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Bramante, S.; Kaufmann, J.K.; Veckman, V.; Liikanen, I.; Nettelbeck, D.M.; Hemminki, O.; Vassilev, L.; Cerullo, V.; Oksanen, M.; Heiskanen, R.; et al. Treatment of melanoma with a serotype 5/3 chimeric oncolytic adenovirus coding for GM-CSF: Results in vitro, in rodents and in humans. Int. J. Cancer 2015, 137, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Nakanishi, T.; Matsumoto, M.; Nomura, M.; Seya, T.; Okabe, M. Disruption of Mouse CD46 Causes an Accelerated Spontaneous Acrosome Reaction in Sperm. Mol. Cell. Biol. 2003, 23, 2614–2622. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Harris, C.L.; Johnson, P.M.; Morgan, B.P. Rat membrane cofactor protein (MCP; CD46) is expressed only in the acrosome of developing and mature spermatozoa and mediates binding to immobilized activated C3. Biol. Reprod. 2004, 71, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Seymour, L.W.; Fisher, K.D. Oncolytic viruses: Finally delivering. Br. J. Cancer 2016, 114, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.C.; Sarver, A.L.; Gavin, K.J.; Thayanithy, V.; Getzy, D.M.; Newman, R.A.; Cutter, G.R.; Lindblad-Toh, K.; Kisseberth, W.C.; Hunter, L.E.; et al. Molecular subtypes of osteosarcoma identified by reducing tumor heterogeneity through an interspecies comparative approach. Bone 2011, 49, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Ritt, M.G.; Wojcieszyn, J.; Modiano, J.F. Functional loss of p21/Waf-1 in a case of benign canine multicentric melanoma. Vet. Pathol. 1998, 35, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Davydova, J.; Yamamoto, M. Oncolytic adenoviruses: Design, generation, and experimental procedures. Curr. Protoc. Hum. Genet. 2013. [Google Scholar] [CrossRef]
- Nettelbeck, D.M.; Rivera, A.A.; Davydova, J.; Dieckmann, D.; Yamamoto, M.; Curiel, D.T. Cyclooxygenase-2 promoter for tumour-specific targeting of adenoviral vectors to melanoma. Melanoma Res. 2003, 13, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, M.A.; Oneal, M.J.; Davydova, J.; Bergert, E.; Yamamoto, M.; Morris, J.C., 3rd. Construction of an MUC-1 promoter driven, conditionally replicating adenovirus that expresses the sodium iodide symporter for gene therapy of breast cancer. BCR 2009, 11, R53. [Google Scholar] [CrossRef] [PubMed]
- Oneal, M.J.; Trujillo, M.A.; Davydova, J.; McDonough, S.; Yamamoto, M.; Morris, J.C. Effect of increased viral replication and infectivity enhancement on radioiodide uptake and oncolytic activity of adenovirus vectors expressing the sodium iodide symporter. Cancer Gene Ther. 2013, 20, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ventrella, D.; Dondi, F.; Barone, F.; Serafini, F.; Elmi, A.; Giunti, M.; Romagnoli, N.; Forni, M.; Bacci, M.L. The biomedical piglet: Establishing reference intervals for haematology and clinical chemistry parameters of two age groups with and without iron supplementation. BMC Vet. Res. 2017, 13, 23. [Google Scholar] [CrossRef]
- Rahman, M.M. Normal Hematological and Biochemical References of Swine Species. Available online: https://www.researchgate.net/publication/307859127_Normal_hematological_and_biochemical_references_of_Swine_species (accessed on 13 December 2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum Chemistry | Control/PBS (d.p.i) | Ad5ΔE3-Luc (d.p.i) | Ad5/3ΔE3-Luc (d.p.i) | Reference | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Measurement | 0 | 2 | 4 | 7 | 0 | 2 | 4 | 7 | 0 | 2 | 4 | 7 | Range |
| Glucose (mg/dL) | 136 | 104 | 99 | 97 | 108 | 150 | 88 | 94 | 111 | 120 | 92 | 90 | 85–150 |
| SDH (U/L) | 35.3 | 4.8 | 3.4 | 22.3 | 12.6 | 10.4 | 8.7 | 82.6 | 24.4 | 8.6 | 18.8 | 12.6 | 0–45 |
| Total Bilirubin (mg/dL) | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0–1 |
| Cholesterol (mg/dL) | 91 | 71 | 78 | 88 | 72 | 83 | 89 | 80 | 102 | 106 | 110 | 94 | 36–54 |
| Total protein (g/dL) | 5.3 | 5.6 | 5.5 | 5.6 | 4.5 | 5.1 | 5.3 | 5.6 | 5.5 | 5.1 | 5.6 | 5.6 | 7.9–8.9 |
| Albumin (g/dL) | 3.5 | 3.7 | 3.9 | 4 | 3.3 | 3.7 | 3.7 | 4 | 3.5 | 3.3 | 3.6 | 3.7 | 1.9–3.3 |
| Urea N (mg/dL) | 10 | 14 | 9 | 9 | 7 | 11 | 7 | 9 | 6 | 12 | 8 | 8 | 10–30 |
| Creatinine (mg/dL) | 0.9 | 0.9 | 0.7 | 0.8 | 0.6 | 0.6 | 0.5 | 0.7 | 0.8 | 0.8 | 0.6 | 0.6 | 1.0–2.7 |
| Phosphorous (mg/dL) | 9.1 | 8 | 7.3 | 6.9 | 9 | 8.6 | 7.9 | 9.1 | 8.9 | 8.4 | 8.1 | 8.5 | 5.3–9.6 |
| Calcium (mg/dL) | 10.2 | 11.1 | 10.5 | 10.7 | 10.4 | 10.8 | 10.4 | 11.2 | 10.7 | 10.4 | 10.9 | 10.6 | 7.1–11.6 |
| Sodium (mmol/L) | 142 | 145 | 144 | 142 | 142 | 145 | 143 | 142 | 142 | 145 | 143 | 141 | 135–150 |
| Potassium (mmol/L) | 3.8 | 4 | 4.3 | 3.8 | 3.9 | 4.9 | 4 | 4 | 3.7 | 4.5 | 4.1 | 4 | 4.4–6.7 |
| Chloride (mmol/L) | 104 | 109 | 107 | 102 | 105 | 102 | 103 | 97 | 102 | 105 | 103 | 98 | 94–106 |
| Bicarbonate (mmol/L) | 29 | 27 | 31 | 32 | 32 | 33 | 32 | 25 | 31 | 32 | 31 | 36 | 18–27 |
| CK (U/L) | 466 | 1082 | 1567 | 2103 | 1035 | 732 | 1247 | 1078 | 513 | 531 | 2761 | 1049 | 61–1251 |
| Gamma-GT (U/L) | 42 | 38 | 40 | 43 | 40 | 33 | 33 | 39 | 30 | 27 | 35 | 35 | 10–60 |
| Globulin (g/dL) | 1.8 | 1.9 | 1.6 | 1.6 | 1.2 | 1.4 | 1.6 | 1.6 | 2 | 1.8 | 2 | 1.9 | 5.3–6.4 |
| A/G Ratio | 1.9 | 1.9 | 2.4 | 2.5 | 2.8 | 2.6 | 2.3 | 2.5 | 1.8 | 1.8 | 1.8 | 1.9 | 0.37–0.51 |
| Urea/Creat ratio | 11.1 | 15.6 | 12.9 | 11.3 | 11.7 | 18.3 | 14 | 12.9 | 7.5 | 15 | 13.3 | 13.3 | N/A |
| ALP (VALKP) (U/L) | 221 | 205 | 195 | 187 | 259 | 190 | 164 | 203 | 250 | 167 | 190 | 197 | 118–395 |
| ALT (GPT) (U/L) | 59 | 61 | 70 | 75 | 68 | 61 | 64 | 77 | 72 | 60 | 74 | 81 | 31–58 |
| AST (GOT) (U/L) | 37 | 47 | 65 | 56 | 61 | 38 | 54 | 56 | 43 | 33 | 83 | 51 | 32–84 |
| LDH (VLDH) (U/L) | 706 | 654 | 640 | 683 | 654 | 615 | 728 | 756 | 693 | 501 | 782 | 536 | 380–634 |
| Triglycerides (VTRIG) | 37 | 45 | 37 | 65 | 29 | 53 | 55 | 76 | 49 | 40 | 35 | 57 | N/A |
| Blood Parameters | Control/PBS (d.p.i) | Ad5ΔE3-Luc (d.p.i) | Ad5/3ΔE3-Luc (d.p.i) | Reference | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Measurement | 0 | 2 | 4 | 7 | 0 | 2 | 4 | 7 | 0 | 2 | 4 | 7 | Range |
| Red Blood Cells (×106/µL) | 5.5 | 5.8 | 5.3 | 5.1 | 5.4 | 6.3 | 5.4 | 5.9 | 6.3 | 6.2 | 5.9 | 5.7 | 5–8 |
| Hemoglobin (g/dL) | 10.0 | 10.4 | 9.7 | 9.1 | 9.8 | 11.5 | 9.7 | 10.9 | 11.0 | 11.0 | 10.4 | 10.1 | 10–16 |
| Hematocrit (%) | 32.7 | 32.5 | 29.7 | 28.7 | 31.8 | 34.8 | 31.0 | 35.2 | 36.8 | 33.0 | 32.1 | 30.6 | 32–50 |
| Platelets (×103/µL) | 357 | 386 | 300 | 412 | 131 | 234 | 400 | 293 | 260 | 222 | 361 | 377 | 325–715 |
| White Blood Cells (×103/µL) | 17.5 | 20.9 | 15.0 | 15.5 | 11.2 | 14.0 | 10.7 | 11.0 | 14.0 | 20.6 | 14.5 | 13.2 | 11–22 |
| Neutrophils (×103/µL) | 6.7 | 11.9 | 7.4 | 7.0 | 4.7 | 6.2 | 4.6 | 3.6 | 5.9 | 14.4 | 5.1 | 5.7 | 0.3–15.2 |
| Lymphocytes (×103/µL) | 10.7 | 7.3 | 6.3 | 7.4 | 5.5 | 5.6 | 4.8 | 6.2 | 7.4 | 4.3 | 8.4 | 6.5 | 3.6–18.5 |
| Monocytes (×103/µL) | 0.2 | 1.7 | 0.3 | 0.5 | 0.8 | 2.0 | 1.1 | 0.7 | 0.3 | 1.2 | 0.2 | 0.4 | 0.0–4.9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koodie, L.; Robertson, M.G.; Chandrashekar, M.; Ruth, G.; Dunning, M.; Bianco, R.W.; Davydova, J. Rodents Versus Pig Model for Assessing the Performance of Serotype Chimeric Ad5/3 Oncolytic Adenoviruses. Cancers 2019, 11, 198. https://doi.org/10.3390/cancers11020198
Koodie L, Robertson MG, Chandrashekar M, Ruth G, Dunning M, Bianco RW, Davydova J. Rodents Versus Pig Model for Assessing the Performance of Serotype Chimeric Ad5/3 Oncolytic Adenoviruses. Cancers. 2019; 11(2):198. https://doi.org/10.3390/cancers11020198
Chicago/Turabian StyleKoodie, Lisa, Matthew G. Robertson, Malavika Chandrashekar, George Ruth, Michele Dunning, Richard W. Bianco, and Julia Davydova. 2019. "Rodents Versus Pig Model for Assessing the Performance of Serotype Chimeric Ad5/3 Oncolytic Adenoviruses" Cancers 11, no. 2: 198. https://doi.org/10.3390/cancers11020198
APA StyleKoodie, L., Robertson, M. G., Chandrashekar, M., Ruth, G., Dunning, M., Bianco, R. W., & Davydova, J. (2019). Rodents Versus Pig Model for Assessing the Performance of Serotype Chimeric Ad5/3 Oncolytic Adenoviruses. Cancers, 11(2), 198. https://doi.org/10.3390/cancers11020198