NKG2D Polymorphism in Melanoma Patients from Southeastern Spain

 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Results
2.1. Clinical and Histological Characteristics of Melanoma Patients
2.2. Frequency of SNPs in the NKC Region
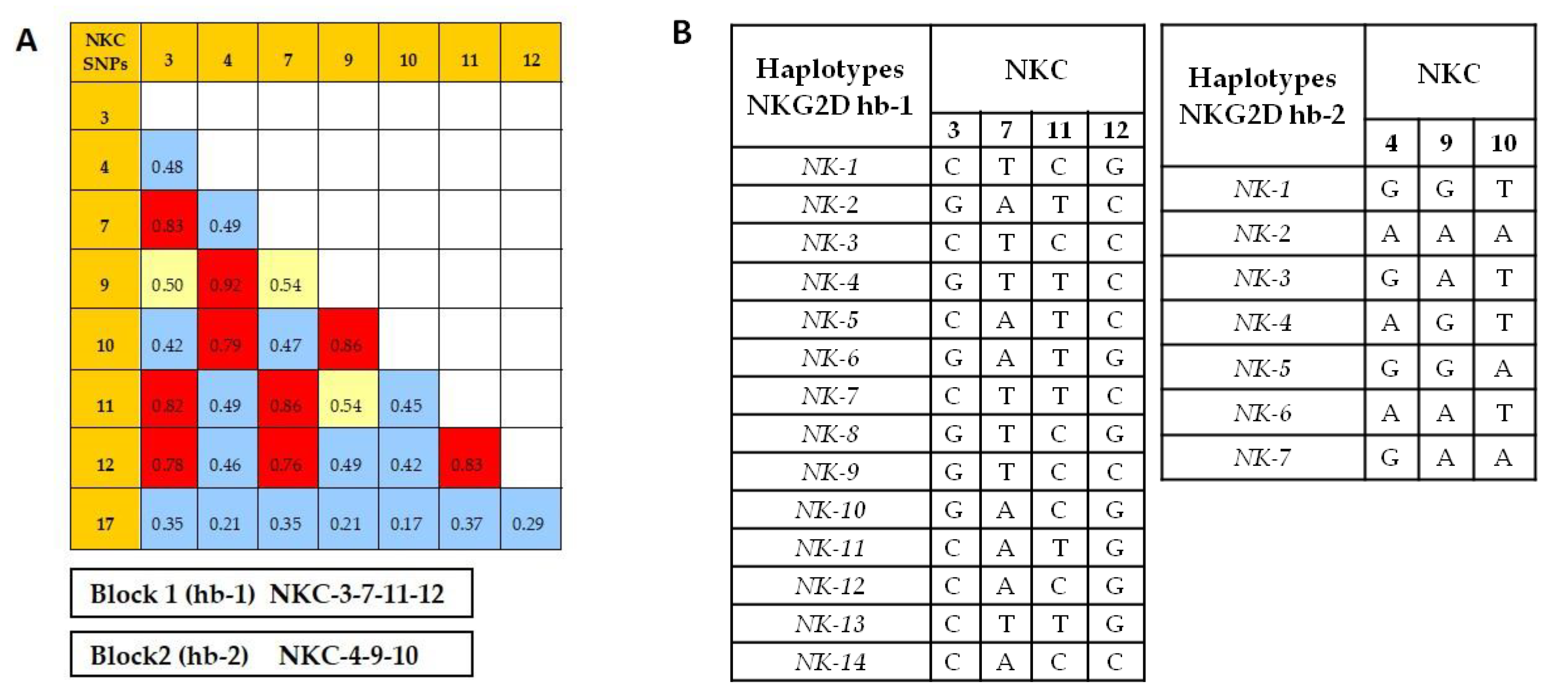
2.3. Identification of NKC Haplotypes
2.4. NKC Haplotype Frequencies
2.5. NKC Haplotypes and Melanoma Risk
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Sample Collection and DNA preparation
4.3. NKC Region Genotyping
4.4. NKC Region Haplotype Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, Risk Factors, Pathogenesis, Diagnosis and Classification. In Vivo 2014, 28, 1005–1012. [Google Scholar] [PubMed]
- Rigel, D.S. Epidemiology of melanoma. Semin. Cutan. Med. Surg. 2010, 29, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Mohammadpour, A.; Derakhshan, M.; Darabi, H.; Hedayat, P.; Momeni, M. Melanoma: Where we are and where we go. J. Cell. Physiol. 2018, 234, 3307–3320. [Google Scholar] [CrossRef] [PubMed]
- Anichini, A.; Vegetti, C.; Mortarini, R. The paradox of T cell-mediated antitumor immunity in spite of poor clinical outcome in human melanoma. Cancer Immunol. Immunother. 2004, 53, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.P.; Yee, C.; Savage, P.A.; Fong, L.; Brockstedt, D.; Weber, J.S.; Johnson, D.; Swetter, S.; Thompson, J.; Greenberg, P.D.; et al. Characterization of circulating T cells specific for tumor-associated antigens in melanoma patients. Nat. Med. 1999, 5, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Groh, V.; Wu, J.; Yee, C.; Spies, T. Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature 2002, 419, 734–738. [Google Scholar] [CrossRef]
- Konjević, G.; MirjacićMartinović, K.; Vuletić, A.; Jović, V.; Jurisić, V.; Babović, N.; Spuzić, I. Low expression of CD161 and NKG2D activating NK receptor is associated with impaired NK cell cytotoxicity in metastatic melanoma patients. Clin. Exp. Metastasis 2007, 24, 1–11. [Google Scholar] [CrossRef]
- Martinovic, K.; MKonjević, G.; Babović, N.; Inić, M. The stage dependent changes in NK cell activity and the expression of activating and inhibitory NK cell receptors in melanoma patients. J. Surg. Res. 2011, 171, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Casado, J.G.; Pawelec, G.; Morgado, S.; Sanchez-Correa, B.; Delgado, E.; Gayoso, I.; Duran, E.; Solana, R.; Tarazona, R. Expression of adhesion molecules and ligands for activating and costimulatory receptors involved in cell-mediated cytotoxicity in a large panel of human melanoma cell lines. Cancer Immunol. Immunother. 2009, 58, 1517–1526. [Google Scholar] [CrossRef]
- Morgado, S.; Sanchez-Correa, B.; Casado, J.G.; Duran, E.; Gayoso, I.; Labella, F.; Solana, R.; Tarazona, R. NK cell recognition and killing of melanoma cells is controlled by multiple activating receptor-ligand interactions. J. Innate Immun. 2011, 3, 365–373. [Google Scholar] [CrossRef]
- Solana, R.; CasadoJ, G.; Delgado, E.; De la Rosa, O.; Marin, J.; Duran, E.; Pawelec, G.; Tarazona, R. Lymphocyte activation in response to melanoma: Interaction of NK-associated receptors and their ligands. Cancer Immunol. Immunother. 2007, 56, 101–110. [Google Scholar] [CrossRef]
- Pende, D.; Rivera, P.; Marcenaro, S.; Chang, C.C.; Biassoni, R.; Conte, R.; Kubin, M.; Cosman, D.; Ferrone, S.; Moretta, L.; et al. Major histocompatibility complex class I-related chain A and UL16-binding protein expression on tumor cell lines of different histotypes: Analysis of tumor susceptibility to NKG2D-dependent natural killer cell cytotoxicity. Cancer Res. 2002, 62, 6178–6186. [Google Scholar] [PubMed]
- Moretta, L.; Bottino, C.; Pende, D.; Castriconi, R.; Mingari, M.C.; Moretta, A. Surface NK receptors and their ligands on tumor cells. Semin. Immunol. 2006, 18, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Seliger, B.; Abken, H.; Ferrone, S. HLA-G and MIC expression in tumors and their role in anti-tumor immunity. Trends Immunol. 2003, 24, 82–87. [Google Scholar] [CrossRef]
- Vetter, C.S.; Groh, V.; Straten, P.T.; Spies, T.; Bröcker, E.B.; Becker, J.C. Expression of stress-induced MHC class I related chain molecules on human melanoma. J. Investig. Dermatol. 2002, 118, 600–605. [Google Scholar] [CrossRef]
- Carrega, P.; Pezzino, G.; Queirolo, P.; Bonaccorsi, I.; Falco, M.; Vita, G.; Pende, D.; Misefari, A.; Moretta, A.; Mingari, M.C.; et al. Susceptibility of human melanoma cells to autologous natural killer (NK) cell killing: HLA-related effector mechanisms and role of unlicensed NK cells. PLoS ONE 2009, 4, e8132. [Google Scholar] [CrossRef]
- Pende, D.; Cantoni, C.; Rivera, P.; Castriconi, M.R.; Marcenaro, S.; Nanni, M.; Biassoni, R.; Bottino, C.; Moretta, A.; Moretta, L. Role of NKG2D in tumor cell lysis mediated by human NK cells: Cooperation with natural cytotoxicity receptors and capability of recognizing tumors of nonepithelial origin. Eur. J. Immunol. 2001, 31, 1076–1086. [Google Scholar] [CrossRef]
- Cosman, D.; Müllberg, J.; Sutherland, C.L.; Chin, W.; Armitage, R.; Fanslow, W.; Kubin, M.; Chalupny, N.J. ULBPs, novel MHC class I-related molecules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity through the NKG2D receptor. Immunity 2001, 14, 123–133. [Google Scholar] [CrossRef]
- Sheppard, S.; Ferry, A.; Guedes, J.; Guerra, N. The paradoxical role of NKG2D in Cancer Immunity. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Raulet, D.H. Roles of the NKG2D immunoreceptor and its ligands. Nat. Rev. Immunol. 2003, 3, 781–790. [Google Scholar] [CrossRef]
- Bauer, S.; Groh, V.; Wu, J.; Steinle, A.; Phillips, J.H.; Lanier, L.L.; Spies, T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 1999, 285, 727–729. [Google Scholar] [CrossRef]
- Wu, J.; Song, Y.; Bakker, A.B.; Bauer, S.; Spies, T.; Lanier, L.L.; Phillips, J.H. An activating immunoreceptor complex formed by NKG2D and DAP10. Science 1999, 285, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Cerwenka, A.; Lanier, L.L. NKG2D ligands: Unconventional MHC class I-like molecules exploited by viruses and cancer. Tissue Antigens 2003, 61, 335–343. [Google Scholar] [CrossRef]
- Kubin, M.; Cassiano, L.; Chalupny, J.; Chin, W.; Cosman, D.; Fanslow, W.; Müllberg, J.; Rousseau, A.M.; Ulrich, D.; Armitage, R. ULBP1, 2, 3: Novel MHC class I-related molecules that bind to human cytomegalovirus glycoprotein UL16, activate NK cells. Eur. J. Immunol. 2001, 31, 1428–1437. [Google Scholar] [CrossRef]
- Groh, V.; Bahram, S.; Bauer, S.; Herman, A.; Beauchamp, M.; Spies, T. Cell stress-regulated human major histocompatibility complex class I gene expressed in gastrointestinal epithelium. Proc. Natl. Acad. Sci. USA 1996, 93, 12445–12450. [Google Scholar] [CrossRef] [PubMed]
- Groh, V.; Steinle, A.; Bauer, S.; Spies, T. Recognition of stress-induced MHC molecules by intestinal epitelialgy T cells. Science 1998, 279, 1737–1740. [Google Scholar] [CrossRef]
- Guillamón, C.F.; Martínez-Sánchez, M.V.; Gimeno, L.; Mrowiec, A.; Martínez-García, J.; Server-Pastor, G.; Martínez-Escribano, J.; Torroba, A.; Ferri, B.; Abellán, D.; et al. NK Cell Education in Tumor Immune Surveillance: DNAM-1/KIR Receptor Ratios as Predictive Biomarkers for Solid Tumor Outcome. Cancer Immunol. Res. 2018, 6, 1537–1547. [Google Scholar] [CrossRef]
- Hayashi, T.; Morishita, Y.; Hayashi, I.; Kusunoki, Y.; Nakachi, K. Identification of the NKG2D haplotypes associated with natural cytotoxic activity of peripheral blood lymphocytes and cancer immunosurveillance. Cancer Res. 2006, 66, 563–570. [Google Scholar] [CrossRef]
- Ma, J.; Guo, X.; Wu, X.; Li, J.; Zhu, X.; Li, Z.; Li, J.; Pan, L.; Li, T.; Li, H.; et al. Association of NKG2D genetic polymorphism with susceptibility to chronic hepatitis B in a Han Chinese population. J. Med. Virol. 2010, 82, 1501–1507. [Google Scholar] [CrossRef]
- Piotrowski, P.; Lianeri, M.; Olesinska, M.; Jagodzinski, P.P. Prevalence of the NKG2D Thr72Ala Polymorphism in patients with systemic lupus erythematosus. Mol. Biol. Rep. 2012, 39, 1343–1347. [Google Scholar] [CrossRef]
- Roszak, A.; Lianeri, M.; Jagodziński, P.P. Prevalence of the NKG2D Thr72Ala polymorphism in patients with cervical carcinoma. Genet. Test. Mol. Biomark. 2012, 16, 841–845. [Google Scholar] [CrossRef]
- Lanier, L.L. NKG2D Receptor and Its Ligands in Host Defense. Cancer Immunol. Res. 2015, 3, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, S.; Lopez-Soto, A.; Suarez-Alvarez, B.; Lopez-Vazquez, A.; Lopez-Larrea, C. NKG2D ligands: Key targets of the immune response. Trends Immunol. 2008, 29, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Kabalak, G.; Thomas, R.M.; Martin, J.; Ortego-Centeno, N.; Jimenez-Alonso, J.; de Ramón, E.; Buyny, S.; Hamsen, S.; Gross, W.L.; Schnarr, S.; et al. Association of an NKG2D gene variant with systemic lupus erythematosus in two populations. Hum. Immunol. 2010, 71, 74–78. [Google Scholar] [CrossRef]
- Ghobadzadeh, S.; Shams, A.; Eslami, G.; Mirghanizadeh, A. Investigation of NKG2D rs1049174G>C Gene Polymorphism in Women with Breast Cancer. J. Shahid Sadoughi Univ. Med. Sci. 2013, 21, 291–299. [Google Scholar]
- Melum, E.; Karlsen, T.H.; Schrumpf, E.; Bergquist, A.; Thorsby, E.; Boberg, K.M.; Lie, B.A. Cholangiocarcinoma in primary sclerosing cholangitis is associated with NKG2D polymorphisms. Hepatology 2008, 47, 90–96. [Google Scholar] [CrossRef]
- Chaiteerakij, R.; Juran, B.D.; Moser, C.D.; Mettler, T.A.; Lazaridis, K.; Roberts, L.R. Lack of Association between NKG2D Polymorphisms and Cholangiocarcinoma (CCA) Risk in Primary Sclerosing Cholangitis (PSC) and Non-PSC Patients. Gastroenterology 2012, 142, S-922. [Google Scholar] [CrossRef]
- Espinoza, J.L.; Takami, A.; Onizuka, M.; Sao, H.; Akiyama, H.; Miyamura, K.; Okamoto, S.; Inoue, M.; Kanda, Y.; Ohtake, S.; et al. Japan Marrow Donor Program. NKG2D gene polymorphism has a significant impact on transplant outcomes after HLA-fully-matched unrelated bone marrow transplantation for standard risk hematologic malignancies. Haematologica 2009, 94, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Furue, H.; Matsuo, K.; Kumimoto, H.; Hiraki, A.; Suzuki, T.; Yatabe, Y.; Komori, K.; Kanemitsu, Y.; Hirai, T.; Kato, T.; et al. Decreased risk of colorectal cancer with the high Natural-Killer (NK)-cell activity NKG2D genotype in Japanese. Carcinogenesis 2008, 29, 316–320. [Google Scholar] [CrossRef]
- Furue, H.; Kumimoto, H.; Matsuo, K.; Suzuki, T.; Hasegawa, Y.; Shinoda, M.; Sugimura, T.; Mitsudo, K.; Tohnai, I.; Ueda, M.; et al. Opposite impact of NKG2D genotype by lifestyle exposure to risk of aerodigestive tract cancer among Japanese. Int. J. Cancer 2008, 123, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Balch, C.M.; Gershenwald, J.E.; Soong, S.J.; Thompson, J.F.; Atkins, M.B.; Byrd, D.R.; Buzaid, A.C.; Cochran, A.J.; Coit, D.G.; Ding, S.; et al. Final version of 2009 AJCC melanoma staging and classification. J. Clin. Oncol. 2009, 27, 6199–6206. [Google Scholar] [CrossRef]
- Gershenwald, J.E.; Scolyer, R.A.; Hess, K.R.; Sondak, V.K.; Long, G.V.; Ross, M.I.; Lazar, A.J.; Faries, M.B.; Kirkwood, J.M.; McArthur, G.A.; et al. Melanoma staging: Evidence-based changes in the American Joint Committee on Cancer eighth edition cancer staging manual. CA Cancer J. Clin. 2017, 67, 472–492. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite v. 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Svejgaard, A.; Ryder, L.P. HLA and disease associations: Detecting the strongest association. Tissue Antigens 1994, 43, 18–27. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Demographic Data | Controls 1 (N = 200) | Melanoma Patients (N = 233) |
|---|---|---|
| Age (years, Mean ± SD) | 51 ± 17 | 53 ± 15 |
| Gender (Male) | 92 (46%) | 112 (48%) |
| Histological subtypes | ||
| SSM | 162 (69%) | |
| NM | 29 (12%) | |
| LMM | 16 (7%) | |
| ALM | 13 (6%) | |
| Other 2 | 13 (6%) | |
| Clinical characteristics | ||
| Tumor thickness | ||
| ≤1 mm | 128 (55%) | |
| >1 mm | 104 (45%) | |
| Ulceration at primary lesion | ||
| Yes | 44 (20%) | |
| No | 173 (80%) | |
| SLN metastasis | ||
| Yes | 22 (16%) | |
| No | 119 (84%) | |
| NK CRegion | Genotype | Controls (N = 200) | Melanoma Patients (N = 233) | ||
|---|---|---|---|---|---|
| Total | SSM | NM | |||
| NKC3 | CC | 83 (42%) | 114 (49%) | 81 (50%) | 13 (45%) |
| CG | 86 (43%) | 94 (40%) | 62 (38%) | 14 (48%) | |
| GG | 31 (15%) | 25 (11%) | 19 (12%) | 2 (7%) | |
| C-allele | 169 (85%) | 208 (89%) | 143 (88%) | 27(93%) | |
| NKC4 | GG | 111 (56%) | 138 (59%) | 97 (60%) | 15 (52%) |
| AG | 72 (36%) | 85 (37%) | 58 (36%) | 13 (45%) | |
| AA | 17 (8%) | 10 (4%) | 7 (4%) | 1 (3%) | |
| G-allele | 183(92%) | 223 (96%) | 155 (96%) | 28 (97%) | |
| NKC7 | AA | 28 (14%) | 22 (9%) | 17 (11%) | 2 (7%) |
| AT | 92 (46%) | 94 (40%) | 62 (38%) | 14 (48%) | |
| TT | 80 (40%) | 117 (51%) 1 | 83 (51%) 2 | 13 (45%) | |
| A-allele | 120 (60%) | 116 (49%) 3 | 79 (49%)4 | 16 (55%) | |
| NKC9 | AA | 16 (8%) | 27 (12%) | 21 (13%) | 3 (10%) |
| AG | 70 (35%) | 58 (25%) 5 | 38 (23%) 6 | 9 (31%) | |
| GG | 114 (57%) | 148 (63%) | 103 (64%) | 17 (59%) | |
| G-allele | 184 (92%) | 206 (88%) | 141 (87%) | 26 (90%) | |
| NKC10 | AA | 14 (7%) | 9 (4%) | 7 (4%) | 1 (3%) |
| AT | 75 (38%) | 76 (33%) | 53 (33%) | 10 (35%) | |
| TT | 111 (55%) | 148 (63%) | 102 (63%) | 18 (62%) | |
| T-allele | 186 (93%) | 155 (96%) | 155 (96%) | 28 (97%) | |
| NKC11 | CC | 80 (40%) | 109 (47%) | 79 (49%) | 12 (41%) |
| CT | 91 (46%) | 99 (42%) | 63 (39%) | 15 (52%) | |
| TT | 29 (14%) | 25 (11%) | 20 (12%) | 2 (7%) | |
| T-allele | 120 (60%) | 124 (53%) | 83 (51%) | 17 (59%) | |
| NKC12 | CC | 32 (16%) | 35 (15%) | 26 (16%) | 3 (10%) |
| CG | 87 (44%) | 100 (43%) | 64 (40%) | 17 (59%) | |
| GG | 81 (40%) | 98 (42%) | 72 (44%) | 9 (31%) | |
| C-allele | 119 (60%) | 135 (58%) | 90 (56%) | 20 (69%) | |
| NKC17 | CC | 35 (17%) | 43 (18%) | 32 (20%) | 7 (24%) |
| CG | 89 (45%) | 112 (48%) | 73 (45%) | 14 (48%) | |
| GG | 76 (38%) | 78 (34%) | 57 (35%) | 8 (28%) | |
| C-allele | 124 (62%) | 155 (66%) | 105 (65%) | 21 (72%) | |
| NKC Region | Genotype | Control N = 200 | Melanoma Patients (N = 233) | |||||
|---|---|---|---|---|---|---|---|---|
| Tumor Thickness | Ulceration | LN Metastasis | ||||||
| ≤1.0 mm N = 128 | >1.0 mm N = 104 | No N = 173 | Yes N = 44 | No N = 119 | Yes N = 22 | |||
| NKC3 | CC | 83 (42%) | 63 (49%) | 51 (49%) | 84 (49%) | 20 (46%) | 58 (49%) | 11 (50%) |
| CG | 86 (43%) | 50 (39%) | 43 (41%) | 70 (41%) | 19 (43%) | 50 (42%) | 6 (27%) | |
| GG | 31 (15%) | 15 (12%) | 10 (10%) | 19 (11%) | 5 (11%) | 11 (9%) | 5 (23%) | |
| C-allele | 169 (85%) | 113 (88%) | 94 (90%) | 154 (90%) | 39 (89%) | 108 (91%) | 17 (77%) | |
| NKC4 | CC | 111 (56%) | 74 (58%) | 63 (61%) | 103 (60%) | 22(50%) | 73 (61%) | 12 (55%) |
| CT | 72 (36%) | 47 (37%) | 38 (37%) | 61(35%) | 21 (48%) | 43 (36%) | 7 (32%) | |
| TT | 17 (8%) | 7 (6%) | 3 (3%) | 9(5%) | 1 (2%) | 3 (2%) | 3 (14%) | |
| C-allele | 183 (92%) | 121 (95%) | 101 (98%) | 164 (95%) | 43 (98%) | 116 (97%) | 19 (87%) 1 | |
| NKC7 | AA | 28 (14%) | 14 (11%) | 8 (8%) | 16 (9%) | 5 (11%) | 9 (8%) | 4 (18%) |
| AT | 92 (46%) | 48 (38%) | 45 (43%) | 68 (39%) | 21 (48%) | 50 (42%) | 7 (32%) | |
| TT | 80 (40%) | 66 (52%) | 51 (49%) | 89 (51%) | 18 (41%) | 60 (50%) | 11 (50%) | |
| A-allele | 120 (60%) | 62 (49%) | 53 (51%) | 84 (48%) | 26 (59%) | 59 (50%) | 11 (50%) | |
| NKC9 | AA | 16 (8%) | 19 (15%) | 8 (8%) | 24 (14%) | 1 (2%) 2 | 19 (16%) | 5 (23%) |
| AG | 70 (35%) | 30 (23%) | 28 (27%) | 41 (24%) | 16 (36%) | 22 (19%) | 5 (23%) 3 | |
| GG | 114 (57%) | 79 (62%) | 68 (65%) | 108 (62%) | 27 (61%) | 78 (65%) | 12 (54%) | |
| G-allele | 184 (92%) | 109 (85%) | 96 (92%) | 149 (86%) | 43(97%) 4 | 100(84%) | 17(77%) 5 | |
| NKC10 | AA | 14 (7%) | 5 (5%) | 3 (3%) | 8 (5%) | 1 (2%) | 3 (2%) | 3 (14%) 6 |
| AT | 75 (38%) | 44 (34%) | 32 (31%) | 58 (33%) | 15 (34%) | 39 (33%) | 7 (32%) | |
| TT | 111 (55%) | 78 (61%) | 69 (66%) | 107 (62%) | 28 (64%) | 77 (65%) | 12 (55%) | |
| T-allele | 186 (93%) | 122 (95%) | 101 (97%) | 165 (95%) | 43 (98%) | 116 (98%) | 19 (87%) 7 | |
| NKC11 | CC | 80 (40%) | 62 (48%) | 47 (45%) | 85 (49%) | 14 (32%) 8 | 54 (45%) | 10 (46%) |
| CT | 91 (46%) | 51 (40%) | 47 (45%) | 69 (40%) | 25 (57%) | 55 (46%) | 7 (32%) | |
| TT | 29 (14%) | 15 (12%) | 10 (10%) | 19 (11%) | 5 (11%) | 10 (8%) | 5 (23%) | |
| T-allele | 120 (60%) | 66 (52%) | 57 (55%) | 88 (51%) | 30 (68%) 9 | 65 (54%) | 12 (55%) | |
| NKC12 | CC | 32 (16%) | 21 (16%) | 14 (13%) | 26 (15%) | 7 (16%) | 14 (12%) | 6 (27%) |
| CG | 87 (44%) | 50 (39%) | 49 (47%) | 70 (40%) | 25 (57%) | 55 (46%) | 7 (32%) | |
| GG | 81 (40%) | 57 (45%) | 41 (39%) | 77 (45%) | 12 (27%) 10 | 50 (42%) | 9 (41%) | |
| C-allele | 119 (60%) | 71 (55%) | 63 (60%) | 96 (55%) | 32 (73%) 11 | 69 (58%) | 13 (59%) | |
| NKC17 | CC | 35 (17%) | 23 (18%) | 20 (19%) | 31 (18%) | 9 (21%) | 23 (19%) | 5 (23%) |
| CG | 89 (45%) | 64 (50%) | 47 (45%) | 87 (50%) | 16 (36%) | 55 (46%) | 8 (36%) | |
| GG | 76 (38%) | 41 (32%) | 37 (36%) | 55 (32%) | 19 (43%) | 41 (35%) | 9 (41%) | |
| C-allele | 124 (62%) | 87 (68%) | 67 (64%) | 118 (68%) | 25 (57%) | 78 (65%) | 13 (59%) | |
| NKG2D Hb-1 Haplotypes | Controls | Melanoma Patients | ||
|---|---|---|---|---|
| Total | p/Pc | (X2) | ||
| NK-1 | 0.587 | 0.619 | 0.33 | (0.94) |
| NK-2 | 0.329 | 0.276 | 0.09 | (2.87) |
| NK-3 | 0.015 1 | 0.048 | 0.007/0.098 | (7.26) |
| NK-4 | 0.010 | 0.011 | 0.93 | (0.008) |
| NK-5 | 0.010 | 0.011 | 0.92 | (0.009) |
| NK-6 | 0.010 | 0.009 | 0.82 | (0.052) |
| NK-7 | 0.005 | 0.013 | 0.23 | (1.45) |
| NK-8 | 0.005 | 0.007 | 0.74 | (0.11) |
| NK-9 | 0.005 | 0.006 | 0.84 | (0.04) |
| NK-10 | 0.010 2 | 0.000 | 0.03/0.42 | (4.56) |
| NK-11 | 0.005 | 0.000 | 0.14 | (2.19) |
| NK-12 | 0.003 | 0.000 | 0.27 | (1.19) |
| NK-13 | 0.003 | 0.000 | 0.31 | (1.04) |
| NK-14 | 0.003 | 0.000 | 0.27 | (1.21) |
| NKG2D Hb-2 Haplotype | Controls | Melanoma Patients | ||
|---|---|---|---|---|
| Total | p/Pc | (X2) | ||
| NK-1 | 0.717 | 0.732 | 0.63 | (0.23) |
| NK-2 | 0.240 | 0.197 | 0.13 | (2.30) |
| NK-3 | 0.000 | 0.039 | 0.00009/0.0006 | (15.25) |
| NK-4 | 0.013 | 0.024 | 0.24 | (1.35) |
| NK-5 | 0.015 | 0.004 | 0.10 | (2.69) |
| NK-6 | 0.012 | 0.004 | 0.19 | (1.67) |
| NK-7 | 0.003 | 0.000 | 0.31 | (1.04) |
| NKG2D Hb-2 Haplotype | Melanoma Patients | |||||||
|---|---|---|---|---|---|---|---|---|
| SSM | NM | Tumor Thickness | Ulceration | SLN Metastasis | ||||
| <1 mm | >1 mm | No | Yes | No | Yes | |||
| NK-3 | 0.040 | 0.034 | 0.056 | 0.024 | 0.046 | 0.000 1 | 0.067 | 0.045 |
| NKG2D Haplotypes and Genotypes | Controls (N = 200) | Melanoma Patients (N = 233) | ||
|---|---|---|---|---|
| Total (N = 191) | SSM (N = 162) | NM (N = 29) | ||
| Hb-1 | ||||
| NK-1 | 163 (81%) | 197 (84%) | 135 (83%) | 26 (90%) |
| NK-2 | 109 (54%) | 110 (47%) | 74 (46%) | 16 (55%) |
| NK-1/NK-1 | 73 (37%) | 92 (40%) | 68 (42%) | 9 (31%) |
| NK-1/NK-2 | 80 (40%) | 84 (36%) | 55 (34%) | 14 (48%) |
| NK-2/NK-2 | 24 (12%) | 20 (9%) | 15 (9%) | 2 (7%) |
| Hb-2 | ||||
| NK-1 | 182 (91%) | 204 (88%) | 139 (86%) | 26 (90%) |
| NK-2 | 84 (42%) | 84 (36%) | 59 (36%) | 11 (38%) |
| NK-1/NK-1 | 106 (53%) | 137 (59%) | 96 (59%) | 15 (52%) |
| NK-1/NK-2 | 65 (33%) | 56 (24%) | 37 (23%) 1 | 8 (28%) |
| NKC | SNP ID | AB TaqMan Assay | Nucleotide Variation * |
|---|---|---|---|
| 3 | rs1049174 | C_9345347_10 | [G/C] |
| 4 | rs2255336 | C_22274476_10 | [A/G] |
| 7 | rs2617160 | C_1841959_10 | [A/T] |
| 9 | rs2246809 | C_1842497_10 | [A/G] |
| 10 | rs2617169 | CUSTOM | [T/A] |
| 11 | rs2617170 | C_1842316_10 | [T/C] |
| 12 | rs2617171 | C_26984346_10 | [C/G] |
| 17 | rs1983526 | C_11919464_10 | [C/G] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gimeno, L.; Martínez-Banaclocha, H.; Bernardo, M.V.; Bolarin, J.M.; Marín, L.; López-Hernández, R.; López-Alvarez, M.R.; Moya-Quiles, M.R.; Muro, M.; Frias-Iniesta, J.F.; et al. NKG2D Polymorphism in Melanoma Patients from Southeastern Spain. Cancers 2019, 11, 438. https://doi.org/10.3390/cancers11040438
Gimeno L, Martínez-Banaclocha H, Bernardo MV, Bolarin JM, Marín L, López-Hernández R, López-Alvarez MR, Moya-Quiles MR, Muro M, Frias-Iniesta JF, et al. NKG2D Polymorphism in Melanoma Patients from Southeastern Spain. Cancers. 2019; 11(4):438. https://doi.org/10.3390/cancers11040438
Chicago/Turabian StyleGimeno, Lourdes, Helios Martínez-Banaclocha, M. Victoria Bernardo, José Miguel Bolarin, Luis Marín, Ruth López-Hernández, M. Rocío López-Alvarez, M. Rosa Moya-Quiles, Manuel Muro, José Francisco Frias-Iniesta, and et al. 2019. "NKG2D Polymorphism in Melanoma Patients from Southeastern Spain" Cancers 11, no. 4: 438. https://doi.org/10.3390/cancers11040438