Independent Negative Prognostic Role of TCF1 Expression within the Wnt/β-Catenin Signaling Pathway in Primary Breast Cancer Patients

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
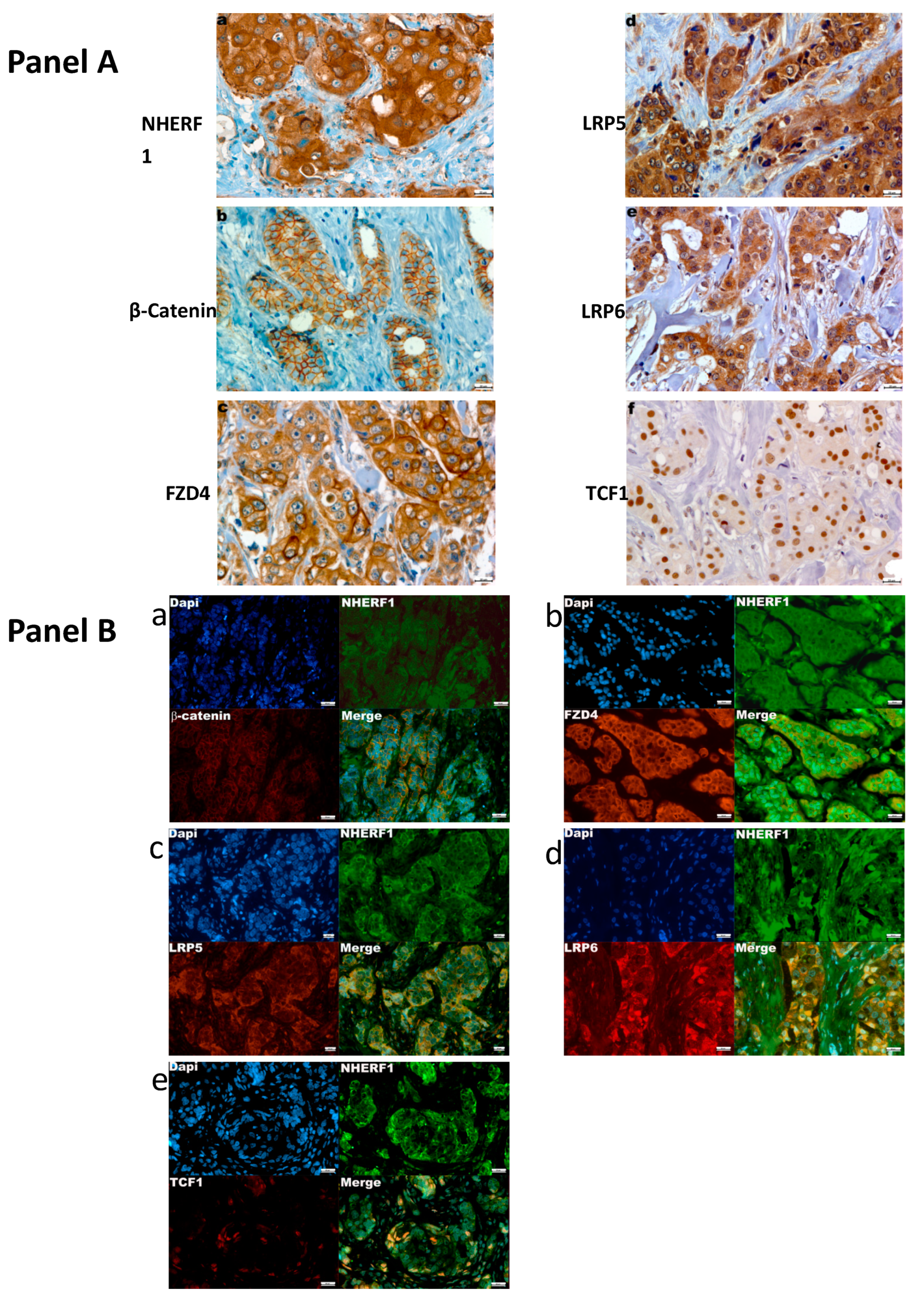
2.1. Profiling of Expression and Relationship between Tumor Markers and Clinicopathological Features
2.2. Analysis of Association between NHERF1 and β-catenin, FZD4, LRP5, LRP6, TCF1 Protein Expression
2.3. Immunofluorescence Co-Localization
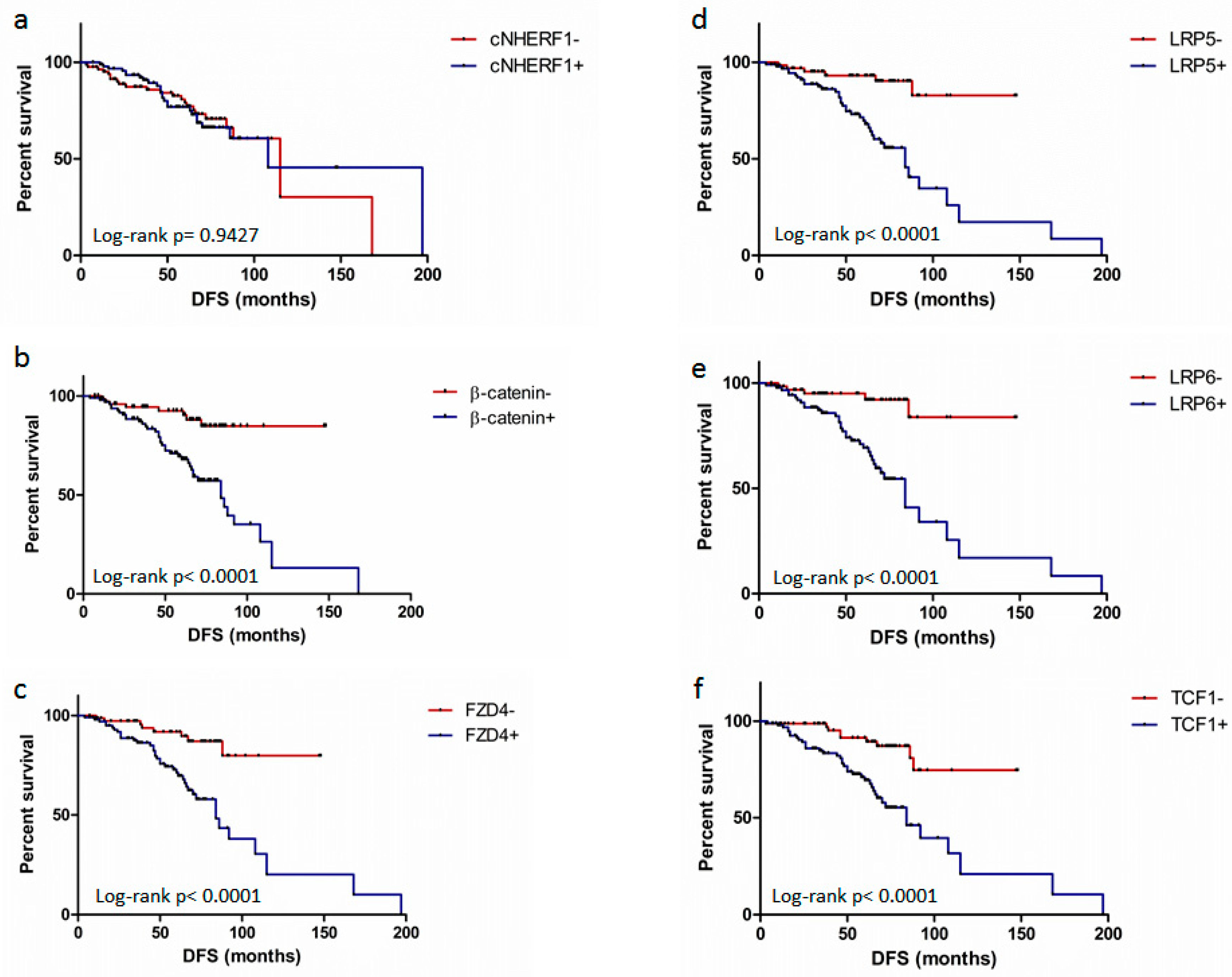
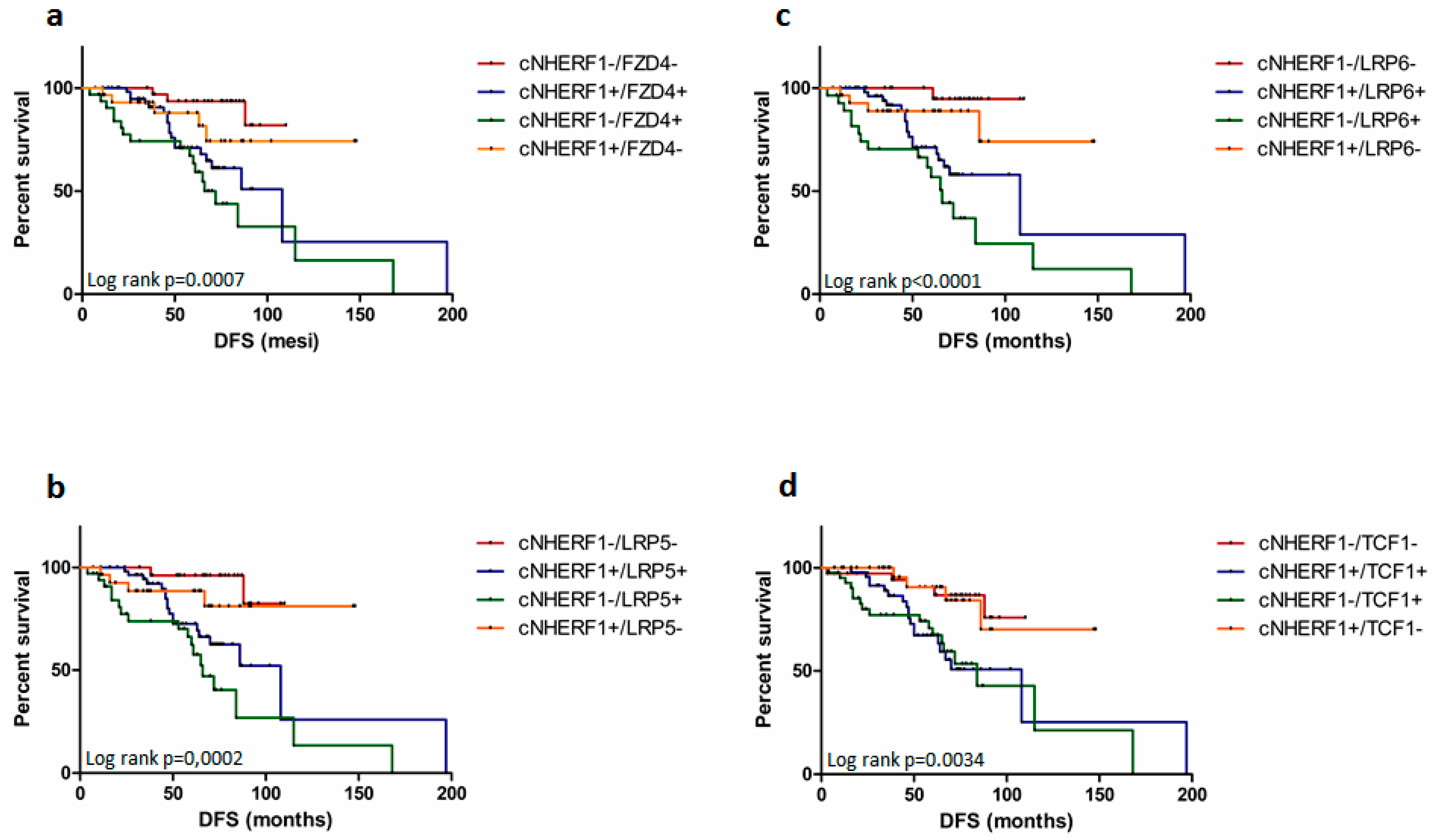
2.4. Expression of Proteins and Patient Clinical Outcome
3. Discussion
4. Materials and Methods
4.1. Patients and Clinicopathological Characteristics
4.2. Tissue Microarray and Immunohistochemistry.
4.3. Immunohistochemical Assessment
4.4. Immunofluorescence
4.5. Follow-Up and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Donowitz, M.; Cha, B.; Zachos, N.C.; Brett, C.L.; Sharma, A.; Tse, C.M.; Li, X. NHERF family and NHE3 regulation. J. Physiol. 2005, 567, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Centonze, M.; Saponaro, C.; Mangia, A. NHERF1 Between Promises and Hopes: Overview on Cancer and Prospective Openings. Transl. Oncol. 2018, 11, 374–390. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, C.; Sergio, S.; Coluccia, A.; De Luca, M.; La Regina, G.; Mologni, L.; Famiglini, V.; Naccarato, V.; Bonetti, D.; Gautier, C.; et al. β-catenin knockdown promotes NHERF1-mediated survival of colorectal cancer cells: Implications for a double-targeted therapy. Oncogene 2018, 37, 3301–3316. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Qin, Q.; Song, R.; Zhao, C.; Liu, H.; Yang, Y.; Gu, S.; Zhou, D.; He, J. NHERF1 inhibits beta-catenin-mediated proliferation of cervical cancer cells through suppression of alpha-actinin-4 expression. Cell Death Dis. 2018, 9, 668. [Google Scholar] [CrossRef] [PubMed]
- Mangia, A.; Caldarola, L.; Dell’Endice, S.; Scarpi, E.; Saragoni, L.; Monti, M.; Santini, D.; Brunetti, O.; Simone, G.; Silvestris, N. The potential predictive role of nuclear NHERF1 expression in advanced gastric cancer patients treated with epirubicin/oxaliplatin/capecitabine first line chemotherapy. Cancer Biol. Ther. 2015, 16, 1140–1147. [Google Scholar] [CrossRef]
- Malfettone, A.; Silvestris, N.; Saponaro, C.; Ranieri, G.; Russo, A.; Caruso, S.; Popescu, O.; Simone, G.; Paradiso, A.; Mangia, A. High density of tryptase-positive mast cells in human colorectal cancer: A poor prognostic factor related to protease-activated receptor 2 expression. J. Cell. Mol. Med. 2013, 17, 1025–1037. [Google Scholar] [CrossRef]
- Malfettone, A.; Silvestris, N.; Paradiso, A.; Mattioli, E.; Simone, G.; Mangia, A. Overexpression of nuclear NHERF1 in advanced colorectal cancer: Association with hypoxic microenvironment and tumor invasive phenotype. Exp. Mol. Pathol. 2012, 92, 296–303. [Google Scholar] [CrossRef]
- Mangia, A.; Partipilo, G.; Schirosi, L.; Saponaro, C.; Galetta, D.; Catino, A.; Scattone, A.; Simone, G. Fine Needle Aspiration Cytology: A Tool to Study NHERF1 Expression as a Potential Marker of Aggressiveness in Lung Cancer. Mol. Biotechnol. 2015, 57, 549–557. [Google Scholar] [CrossRef]
- Greco, M.R.; Bon, E.; Rubino, R.; Guerra, L.; Bernabe-Garcia, M.; Cannone, S.; Cayuela, M.L.; Ciaccia, L.; Marionneau-Lambot, S.; Oullier, T.; et al. Phosphorylation of NHERF1 S279 and S301 differentially regulates breast cancer cell phenotype and metastatic organotropism. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 26–37. [Google Scholar] [CrossRef]
- Saponaro, C.; Vagheggini, A.; Scarpi, E.; Centonze, M.; Catacchio, I.; Popescu, O.; Pastena, M.I.; Giotta, F.; Silvestris, N.; Mangia, A. NHERF1 and tumor microenvironment: A new scene in invasive breast carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 96. [Google Scholar] [CrossRef] [PubMed]
- Mangia, A.; Scarpi, E.; Partipilo, G.; Schirosi, L.; Opinto, G.; Giotta, F.; Simone, G. NHERF1 together with PARP1 and BRCA1 expression as a new potential biomarker to stratify breast cancer patients. Oncotarget 2017, 8, 65730–65742. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, A.; Scarpi, E.; Malfettone, A.; Addati, T.; Giotta, F.; Simone, G.; Amadori, D.; Mangia, A. Nuclear NHERF1 expression as a prognostic marker in breast cancer. Cell Death Dis. 2013, 4, e904. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.S.; Barrick, S.R.; Grubisha, M.J.; Brufsky, A.M.; Friedman, P.A.; Romero, G. Direct interaction between NHERF1 and Frizzled regulates β-catenin signaling. Oncogene 2011, 30, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Schirosi, L.; Mazzotta, A.; Opinto, G.; Pinto, R.; Graziano, G.; Tommasi, S.; Fucci, L.; Simone, G.; Mangia, A. β-catenin interaction with NHERF1 and RASSF1A methylation in metastatic colorectal cancer patients. Oncotarget 2016, 7, 67841–67850. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y. Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Front. Immunol. 2018, 9, 745. [Google Scholar] [CrossRef]
- Jönsson, M.; Borg, A.; Nilbert, M.; Andersson, T. Involvement of adenomatous polyposis coli (APC)/beta-catenin signalling in human breast cancer. Eur. J. Cancer 2000, 36, 242–248. [Google Scholar] [CrossRef]
- Georgescu, M.M.; Gagea, M.; Cote, G. NHERF1/EBP50 Suppresses Wnt-β-Catenin Pathway-Driven Intestinal Neoplasia. Neoplasia 2016, 18, 512–523. [Google Scholar] [CrossRef]
- MacDonald, B.T.; He, X. Frizzled and LRP5/6 receptors for Wnt/β-catenin signaling. Cold Spring Harb. Perspect. Biol. 2012, 4, a007880. [Google Scholar] [CrossRef]
- May, P.; Woldt, E.; Matz, R.L.; Boucher, P. The LDL receptor-related protein (LRP) family: An old family of proteins with new physiological functions. Ann. Med. 2007, 39, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Wang, W.; Zhang, Z.; Bai, Y.; Gao, J.; Zhao, C. Wnt signaling in human and mouse breast cancer: Focusing on Wnt ligands, receptors and antagonists. Cancer Sci. 2018, 109, 3368–3375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.P.; Li, X.W.; Lang, J.H. Prognostic Value of β-catenin Expression in Breast Cancer Patients: A Meta-analysis. Asian Pac. J. Cancer Prev. 2015, 16, 5625–5633. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kreimann, E.L.; Morales, F.C.; de Orbeta-Cruz, J.; Takahashi, Y.; Adams, H.; Liu, T.J.; McCrea, P.D.; Georgescu, M.M. Cortical stabilization of beta-catenin contributes to NHERF1/EBP50 tumor suppressor function. Oncogene 2007, 26, 5290–5299. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, M.M.; Morales, F.C.; Molina, J.R.; Hayashi, Y. Roles of NHERF1/EBP50 in cancer. Curr. Mol. Med. 2008, 8, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Mangia, A.; Chiriatti, A.; Bellizzi, A.; Malfettone, A.; Stea, B.; Zito, F.A.; Reshkin, S.J.; Simone, G.; Paradiso, A. Biological role of NHERF1 protein expression in breast cancer. Histopathology 2009, 55, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Cowin, P.; Rowlands, T.M.; Hatsell, S.J. Cadherins and catenins in breast cancer. Curr. Opin. Cell Biol. 2005, 17, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Dubois, L.; Lecourtois, M.; Alexandre, C.; Hirst, E.; Vincent, J.P. Regulated endocytic routing modulates wingless signaling in Drosophila embryos. Cell 2001, 105, 613–624. [Google Scholar] [CrossRef]
- Maubant, S.; Tahtouh, T.; Brisson, A.; Maire, V.; Némati, F.; Tesson, B.; Ye, M.; Rigaill, G.; Noizet, M.; Dumont, A.; et al. LRP5 regulates the expression of STK40, a new potential target in triple-negative breast cancers. Oncotarget 2018, 9, 22586–22604. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, C.C.; Prior, J.; Piwnica-Worms, D.; Bu, G. LRP6 overexpression defines a class of breast cancer subtype and is a target for therapy. Proc. Natl. Acad. Sci. USA 2010, 107, 5136–5141. [Google Scholar] [CrossRef]
- Karn, T.; Ruckhäberle, E.; Hanker, L.; Müller, V.; Schmidt, M.; Solbach, C.; Gätje, R.; Gehrmann, M.; Holtrich, U.; Kaufmann, M.; et al. Gene expression profiling of luminal B breast cancers reveals NHERF1 as a new marker of endocrine resistance. Breast Cancer Res. Treat. 2011, 130, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Cardone, R.A.; Bellizzi, A.; Busco, G.; Weinman, E.J.; Dell’Aquila, M.E.; Casavola, V.; Azzariti, A.; Mangia, A.; Paradiso, A.; Reshkin, S.J. The NHERF1 PDZ2 domain regulates PKA-RhoA-p38-mediated NHE1 activation and invasion in breast tumor cells. Mol. Biol. Cell 2007, 18, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- El-Tanani, M.; Fernig, D.G.; Barraclough, R.; Green, C.; Rudland, P. Differential modulation of transcriptional activity of estrogen receptors by direct protein-protein interactions with the T cell factor family of transcription factors. J. Biol. Chem. 2001, 276, 41675–41682. [Google Scholar] [CrossRef]
- López-Knowles, E.; Zardawi, S.J.; McNeil, C.M.; Millar, E.K.; Crea, P.; Musgrove, E.A.; Sutherland, R.L.; O’Toole, S.A. Cytoplasmic localization of beta-catenin is a marker of poor outcome in breast cancer patients. Cancer Epidemiol. Biomark. Prev. 2010, 19, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.W.; Liu, Y.F.; Yu, J.C.; Wang, H.W.; Ding, S.L.; Hsiung, C.N.; Hsu, H.M.; Shieh, J.C.; Wu, P.E.; Shen, C.Y. Prognostic significance of cyclin D1, β-catenin, and MTA1 in patients with invasive ductal carcinoma of the breast. Ann. Surg. Oncol. 2012, 19, 4129–4139. [Google Scholar] [CrossRef] [PubMed]
- Tortelote, G.G.; Reis, R.R.; de Almeida Mendes, F.; Abreu, J.G. Complexity of the Wnt/β-catenin pathway: Searching for an activation model. Cell. Signal. 2017, 40, 30–43. [Google Scholar] [CrossRef]
- Mangia, A.; Malfettone, A.; Saponaro, C.; Tommasi, S.; Simone, G.; Paradiso, A. Human epidermal growth factor receptor 2, Na+/H+ exchanger regulatory factor 1, and breast cancer susceptibility gene-1 as new biomarkers for familial breast cancers. Hum. Pathol. 2011, 42, 1589–1595. [Google Scholar] [CrossRef]
- Wolff, A.C.; Hammond, M.E.; Schwartz, J.N.; Hagerty, K.L.; Allred, D.C.; Cote, R.J.; Dowsett, M.; Fitzgibbons, P.L.; Hanna, W.M.; Langer, A.; et al. American Society of Clinical Oncology; College of American Pathologists. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for human epidermal growth factor receptor 2 testing in breast cancer. J. Clin. Oncol. 2007, 25, 118–145. [Google Scholar] [CrossRef]
- Nagayama, S.; Yamada, E.; Kohno, Y.; Aoyama, T.; Fukukawa, C.; Kubo, H.; Watanabe, G.; Katagiri, T.; Nakamura, Y.; Sakai, Y.; et al. Inverse correlation of the up-regulation of FZD10 expression and the activation of beta-catenin in synchronous colorectal tumors. Cancer Sci. 2009, 100, 405–412. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| cNHERF1 | nNHERF1 | β-catenin | FZD 4 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Negative | Positive | p-Value | Negative | Positive | p-Value | Negative | Positive | p-Value | Negative | Positive | p-Value | |||||||||
| Characteristics | n | (%) | n | (%) | n | (%) | n | (%) | n | (%) | n | (%) | n | (%) | n | (%) | ||||
| Patient age | ||||||||||||||||||||
| ≤54years | 49 | (26.1) | 50 | (26.6) | 0.0865 | 87 | (46) | 12 | (6.4) | 0.9831 | 39 | (20.2) | 58 | (30) | 0.4298 | 46 | (23.8) | 51 | (26.4) | 0.1632 |
| >54 years | 33 | (17.5) | 56 | (29.8) | 79 | (41.8) | 11 | (5.8) | 44 | (22.8) | 52 | (27) | 36 | (18.7) | 60 | (31.1) | ||||
| Histological type | ||||||||||||||||||||
| IDC | 69 | (36.7) | 92 | (48.9) | 0.7512 | 141 | (74.6) | 21 | (11.1) | 0.2613 | 69 | (35.8) | 101 | (52.3) | 0.0138 | 65 | (33.7) | 103 | (53.4) | 0.0199 |
| ILC | 7 | (3.7) | 9 | (4.9) | 16 | (8.5) | 0 | / | 10 | (5.2) | 2 | (1) | 9 | (4.7) | 5 | (2.6) | ||||
| Other | 6 | (3.2) | 5 | (2.6) | 9 | (4.8) | 2 | (1) | 4 | (2) | 7 | (3.7) | 8 | (4.1) | 3 | (1.5) | ||||
| Tumor size | ||||||||||||||||||||
| ≤2 cm | 58 | (32.2) | 47 | (26.2) | 0.0020 | 94 | (52) | 12 | (6.6) | 0.5055 | 50 | (26.8) | 60 | (32.2) | 0.2420 | 57 | (30.6) | 53 | (28.5) | 0.0010 |
| >2 cm | 24 | (13.3) | 51 | (28.3) | 64 | (35.4) | 11 | (6) | 28 | (15) | 48 | (26) | 21 | (11.3) | 55 | (29.6) | ||||
| Lymph node status | ||||||||||||||||||||
| Negative | 50 | (26.9) | 61 | (32.8) | 0.7486 | 98 | (53.7) | 13 | (7) | 0.7417 | 51 | (27.3) | 61 | (32.6) | 0.3521 | 49 | (26.2) | 61 | (32.6) | 0.5599 |
| Positive | 32 | (17.2) | 43 | (23.1) | 65 | (35) | 10 | (5.3) | 29 | (15.5) | 46 | (24.6) | 31 | (16.6) | 46 | (24.6) | ||||
| Histological grade | ||||||||||||||||||||
| G1–2 | 49 | (26.8) | 49 | (26.8) | 0.1294 | 82 | (44.6) | 17 | (9.2) | 0.0386 | 49 | (26) | 54 | (28,7) | 0.0896 | 56 | (30) | 47 | (25) | 0.0004 |
| G3 | 33 | (18) | 52 | (28.4) | 79 | (42.9) | 6 | (3.3) | 30 | (16) | 55 | (29,3) | 24 | (12.8) | 60 | (32.2) | ||||
| Estrogen Receptor | ||||||||||||||||||||
| ER-negative (≤10%) | 26 | (14) | 31 | (16.7) | 0.7802 | 51 | (27.3) | 7 | (3.7) | 0.9487 | 13 | (6.9) | 48 | (25.4) | <0.0001 | 20 | (10.6) | 39 | (20.6) | 0.1141 |
| ER-positive (>10%) | 56 | (30.1) | 73 | (39.2) | 113 | (60.4) | 16 | (8.6) | 67 | (35.4) | 61 | (32.3) | 60 | (31.7) | 70 | (37.1) | ||||
| Progesterone Receptor | ||||||||||||||||||||
| PR-negative (≤10%) | 37 | (19.9) | 49 | (26.3) | 0.7866 | 77 | (41.2) | 10 | (5.3) | 0.7545 | 29 | (15) | 62 | (33) | 0.0050 | 34 | (18) | 54 | (28.6) | 0.3376 |
| PR-positive (>10%) | 45 | (24.2) | 55 | (29.6) | 87 | (46.5) | 13 | (7) | 51 | (27) | 47 | (25) | 46 | (24.3) | 55 | (29.1) | ||||
| HER2/neu | ||||||||||||||||||||
| Negative (0,1+) | 70 | (38.2) | 89 | (48.6) | 0.8281 | 143 | (77.8) | 17 | (9.2) | 0.7832 | 68 | (39.3) | 94 | (54.3) | 0.0002 | 66 | (35.5) | 96 | (51.6) | 0.3909 |
| Positive (3+) | 10 | (5.5) | 14 | (7.7) | 21 | (11.4) | 3 | (1.6) | 11 | (6.4) | 0 | / | 12 | (6.4) | 12 | (6.4) | ||||
| Ki67 | ||||||||||||||||||||
| Negative (≤ 20%) | 48 | (26) | 54 | (29.1) | 0.4065 | 91 | (49) | 12 | (6.4) | 0.7414 | 54 | (28.7) | 52 | (27.7) | 0.0082 | 52 | (27.6) | 55 | (29.3) | 0.0540 |
| Positive (> 20%) | 34 | (18.4) | 49 | (26.5) | 72 | (38.7) | 11 | (5.9) | 26 | (13.8) | 56 | (29.8) | 28 | (14.9) | 53 | (28.2) | ||||
| LRP5 | LRP6 | TCF1 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Negative | Positive | p-Value | Negative | Positive | p-Value | Negative | Positive | p-Value | |||||||
| Characteristics | n | (%) | n | (%) | n | (%) | n | (%) | n | (%) | n | (%) | |||
| Patient age | |||||||||||||||
| ≤54 years | 37 | (20.6) | 54 | (30) | 0.9771 | 45 | (26.6) | 44 | (26) | 0.0175 | 45 | (23) | 59 | (30) | 0.9014 |
| >54 years | 36 | (20) | 53 | (29.4) | 26 | (15.4) | 54 | (32) | 39 | (20) | 53 | (27) | |||
| Histological type | |||||||||||||||
| IDC | 61 | (33.9) | 96 | (53.3) | 0.4697 | 59 | (35) | 89 | (52.6) | 0.1696 | 74 | (37.8) | 94 | (48) | 0.1396 |
| ILC | 7 | (3.9) | 6 | (3.3) | 5 | (3) | 6 | (3.5) | 3 | (1.5) | 12 | (6.1) | |||
| Other | 5 | (2.8) | 5 | (2.8) | 7 | (4.1) | 3 | (1.8) | 7 | (3.6) | 6 | (3) | |||
| Tumor size | |||||||||||||||
| ≤2 cm | 54 | (31) | 51 | (29.3) | 0.0009 | 51 | (31.3) | 49 | (30) | 0.0089 | 50 | (26.5) | 64 | (33.8) | 0.8714 |
| >2 cm | 18 | (10.3) | 51 | (29.3) | 19 | (11.7) | 44 | (27) | 32 | (17) | 43 | (22.7) | |||
| Lymph node status | |||||||||||||||
| Negative | 42 | (24.1) | 63 | (36.2) | 0.6486 | 42 | (25.6) | 59 | (36) | 0.7187 | 50 | (26.3) | 60 | (31.6) | 0.5640 |
| Positive | 30 | (17.2) | 39 | (22.5) | 28 | (17) | 35 | (21.4) | 33 | (17.4) | 47 | (24.7) | |||
| Histological grade | |||||||||||||||
| G1–2 | 50 | (28.7) | 42 | (24.1) | 0.0002 | 47 | (28.7) | 40 | (24.4) | 0.0010 | 49 | (25.8) | 56 | (29.5) | 0.2778 |
| G3 | 22 | (12.7) | 60 | (34.5) | 22 | (13.4) | 55 | (33.5) | 33 | (17.4) | 52 | (27.3) | |||
| Estrogen Receptor | |||||||||||||||
| ER-negative (≤10%) | 19 | (10.8) | 38 | (21.6) | 0.1571 | 12 | (7.2) | 39 | (23.5) | 0.0012 | 20 | (10.4) | 42 | (21.9) | 0.0432 |
| ER-positive (>10%) | 53 | (30.1) | 66 | (37.5) | 58 | (35) | 57 | (34.3) | 62 | (32.3) | 68 | (35.4) | |||
| Progesterone Receptor | |||||||||||||||
| PR-negative (≤10%) | 27 | (15.3) | 53 | (30.1) | 0.0778 | 23 | (13.8) | 53 | (31.9) | 0.0043 | 34 | (17.7) | 57 | (29.7) | 0.1552 |
| PR-positive (>10%) | 45 | (25.6) | 51 | (29) | 47 | (28.3) | 43 | (26) | 48 | (25) | 53 | (27.6) | |||
| HER2/neu | |||||||||||||||
| Negative (0,1+) | 58 | (33.3) | 94 | (54) | 0.0619 | 60 | (36.1) | 84 | (50.6) | 0.7376 | 63 | (33.3) | 100 | (53) | 0.0670 |
| Positive (3+) | 13 | (7.5) | 9 | (5.2) | 10 | (6) | 12 | (7.3) | 15 | (8) | 11 | (5.8) | |||
| Ki67 | |||||||||||||||
| Negative (≤20%) | 51 | (29.1) | 51 | (29.1) | 0.0049 | 47 | (28.5) | 47 | (28.5) | 0.0235 | 42 | (22) | 66 | (34.5) | 0.1979 |
| Positive (>20%) | 21 | (12) | 52 | (29.8) | 23 | (14) | 48 | (29) | 40 | (21) | 43 | (22.5) | |||
| β-catenin | FZD4 | LRP5 | LRP6 | TCF1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| cNHERF1 | 0.01 | 0.883 | 0.34 | <0.0001 | 0.24 | 0.0022 | 0.26 | 0.0010 | 0.18 | 0.0169 |
| nNHERF1 | 0.047 | 0.5394 | −0.12 | 0.1110 | −0.21 | 0.0079 | −0.25 | 0.0017 | −0.21 | 0.0048 |
| DFS | OS | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Characteristics | N. pts | N. events | 5-yrs % DFS | p-Value | HR (95% CI) | p-Value | N. events | 5-yrs % OS | p-Value | HR (95% CI) | p-Value |
| Overall | 200 | 51 | 80 | - | - | - | 10 | 95 | - | - | - |
| β-catenin | |||||||||||
| <5 | 75 | 9 | 92 | 1.00 | 2 | 98 | 1.00 | ||||
| ≥5 | 101 | 41 | 67 | <0.0001 | 4.70 (2.20–10.04) | <0.0001 | 8 | 92 | 0.139 | 3.03 (0.64–14.28) | 0.161 |
| FZD4 | |||||||||||
| <77 | 76 | 9 | 91 | 1.00 | 3 | 97 | 1.00 | ||||
| ≥77 | 101 | 39 | 71 | 0.0001 | 3.76 (1.81–7.82) | 0.0004 | 7 | 93 | 0.399 | 1.77 (0.46–6.87) | 0.406 |
| LRP5 | |||||||||||
| <83 | 66 | 6 | 93 | 1.00 | 1 | 98 | 1.00 | ||||
| ≥83 | 99 | 39 | 70 | <0.0001 | 5.44 (2.29–12.96) | 0.0001 | 8 | 91 | 0.068 | 5.56 (0.70–44.51) | 0.106 |
| LRP6 | |||||||||||
| <74 | 65 | 5 | 95 | 1.00 | 2 | 98 | 1.00 | ||||
| ≥74 | 92 | 38 | 68 | <0.0001 | 6.14 (2.39–15.75) | 0.0002 | 7 | 91 | 0.290 | 2.28 (0.47–10.98) | 0.304 |
| TCF1 | |||||||||||
| <16 | 78 | 9 | 92 | 1.00 | 2 | 100 | 1.00 | ||||
| ≥16 | 98 | 40 | 68 | <0.0001 | 4.15 (2.00–8.61) | 0.0001 | 7 | 91 | 0.165 | 2.89 (0.60–13.96) | 0.185 |
| cNHERF1 | |||||||||||
| <80 | 85 | 23 | 80 | 1.00 | 7 | 92 | 1.00 | ||||
| ≥80 | 96 | 25 | 77 | 0.981 | 1.01 (0.57–1.79) | 0.981 | 3 | 97 | 0.160 | 0.39 (0.10–1.52) | 0.177 |
| nNHERF1 | |||||||||||
| 0 | 159 | 43 | 80 | 1.00 | 7 | 96 | 1.00 | ||||
| >0 | 22 | 5 | 84 | 0.621 | 0.79 (0.31–2.01) | 0.623 | 3 | 89 | 0.124 | 2.77 (0.71–10.74) | 0.141 |
| Age (years) | |||||||||||
| ≤54 | 104 | 33 | 78 | 1.00 | 6 | 95 | 1.00 | ||||
| >54 | 96 | 18 | 81 | 0.339 | 0.75 (0.42–1.36) | 0.342 | 4 | 95 | 0.707 | 0.78 (0.22–2.78) | 0.708 |
| Histotype | |||||||||||
| Ductal | 172 | 45 | 79 | 1.00 | 0.673 | 9 | 95 | 1.00 | 0.727 | ||
| Lobular | 18 | 4 | 84 | 0.59 (0.18–1.94) | 0 | 100 | 0 | ||||
| Other | 10 | 2 | 87 | 0.666 | 0.85 (0.20–3.54) | 1 | 83 | 0.446 | 2.32 (0.29–18.43) | ||
| Tumor size (cm) | |||||||||||
| ≤2.0 | 117 | 19 | 88 | 1.00 | 4 | 97 | 1.00 | ||||
| >2.0 | 75 | 29 | 68 | 0.002 | 2.44 (1.36–4.37) | 0.003 | 6 | 92 | 0.165 | 2.38 (0.67–8.45) | 0.179 |
| Node | |||||||||||
| Negative | 121 | 21 | 87 | 1.00 | 5 | 96 | 1.00 | ||||
| Positive | 75 | 26 | 72 | 0.001 | 2.58 (1.42–4.67) | 0.002 | 5 | 93 | 0.359 | 1.77 (0.51–6.12) | 0.366 |
| Grade | |||||||||||
| 1-2 | 108 | 7 | 94 | 1.00 | 2 | 99 | 1.00 | ||||
| 3 | 87 | 42 | 63 | <0.0001 | 7.50 (3.35–16.78) | <0.0001 | 8 | 90 | 0.026 | 4.87 (1.03–22.93) | 0.045 |
| ER (%) | |||||||||||
| ≤10 | 63 | 36 | 58 | 1.00 | 7 | 88 | 1.00 | ||||
| >10 | 135 | 13 | 92 | <0.0001 | 0.19 (0.10–0.37) | <0.0001 | 3 | 99 | 0.024 | 0.24 (0.06–0.93) | 0.039 |
| PR (%) | |||||||||||
| ≤10 | 92 | 38 | 68 | 1.00 | 8 | 91 | 1.00 | ||||
| >10 | 106 | 11 | 90 | <0.0001 | 0.28 (0.14–0.55) | 0.0002 | 2 | 99 | 0.055 | 0.25 (0.05–1.16) | 0.077 |
| Ki67 (%) | |||||||||||
| ≤20 | 113 | 12 | 90 | 1.00 | 1 | 99 | 1.00 | ||||
| >20 | 84 | 36 | 68 | <0.0001 | 3.68 (1.90–7.13) | 0.0001 | 9 | 90 | 0.004 | 11.25 (1.42–88.83) | 0.022 |
| HER2 | |||||||||||
| Negative | 173 | 48 | 78 | 1.00 | 10 | 94 | 1.00 | ||||
| Positive | 26 | 2 | 88 | 0.137 | 0.36 (0.09–1.48) | 0.155 | 0 | 100 | 0.275 | 0 | 0.993 |
| DFS | OS | |||
|---|---|---|---|---|
| HR (95% CI) | p-Value | HR (95% CI) | p-Value | |
| Grade (3 vs. 1–2) | 5.28 (1.60–17.41) | 0.006 | - | - |
| β-catenin | 3.26 (1.14–9.33) | 0.027 | - | - |
| TCF1 | 4.86 (1.47–16.05) | 0.009 | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saponaro, C.; Scarpi, E.; Zito, F.A.; Giotta, F.; Silvestris, N.; Mangia, A. Independent Negative Prognostic Role of TCF1 Expression within the Wnt/β-Catenin Signaling Pathway in Primary Breast Cancer Patients. Cancers 2019, 11, 1035. https://doi.org/10.3390/cancers11071035
Saponaro C, Scarpi E, Zito FA, Giotta F, Silvestris N, Mangia A. Independent Negative Prognostic Role of TCF1 Expression within the Wnt/β-Catenin Signaling Pathway in Primary Breast Cancer Patients. Cancers. 2019; 11(7):1035. https://doi.org/10.3390/cancers11071035
Chicago/Turabian StyleSaponaro, Concetta, Emanuela Scarpi, Francesco Alfredo Zito, Francesco Giotta, Nicola Silvestris, and Anita Mangia. 2019. "Independent Negative Prognostic Role of TCF1 Expression within the Wnt/β-Catenin Signaling Pathway in Primary Breast Cancer Patients" Cancers 11, no. 7: 1035. https://doi.org/10.3390/cancers11071035
APA StyleSaponaro, C., Scarpi, E., Zito, F. A., Giotta, F., Silvestris, N., & Mangia, A. (2019). Independent Negative Prognostic Role of TCF1 Expression within the Wnt/β-Catenin Signaling Pathway in Primary Breast Cancer Patients. Cancers, 11(7), 1035. https://doi.org/10.3390/cancers11071035