Ductal Carcinoma In Situ Progression in Dog Model of Breast Cancer

 , , ,
, , ,
Abstract
:1. Introduction
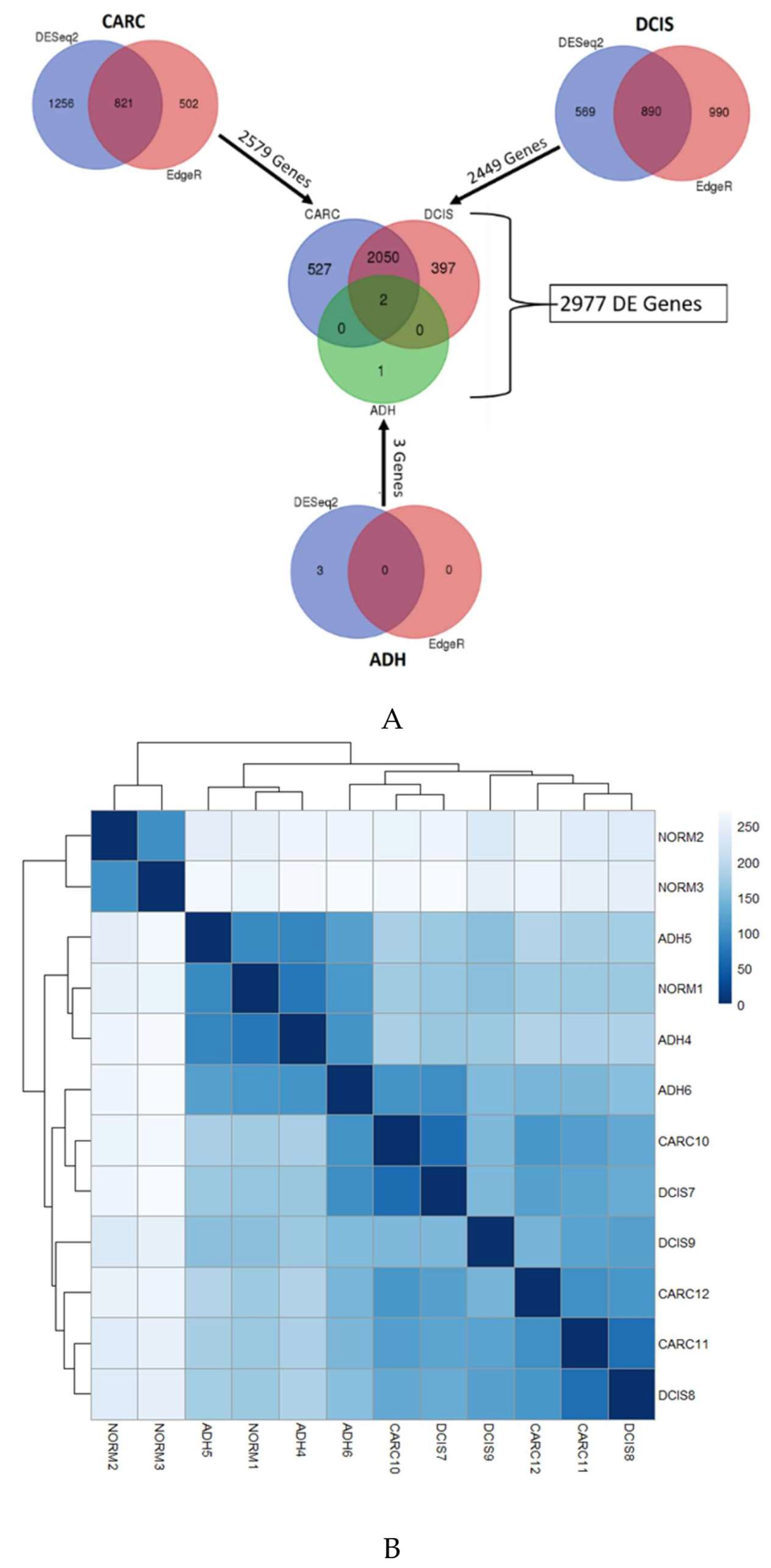
2. Results
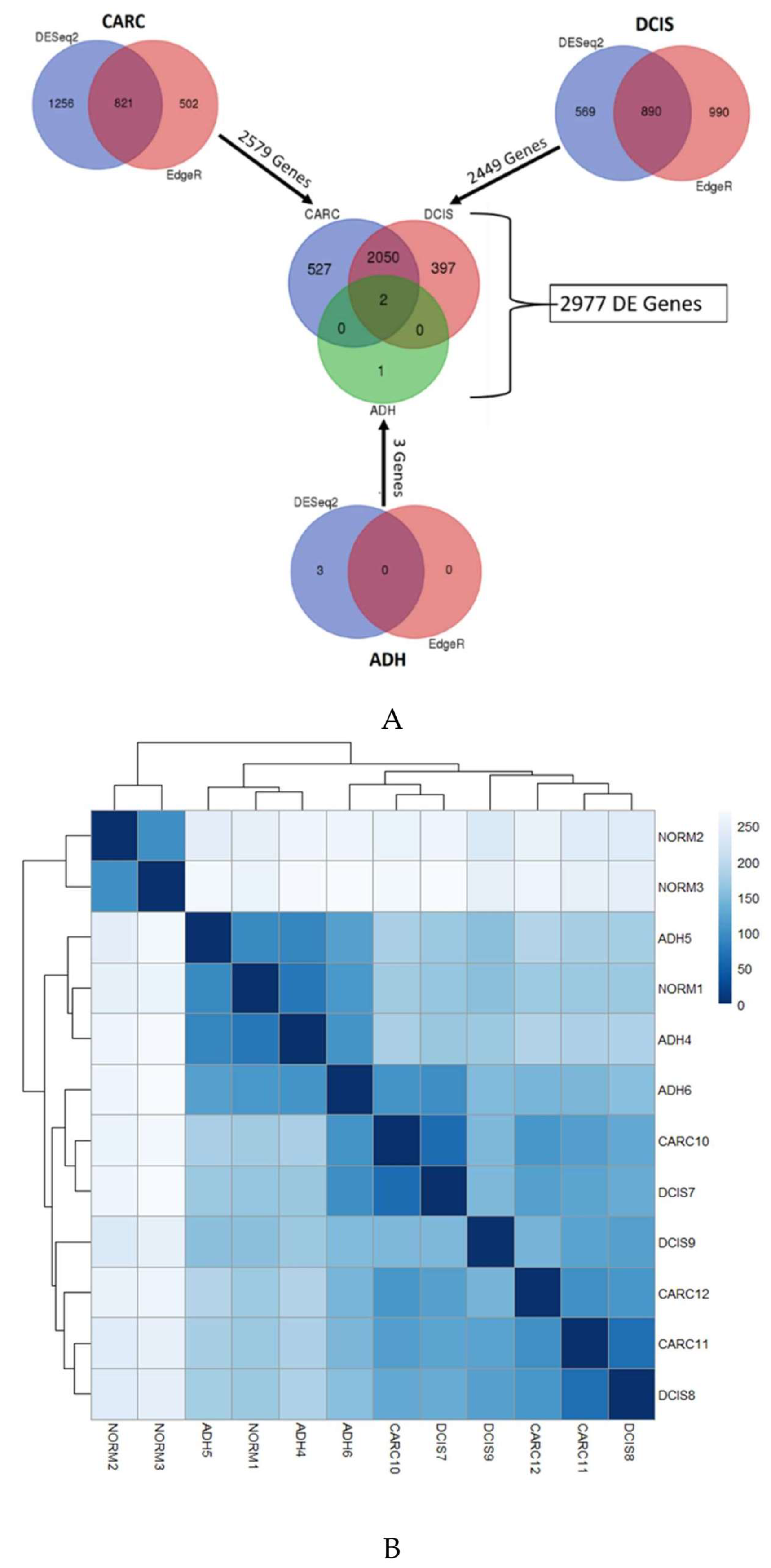
2.1. Gene Expression Profiles of ADH, DCIS and Invasive Cancer
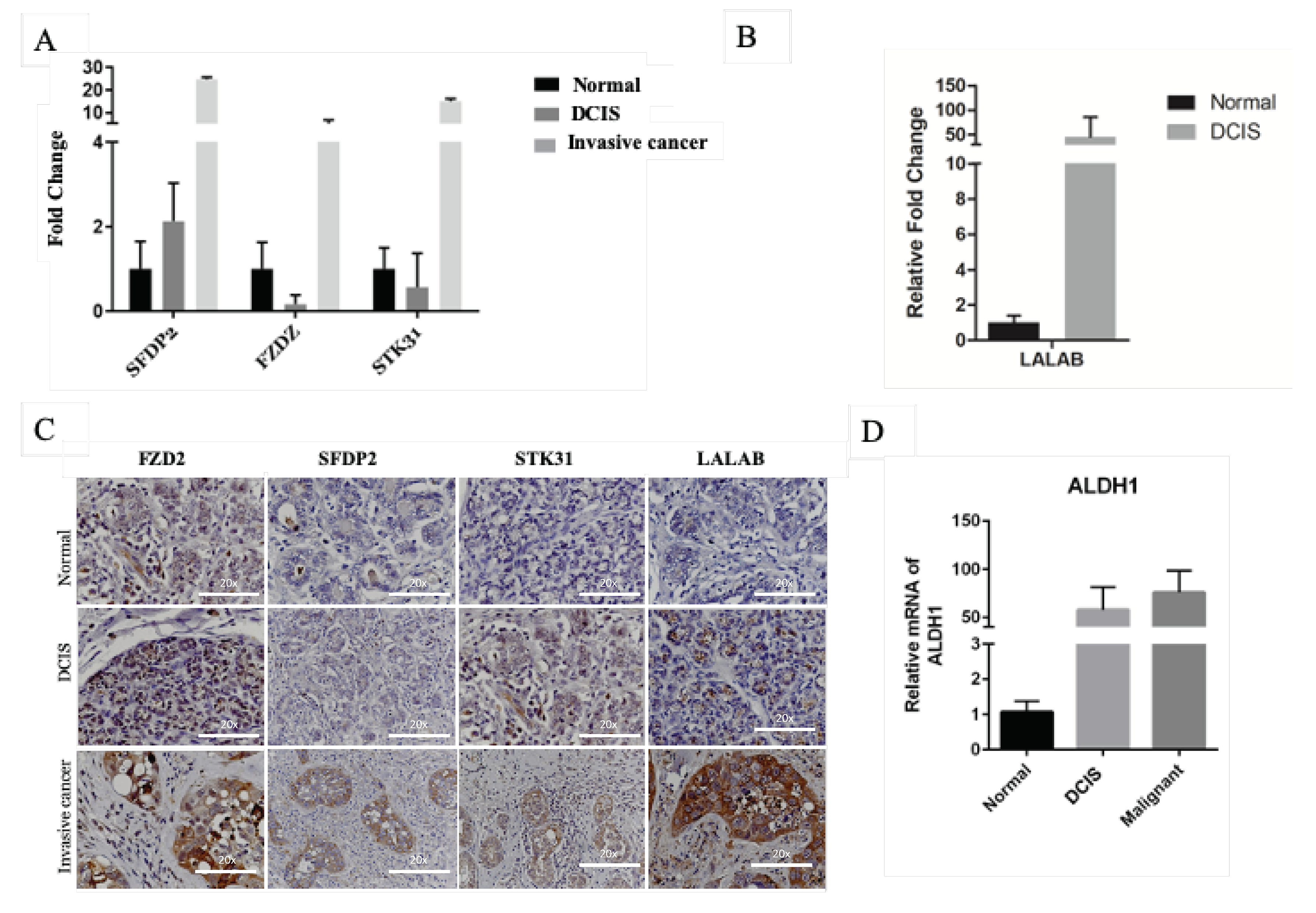
2.2. Identification of Genes Associated with DCIS Progression to Invasive Cancer
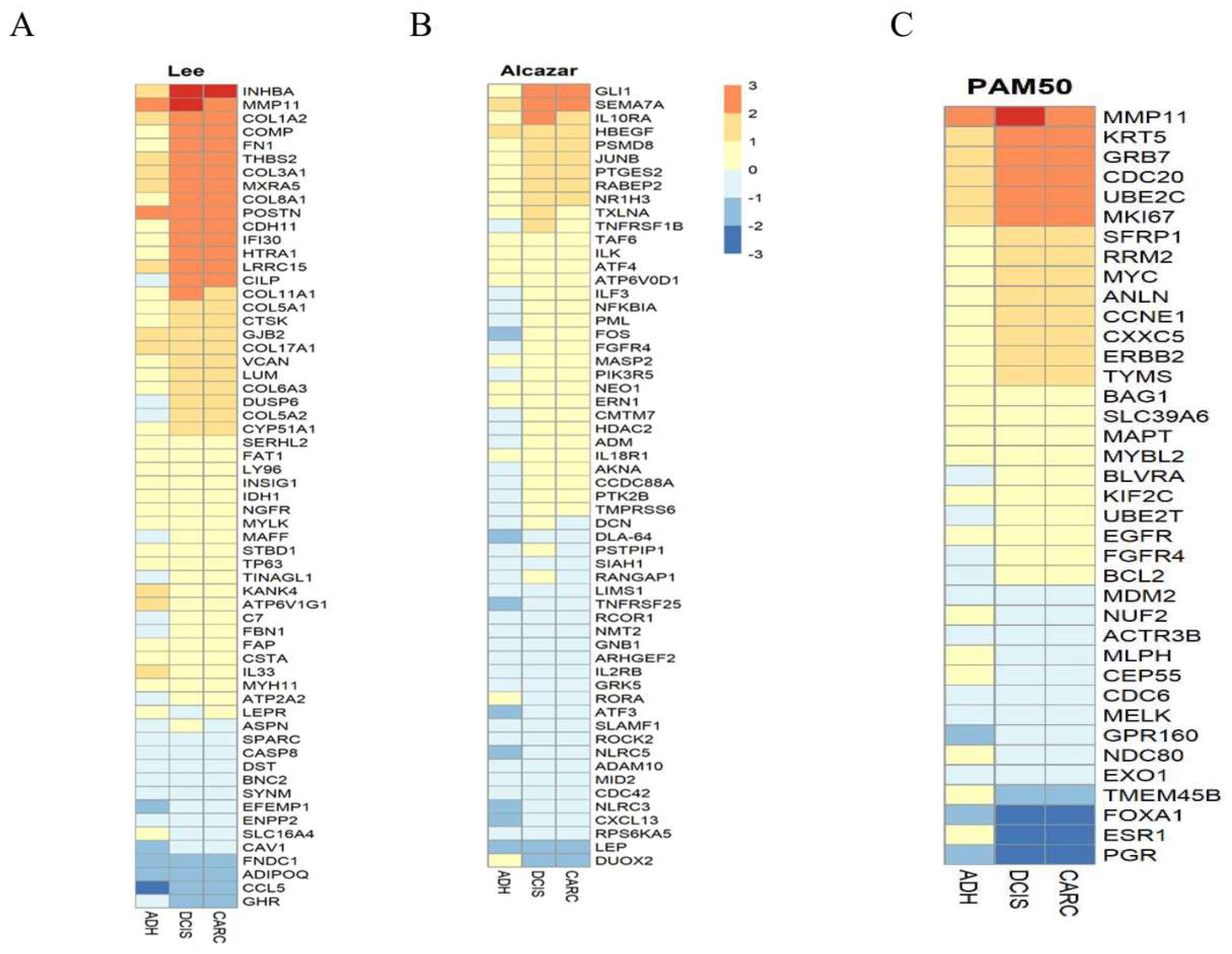
2.3. Genes Expression Similarities between Canine and Humans DCIS and Invasive Cancer
3. Discussion
4. Materials and Methods
4.1. Clinical Specimens
4.2. RNA Isolation
4.3. Next-Generation Sequencing
4.4. Bioinformatics Analysis
4.5. Pathway Enrichment Analysis
4.6. Quantitative Real-Time RT-PCR (qRT-PCR)
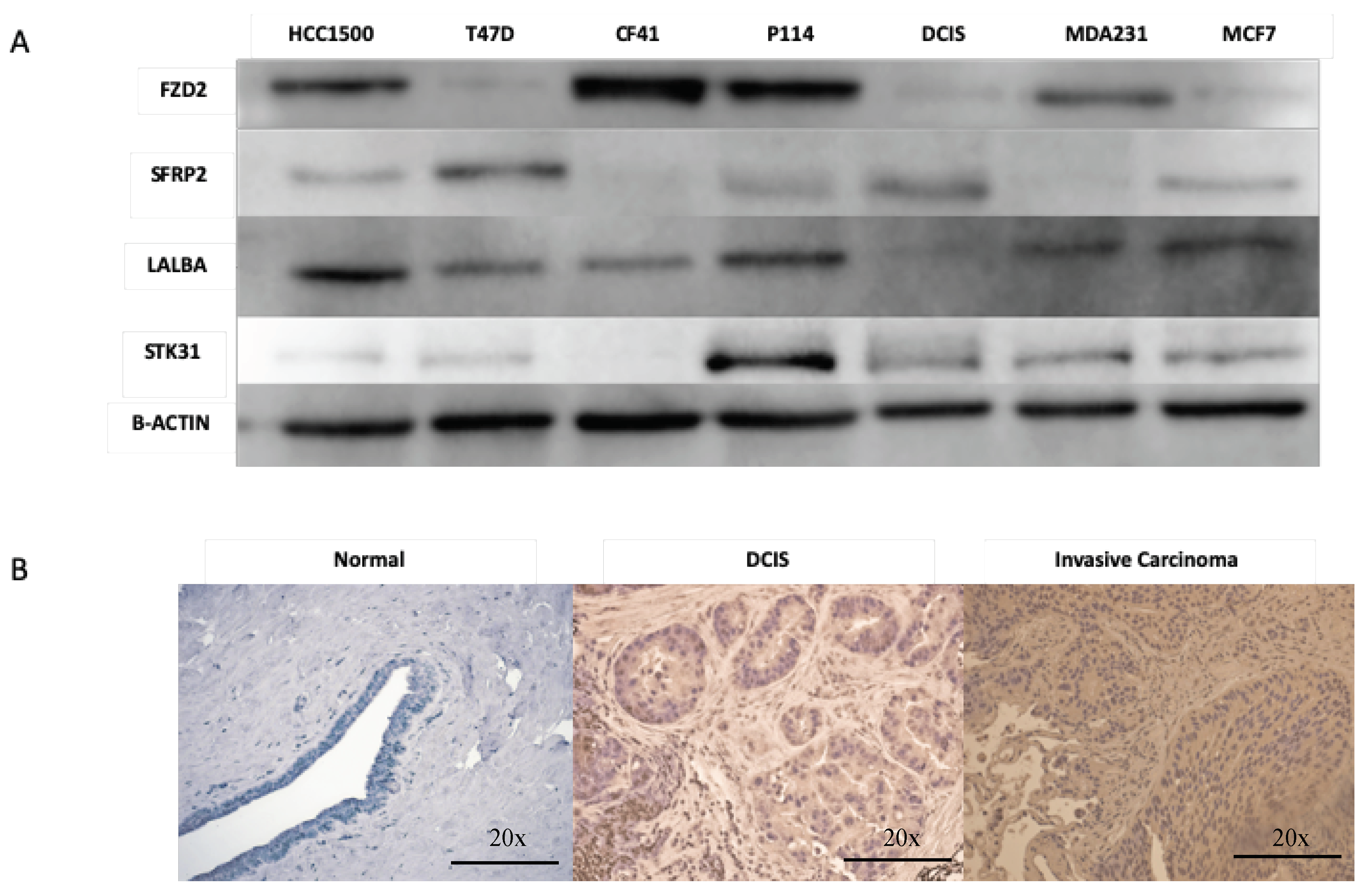
4.7. Immunohistochemistry
4.8. Western Blot Analysis
4.9. Kaplan Meier Srvivial Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data availability
References
- Gorringe, K.L.; Fox, S.B. Ductal Carcinoma In Situ Biology, Biomarkers, and Diagnosis. Front. Oncol. 2017, 7, e248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, E.M.; DeSantis, C.E.; Lin, C.C.; Kramer, J.L.; Jemal, A.; Kohler, B.; Brawley, O.W.; Gansler, T. Cancer Statistics: Breast Cancer in Situ. CA. Cancer J. Clin. 2015, 65, 481–495. [Google Scholar] [CrossRef]
- Ernster, V.L.; Barclay, J.; Kerlikowske, K.; Wilkie, H.; Ballard-Barbash, R. Mortality among Women with Ductal Carcinoma in Situ of the Breast in the Population-Based Surveillance, Epidemiology and End Results Program. Arch. Intern. Med. 2000, 160, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.; Feig, S.A.; Drexler, M.; Lehman, C. Implications of Overdiagnosis: Impact on Screening Mammography Practices. Popul. Health Manag. 2015, 18, S3–S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Garg, C.; Löhler, J.; Gocht, A.; Deppert, W. A Transgenic Mouse Model for the Ductal Carcinoma in Situ (DCIS) of the Mammary Gland. Oncogene 2000, 19, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Souter, L.H.; Andrews, J.D.; Zhang, G.; Cook, A.C.; Postenka, C.O.; Al-Katib, W.; Leong, H.S.; Rodenhiser, D.I.; Chambers, A.F.; Tuck, A.B. Human 21T Breast Epithelial Cell Lines Mimic Breast Cancer Progression in Vivo and in Vitro and Show Stage-Specific Gene Expression Patterns. Lab. Investig. 2010, 90, 1247–1258. [Google Scholar] [CrossRef] [Green Version]
- Lampreht Tratar, U.; Horvat, S.; Cemazar, M. Transgenic Mouse Models in Cancer Research. Front. Oncol. 2018, 8, e268. [Google Scholar] [CrossRef] [Green Version]
- National Academies of Sciences, Engineering, and Medicine. The Role of Clinical Studies for Pets with Naturally Occurring Tumors in Translational Cancer Research: Workshop Summary; National Academy Press: Washington, DC, USA, 2015. [Google Scholar] [CrossRef]
- Vail, D.M.; Macewen, E.G. Spontaneously Occurring Tumors of Companion Animals as Models for Human Cancer. Cancer Invest. 2000, 18, 781–792. [Google Scholar] [CrossRef]
- Owen, L. A Comparative Study of Canine and Human Breast Cancer. Invest. Cell Pathol. 1979, 2, 257–275. [Google Scholar]
- Gilbertson, S.R.; Kurzman, I.D.; Zachrau, R.E.; Hurvitz, A.I.; Black, M.M. Canine Mammary Epithelial Neoplasms: Biologic Implications of Morphologic Characteristics Assessed in 232 Dogs. Vet. Pathol. 1983, 20, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Warner, M. Age Incidence and Site Distribution of Mammary Dysplasias in Young Beagle Bitches. J. Natl. Cancer Inst. 1976, 57, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Perez Alenza, M.D.; Peña, L.; del Castillo, N.; Nieto, A.I. Factors Influencing the Incidence and Prognosis of Canine Mammary Tumours. J. Small Anim. Pract. 2000, 41, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Antuofermo, E.; Miller, M.A.; Pirino, S.; Xie, J.; Badve, S.; Mohammed, S.I. Spontaneous Mammary Intraepithelial Lesions in Dogs--a Model of Breast Cancer. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 2247–2256. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, S.I.; Meloni, G.B.; Pinna Parpaglia, M.L.; Marras, V.; Burrai, G.P.; Meloni, F.; Pirino, S.; Antuofermo, E. Mammography and Ultrasound Imaging of Preinvasive and Invasive Canine Spontaneous Mammary Cancer and Their Similarities to Human Breast Cancer. Cancer Prev. Res. 2011, 4, 1790–1798. [Google Scholar] [CrossRef] [Green Version]
- Mouser, P.; Miller, M.A.; Antuofermo, E.; Badve, S.S.; Mohammed, S.I. Prevalence and Classification of Spontaneous Mammary Intraepithelial Lesions in Dogs without Clinical Mammary Disease. Vet. Pathol. 2010, 47, 275–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorenmo, K.; Kristiansen, V.; Cofone, M.; Shofer, F.; Breen, A.; Langeland, M.; Mongil, C.; Grondahl, A.; Teige, J.; Goldschmidt, M. Canine Mammary Gland Tumours; a Histological Continuum from Benign to Malignant; Clinical and Histopathological Evidence. Vet. Comp. Oncol. 2009, 7, 162–172. [Google Scholar] [CrossRef]
- Carvalho, M.I.; Pires, I.; Prada, J.; Queiroga, F.L. A Role for T-Lymphocytes in Human Breast Cancer and in Canine Mammary Tumors. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, e550. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential Expression Analysis of Multifactor RNA-Seq Experiments with Respect to Biological Variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Gujral, T.S.; Chan, M.; Peshkin, L.; Sorger, P.K.; Kirschner, M.W.; Macbeath, G. A Noncanonical Frizzled2 Pathway Regulates Epithelial-Mesenchymal Transition and Metastasis. Cell 2014, 159, 844–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Luo, J. Alcohol and Cancer Stem Cells. Cancers 2017, 9, e158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, E.S.; Ertel, A.; Davicioni, E.; Kline, J.; Schwartz, G.F.; Witkiewicz, A.K. Progression of Ductal Carcinoma in Situ to Invasive Breast Cancer Is Associated with Gene Expression Programs of EMT and Myoepithelia. Breast Cancer Res. 2012, 133, 1009–1024. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.J.; Salunga, R.; Tuggle, J.T.; Gaudet, J.; Enright, E.; McQuary, P.; Payette, T.; Pistone, M.; Stecker, K.; Zhang, B.M.; et al. Gene Expression Profiles of Human Breast Cancer Progression. Proc. Natl. Acad. Sci. USA 2003, 100, 5974–5979. [Google Scholar] [CrossRef] [Green Version]
- Schuetz, C.S.; Bonin, M.; Clare, S.E.; Nieselt, K.; Sotlar, K.; Walter, M.; Fehm, T.; Solomayer, E.; Riess, O.; Wallwiener, D.; et al. Progression-Specific Genes Identified by Expression Profiling of Matched Ductal Carcinomas in Situ and Invasive Breast Tumors, Combining Laser Capture Microdissection and Oligonucleotide Microarray Analysis. Cancer Res. 2006, 66, 5278–5286. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Stewart, S.; Nagtegaal, I.; Luo, J.; Wu, Y.; Colditz, G.; Medina, D.; Allred, D.C. Differentially Expressed Genes Regulating the Progression of Ductal Carcinoma in Situ to Invasive Breast Cancer. Cancer Res. 2012, 72, 4574–4586. [Google Scholar] [CrossRef] [Green Version]
- Gil Del Alcazar, C.R.; Huh, S.J.; Ekram, M.B.; Trinh, A.; Liu, L.L.; Beca, F.; Zi, X.; Kwak, M.; Bergholtz, H.; Su, Y.; et al. Immune Escape in Breast Cancer during in Situ to Invasive Carcinoma Transition. Cancer Discov. 2017, 7, 1098–1115. [Google Scholar] [CrossRef] [Green Version]
- Wallden, B.; Storhoff, J.; Nielsen, T.; Dowidar, N.; Schaper, C.; Ferree, S.; Liu, S.; Leung, S.; Geiss, G.; Snider, J.; et al. Development and Verification of the PAM50-Based Prosigna Breast Cancer Gene Signature Assay. BMC Med. Genomics 2015, 8, e54. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-Responsive Genes Involved in Oxidative Phosphorylation Are Coordinately Downregulated in Human Diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Page, D.L.; Dupont, W.D.; Rogers, L.W.; Jensen, R.A.; Schuyler, P.A. Continued Local Recurrence of Carcinoma 15–25 Years after a Diagnosis of Low Grade Ductal Carcinoma in Situ of the Breast Treated Only by Biopsy. Cancer 1995, 76, 1197–1200. [Google Scholar] [CrossRef]
- Moore, E.; Magee, H.; Coyne, J.; Gorey, T.; Dervan, P.A. Widespread Chromosomal Abnormalities in High-Grade Ductal Carcinoma in Situ of the Breast. Comparative Genomic Hybridization Study of Pure High- Grade DCIS. J. Pathol. 1999, 187, 403–409. [Google Scholar] [CrossRef]
- Felipe Lima, J.; Nofech-Mozes, S.; Bayani, J.; Bartlett, J. EMT in Breast Carcinoma—A Review. J. Clin. Med. 2016, 5, e65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wei, H.; Liu, F.; Guan, J.-L. Hyper-Activation of MTORC1 Promotes Breast Cancer Progression through Enhancing Glucose Starvation-Induced Autophagy and Akt Signaling. J. Biol. Chem. 2013. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Tao, D.; Fang, Y.; Deng, C.; Xu, Q.; Zhou, J. TNF-Alpha Promotes Invasion and Metastasis via NF-Kappa B Pathway in Oral Squamous Cell Carcinoma. Med. Sci. Monit. Basic Res. 2017, 23, e141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, A.P.; Lièvre, M.; Thomas, C.; Hinkal, G.; Ansieau, S.; Puisieux, A. Generation of Breast Cancer Stem Cells through Epithelial-Mesenchymal Transition. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Seth, A.; Kitching, R.; Landberg, G.; Xu, J.; Zubovits, J.; Burger, A.M. Gene Expression Profiling of Ductal Carcinomas in Situ and Invasive Breast Tumors. Anticancer Res. 2003, 23, 2043–2051. [Google Scholar]
- Porter, D.; Lahti-Domenici, J.; Keshaviah, A.; Bae, Y.; Argani, P.; Marks, J.; Richardson, A.; Cooper, A.; Strausberg, R.; Riggins, G.; et al. Molecular Markers in Ductal Carcinoma in Situ of the Breast. Mol. Cancer Res. 2003, 1, 362–375. [Google Scholar]
- Porter, D.A.; Krop, I.E.; Polyak, K.; Porter, D.A.; Krop, I.E.; Nasser, S.; Sgroi, D.; Kaelin, C.M.; Polyak, K.; Nasser, S.; et al. A SAGE (Serial Analysis of Gene Expression) View of Breast Tumor Progression. Cancer Res. 2001, 61, 5697–5702. [Google Scholar]
- Hannemann, J.; Velds, A.; Halfwerk, J.B.G.; Kreike, B.; Peterse, J.L.; Van de Vijver, M.J. Classification of Ductal Carcinoma in Situ by Gene Expression Profiling. Breast Cancer Res. 2006, 8, R61. [Google Scholar] [CrossRef] [Green Version]
- Ein-Dor, L.; Kela, I.; Getz, G.; Givol, D.; Domany, E. Outcome Signature Genes in Breast Cancer: Is There a Unique Set? Bioinformatics 2005, 21, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Lo, P.K.; Zhang, Y.; Yao, Y.; Wolfson, B.; Yu, J.; Han, S.Y.; Duru, N.; Zhou, Q. Tumor-Associated Myoepithelial Cells Promote the Invasive Progression of Ductal Carcinoma in Situ through Activation of TGFβ Signaling. J. Biol. Chem. 2017, 292, 11466–11484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, R.; Mihich, E. Eighteenth Annual Pezcoller Symposium: Tumor Microenvironment and Heterotypic Interactions. Cancer Res. 2006, 66, 11550–11553. [Google Scholar] [CrossRef] [Green Version]
- Grigoriadis, A.; Mackay, A.; Reis-Filho, J.S.; Steele, D.; Iseli, C.; Stevenson, B.J.; Jongeneel, C.V.; Valgeirsson, H.; Fenwick, K.; Iravani, M.; et al. Establishment of the Epithelial-Specific Transcriptome of Normal and Malignant Human Breast Cells Based on MPSS and Array Expression Data. Breast Cancer Res. 2006, 8, R56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Watkinson, J.; Varadan, V.; Anastassiou, D. Multi-Cancer Computational Analysis Reveals Invasion-Associated Variant of Desmoplastic Reaction Involving INHBA, THBS2 and COL11A1. BMC Med. Genomics 2010, 3, e51. [Google Scholar] [CrossRef] [Green Version]
- Vincent, K.M.; Postovit, L.M. A Pan-Cancer Analysis of Secreted Frizzled-Related Proteins: Re-Examining Their Proposed Tumour Suppressive Function. Sci. Rep. 2017, 7, e42719. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Wang, W.; Zhang, Z.; Bai, Y.; Gao, J.; Zhao, C. Wnt Signaling in Human and Mouse Breast Cancer: Focusing on Wnt Ligands, Receptors and Antagonists. Cancer Sci. 2018, 109, 3368–3375. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Orte, E.; Sáenz-Narciso, B.; Moreno, S.; Cabello, J. Multiple Functions of the Noncanonical Wnt Pathway. Trends Genet. 2013, 29, 545–553. [Google Scholar] [CrossRef]
- Kuo, P.L.; Huang, Y.L.; Hsieh, C.C.J.; Lee, J.C.; Lin, B.W.; Hung, L.Y. STK31 Is a Cell-Cycle Regulated Protein That Contributes to the Tumorigenicity of Epithelial Cancer Cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Coombes, R.C.; Caballero, O.L.; Shousha, S.; Ghaem-Maghami, S.; Woodley-Barker, L.; Wilhelm-Benartzi, C.S.; Neville, A.M. NY-ESO-1 Expression in DCIS: A New Predictor of Good Prognosis. Oncoscience 2017, 4, e33. [Google Scholar] [CrossRef]
- Vilotte, J.L.; Soulier, S.; Mercier, J.C. Sequence of the Murine α-Lactalbumin-Encoding CDNA: Interspecies Comparison of the Coding Frame and Deduced Preprotein. Gene 1992, 112, 251–255. [Google Scholar] [CrossRef]
- Lee, A.; Rosen, P.; DeLellis, R.; Saigo, P.; Gangi, M.; Groshen, S.; Bagin, R.; Wolfe, H. Tumor Marker Expression in Breast Carcinomas and Relationship to Prognosis. An Immunohistochemical Study. Am. J. Clin. Pathol. 1985, 84, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq-A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Group | Replicate Name | Filename | Total Reads | Quality Control Reads | % Reads Passing Quality Control | Reads went into mapping | Total Reads Mapped | Overall Mapping Rate |
|---|---|---|---|---|---|---|---|---|
| NORM | NORM1 | NORM1T_1.fastq | 15,190,966 | 12,695,551 | 83.57 | 25,389,212 | 21,810,532 | 85.90 |
| NORM1T_2.fastq | 15,190,966 | 12,695,970 | 83.58 | |||||
| NORM2 | NORM2T_1.fastq | 48,392,239 | 48,392,019 | 100.00 | 96,783,168 | 93,500,856 | 96.61 | |
| NORM2T_2.fastq | 48,392,239 | 48,391,804 | 100.00 | |||||
| NORM3 | NORM3T_1.fastq | 55,636,922 | 55,636,682 | 100.00 | 111,272,404 | 106,104,220 | 95.36 | |
| NORM3T_2.fastq | 55,636,922 | 55,636,437 | 100.00 | |||||
| ADH | ADH4 | ADH4T_1.fastq | 12,072,501 | 9,985,381 | 82.71 | 19,969,542 | 16,901,852 | 84.64 |
| ADH4T_2.fastq | 12,072,501 | 9,985,596 | 82.71 | |||||
| ADH5 | ADH5T_1.fastq | 19,656,483 | 16,956,030 | 86.26 | 33,910,162 | 29,749,778 | 87.73 | |
| ADH5T_2.fastq | 19,656,483 | 16,956,364 | 86.26 | |||||
| ADH6 | ADH6T_1.fastq | 10,597,768 | 9,232,178 | 87.11 | 18,461,792 | 15,062,516 | 81.59 | |
| ADH6T_2.fastq | 10,597,768 | 9,232,503 | 87.12 | |||||
| DCIS | DCIS7 | DCIS7T_1.fastq | 15,265,327 | 12,725,074 | 83.36 | 25,447,108 | 20,483,162 | 80.49 |
| DCIS7T_2.fastq | 15,265,327 | 12,725,528 | 83.36 | |||||
| DCIS8 | DCIS8T_1.fastq | 21,202,092 | 17,943,923 | 84.63 | 35,886,362 | 32,592,522 | 90.82 | |
| DCIS8T_2.fastq | 21,202,092 | 17,944,297 | 84.63 | |||||
| DCIS9 | DCIS9T_1.fastq | 17,680,467 | 15,266,076 | 86.34 | 30,530,820 | 27,452,402 | 89.92 | |
| DCIS9T_2.fastq | 17,680,467 | 15,266,383 | 86.35 | |||||
| CARC | CARC10 | CARC10T_1.fastq | 11,964,706 | 9,594,034 | 80.19 | 19,186,476 | 16,066,992 | 83.74 |
| CARC10T_2.fastq | 11,964,706 | 9,594,135 | 80.19 | |||||
| CARC11 | CARC11T_1.fastq | 12,749,664 | 10,291,360 | 80.72 | 20,581,584 | 18,862,732 | 91.65 | |
| CARC11T_2.fastq | 12,749,664 | 10,291,519 | 80.72 | |||||
| CARC12 | CARC12T_1.fastq | 13,512,440 | 10,072,537 | 74.54 | 20,143,324 | 18,112,260 | 89.92 | |
| CARC12T_2.fastq | 13,512,440 | 10,072,720 | 74.54 |
| Gene ID | Gene Name | DE in Stage | DCIS log2FC | CARC log2FC | Gene Description |
|---|---|---|---|---|---|
| ENSCAFG00000000473 | TRHDE | CARC DCIS | −4.20 | −3.93 | thyrotropin releasing hormone degrading enzyme |
| ENSCAFG00000002606 | NA | ADH CARC DCIS | −4.88 | −4.83 | NA |
| ENSCAFG00000002714 | EPHA5 | CARC DCIS | −4.00 | −3.92 | EPH receptor A5 |
| ENSCAFG00000002795 | STK31 | CARC DCIS | −4.14 | −3.93 | serine/threonine kinase 31 |
| ENSCAFG00000006357 | CNTN3 | CARC DCIS | −4.82 | −4.72 | contactin 3 |
| ENSCAFG00000008210 | GREM1 | CARC DCIS | 4.75 | 4.73 | gremlin 1, DAN family BMP antagonist |
| ENSCAFG00000008337 | TMPRSS15 | CARC DCIS | −4.06 | −3.98 | transmembrane protease, serine 15 |
| ENSCAFG00000008353 | SFRP2 | CARC DCIS | 5.20 | 5.18 | secreted frizzled related protein 2 |
| ENSCAFG00000009842 | OPCML | CARC DCIS | −4.28 | −3.76 | opioid binding protein/cell adhesion molecule like |
| ENSCAFG00000010878 | TCERG1L | CARC DCIS | −4.32 | −3.93 | transcription elongation regulator 1 like |
| ENSCAFG00000013453 | NA | CARC DCIS | −4.23 | −3.83 | NA |
| ENSCAFG00000013459 | KLRD1 | CARC DCIS | −4.44 | −4.39 | Natural killer cells antigen CD94 |
| ENSCAFG00000013703 | PTH2R | CARC DCIS | −4.06 | −3.91 | parathyroid hormone 2 receptor |
| ENSCAFG00000013753 | NA | CARC DCIS | −4.48 | −4.26 | NA |
| ENSCAFG00000014134 | FZD2 | CARC DCIS | 4.41 | 4.35 | frizzled class receptor 2 |
| ENSCAFG00000015538 | PCDH15 | CARC DCIS | −4.30 | −4.07 | protocadherin related 15 |
| ENSCAFG00000017227 | GABRG2 | CARC DCIS | −4.03 | −3.79 | gamma-aminobutyric acid type A receptor gamma2 subunit |
| ENSCAFG00000019105 | NA | CARC DCIS | −4.00 | −3.66 | NA |
| ENSCAFG00000023324 | S100A12 | CARC DCIS | 4.15 | 4.11 | S100 calcium binding protein A12 |
| ENSCAFG00000023449 | KRT13 | CARC DCIS | 4.62 | 4.62 | keratin 13 |
| ENSCAFG00000025443 | NA | CARC DCIS | −4.13 | −4.14 | NA |
| ENSCAFG00000028937 | NA | CARC DCIS | −4.21 | −3.90 | NA |
| ENSCAFG00000029431 | SYNPR | CARC DCIS | −4.51 | −4.09 | Synaptoporin |
| ENSCAFG00000029470 | NA | CARC DCIS | 4.28 | 4.24 | NA |
| ENSCAFG00000030453 | FAM19A1 | CARC DCIS | −4.14 | −3.86 | family with sequence similarity 19 member A1, C-C motif chemokine like |
| ENSCAFG00000031931 | HIST1H2BJ | CARC DCIS | 4.16 | 4.10 | histone cluster 1 H2B family member j |
| ENSCAFG00000032610 | NA | CARC DCIS | −4.04 | −3.91 | NA |
| ENSCAFG00000032615 | NA | CARC | 4.36 | 4.11 | NA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, S.I.; Utturkar, S.; Lee, M.; Yang, H.H.; Cui, Z.; Atallah Lanman, N.; Zhang, G.; Ramos Cardona, X.E.; Mittal, S.K.; Miller, M.A. Ductal Carcinoma In Situ Progression in Dog Model of Breast Cancer. Cancers 2020, 12, 418. https://doi.org/10.3390/cancers12020418
Mohammed SI, Utturkar S, Lee M, Yang HH, Cui Z, Atallah Lanman N, Zhang G, Ramos Cardona XE, Mittal SK, Miller MA. Ductal Carcinoma In Situ Progression in Dog Model of Breast Cancer. Cancers. 2020; 12(2):418. https://doi.org/10.3390/cancers12020418
Chicago/Turabian StyleMohammed, Sulma I., Sagar Utturkar, Maxwell Lee, Howard H. Yang, Zhibin Cui, Nadia Atallah Lanman, GuangJun Zhang, Xavier E. Ramos Cardona, Suresh K. Mittal, and Margaret A. Miller. 2020. "Ductal Carcinoma In Situ Progression in Dog Model of Breast Cancer" Cancers 12, no. 2: 418. https://doi.org/10.3390/cancers12020418