
The Influence of the Microbiome on Immunotherapy for Gastroesophageal Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
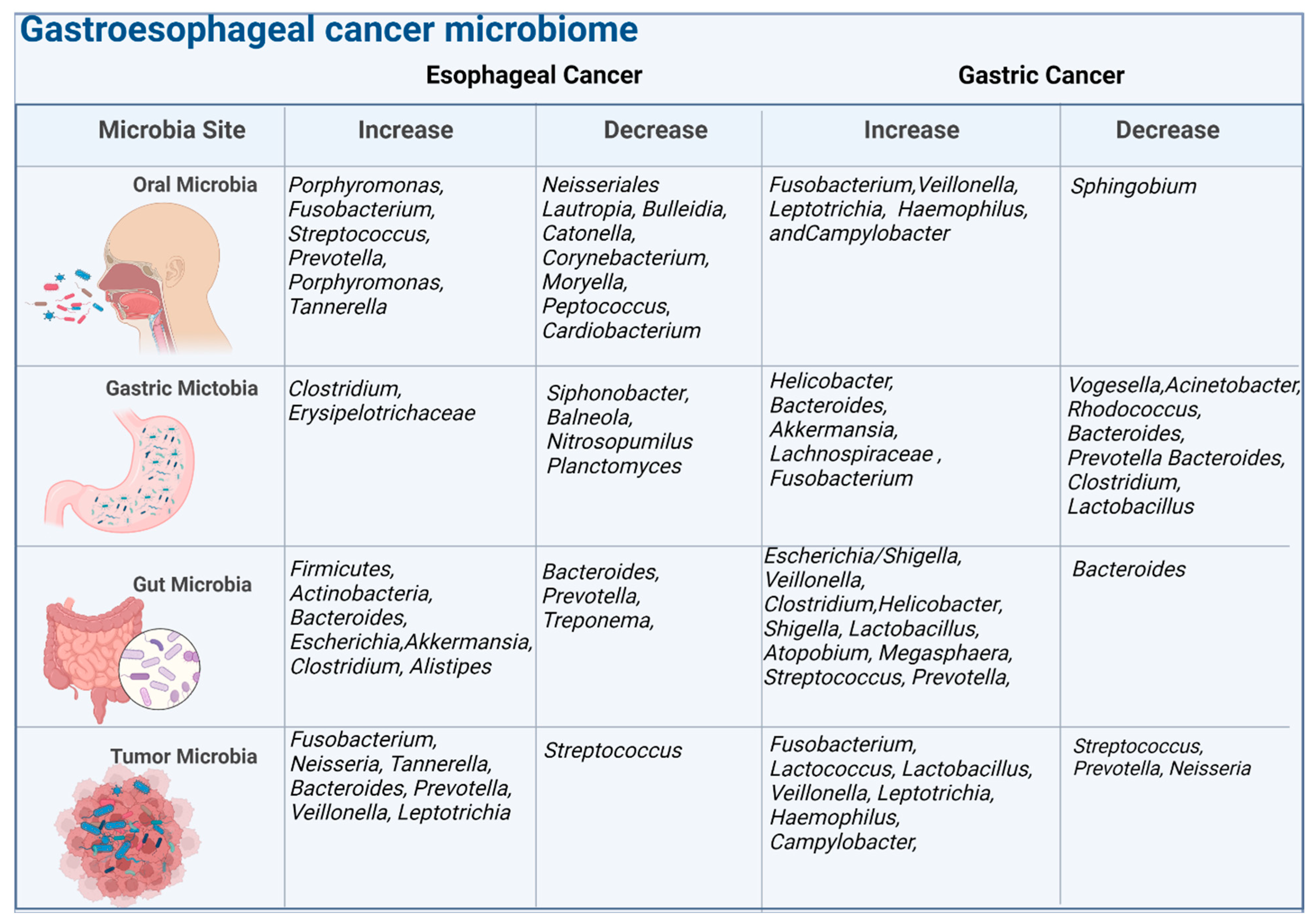
2. Tumor and Host Microbiome on the Pathogenesis of EC and GC
2.1. Microbiome in Esophageal Cancer
2.2. Microbiome in Gastric Cancer
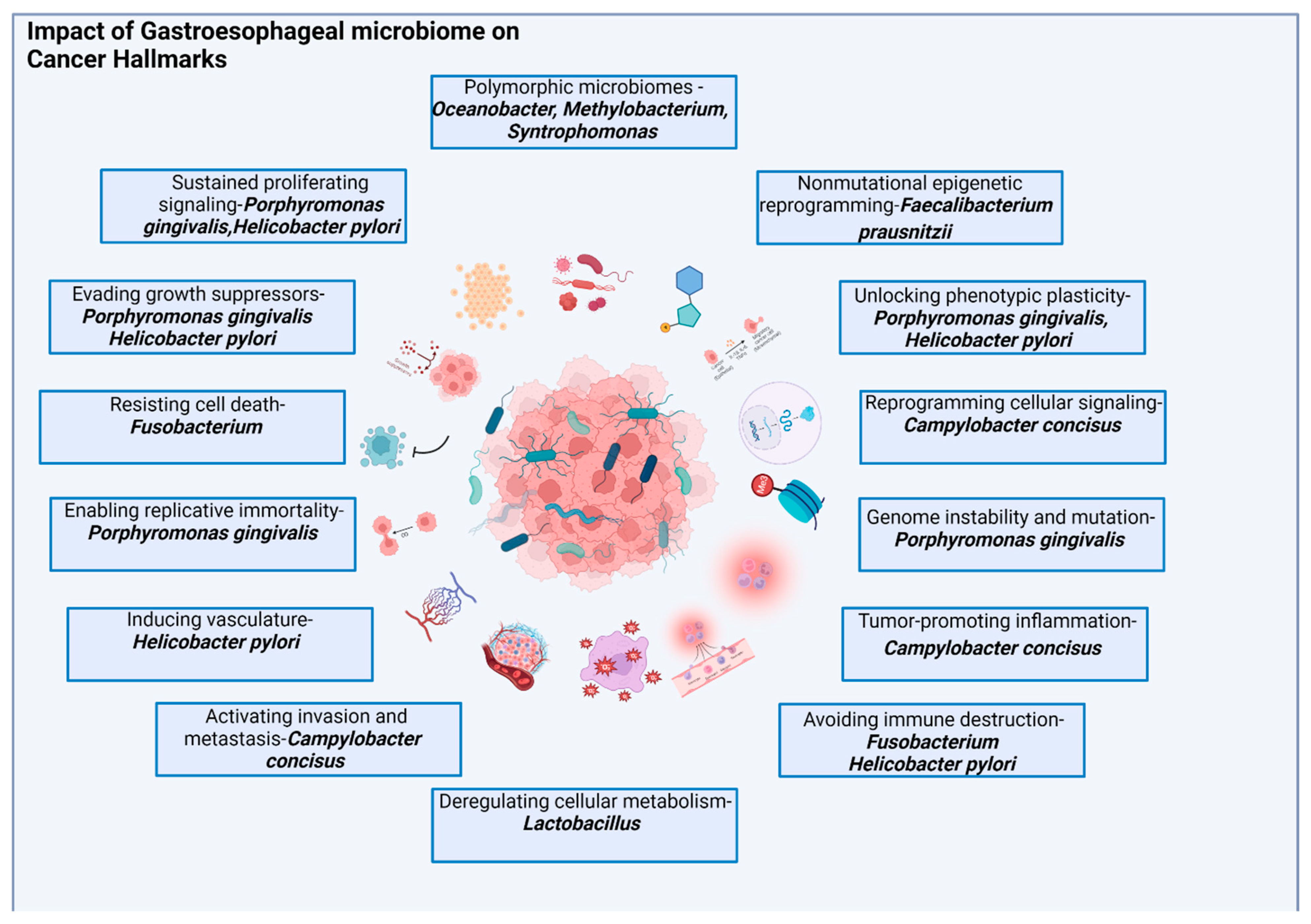
3. Mechanisms of Microbiome Impact on Esophageal and Gastric Carcinogenesis
4. Mechanism of Microbiota’s Impact on Host Immune Response
5. Current Approaches to Treating Advanced Gastroesophageal Cancers and Clinical Trials Involving Checkpoint Inhibitors
6. Connection between Microbiota, the Immune System Response of the Host, and the Impact on Cancer Immunotherapy and Treatment Toxicity
7. Summary and Recommendations for Improving the Effectiveness of Microbiota in Cancer Immunotherapy
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hyland, P.L.; Hu, N.; Rotunno, M.; Su, H.; Wang, C.; Wang, L.; Pfeiffer, R.M.; Gherman, B.; Giffen, C.; Dykes, C.; et al. Global changes in gene expression of Barrett’s esophagus compared to normal squamous esophagus and gastric cardia tissues. PLoS ONE 2014, 9, e93219. [Google Scholar]
- Jardim, S.R.; de Souza, L.M.P.; de Souza, H.S.P. The Rise of Gastrointestinal Cancers as a Global Phenomenon: Unhealthy Behavior or Progress? Int. J. Environ. Res. Public Health 2023, 20, 3640. [Google Scholar] [PubMed]
- Sheikh, M.; Roshandel, G.; McCormack, V.; Malekzadeh, R. Current Status and Future Prospects for Esophageal Cancer. Cancers 2023, 15, 765. [Google Scholar] [CrossRef]
- Rawla, P.; Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Gastroenterol. Rev./Przegląd Gastroenterol. 2019, 14, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.D.; Lordick, F.; Grabsch, H.I.; Terashima, M.; Terada, M.; Yoshikawa, T.; Boku, N.; Kataoka, K.; Smyth, E.C.; Mauer, M.; et al. Multidisciplinary management of stage II-III gastric and gastro-oesophageal junction cancer. Eur. J. Cancer 2019, 124, 67–76. [Google Scholar] [CrossRef]
- Ajani, J.A.; D’Amico, T.A.; Bentrem, D.J.; Chao, J.; Cooke, D.; Corvera, C.; Das, P.; Enzinger, P.C.; Enzler, T.; Fanta, P.; et al. Gastric cancer, version 2.2022, NCCN clinical practice guidelines in oncology. J. Natl. Compr. Cancer Netw. 2022, 20, 167–192. [Google Scholar]
- Lau, H.C.H.; Sung, J.J.-Y.; Yu, J. Gut microbiota: Impacts on gastrointestinal cancer immunotherapy. Gut Microbes 2021, 13, 1869504. [Google Scholar] [CrossRef]
- Naqash, A.R.; Kihn-Alarcón, A.J.; Stavraka, C.; Kerrigan, K.; Vareki, S.M.; Pinato, D.J.; Puri, S. The role of gut microbiome in modulating response to immune checkpoint inhibitor therapy in cancer. Ann. Transl. Med. 2021, 9, 1034. [Google Scholar] [CrossRef]
- Schupack, D.A.; Mars, R.A.T.; Voelker, D.H.; Abeykoon, J.P.; Kashyap, P.C. The promise of the gut microbiome as part of individualized treatment strategies. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 7–25. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, R.; Shohag, S.; Ahasan, T.; Sarkar, N.; Khan, H.; Hasan, A.M.; Cavalu, S.; Rauf, A. Microbiome in cancer: Role in carcinogenesis and impact in therapeutic strategies. BioMedicine 2022, 149, 112898. [Google Scholar] [CrossRef]
- Kim, N. Sex Difference of Esophageal Cancer: Esophageal Squamous Cell Carcinoma vs. Esophageal Adenocarcinoma. In Sex/Gender-Specific Medicine in the Gastrointestinal Diseases; Springer: Berlin/Heidelberg, Germany, 2022; pp. 69–92. [Google Scholar] [CrossRef]
- Arnold, M.; Soerjomataram, I.; Ferlay, J.; Forman, D. Global incidence of oesophageal cancer by histological subtype in 2012. Gut 2015, 64, 381–387. [Google Scholar] [CrossRef]
- Gregson, E.M.; Bornschein, J.; Fitzgerald, R. Genetic progression of Barrett’s oesophagus to oesophageal adenocarcinoma. Br. J. Cancer 2018, 115, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, X.; Zhuang, M.; Yuan, Z.; Nie, S.; Lu, M.; Jin, L.; Ye, W. Smoking and alcohol drinking in relation to the risk of esophageal squamous cell carcinoma: A population-based case-control study in China. Sci. Rep. 2017, 7, 17249. [Google Scholar] [CrossRef] [PubMed]
- Simba, H.; Menya, D.; Mmbaga, B.T.; Dzamalala, C.; Finch, P.; Mlombe, Y.; Mremi, A.; Narh, C.T.; Schüz, J.; McCormack, V. The contribution of smoking and smokeless tobacco to oesophageal squamous cell carcinoma risk in the African oesophageal cancer corridor: Results from the ESCCAPE multicentre case-control studies. Int. J. Cancer 2023, 152, 2269–2282. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- La Rosa, G.R.; Gattuso, G.; PEduLLà, E.; Rapisarda, E.; Nicolosi, D.; Salmeri, M. Association of oral dysbiosis with oral cancer development. Oncol. Lett. 2020, 19, 3045–3058. [Google Scholar] [CrossRef]
- Muszyński, D.; Kudra, A.; Sobocki, B.K.; Folwarski, M.; Vitale, E.; Filetti, V.; Dudzic, W.; Kaźmierczak-Siedlecka, K.; Połom, K. Esophageal cancer and bacterial part of gut microbiota—A multidisciplinary point of view. Front. Cell. Infect. Microbiol. 2022, 12, 1057668. [Google Scholar] [CrossRef]
- Yano, Y.; Etemadi, A.; Abnet, C.C. Microbiome and Cancers of the Esophagus: A Review. Microorganisms 2021, 9, 1764. [Google Scholar] [CrossRef]
- Snider, E.J.; Compres, G.; Freedberg, D.E.; Giddins, M.J.; Khiabanian, H.; Lightdale, C.J.; Nobel, Y.R.; Toussaint, N.C.; Uhlemann, A.-C.; Abrams, J.A. Barrett’s esophagus is associated with a distinct oral microbiome. Clin. Transl. Gastroenterol. 2018, 9, e135. [Google Scholar] [CrossRef]
- Li, H.; Luo, Z.; Zhang, H.; Huang, N.; Li, D.; Luo, C.; Wang, T. Characteristics of Oral Microbiota in Patients with Esophageal Cancer in China. BioMed Res. Int. 2021, 2021, 2259093. [Google Scholar] [CrossRef]
- Chen, X.; Winckler, B.; Lu, M.; Cheng, H.; Yuan, Z.; Yang, Y.; Jin, L.; Ye, W. Oral Microbiota and Risk for Esophageal Squamous Cell Carcinoma in a High-Risk Area of China. PLoS ONE 2015, 10, e0143603. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Wu, J.; Pei, Z.; Yang, L.; Purdue, M.P.; Freedman, N.D.; Jacobs, E.J.; Gapstur, S.M.; Hayes, R.B.; Ahn, J. Oral Microbiome Composition Reflects Prospective Risk for Esophageal Cancers. Cancer Res. 2017, 77, 6777–6787. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Yang, T.; Yan, Y.; Zhang, Y.; Li, Z.; Wang, Y.; Yang, J.; Xia, Y.; Xiao, H.; Han, H.; et al. Alterations of Oral Microbiota in Chinese Patients with Esophageal Cancer. Front. Cell. Infect. Microbiol. 2020, 10, 541144. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.J.; Zhang, Y.P.; Zheng, Q.Q.; Jin, H.C.; Wang, F.L.; Chen, M.; Shao, L.; Zou, D.H.; Yu, X.M.; Mao, W.M. Helicobacter pylori infection and esophageal cancer risk: An updated meta-analysis. World J. Gastroenterol. 2013, 19, 6098. [Google Scholar] [CrossRef]
- Nasrollahzadeh, D.; Malekzadeh, R.; Ploner, A.; Shakeri, R.; Sotoudeh, M.; Fahimi, S.; Nasseri-Moghaddam, S.; Kamangar, F.; Abnet, C.C.; Winckler, B.; et al. Variations of gastric corpus microbiota are associated with early esophageal squamous cell carcinoma and squamous dysplasia. Sci. Rep. 2015, 5, srep08820. [Google Scholar] [CrossRef]
- Liu, A.Q.; Vogtmann, E.; Shao, D.T.; Abnet, C.C.; Dou, H.Y.; Qin, Y.; Su, Z.; Wei, W.Q.; Chen, W. A Comparison of Biopsy and Mucosal Swab Specimens for Examining the Microbiota of Upper Gastrointestinal CarcinomaSpecimen Comparison for Upper Gastrointestinal Microbiota. Cancer Epidemiol. Biomark. Prev. 2019, 28, 2030–2037. [Google Scholar] [CrossRef]
- Lv, J.; Guo, L.; Liu, J.J.; Zhao, H.P.; Zhang, J.; Wang, J.H. Alteration of the esophageal microbiota in Barrett’s esophagus and esophageal adenocarcinoma. World J. Gastroenterol. 2019, 25, 2149. [Google Scholar] [CrossRef]
- Münch, N.S.; Fang, H.-Y.; Ingermann, J.; Maurer, H.C.; Anand, A.; Kellner, V.; Sahm, V.; Wiethaler, M.; Baumeister, T.; Wein, F.; et al. High-Fat Diet Accelerates Carcinogenesis in a Mouse Model of Barrett’s Esophagus via Interleukin 8 and Alterations to the Gut Microbiome. Gastroenterology 2019, 157, 492–506.e2. [Google Scholar] [CrossRef]
- Yang, W.; Chen, C.-H.; Jia, M.; Xing, X.; Gao, L.; Tsai, H.-T.; Zhang, Z.; Liu, Z.; Zeng, B.; Yeung, S.-C.J.; et al. Tumor-Associated Microbiota in Esophageal Squamous Cell Carcinoma. Front. Cell Dev. Biol. 2021, 9, 641270. [Google Scholar] [CrossRef]
- Deng, Y.; Tang, D.; Hou, P.; Shen, W.; Li, H.; Wang, T.; Liu, R. Dysbiosis of gut microbiota in patients with esophageal cancer. Microb. Pathog. 2021, 150, 104709. [Google Scholar] [CrossRef]
- Li, N.; Bai, C.; Zhao, L.; Sun, Z.; Ge, Y.; Li, X. The Relationship Between Gut Microbiome Features and Chemotherapy Response in Gastrointestinal Cancer. Front. Oncol. 2021, 11, 781697. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.; Okamura, A.; Kanie, Y.; Sakamoto, K.; Fujiwara, D.; Kanamori, J.; Imamura, Y.; Takeda, K.; Watanabe, M. Fecal microbes associated with the outcomes after esophagectomy in patients with esophageal cancer. Ann. Surg. Oncol. 2022, 29, 7448–7457. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Shao, D.; Zhou, J.; Gu, J.; Qin, J.; Chen, W.; Wei, W. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma. Chin. J. Cancer Res. 2020, 32, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Neto, A.G.; Whitaker, A.; Pei, Z. Microbiome and potential targets for chemoprevention of esophageal adenocarcinoma. Semin. Oncol. 2016, 43, 86–96. [Google Scholar] [CrossRef]
- Liu, N.; Ando, T.; Ishiguro, K.; Maeda, O.; Watanabe, O.; Funasaka, K.; Nakamura, M.; Miyahara, R.; Ohmiya, N.; Goto, H. Characterization of bacterial biota in the distal esophagus of Japanese patients with reflux esophagitis and Barrett’s esophagus. BMC Infect. Dis. 2013, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lu, X.; Nossa, C.W.; Francois, F.; Peek, R.M.; Pei, Z. Inflammation and Intestinal Metaplasia of the Distal Esophagus Are Associated with Alterations in the Microbiome. Gastroenterology 2009, 137, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Gupta, A.; Chauhan, R.; Bhat, A.A.; Nisar, S.; Hashem, S.; Akhtar, S.; Ahmad, A.; Haris, M.; Singh, M.; et al. Cross-talk between the microbiome and chronic inflammation in esophageal cancer: Potential driver of oncogenesis. Cancer Metastasis Rev. 2022, 41, 281–299. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, Z.; Lin, Y.; Chen, Y.; Peng, X.-E.; He, F.; Liu, S.; Yan, S.; Huang, L.; Lu, W.; et al. Streptococcus and Prevotella are associated with the prognosis of oesophageal squamous cell carcinoma. J. Med. Microbiol. 2018, 67, 1058–1068. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Severgnini, M.; Pecere, S.; Ponziani, F.R.; Boskoski, I.; Larghi, A.; Quaranta, G.; Masucci, L.; Ianiro, G.; Camboni, T.; et al. Esophageal microbiome signature in patients with Barrett’s esophagus and esophageal adenocarcinoma. PLoS ONE 2020, 15, e0231789. [Google Scholar] [CrossRef]
- Wang, Q.; Rao, Y.; Guo, X.; Liu, N.; Liu, S.; Wen, P.; Li, S.; Li, Y. Oral Microbiome in Patients with Oesophageal Squamous Cell Carcinoma. Sci. Rep. 2019, 9, 19055. [Google Scholar] [CrossRef]
- Li, D.; He, R.; Hou, G.; Ming, W.; Fan, T.; Chen, L.; Zhang, L.; Jiang, W.; Wang, W.; Lu, Z.; et al. Characterization of the Esophageal Microbiota and Prediction of the Metabolic Pathways Involved in Esophageal Cancer. Front. Cell. Infect. Microbiol. 2020, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Tang, D.; Wan, P.; Peng, Z.; Sun, M.; Guo, X.; Liu, R. Identification of tissue-specific microbial profile of esophageal squamous cell carcinoma by full-length 16S rDNA sequencing. Appl. Microbiol. Biotechnol. 2022, 106, 3215–3229. [Google Scholar] [CrossRef] [PubMed]
- Liao, O.; Ye, G.; Du, Q.; Ye, J. Gastric microbiota in gastric cancer and precancerous stages: Mechanisms of carcinogenesis and clinical value. Helicobacter 2023, 28, e12964. [Google Scholar] [CrossRef] [PubMed]
- Kupcinskas, J.; Wex, T.; Link, A.; Bartuseviciute, R.; Dedelaite, M.; Kevalaite, G.; Leja, M.; Skieceviciene, J.; Kiudelis, G.; Jonaitis, L. PSCA and MUC1 gene polymorphisms are linked with gastric cancer and pre-malignant gastric conditions. Anticancer Res. 2014, 34, 7167–7175. [Google Scholar]
- Petkevicius, V.; Salteniene, V.; Juzenas, S.; Wex, T.; Link, A.; Leja, M.; Steponaitiene, R.; Skieceviciene, J.; Kupcinskas, L.; Jonaitis, L. Polymorphisms of microRNA target genes IL12B, INSR, CCND1 and IL10 in gastric cancer. World J. Gastroenterol. 2017, 23, 3480. [Google Scholar] [CrossRef]
- Liu, X.; Shao, L.; Liu, X.; Ji, F.; Mei, Y.; Cheng, Y.; Liu, F.; Yan, C.; Li, L.; Ling, Z. Alterations of gastric mucosal microbiota across different stomach microhabitats in a cohort of 276 patients with gastric cancer. EBioMedicine 2019, 40, 336–348. [Google Scholar] [CrossRef]
- Castaño-Rodríguez, N.; Goh, K.-L.; Fock, K.M.; Mitchell, H.M.; Kaakoush, N.O. Dysbiosis of the microbiome in gastric carcinogenesis. Sci. Rep. 2017, 7, 15957. [Google Scholar] [CrossRef]
- Tseng, C.-H.; Lin, J.-T.; Ho, H.J.; Lai, Z.-L.; Wang, C.-B.; Tang, S.-L.; Wu, C.-Y. Gastric microbiota and predicted gene functions are altered after subtotal gastrectomy in patients with gastric cancer. Sci. Rep. 2016, 6, 20701. [Google Scholar] [CrossRef]
- Park, J.Y.; Seo, H.; Kang, C.S.; Shin, T.S.; Kim, J.W.; Park, J.M.; Kim, J.G.; Kim, Y.K. Dysbiotic change in gastric microbiome and its functional implication in gastric carcinogenesis. Sci. Rep. 2022, 12, 4285. [Google Scholar] [CrossRef]
- Chen, C.; Chen, L.; Lin, L.; Jin, D.; Du, Y.; Lyu, J. Research progress on gut microbiota in patients with gastric cancer, esophageal cancer, and small intestine cancer. Appl. Microbiol. Biotechnol. 2021, 105, 4415–4425. [Google Scholar] [CrossRef]
- Liang, W.; Yang, Y.; Wang, H.; Wang, H.; Yu, X.; Lu, Y.; Shen, S.; Teng, L. Gut microbiota shifts in patients with gastric cancer in perioperative period. Medicine 2019, 98, e16626. [Google Scholar] [CrossRef]
- Chen, X.-H.; Wang, A.; Chu, A.-N.; Gong, Y.-H.; Yuan, Y. Mucosa-Associated Microbiota in Gastric Cancer Tissues Compared With Non-cancer Tissues. Front. Microbiol. 2019, 10, 1261. [Google Scholar] [CrossRef]
- Gao, J.-J.; Zhang, Y.; Gerhard, M.; Mejias-Luque, R.; Zhang, L.; Vieth, M.; Ma, J.-L.; Bajbouj, M.; Suchanek, S.; Liu, W.-D.; et al. Association Between Gut Microbiota and Helicobacter pylori-Related Gastric Lesions in a High-Risk Population of Gastric Cancer. Front. Cell. Infect. Microbiol. 2018, 8, 202. [Google Scholar] [CrossRef]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y.; et al. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut 2018, 67, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.-Y.; Tung, S.-Y.; Pan, H.-Y.; Yen, C.-W.; Xu, H.-W.; Lin, Y.-J.; Deng, Y.-F.; Hsu, W.-T.; Wu, C.-S.; Li, C. Increased Abundance of Clostridium and Fusobacterium in Gastric Microbiota of Patients with Gastric Cancer in Taiwan. Sci. Rep. 2018, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, X.; Yin, J.; Li, Y.; Hou, B.; Zhang, Z. A screening method for gastric cancer by oral microbiome detection. Oncol. Rep. 2018, 39, 2217–2224. [Google Scholar] [CrossRef]
- Wu, Z.-F.; Zou, K.; Wu, G.-N.; Jin, Z.-J.; Xiang, C.-J.; Xu, S.; Wang, Y.-H.; Wu, X.-Y.; Chen, C.; Xu, Z.; et al. A Comparison of Tumor-Associated and Non-Tumor-Associated Gastric Microbiota in Gastric Cancer Patients. Dig. Dis. Sci. 2021, 66, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ng, S.-K.; Ding, Y.; Lin, Y.; Liu, W.; Wong, S.H.; Sung, J.J.-Y.; Yu, J. Meta-analysis of mucosal microbiota reveals universal microbial signatures and dysbiosis in gastric carcinogenesis. Oncogene 2022, 41, 3599–3610. [Google Scholar] [CrossRef] [PubMed]
- Kamboj, K.A.; Cotter, T.G.; Oxentenko, A.S. Helicobacter pylori: The past, present, and future in management. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Salvatori, S.; Marafini, I.; Laudisi, F.; Monteleone, G.; Stolfi, C. Helicobacter pylori and Gastric Cancer: Pathogenetic Mechanisms. Int. J. Mol. Sci. 2023, 24, 2895. [Google Scholar] [CrossRef]
- Muhammad, J.S.; Manzoor, S.; Cui, Z.-G.; Khoder, G. DNA Methylation-Mediated Overexpression of CXCL1 in Helicobacter pylori-Induced Gastric Cancer: In Silico- and In Vitro-Based Identification of a Potential Biomarker for Carcinogenesis. Int. J. Mol. Sci. 2023, 24, 795. [Google Scholar] [CrossRef] [PubMed]
- Correa, P.; Haenszel, W.; Cuello, C.; Tannenbaum, S.; Archer, M. A model for gastric cancer epidemiology. Lancet 1975, 306, 58–60. [Google Scholar] [CrossRef]
- Polk, D.B.; Peek, R.M., Jr. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Takahashi, A.; Suzuki, K.; Kurusu-Kanno, M.; Yamaguchi, K.; Fujiki, H.; Suganuma, M. Epithelial–mesenchymal transition in human gastric cancer cell lines induced by TNF-α-inducing protein of Helicobacter pylori. Int. J. Cancer 2014, 134, 2373–2382. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Sajib, S.; Zahra, F.T.; Lionakis, M.S.; German, N.A.; Mikelis, C.M. Mechanisms of angiogenesis in microbe-regulated inflammatory and neoplastic conditions. Angiogenesis 2018, 21, 1–14. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, Y.; Li, H.; Shao, S. Helicobacter pylori promotes gastric cancer progression through the tumor microenvironment. Appl. Microbiol. Biotechnol. 2022, 106, 4375–4385. [Google Scholar] [CrossRef]
- Suzuki, M.; Mimuro, H.; Kiga, K.; Fukumatsu, M.; Ishijima, N.; Morikawa, H.; Nagai, S.; Koyasu, S.; Gilman, R.H.; Kersulyte, D.; et al. Helicobacter pylori CagA Phosphorylation-Independent Function in Epithelial Proliferation and Inflammation. Cell Host Microbe 2009, 5, 23–34. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, L.; Liang, X.; Li, S.; Ma, L.; Zheng, L.; Li, T.; Yu, H.; Chan, H.; Chen, C.; et al. Helicobacter pylori-induced YAP1 nuclear translocation promotes gastric carcinogenesis by enhancing IL-1β expression. Cancer Med. 2019, 8, 3965–3980. [Google Scholar] [CrossRef]
- Mayerle, J.; den Hoed, C.M.; Schurmann, C.; Stolk, L.; Homuth, G.; Peters, M.J.; Capelle, L.G.; Zimmermann, K.; Rivadeneira, F.; Gruska, S.; et al. Identification of Genetic Loci Associated with Helicobacter pylori Serologic Status. JAMA 2013, 309, 1912–1920. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef]
- Guo, Y.; Cao, X.-S.; Zhou, M.-G.; Yu, B. Gastric microbiota in gastric cancer: Different roles of Helicobacter pylori and other microbes. Front. Cell. Infect. Microbiol. 2023, 12, 1975. [Google Scholar] [CrossRef] [PubMed]
- Gunathilake, M.; Lee, J.; Choi, I.J.; Kim, Y.-I.; Kim, J. Effect of the Interaction between Dietary Patterns and the Gastric Microbiome on the Risk of Gastric Cancer. Nutrients 2021, 13, 2692. [Google Scholar] [CrossRef]
- Wu, B.; Yang, D.; Yang, S.; Zhang, G. Dietary Salt Intake and Gastric Cancer Risk: A Systematic Review and Meta-Analysis. Front. Nutr. 2021, 8, 801228. [Google Scholar] [CrossRef]
- Odhong’, C.; Wilkes, A.; van Dijk, S.; Vorlaufer, M.; Ndonga, S.; Sing’ora, B.; Kenyanito, L. Financing large-scale mitigation by smallholder farmers: What roles for public climate finance? Front. Sustain. Food Syst. 2019, 3, 3. [Google Scholar] [CrossRef]
- Arita, S.; Inagaki-Ohara, K. High-fat-diet–induced modulations of leptin signaling and gastric microbiota drive precancerous lesions in the stomach. Nutrition 2019, 67–68, 110556. [Google Scholar] [CrossRef]
- Faubert, B.; Li, K.Y.; Cai, L.; Hensley, C.T.; Kim, J.; Zacharias, L.G.; Yang, C.; Do, Q.N.; Doucette, S.; Burguete, D.; et al. Lactate Metabolism in Human Lung Tumors. Cell 2017, 171, 358–371.e9. [Google Scholar] [CrossRef] [PubMed]
- San-Millán, I.; Brooks, G.A. Reexamining cancer metabolism: Lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis 2017, 38, 119–133. [Google Scholar] [CrossRef]
- Wang, C.; Song, X.; Han, Z.; Li, X.; Xu, Y.; Xiao, Y. Monitoring Nitric Oxide in Subcellular Compartments by Hybrid Probe Based on Rhodamine Spirolactam and SNAP-tag. ACS Chem. Biol. 2016, 11, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Tsugane, S.; Sasazuki, S. Diet and the risk of gastric cancer: Review of epidemiological evidence. Gastric Cancer 2007, 10, 75–83. [Google Scholar] [CrossRef]
- Wagner, W.; Ciszewski, W.M.; Kania, K.D. L- and D-lactate enhance DNA repair and modulate the resistance of cervical carcinoma cells to anticancer drugs via histone deacetylase inhibition and hydroxycarboxylic acid receptor 1 activation. Cell Commun. Signal. 2015, 13, 36. [Google Scholar] [CrossRef]
- Brand, A.; Singer, K.; Koehl, G.E.; Kolitzus, M.; Schoenhammer, G.; Thiel, A.; Matos, C.; Bruss, C.; Klobuch, S.; Peter, K.; et al. LDHA-Associated Lactic Acid Production Blunts Tumor Immunosurveillance by T and NK Cells. Cell Metab. 2016, 24, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Vinasco, K.; Mitchell, H.M.; Kaakoush, N.O.; Castaño-Rodríguez, N. Microbial carcinogenesis: Lactic acid bacteria in gastric cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2019, 1872, 188309. [Google Scholar] [CrossRef] [PubMed]
- Yee, M.; Kim, S.; Sethi, P.; Düzgüneş, N.; Konopka, K. Porphyromonas gingivalis stimulates IL-6 and IL-8 secretion in GMSM-K, HSC-3 and H413 oral epithelial cells. Anaerobe 2014, 28, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, H.; Duan, X.; Jotwani, R.; Vuddaraju, H.; Liang, S.; Scott, D.A.; Lamont, R.J. Porphyromonas gingivalis-Induced Reactive Oxygen Species Activate JAK2 and Regulate Production of Inflammatory Cytokines through c-Jun. Infect. Immun. 2014, 82, 4118–4126. [Google Scholar] [CrossRef]
- Wang, H.; Kumar, A.; Lamont, R.J.; Scott, D.A. GSK3β and the control of infectious bacterial diseases. Trends Microbiol. 2014, 22, 208–217. [Google Scholar] [CrossRef]
- Chen, M.-F.; Lu, M.-S.; Hsieh, C.-C.; Chen, W.-C. Porphyromonas gingivalis promotes tumor progression in esophageal squamous cell carcinoma. Cell. Oncol. 2021, 44, 373–384. [Google Scholar] [CrossRef]
- Gao, S.; Li, S.; Ma, Z.; Liang, S.; Shan, T.; Zhang, M.; Zhu, X.; Zhang, P.; Liu, G.; Zhou, F.; et al. Presence of Porphyromonas gingivalis in esophagus and its association with the clinicopathological characteristics and survival in patients with esophageal cancer. Infect. Agents Cancer 2016, 11, 3. [Google Scholar] [CrossRef]
- Yilmaz, Ö.; Yao, L.; Maeda, K.; Rose, T.M.; Lewis, E.L.; Duman, M.; Lamont, R.J.; Ojcius, D.M. ATP scavenging by the intracellular pathogen Porphyromonas gingivalis inhibits P2X7-mediated host-cell apoptosis. Cell. Microbiol. 2008, 10, 863–875. [Google Scholar] [CrossRef]
- Morandini, A.C.; Ramos-Junior, E.S.; Potempa, J.; Nguyen, K.-A.; Oliveira, A.C.; Bellio, M.; Ojcius, D.M.; Scharfstein, J.; Coutinho-Silva, R. Porphyromonas gingivalis Fimbriae Dampen P2X7-Dependent Interleukin-1β Secretion. J. Innate Immun. 2014, 6, 831–845. [Google Scholar] [CrossRef]
- Kuboniwa, M.; Hasegawa, Y.; Mao, S.; Shizukuishi, S.; Amano, A.; Lamont, R.J.; Yilmaz, O. P. gingivalis accelerates gingival epithelial cell progression through the cell cycle. Microbes Infect. 2008, 10, 122–128. [Google Scholar] [CrossRef]
- Yao, Á.; Jermanus, C.; Barbetta, B.; Choi, C.; Verbeke, P.; Ojcius, D.M.; Yilmaz, Ö. Porphyromonas gingivalis infection sequesters pro-apoptotic Bad through Akt in primary gingival epithelial cells. Mol. Oral Microbiol. 2010, 25, 89–101. [Google Scholar] [CrossRef]
- Whitmore, S.E.; Lamont, R.J. Oral Bacteria and Cancer. PLoS Pathog. 2014, 10, e1003933. [Google Scholar] [CrossRef]
- Salaspuro, M.P. Acetaldehyde, microbes, and cancer of the digestive tract. Crit. Rev. Clin. Lab. Sci. 2003, 40, 183–208. [Google Scholar] [CrossRef]
- Man, S.M.; Kaakoush, N.O.; Leach, S.T.; Nahidi, L.; Lu, H.K.; Norman, J.; Day, A.S.; Zhang, L.; Mitchell, H.M. Host attachment, invasion, and stimulation of proinflammatory cytokines by Campylobacter concisus and other Non-Campylobacter jejuni Campylobacter species. J. Infect. Dis. 2010, 202, 1855–1865. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Deshpande, N.P.; Man, S.M.; Burgos-Portugal, J.A.; Khattak, F.A.; Raftery, M.J.; Wilkins, M.R.; Mitchell, H.M. Transcriptomic and Proteomic Analyses Reveal Key Innate Immune Signatures in the Host Response to the Gastrointestinal Pathogen Campylobacter concisus. Infect. Immun. 2015, 83, 832–845. [Google Scholar] [CrossRef]
- Asili, P.; Mirahmad, M.; Rezaei, P.; Mahdavi, M.; Larijani, B.; Tavangar, S.M. The Association of Oral Microbiome Dysbiosis with Gastrointestinal Cancers and Its Diagnostic Efficacy. J. Gastrointest. Cancer 2023, 1–20. [Google Scholar] [CrossRef]
- Blackett, K.L.; Siddhi, S.S.; Cleary, S.; Steed, H.; Miller, M.H.; Macfarlane, S.; Macfarlane, G.T.; Dillon, J.F. Oesophageal bacterial biofilm changes in gastro-oesophageal reflux disease, Barrett’s and oesophageal carcinoma: Association or causality? Aliment. Pharmacol. Ther. 2013, 37, 1084–1092. [Google Scholar] [CrossRef]
- Nasr, R.; Shamseddine, A.; Mukherji, D.; Nassar, F.; Temraz, S. The Crosstalk between Microbiome and Immune Response in Gastric Cancer. Int. J. Mol. Sci. 2020, 21, 6586. [Google Scholar] [CrossRef]
- Rizzello, V.; Bonaccorsi, I.; Dongarrà, M.L.; Fink, L.N.; Ferlazzo, G. Role of Natural Killer and Dendritic Cell Crosstalk in Immunomodulation by Commensal Bacteria Probiotics. J. Biomed. Biotechnol. 2011, 2011, 473097. [Google Scholar] [CrossRef]
- Wakasugi, H.; Oshimi, K.; Miyata, M.; Morioka, Y. Augmentation of natural killer (NK) cell activity by a streptococcal preparation, OK-432, in patients with malignant tumors. J. Clin. Immunol. 1981, 1, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Nie, S.; Wang, A.; Yuan, Y. Comparison of clinicopathological parameters, prognosis, micro-ecological environment and metabolic function of Gastric Cancer with or without Fusobacterium sp. Infection. J. Cancer 2021, 12, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Vickers, N.J. Animal communication: When i’m calling you, will you answer too? Curr. Biol. 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 Protein of Fusobacterium nucleatum to Human Inhibitory Receptor TIGIT Protects Tumors from Immune Cell Attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef]
- Peng, R.; Liu, S.; You, W.; Huang, Y.; Hu, C.; Gao, Y.; Jia, X.; Li, G.; Xu, Z.; Chen, Y. Gastric Microbiome Alterations Are Associated with Decreased CD8+ Tissue-Resident Memory T Cells in the Tumor Microenvironment of Gastric Cancer. Cancer Immunol. Res. 2022, 10, 1224–1240. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, N.; Su, X.; Gao, Y.; Yang, R. Gut-Microbiota-Derived Metabolites Maintain Gut and Systemic Immune Homeostasis. Cells 2023, 12, 793. [Google Scholar] [CrossRef]
- Uribe-Herranz, M.; Beghi, S.; Ruella, M.; Parvathaneni, K.; Salaris, S.; Kostopoulos, N.; George, S.S.; Pierini, S.; Krimitza, E.; Costabile, F.; et al. Modulation of the gut microbiota engages antigen cross-presentation to enhance antitumor effects of CAR T cell immunotherapy. Mol. Ther. 2023, 31, 686–700. [Google Scholar] [CrossRef]
- Dzutsev, A.; Goldszmid, R.S.; Viaud, S.; Zitvogel, L.; Trinchieri, G. The role of the microbiota in inflammation, carcinogenesis, and cancer therapy. Eur. J. Immunol. 2015, 45, 17–31. [Google Scholar] [CrossRef]
- Ahlawat, S.; Kumar, P.; Mohan, H.; Goyal, S.; Sharma, K.K. Inflammatory bowel disease: Tri-directional relationship between microbiota, immune system and intestinal epithelium. Crit. Rev. Microbiol. 2021, 47, 254–273. [Google Scholar] [CrossRef]
- Ervin, S.M. Structure, Function, and Inhibition of Gut Microbial Enzymes. Ph.D. Thesis, The University of North, Chapel Hill, NC, USA, 2020. [Google Scholar]
- Mayorgas, A.; Dotti, I.; Salas, A. Microbial metabolites, postbiotics, and intestinal epithelial function. Mol. Nutr. Food Res. 2021, 65, 2000188. [Google Scholar] [CrossRef]
- Peng, G.; Fadeel, B. Understanding the bidirectional interactions between two-dimensional materials, microorganisms, and the immune system. Adv. Drug Deliv. Rev. 2022, 188, 114422. [Google Scholar] [CrossRef]
- Kojima, T.; Muro, K.; Francois, E.; Hsu, C.-H.; Moriwaki, T.; Kim, S.-B.; Lee, S.-H.; Bennouna, J.; Kato, K.; Lin, S.; et al. Pembrolizumab versus chemotherapy as second-line therapy for advanced esophageal cancer: Phase III KEYNOTE-181 study. J. Clin. Oncol. 2019, 37, 4138–4148. [Google Scholar] [CrossRef]
- Park, R.; Da Silva, L.L.; Saeed, A. Immunotherapy Predictive Molecular Markers in Advanced Gastroesophageal Cancer: MSI and Beyond. Cancers 2021, 13, 1715. [Google Scholar] [CrossRef] [PubMed]
- Shitara, K.; Ajani, J.A.; Moehler, M.; Garrido, M.; Gallardo, C.; Shen, L.; Yamaguchi, K.; Wyrwicz, L.; Skoczylas, T.; Bragagnoli, A.C.; et al. Nivolumab plus chemotherapy or ipilimumab in gastro-oesophageal cancer. Nature 2022, 603, 942–948. [Google Scholar] [CrossRef]
- Kato, K.; Shah, M.A.; Enzinger, P.; Bennouna, J.; Shen, L.; Adenis, A.; Sun, J.-M.; Cho, B.C.; Özgüroğlu, M.; Kojima, T.; et al. KEYNOTE-590: Phase III study of first-line chemotherapy with or without pembrolizumab for advanced esophageal cancer. Future Oncol. 2019, 15, 1057–1066. [Google Scholar] [CrossRef]
- Janjigian, Y.Y.; Shitara, K.; Moehler, M.; Garrido, M.; Salman, P.; Shen, L.; Wyrwicz, L.; Yamaguchi, K.; Skoczylas, T.; Bragagnoli, A.C.; et al. First-line nivolumab plus chemotherapy versus chemotherapy alone for advanced gastric, gastro-oesophageal junction, and oesophageal adenocarcinoma (CheckMate 649): A randomised, open-label, phase 3 trial. Lancet 2021, 3, 27–40. [Google Scholar] [CrossRef]
- Doki, Y.; Ajani, J.A.; Kato, K.; Xu, J.; Wyrwicz, L.; Motoyama, S.; Ogata, T.; Kawakami, H.; Hsu, C.-H.; Adenis, A.; et al. Nivolumab Combination Therapy in Advanced Esophageal Squamous-Cell Carcinoma. N. Engl. J. Med. 2022, 386, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Takei, S.; Kawazoe, A.; Shitara, K. The New Era of Immunotherapy in Gastric Cancer. Cancers 2022, 14, 1054. [Google Scholar] [CrossRef]
- Zeng, Z.; Yang, B.; Liao, Z. Biomarkers in Immunotherapy-Based Precision Treatments of Digestive System Tumors. Front. Oncol. 2021, 11, 650481. [Google Scholar] [CrossRef]
- Kahraman, S.; Yalcin, S. Recent Advances in Systemic Treatments for HER-2 Positive Advanced Gastric Cancer. OncoTargets Ther. 2021, 14, 4149–4162. [Google Scholar] [CrossRef]
- Andre, T.; Berton, D.; Curigliano, G.; Ellard, S.; Pérez, J.M.T.; Arkenau, H.-T.; Abdeddaim, C.; Moreno, V.; Guo, W.; Im, E.; et al. Safety and efficacy of anti–PD-1 antibody dostarlimab in patients (pts) with mismatch repair-deficient (dMMR) solid cancers: Results from GARNET study. J. Clin. Oncol. 2021, 39, 9. [Google Scholar] [CrossRef]
- Li, S.; Yu, W.; Xie, F.; Luo, H.; Liu, Z.; Lv, W.; Shi, D.; Yu, D.; Gao, P.; Chen, C.; et al. Neoadjuvant therapy with immune checkpoint blockade, antiangiogenesis, and chemotherapy for locally advanced gastric cancer. Nat. Commun. 2023, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Aisa, A.; Weng, S.; Li, X.; Zhang, D.; Yuan, Y. Immune checkpoint inhibitors combined with HER-2 targeted therapy in HER-2 positive gastroesophageal cancer. Crit. Rev. Oncol. 2022, 180, 103864. [Google Scholar] [CrossRef] [PubMed]
- Liatsos, C.; Papaefthymiou, A.; Kyriakos, N.; Galanopoulos, M.; Doulberis, M.; Giakoumis, M.; Petridou, E.; Mavrogiannis, C.; Rokkas, T.; Kountouras, J. Helicobacter pylori, gastric microbiota and gastric cancer relationship: Unrolling the tangle. World J. Gastrointest. Oncol. 2022, 14, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Sundar, R.; Smyth, E.C.; Peng, S.; Yeong, J.P.S.; Tan, P. Predictive Biomarkers of Immune Checkpoint Inhibition in Gastroesophageal Cancers. Front. Oncol. 2020, 10, 763. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Chen, F. Common strategies for effective immunotherapy of gastroesophageal cancers using immune checkpoint inhibitors. Pathol.-Res. Pract. 2022, 238, 154110. [Google Scholar] [CrossRef]
- Yoon, H.H.; Jin, Z.; Kour, O.; Fonkoua, L.A.; Shitara, K.; Gibson, M.K.; Prokop, L.J.; Moehler, M.; Kang, Y.K.; Shi, Q.; et al. Association of PD-L1 expression and other variables with benefit from immune checkpoint inhibition in advanced gastroesophageal cancer: Systematic review and meta-analysis of 17 phase 3 randomized clinical trials. JAMA Oncol. 2022, 8, 1456–1465. [Google Scholar] [CrossRef]
- Nagata, Y.; Yamamoto, S.; Kato, K. Immune checkpoint inhibitors in esophageal cancer: Clinical development and perspectives. Hum. Vaccines Immunother. 2022, 18, 2143177. [Google Scholar] [CrossRef]
- Hooper, L.V.; Macpherson, A.J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat. Rev. Immunol. 2010, 10, 159–169. [Google Scholar] [CrossRef]
- Jiang, H.; Ye, Y.; Wang, M.; Sun, X.; Sun, T.; Chen, Y.; Li, P.; Zhang, M.; Wang, T. The progress on the relationship between gut microbiota and immune checkpoint blockade in tumors. Biotechnol. Genet. Eng. Rev. 2023, 1–20. [Google Scholar] [CrossRef]
- Ting, N.L.-N.; Yu, J. Microbiota in Cancer Immunotherapy: The Next Milestone of Immuno-oncology. In Microbiome in Gastrointestinal Cancer; Springer: Berlin/Heidelberg, Germany, 2023; pp. 269–287. [Google Scholar] [CrossRef]
- Rezasoltani, S.; Yadegar, A.; Aghdaei, H.A.; Zali, M.R. Modulatory effects of gut microbiome in cancer immunotherapy: A novel paradigm for blockade of immune checkpoint inhibitors. Cancer Med. 2021, 10, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Hsu, R.; Hacein-Bey, C.; Zhang, W.; Gao, L.; Kurth, M.J.; Zhao, H.; Shuai, Z.; Leung, P.S.C. The Evolving Landscape of Fecal Microbial Transplantation. Clin. Rev. Allergy Immunol. 2023, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Chyuan, I.-T.; Chu, C.-L.; Hsu, P.-N. Targeting the Tumor Microenvironment for Improving Therapeutic Effectiveness in Cancer Immunotherapy: Focusing on Immune Checkpoint Inhibitors and Combination Therapies. Cancers 2021, 13, 1188. [Google Scholar] [CrossRef] [PubMed]
- Hayase, E.; Jenq, R.R. Role of the intestinal microbiome and microbial-derived metabolites in immune checkpoint blockade immunotherapy of cancer. Genome Med. 2021, 13, 107. [Google Scholar] [CrossRef] [PubMed]
- de Clercq, N.C.; van den Ende, T.; Prodan, A.; Hemke, R.; Davids, M.; Pedersen, H.K.; Nielsen, H.B.; Groen, A.K.; de Vos, W.M.; van Laarhoven, H.W.; et al. Fecal Microbiota Transplantation from Overweight or Obese Donors in Cachectic Patients with Advanced Gastroesophageal Cancer: A Randomized, Double-blind, Placebo-Controlled, Phase II StudyFecal Microbiota Transplantation for Gastroesophageal Cancer. Clin. Cancer Res. 2021, 27, 3784–3792. [Google Scholar] [CrossRef]
- Samanta, S. Potential impacts of prebiotics and probiotics on cancer prevention. Anti-Cancer Agents Med. Chem. (Formerly Curr. Med. Chem.-Anti-Cancer Agents) 2022, 22, 605–628. [Google Scholar] [CrossRef]
- Hong, J.; Fang, J.-Y. Gut Microbiota Impacts on the Efficacy of Anticancer Treatment of Colorectal Cancer. In Microbiome in Gastrointestinal Cancer; Springer: Berlin/Heidelberg, Germany, 2023; pp. 237–249. [Google Scholar] [CrossRef]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Cramer, P.; Bresalier, R.S. Gastrointestinal and Hepatic Complications of Immune Checkpoint Inhibitors. Curr. Gastroenterol. Rep. 2017, 19, 3. [Google Scholar] [CrossRef]
- Rani, V.; Singhal, S.; Sharma, K.; Vaid, R.; Aggarwal, K.; Bhadana, R.; Agarwal, R.; Atale, N. Human Gut Microbiome: A New Frontier in Cancer Diagnostics & Therapeutics. Curr. Pharm. Des. 2021, 27, 4578–4592. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, J.; Liu, T.; Xing, J.; Zhang, H.; Wang, D.; Tang, D. Bidirectional effects of intestinal microbiota and antibiotics: A new strategy for colorectal cancer treatment and prevention. J. Cancer Res. Clin. Oncol. 2022, 148, 2387–2404. [Google Scholar] [CrossRef]
- Li, X.; Zhang, S.; Guo, G.; Han, J.; Yu, J. Gut microbiome in modulating immune checkpoint inhibitors. EBioMedicine 2022, 82, 104163. [Google Scholar] [CrossRef]
- Deng, R.; Zheng, H.; Cai, H.; Li, M.; Shi, Y.; Ding, S. Effects of helicobacter pylori on tumor microenvironment and immunotherapy responses. Front. Immunol. 2022, 13, 923477. [Google Scholar] [CrossRef]
- Shi, Y.; Zheng, H.; Guo, Z.; Deng, R.; Yu, W.; Song, Y.; Ding, S. Effect of Helicobacter pylori on immunotherapy is gaining more attention. Helicobacter 2022, 27, e12925. [Google Scholar] [CrossRef] [PubMed]
- Che, H.; Xiong, Q.; Ma, J.; Chen, S.; Wu, H.; Xu, H.; Hou, B. Association of Helicobacter pylori infection with survival outcomes in advanced gastric cancer patients treated with immune checkpoint inhibitors. BMC Cancer 2022, 22, 904. [Google Scholar] [CrossRef] [PubMed]
- Sunakawa, Y.; Matoba, R.; Inoue, E.; Sakamoto, Y.; Kawabata, R.; Ishiguro, A.; Akamaru, Y.; Kito, Y.; Takahashi, M.; Matsuyama, J.; et al. Genomic pathway of gut microbiome to predict efficacy of nivolumab in advanced gastric cancer: DELIVER trial (JACCRO GC-08). J. Clin. Oncol. 2021, 39, 161. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, S.; Zou, H.; Han, Z.; Xie, T.; Zhang, B.; Dai, D.; Yin, X.; Liang, Y.; Kou, Y.; et al. Correlation of the gut microbiome and immune-related adverse events in gastrointestinal cancer patients treated with immune checkpoint inhibitors. Front. Cell. Infect. Microbiol. 2023, 13, 1099063. [Google Scholar] [CrossRef]
- Song, L.; Lu, L.; Pu, Y.; Yin, H.; Zhang, K. Nanomaterials-Based Tumor Microenvironment Modulation for Magnifying Sonodynamic Therapy. Accounts Mater. Res. 2022, 3, 971–985. [Google Scholar] [CrossRef]
- Hu, M.; Huang, L. Strategies targeting tumor immune and stromal microenvironment and their clinical relevance. Adv. Drug Deliv. Rev. 2022, 183, 114137. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name of the Trial | Disease Site | Trial Phase | Arms of the Study | Primary Endpoint | Results |
|---|---|---|---|---|---|
| KEYNOTE-590 | Advanced Esophageal or GEJ SCC | Phase III | Chemo + Pembro vs. Chemo + Placebo | OS, PFS | OS: 13.9 vs. 8.8 months |
| CHECKMATE-648 | Advanced Esophageal or GEJ SCC | Phase III | Chemo + Nivo vs. Ipi + Nivo vs. Chemo alone | OS, PFS | OS: 13.2 vs. 12.7 vs. 10.7 months |
| CHECKMATE-649 | Advanced Esophageal or GEJ AC | Phase III | Chemo + Nivo vs. Ipi + Nivo vs. Chemo alone | OS, PFS | OS: 13.1 vs. 11.1 months |
| ATTRACTION-3 | Advanced Esophageal or GEJ SCC | Phase III | Nivo vs. Chemo | OS | OS: 10.9 vs. 8.4 months |
| KEYNOTE-811 | Advanced gastric or GEJ AC, Her2 positive | Phase III | (Chemo + Trastuzumab) + Pembro vs. Placebo | OS, PFS | Study ongoing |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dadgar, N.; Edlukudige Keshava, V.; Raj, M.S.; Wagner, P.L. The Influence of the Microbiome on Immunotherapy for Gastroesophageal Cancer. Cancers 2023, 15, 4426. https://doi.org/10.3390/cancers15184426
Dadgar N, Edlukudige Keshava V, Raj MS, Wagner PL. The Influence of the Microbiome on Immunotherapy for Gastroesophageal Cancer. Cancers. 2023; 15(18):4426. https://doi.org/10.3390/cancers15184426
Chicago/Turabian StyleDadgar, Neda, Vinay Edlukudige Keshava, Moses S. Raj, and Patrick L. Wagner. 2023. "The Influence of the Microbiome on Immunotherapy for Gastroesophageal Cancer" Cancers 15, no. 18: 4426. https://doi.org/10.3390/cancers15184426