1. Introduction
Laryngeal cancer accounts for approximately one-third of all head and neck cancers. The American Cancer Society estimates men to be four times more likely to be diagnosed with laryngeal cancer and die from it compared to women in the United States [
1]. Approximately 60% of patients are diagnosed at advanced stages (stage III or IV), at which point surgery with adjuvant chemotherapy or radiotherapy is predominantly used [
1,
2]. The constant use of aggressive treatments stresses the need for further research into alternative therapies and early-stage biomarkers.
Previous epidemiological studies, along with the significant difference in sex susceptibility in laryngeal cancer, have raised questions about the role of sex hormones in malignancy. At the same time, many studies have shown that smoking and alcoholism are some of the most common contributing factors to laryngeal cancer [
3]. However, global incidence was demonstrated to be seven times higher in men compared to women regardless of alcohol or tobacco usage [
4,
5]. The evident gender dispositions found in the larynx, physiological changes observed during puberty, and the organ characterized as a secondary sex organ imply that sex hormones or endocrine factors could play a significant role in the tumorigenesis of laryngeal cancer [
3,
6]. However, it is still widely debated as a sex hormone-dependent cancer.
Researchers observed the potential role of estrogen in laryngeal cancer when examining the change in estrogen levels in men over the age of 60, correlating to the age at which men have an increased risk of incidence [
7]. Other studies demonstrated a higher probability of abnormal estrogen metabolism in a cohort with 60% laryngeal cancer patients compared to healthy patients [
8]. The presence of functional estrogen receptors in laryngeal cancer and its estrogen dependence have been contested for years [
9,
10,
11]. They were first described in laryngeal cancer over 30 years ago, when studies showed an increase in the reactivity of sex hormone receptors, specifically ERα, in malignant laryngeal tissue compared to adjacent healthy tissue [
6,
12,
13], and many studies have shown that laryngeal cancers respond to estrogen [
14,
15,
16]. The controversy involving laryngeal cancer as an estrogen-responsive cancer could be due to the presence or absence of ERα66 and/or its splice variants ERα36 and ERα46.
The role of estrogen in tumorigenesis is well established in hormone-responsive cancers like breast and prostate cancers [
17]. Estrogen regulates proliferation, survivability, and metastasis via its receptor, estrogen receptor alpha-66 (ERα66). As a result, ERα66 is currently used as a critical molecular biomarker in diagnostics, predicting prognoses and influencing treatment [
1,
16,
18].
ERα66 has been thoroughly investigated in breast cancer, demonstrating its role in mediating the effects of estrogen and specific hormone therapies on tumorigenesis [
17,
19]. Most commonly, clinicians use the presence of ERα66 in breast cancer as a well-established prognostic factor for aggression and responsiveness to endocrine therapy [
9,
20]. It is also a common target for hormone therapy treatments like tamoxifen [
14]. On the other hand, ERα66-negative breast cancers are more aggressive and tend to be more resistant to hormone therapy treatment and, therefore, require other treatment modalities.
Recently, studies have investigated the role of ERα66 splice variants in mediating breast cancer response to estrogen [
15,
17,
19]. ERα36 contains ligand binding domains similar to ERα66 but does not include the transcriptionally active binding domains AF-1 and AF-2 [
19,
21,
22]. It is located in the plasma membrane and is expressed in both ERα66-positive and ERα66-negative breast cancers. Estrogen binding to ERα36 induces phospholipase C (PLC)-dependent protein kinase C (PKC) signaling through a caveolae membrane-mediated mechanism to enhance metastasis and regulate tamoxifen resistance [
21,
22,
23]. ERα46 is a lesser studied isoform due to its similarity in structure to ERα66, but it does not include the AF-1 transactivation domain [
19,
22,
24]. Studies suggest that its similar structure contributes to its function in that it may act as a competitive inhibitor of ERα66, but more studies are needed to confirm this hypothesis [
22,
24].
Similar to breast cancers [
15,
17,
19], the presence of ERα66 in laryngeal cancers can determine the tumor’s aggressiveness and how the cells will respond to treatments with estrogen [
16,
25]. In a previous study, ERα66-positive UM-SCC-12 cells and ERα66-negative UM-SCC-11A cells, which came from male donors, exhibited differential responsiveness to 17β-estradiol; the resulting changes in tumor aggressiveness were dependent on the expression of ERα66 [
16,
25]. In human laryngeal tissue samples, high levels of ERα66 were measured in samples of earlier stage tumors, whereas stage IV tumor samples exhibited the lowest ERα66 levels. There was no correlation between staging and ERα36 expression, even though the receptor was shown to play a role in estrogen-mediated tumorigenesis and chemoresistance [
15,
25]. These data suggest that ERα66 plays a critical role in tumor aggression, particularly in early-stage cancers.
In vitro studies using ER-positive UM-SCC-12 cells and ER-negative UM-SCC-11A cells support this hypothesis [
16,
25]. 17β-estradiol increased proliferation in UM-SCC-12 cells but reduced proliferation in UM-SCC-11A cells. Treatment inhibited apoptosis in UM-SCC-12 cells, but there was no change in apoptosis in UM-SCC-11A cells. These data demonstrate the variability in response to estrogen in laryngeal cancer cells and suggest that this may be due at least in part to the difference in ERα66 expression [
16]. Therefore, ERα66 may be a promising prognostic and diagnostic biomarker in laryngeal cancer similar to the findings in breast cancer.
The seco-steroid vitamin D
3 also impacts tumor growth and metastasis. Vitamin D
3 deficiency is associated with an increased risk of malignancy in multiple cancers [
26,
27,
28]. In addition, vitamin D
3 supplementation (25(OH)D
3) is used as a therapeutic agent in various cancers, like breast, prostate, colorectal, and ovarian cancer. Evidence shows that vitamin D
3 can affect tumorigenesis by regulating the innate and adaptive immune systems [
28]. Vitamin D deficiency could explain the racial disparities in laryngeal cancer, suggesting that vitamin D
3 could also be a risk factor for early-age diagnosis. African American men are at a higher risk of diagnosis at a younger age and death than Caucasian men [
2]. Randomized trials demonstrated that despite similar smoking rates compared to Caucasian men, a common risk factor for laryngeal cancer, African American men were at the highest risk of vitamin D
3 deficiency [
29].
These studies suggest that vitamin D
3 supplementation may be a promising therapeutic agent in laryngeal cancer treatment. Whereas many reports demonstrate that vitamin D deficiency is not a risk factor for laryngeal cancer [
30,
31], other reports show that high levels of circulating 25(OH)D
3 are associated with a lower incidence of malignancy [
32]. Several factors may contribute to this difference, including hormone receptor expression, seasonal measurements, downstream vitamin D
3 metabolism, and localized metabolite production in the tissue [
31]. These factors need to be assessed to determine the therapeutic potential of vitamin D
3 in laryngeal cancer.
Previous studies in breast cancer showed that the vitamin D
3 metabolite, 24R,25(OH)
2D
3, can regulate tumorigenesis without destabilizing serum calcium levels, unlike 1α,25(OH)
2D
3. The effect of 24R,25(OH)
2D
3 on tumorigenesis depended on the expression of ERα66. 24R,25(OH)
2D
3 inhibited tumorigenesis in ERα66-positive MCF7 cells but increased tumorigenesis in the ERα66-negative HCC38 cell line [
33]. We examined the potential for autocrine-based effects causing the difference in response by evaluating the expression of hydroxylase enzymes and the production of vitamin D
3 metabolites. The expression of CYP27B1, the hydroxylase that is responsible for converting 25(OH)D
3 into 1,25(OH)
2D
3, in healthy tissue indicated that breast cancer cells could locally produce metabolites of vitamin D
3, suggesting that vitamin D
3 signaling could be regulated in an autocrine or paracrine manner [
33,
34,
35]. However, it was unclear how the 24-hydroxylase, CYP24A1, the enzyme responsible for converting 25(OH)D
3 into 24,25(OH)
2D
3, was expressed or how the expression of both enzymes was related to ERα expression.
To elucidate the conflicting findings between serum vitamin D levels and laryngeal cancer progression, the present study aimed to evaluate the effect of the active vitamin D3 metabolite, 24R,25(OH)2D3, on markers of laryngeal tumorigenesis, determine if the expression of ERα66 is correlated with this effect in two different laryngeal cancer cell lines, and ascertain if these cells can regulate their tumorigenesis in an autocrine manner. We first examined the protein levels of ERα isoforms in both cell types to confirm a difference in ERα66 mRNA levels. We then treated the ERα66-positive and ERα66-negative cells with 24R,25(OH)2D3 to determine their response. We assessed the possibility of an autocrine effect by examining the presence of hydroxylase enzymes responsible for the hydroxylation of 25(OH)D3 and whether their levels are affected by estrogen. Finally, we examined the production of two vitamin D3 metabolites, 24,25(OH)2D3 and 1,25(OH)2D3.
2. Materials and Methods
2.1. Cell Culture
Human male laryngeal squamous-cell carcinoma cell lines, UM-SCC12 (RRID: CVCL7717) and UM-SCC-11A (RRID: CVCL_7715), were developed in the Carey Laboratory of the Department of Otolaryngology/Head and Neck Surgery at the University of Michigan and purchased from the university. Six weeks prior to receiving them, the cells were sent to the American Type Culture Collection (ATCC, Manassas, VA, USA) for authentication via short tandem repeat testing (STR) and mycoplasma detection. Once received, the cells were expanded and passaged at least 3 times before being used for experiments. Cells were cultured in high-glucose Dulbecco’s Modified Eagle Medium (DMEM; GIBCO™, Fisher Scientific, Hampton, NH, USA) lacking phenol red and supplemented with 10% heat-inactivated fetal bovine serum (FBS), 2mM L-glutamine, 1% non-essential amino acids, and 100 U/mL penicillin-streptomycin (Thermo Fisher Scientific, Waltham, MA, USA). Male rat kidney epithelial cells (NRK-52E; RRID: CVCL_0468) were purchased from ATCC (CRL-1571) and cultured in high-glucose DMEM supplemented with 10% heat-inactivated FBS and 1% penicillin/streptomycin. Once received, the cells were expanded and passaged 3 times prior to use for experimentation.
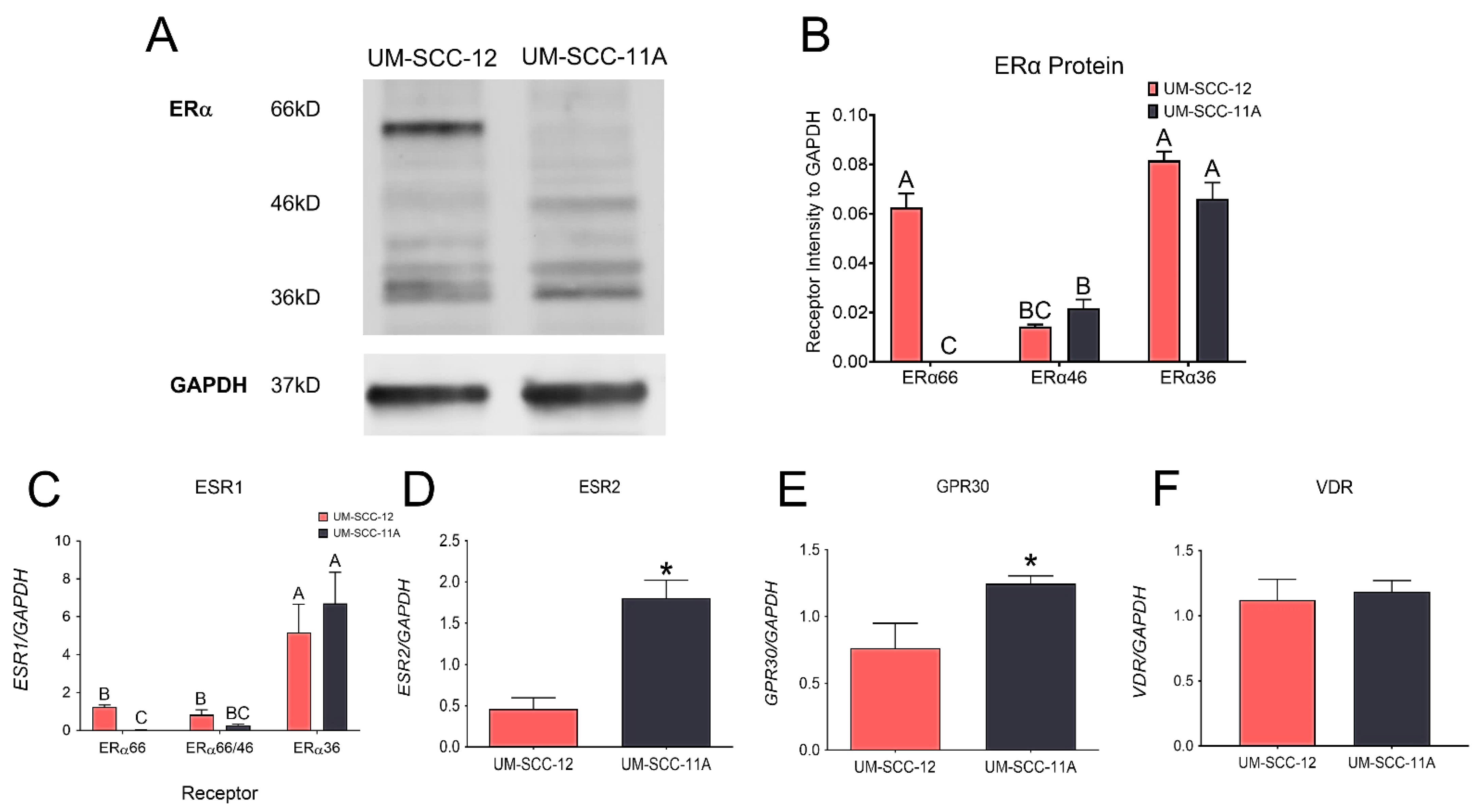
2.2. Characterization of Estrogen Receptors
Cells were seeded in a 6-well plate at 10,000 cells/cm2 (n = 3 wells/cell type) and grown until fully confluent. At confluence, media were replaced one last time. After 24 h, the cells were washed and lysed in a 200 μL/well of radioimmunoprecipitation assay (RIPA) buffer. The cell lysate proteins were resolved on 4–20% Tris-glycine extended gels by means of gel electrophoresis (Bio-Rad, Hercules, CA, USA) and transferred onto a PVDF membrane using the Bio-Rad Mini Trans-blot® Electrophoretic Transfer Cell. The membranes were then blocked and probed with a polyclonal ERα antibody (Thermo Fisher Scientific, RRID: AB_325814, CAT #PA1-309) along with a monoclonal GAPDH antibody (Millipore-Sigma, St. Louis, MO, USA, CAT #MAB374, Clone #6C5). Li-Cor IRDye® Goat anti-rabbit 800 CW (CAT# 926-32211, RRID: AB_621843) and Goat anti-mouse 680RD (CAT# 926-68070, RRID AB_10956588) secondary antibodies (Li-COR Biosciences, Lincoln, NE, USA) were used to visualize the proteins. The membranes were imaged using the LiCor-Biosciences Odyssey® CLx Infrared Imaging System.
The mRNA expression of estrogen-associated receptors and the vitamin D receptor were also measured as previously described [
31]. UM-SCC-12 and UM-SCC-11A cells were plated on 24-well plates (n = 6) and cultured until confluence. At confluence, media were changed one last time, and then the cells were harvested 12 h later. RNA was extracted using Trizol (Invitrogen, Waltham, MA, USA), and quantified using the Take3 Microvolume Plate (Biotek, Winooski, VT, USA). The RNA was then used to synthesize cDNA libraries (Life Technologies, Carlsbad, CA, USA, CAT#4368813). The levels of ERα66 were measured using a primer that amplifies the first 173 amino acids of exon 1 in ESR1 [
36]. Due to the structure of ERα46, it is not possible to distinguish ERα46 from ERα66; therefore, another primer was used, designed to amplify exons 7 and 8, found in both ERα46 and ERα66 but not in ERα36 [
36]. A primer that amplified the 9th exon of ESR1 measured the levels of ERα36. The expression levels of ESR2 (ERβ), GPR30, and the classical vitamin D receptor (VDR) were also measured. Levels were quantified by means of real-time quantitative PCR using Power SybrGreen
® Master Mix (Applied Biosystems, Walthalm, MA, USA) and its associated primers designed through Thermo Fisher Scientific (
Supplementary Table S1). The expression of all receptors was normalized to GAPDH expression.
2.3. Response to 24R,25(OH)2D3
24R,25(OH)2D3 was purchased from Enzo Life Sciences (Farmingdale, NY, USA, BML-DM300) to assess the effect of 24R,25(OH)2D3 on markers of tumorigenicity. UM-SCC-12 and UM-SCC-11A cell monolayers were first cultured on 96-well or 24-well plates (n = 6 wells/group). At the time of treatment, the cells were treated with 10−7 M, 10−8 M, 10−9 M 24R,25(OH)2D3, or vehicle for 15 min, at which point the media were replaced with fresh full media for 24 h. The cells were then assessed for proliferation via the EdU DNA synthesis assay (Thermo Fisher Scientific), and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, (MTT) (Sigma-Aldrich, St. Louis, MO, USA). Apoptosis was measured via the p53 assay (R&D Systems, Minneapolis, MN, USA), DNA fragmentation (TUNEL assay) (Trevigen, Gaithersburg, MD, USA), BAX/BCL2 mRNA, and Caspase 9 Activity (Promega, Madison, WI, USA). Metastasis was measured through the Western blot analysis of Nidogen-1 and the ratio of osteoprotegerin to RANKL, two lung and bone metastasis markers, respectively.
2.3.1. Cell Proliferation
DNA Synthesis
UM-SCC-12 and UM-SCC-11A cells were cultured in 96-well plates until 70% confluence and then serum-starved in full media lacking FBS for 48 h. After 48 h of serum starvation, the cells were treated as described above and incubated in fresh media for 20 h. At that point, the cells were pulsed with 10 μL/well of 1:100 diluted 5-ethynyl-2′-deoxyuridine (EdU) and then incubated for another 4 h. Finally, the cells were harvested and assessed for proliferation according to the manufacturer’s protocol (Thermo Fisher Scientific).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide-MTT Assay
In a separate experiment assessing the effects of 24R,25(OH)2D3 on cell viability, UM-SCC-12 and UM-SCC-11A cells were cultured in 96-well plates until confluence and treated as stated previously. After 24 h, the cells were harvested and examined for formazan crystal solubilization according to the MTT assay protocol (Sigma-Aldrich).
2.3.2. Apoptosis
TUNEL Assay
Cells were cultured in 96-well plates (n = 6) until confluence and treated as described above. Twenty-four hours after treatment, DNA fragmentation was measured via colorimetric terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling (Trevigen, TiterTAC™ in situ microplate TUNEL assay) assay according to the manufacturer’s protocol.
BAX/BCL2 Gene Expression
The ratio of apoptotic markers B cell lymphoma protein-2-associated X protein (BAX) and B cell lymphoma protein 2 (BCL2) demonstrates the life or death response of cells to an apoptotic stimulus [
31]. An increase in the BAX/BCL2 ratio indicates a decrease in resistance to apoptotic stimuli, leading to an increase in cell death [
37]. Cells were harvested 12 h after treatment and assessed for gene expression as described above. BAX and BCL2 mRNA expression were first normalized to GAPDH and then the ratio of BAX to BCL2 was calculated.
Total p53 Content
Total p53 content was measured to evaluate apoptosis. UM-SCC-12 and UM-SCC-11A cells were plated in 24-well plates and treated as described above. Twenty-four hours after treatment, the cells were lysed in 500 μL 1mM EDTA, 0.05% Triton™X-100. The total p53 content was measured in the cell lysate and normalized to total protein content (Pierce 660nM BCA Protein Assay, Thermo Fisher Scientific) according to the manufacturer’s protocol.
Caspase 9 Activity
The initiator caspase, caspase-9, plays a critical role in the intrinsic pathway, activating effector caspases, which initiate the degradation phase of apoptosis [
38]. Caspase 9 activity was measured 24 h after treatment using the Caspase-Glo
® 9 Assay (Promega, CAT #G8210) according to the manufacturer’s protocol.
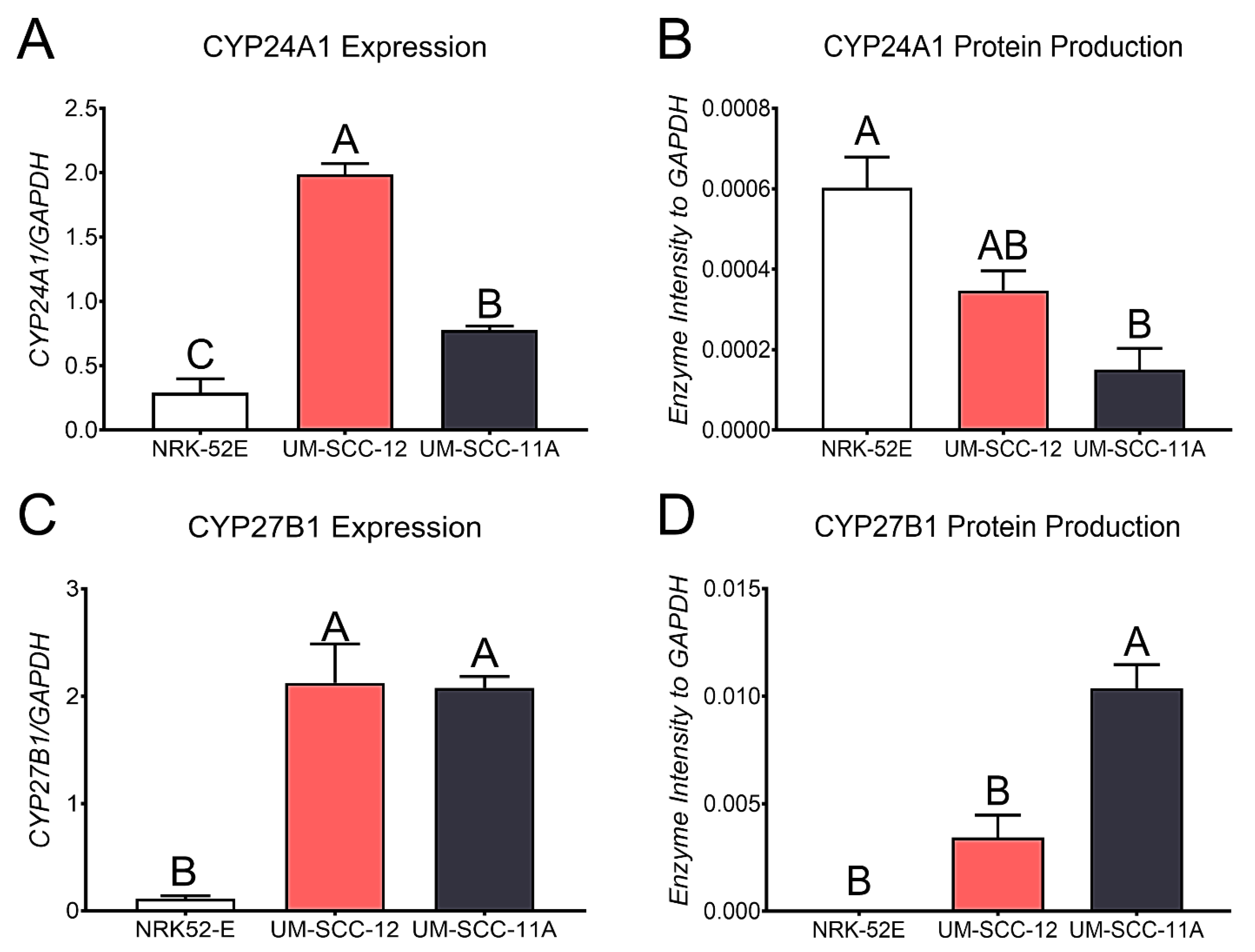
24-Hydroxylase and 1-Hydroxylase Measurements
Quantitative PCR was used to examine the levels of CYP24A1 and CYP27B1 in both ERα66-positive UM-SCC-12 and ERα66-negative UM-SCC-11A cells. 25(OH)D
3 is hydroxylated to 1,25(OH)
2D
3 or 24,25(OH)
2D
3 in the kidneys; therefore, NRK-52E rat kidney epithelial cells were used as a positive control for baseline levels of the 24-hydroxylase CYP24A1 and the 1-hydroxylase CYP27B1 enzymes as described above [
31]. CYP24A1 and CYP27B1 expression levels in confluent cultures were normalized to the expression levels of GAPDH.
To measure the amounts of each hydroxylase, UM-SCC-12, UM-SCC-11A, and NRK-52E cells were cultured to confluence. The cells were harvested after medium change. Transfer membranes of the cell lysates were probed with a CYP24A1 polyclonal antibody (Thermo Fisher Scientific, RRID: AB_11154312, CAT #PA521704), or a CYP27B1 polyclonal antibody (Abcam, Cambridge, UK, CAT #ab206655, Clone #EPR20271) and a monoclonal GAPDH antibody. Proteins were normalized to GAPDH.
2.4. Local Production of Vitamin D3 Metabolites
2.4.1. Vitamin D Metabolites
25-hydroxyvitamin D3-deuterated 3 used for internal standards was ordered from Cambridge Isotope Laboratories (Tewksbury, MA, USA). 1,25-dihydroxyvitamin D3-d3 was used as another internal standard (Sigma Aldrich). 24R,25-dihydroxyvitamin D3-d6, 25-hydroxyvitamin D3 monohydrate-d6, and 1,25-dihydroxyvitamin D3-d6 were used for the calibration standard curves (Sigma Aldrich). Liquid chromatography–mass spectroscopy (LCMS)-grade methanol and water were used to make mobile phases and to extract and purify samples (Thermo Fisher Scientific, Waltham, WA). Ammonium acetate, formic acid, and molecular grade ethanol were used to make mobile phases (Sigma Aldrich).
2.4.2. Cell Treatment
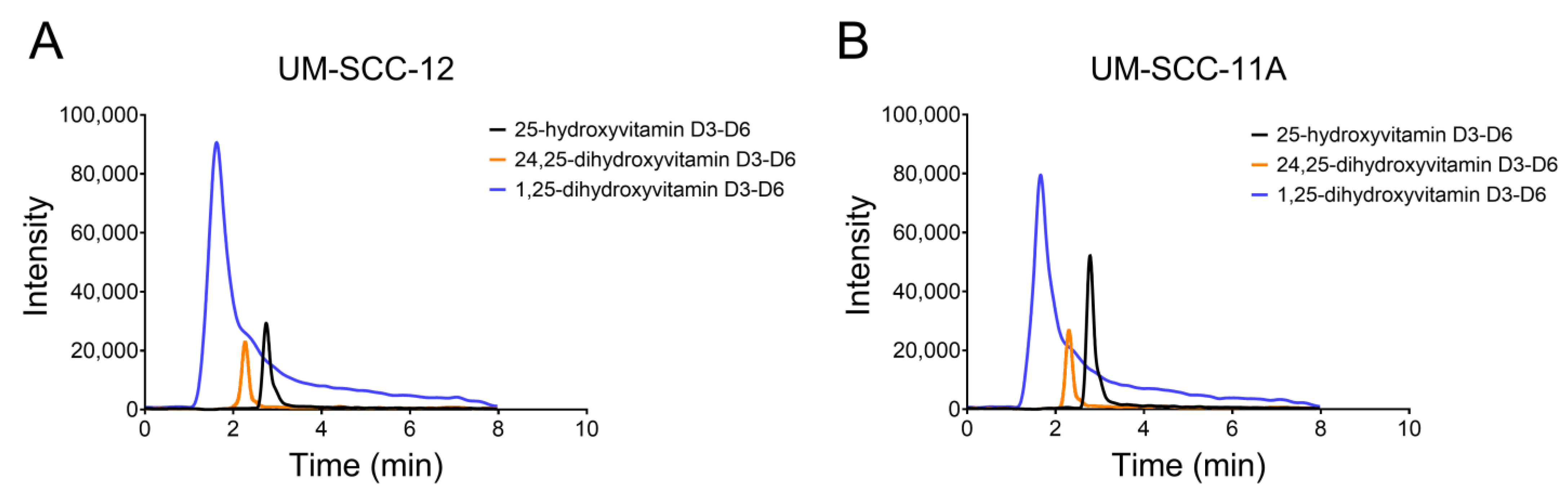
NRK-52E, UM-SCC-12, and UM-SCC-11A cells were cultured to confluence in 6 T75 flasks seeded at 10,000 cells/cm2. At confluence, the cells were treated with 5 × 10−7 M (0.203 μg/mL) 25(OH)D3-d6 for 24 h. The conditioned media were collected and extracted for HPLC to analyze the local secretion of vitamin D3 metabolites. Cell layers were lysed with Triton X-100 in 1x PBS to calculate DNA content.
2.4.3. Analytical Methods
The production of 1,25(OH)
2D
3 and 24,25(OH)
2D
3 was measured using high-performance liquid chromatography–mass spectrometry as previously described [
31]. Briefly, a 9-point standard curve was created from a range of 0.2-250 ng/mL from stock solutions of 24R,25-dihydroxyvitamin D3-d6, 25-hydroxyvitamin D3 monohydrate-d6, and 1,25-dihydroxyvitamin D3-d6. 1,25-dihydroxyvitamin D3-d3 and 25-hydroxyvitamin D
3-d3 were used as internal standards. Calibration curves are shown in
Supplemental Figure S1.
The equations used to calculate vitamin D3 metabolites are in the supplementary methods. Measurements of 25(OH)D3, 1,25(OH)2D3, and 24,25(OH)2D3 in the conditioned media were compared to the 9-point standard curve equation, described below. The percent conversion of 25(OH)D3 to 1,25(OH)2D3 or 24,25(OH)2D3 was calculated as follows: % conversion = [(ng/mL metabolite)/(total 25(OH)D3 + 1,25(OH)2D3 + 24,25(OH)2D3)] × 100. The molarity of 1,25(OH)2D3 or 24,25(OH)2D3 in the conditioned media was calculated as follows: Molarity = grams/(volume × molecular weight).
Standard curves were generated as described below. Mass-to-charge ratios (423.55 > 121.18, 437.76 > 138.01, 407.47 > 107.21, 440.75 > 135.01) were used for quantification with cone and collision voltages between 13 and 17V and 14–18 V, respectively [
26]. The calibration curve was calculated through weighted linear regression. The normalized peak area ratio of analyte to internal standard was plotted against the concentration of the 3 metabolites, 25(OH)D
3, 1,25(OH)
2D
3, and 24,25(OH)
2D
3 [
31]. The transitions were monitored for 62 ms.
2.5. Statistical Analysis
All data graphed are shown as means ± SEM. Treatment vs. control was assessed by means of the Wilcoxon matched-pairs signed rank test, with p-values ≤ 0.05 determined as significant. Baseline enzyme levels and vitamin D3 metabolite levels were determined significantly different between groups using one-way ANOVA with a Tukey post-test correction, with p-values ≤ 0.05 considered significant. Timepoint analyses were evaluated using a two-way ANOVA and a Tukey post-test, and p-values ≤ 0.05 were considered significant.
4. Discussion
Vitamin D
3 supplementation has attracted much attention in cancer therapeutics, although its potential in laryngeal cancer has not been investigated. The contradictory reports of laryngeal cancer as a hormone-responsive cancer and vitamin D
3 deficiency as a risk factor make distinguishing the potential for vitamin D
3 treatment challenging. This study and previous studies show that many factors are at play to account for these contradictory findings. In female breast cancer, the presence or absence of ERα66 changes the cell’s response to treatment with vitamin D
3 metabolites [
33]. Our lab showed that some male laryngeal cancers are responsive to estrogen, opening the possibility of this factor playing a significant role in the response of male cells to metabolites of vitamin D
3 [
16]. Classifying laryngeal cancer as estrogen-responsive makes using ERα66 as an early diagnosis marker and a prognostic marker plausible. The high reactivity of estrogen receptor alpha was found only in malignant laryngeal tissue and not the surrounding tissue [
6,
13]. These findings make ERα66 a promising candidate as a marker for malignancy in the future.
ERα66 has long been used as a critical biomarker for determining aggression and the course of treatment in breast cancer. The prognostic value of ERα in male laryngeal cancer is becoming more apparent. More recent studies demonstrate the role of ERα36 in tumorigenesis by regulating metastasis and invasion, as well as resistance to hormone therapies like tamoxifen. Finally, the role of ERα46 in tumorigenesis needs further investigation.
We hypothesize that estrogen-responsive cancers will behave similarly and that the expression of ERα66 will change the response to vitamin D
3 treatment. Previous studies in our lab have already demonstrated the potential of ERα66 as an indicator of aggression in male laryngeal cancer, similar to female breast cancer [
16,
25]. Our Western blot analysis showed that different laryngeal squamous-cell carcinoma cells produce different levels of ERα66. To evaluate the role of ERα66 in mediating the effects of 24R,25(OH)
2D
3, we used two different male laryngeal cancer cell lines that produce similar levels of ERα36 and ERα46, but one cell type, UM-SCC-12 cells, also has ERα66. The differential expression of ERα66 emphasizes its role in mediating the effects observed because there is no change in the levels of ERα36 or ERα46. We observed a difference in expression of ERβ and GPR30, so the role of these receptors in mediating the effects cannot be ruled out. The similar expression of VDR observed in both cell types makes it unlikely that VDR plays a role in mediating the effect of 24R,25(OH)
2D
3. However, as we have observed in breast cancer, cells that differentially express ERα66 respond differently to the vitamin D
3 metabolite, 24R,25(OH)
2D
3 [
31], making its role in mediating laryngeal cancer progression more plausible. Therefore, we first wanted to evaluate the cells’ response to 24R,25(OH)
2D
3.
We examined the therapeutic potential of 24R,25(OH)
2D
3 in laryngeal cancer cells by treating UM-SCC-12 and UM-SCC-11A cells with vehicle or 24R,25(OH)
2D
3. 24R,25(OH)
2D
3 increased markers of tumorigenesis in the ERα66-positive UM-SCC-12 cells by increasing indicators of proliferation and metastasis and reducing apoptosis. In contrast, 24R,25(OH)
2D
3 reduced markers of tumorigenesis in the ERα66-negative UM-SCC-11A through a reduction in markers of proliferation and metastasis and an increase in apoptosis. Overall, because these cells differentially express ERα66, these data demonstrate that the effect of 24R,25(OH)
2D
3 on laryngeal cancer is related to the expression of ERα66. However, we also observed a difference in the expression of ERβ and GPR30, which may also play a role. Interestingly, in ERα66-positive MCF7 breast cancer cells treated with 24R,25(OH)
2D
3, although proliferation was stimulated, the increase in apoptosis and the reduction in various metastatic markers demonstrated an overall antitumorigenic effect, while in ERα66-negative HCC38 breast cancer, 24R,25(OH)
2D
3 increased tumorigenesis [
33]. The similar expression levels of ERβ and GPR30 in both breast cancer cells further suggest that ERa66 is the major player regulating the response to 24R,25(OH)
2D
3, which could be responsible for the difference in both cancers. The opposite effects observed in laryngeal cancer versus breast cancer may be due to the difference in male cells compared to female cells.
These data suggest that the expression of ERα66 plays a critical role in the cellular response to the vitamin D3 metabolite. However, while there was a difference in cellular response to 24R,25(OH)2D3 based on ERα66 expression, it was unclear as to why these responses were different in these two different cancer types even when expressing similar levels of ERα66. This could reflect a fundamental difference between breast cancer and laryngeal cancer. Another possible reason for the difference could be sex-dependent; the breast cancer cells originated from female donors and the laryngeal cancer cells were taken from male donors. Alternatively, factors other than ERα66 may contribute to the local regulation of 24R,25(OH)2D3 through autocrine signaling to mediate its effect on tumorigenesis. For example, ERβ and GPR30 were present in the cells and may interact with ERα isoforms; similarly, the androgen receptor may play a role.
Based on previous data, demonstrating the regulation of vitamin D3 hydroxylases by estrogen [
42], as well as the ability of laryngeal cancer cells to produce their own estrogen [
25], suggested the potential for an autocrine-mediated response to 24R,25(OH)
2D
3. Therefore, we aimed to determine the if the laryngeal cancer cells can produce vitamin D
3 metabolites locally, which could induce an autocrine effect on the cells. Our results confirm that laryngeal cancer cells can convert 25(OH)D
3 into active metabolites. Both cell types expressed the 24-hydroxylase, CYP24A1, and the 1−hydroxylase, CYP27B1, demonstrating the potential for producing active vitamin D
3 metabolites. CYP24A1 is not only responsible for the hydroxylation of 25(OH)D
3 to 24,25(OH)
2D
3 but also the hydroxylation of 1,25(OH)
2D
3 into 1,24,25(OH)
3D
3. A high expression of CYP24A1 is known as the initial step in the degradation of the 1,25(OH)
2D
3 pathway [
31,
35,
43]. The ERα66-positive UM-SCC-12 cells expressed significantly higher levels than the ERα66-negative UM-SCC-11A cells, and both cell types expressed more elevated levels than the positive control NRK-52E cells. However, when evaluating the cells for protein production, NRK-52E cells produced the most CYP24A1, significantly more than UM-SCC-11A cells. CYP27B1 expression showed higher levels in both laryngeal cancer cells than NRK-52E cells. However, the expression did not change with ERα66 expression. In contrast, the protein levels of CYP27B1 increased in ERα66-negative UM-SCC-11A cells compared to UM-SCC-12 cells. Together, these data suggest that laryngeal cancer’s ability to produce vitamin D
3 metabolites in response to 25(OH)D
3 may depend on the expression of ERα66. The local production of active vitamin D
3 metabolites can affect the cells through autocrine/paracrine signaling [
34,
35]. It is also interesting to note that the cells examined show similar levels of 1,25(OH)
2D
3 production, while the production of 24R,25(OH)
2D
3 was cell type-dependent, which may indicate the vital role of 24R,25(OH)
2D
3 on cell tumorigenesis and as an endocrine factor.
Laryngeal cancer cells and breast cancer cells produced 24,25(OH)
2D
3 and 1,25(OH)
2D
3 at levels that were greater than are present in serum [
31,
44,
45]. Moreover, the locally produced metabolites were at levels corresponding to the concentrations at which we noted cellular responses [
45]. Both laryngeal cancer cell lines produced significantly more 24,25(OH)
2D
3 than NRK-52E cells in response to 25(OH)D
3, and UM-SCC-11A cells produced substantially more 24,25(OH)
2D
3 compared to UM-SCC-12 cells. All three cell lines produced similar levels of 1,25(OH)
2D
3, significantly higher than the 30 pmol/L serum levels measured in patients. UM-SCC-11A cells produced the highest amount of 24,25(OH)
2D
3, similar to MCF7 cells [
31]. Interestingly, while MCF7 cells are ERα66-positive, they responded similarly to 24R,25(OH)
2D
3 as UM-SCC-11A cells in that tumorigenesis was reduced [
33]. UM-SCC-12 cells, while ERα66-positive, responded similarly to the ERα66-negative breast cancer cell line HCC38.
It is important to note that the expression and protein levels of the CYP24A1 and CYP27B1 enzymes are not directly correlated with the production of the local metabolites. The expression and protein levels do not measure enzyme activity and, therefore, cannot be compared to the production of the vitamin D3 metabolites. The presence of the enzymes is used to indicate the cells’ ability to produce metabolites. These findings suggest that the response to 24R,25(OH)2D3 may be dependent on the local production of the metabolite which is related to the presence of ERα66 and the sex of the cells.
In summary, 24R,25(OH)2D3 can regulate markers of laryngeal cancer tumorigenesis and its effect is dependent on the expression of ERα66, similar to that observed in breast cancer. These findings demonstrate a potential therapeutic agent for aggressive laryngeal cancer cells that do pose a risk to calcium levels, making it more plausible as an anti-tumor agent. Through the expression of CYP24A1 and CYP27B1, laryngeal cancer cells can produce vitamin D3 metabolites 1,25(OH)2D3 and 24,25(OH)2D3 at biologically relevant levels. The local production of these metabolites suggests the potential autocrine or paracrine effects of these metabolites on tumorigenesis. 24R,25(OH)2D3 may be used as a potential therapeutic agent in some laryngeal cancers; however, the local production and the level of ERα66 must be considered. This study did not measure the production of other vitamin D3 metabolites, so their role in laryngeal cancer tumorigenesis cannot be ruled out. The similar expression of ERα36 and ERα46 in the two cell lines eliminates the possibility of these receptors playing a role in this effect, but the role of the other active receptor, ERβ, cannot be dismissed. Further investigation is needed to address these possibilities. To date, the use of vitamin D3 supplementation in laryngeal cancer has not yet been examined.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





