
Assessing the Effects of Curcumin and 450 nm Photodynamic Therapy on Oxidative Metabolism and Cell Cycle in Head and Neck Squamous Cell Carcinoma: An In Vitro Study

 ,
,  ,
,  ,
,  ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Experimental Setup, Curcumin Treatments, and Irradiated Parameters
2.3. ATP Synthase Activity Assay
2.4. Oxygen Consumption Assay
2.5. Cell Homogenate Preparation
2.6. Assay of Antioxidant Enzyme Activities
2.7. Malondialdehyde Assay
2.8. Evaluation of Cellular Growth and Cell Cycle
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Result
3.1. Curcumin in Higher Doses Alone or with Laser Light Irradiation Produces a Decrease in Aerobic ATP Synthesis and an Increase in Oxygen Consumption by Inducing the Uncoupling of Oxidative Phosphorylation
3.2. Curcumin in Higher Doses Alone or with Laser Light Irradiation Causes a Further Decrease in Cellular Energetics Compared to the Native Molecule
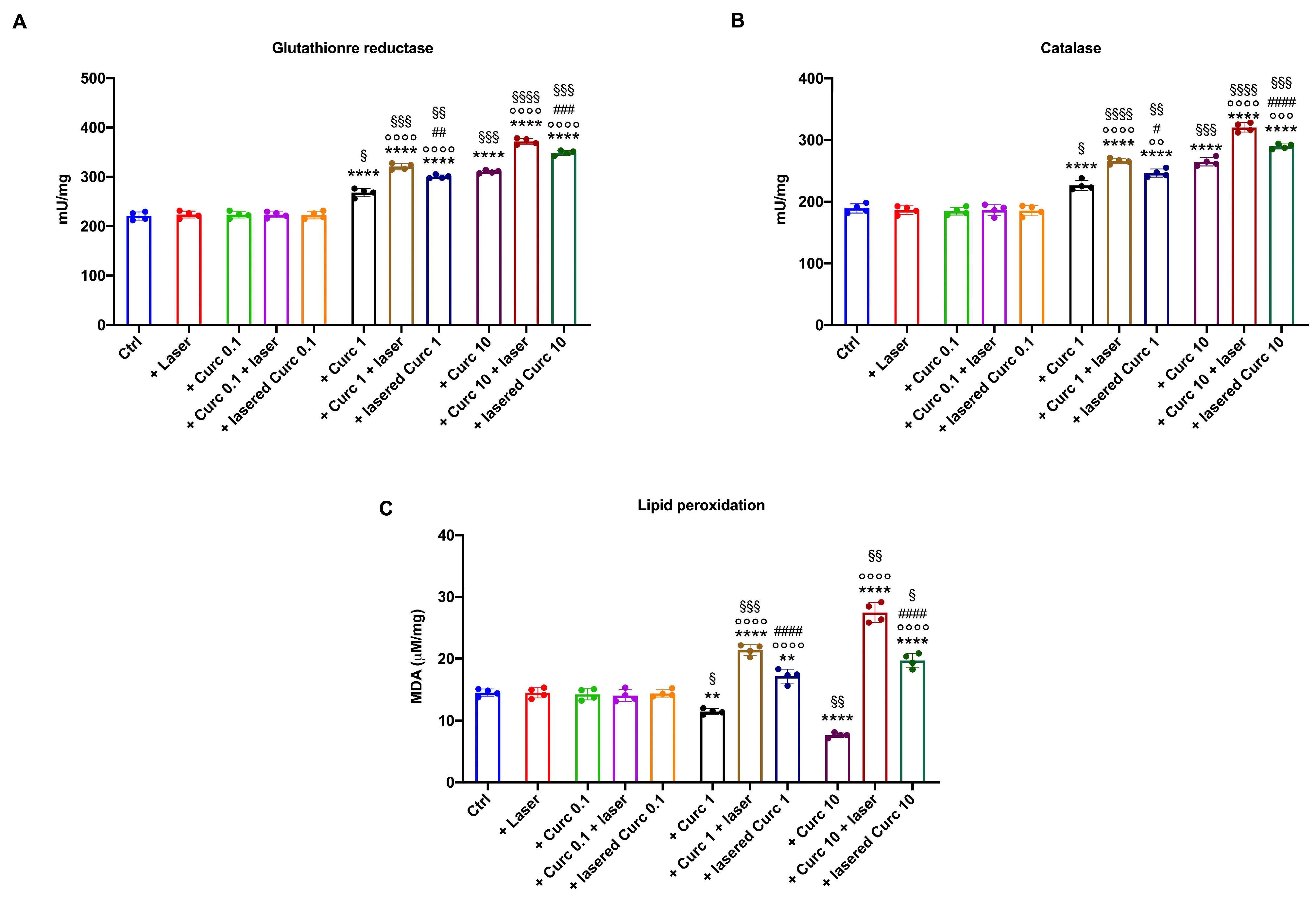
3.3. Curcumin in Higher Doses Alone or with Laser Light Irradiation Induces an Antioxidant Response That Is Insufficient to Limit Oxidative Damage
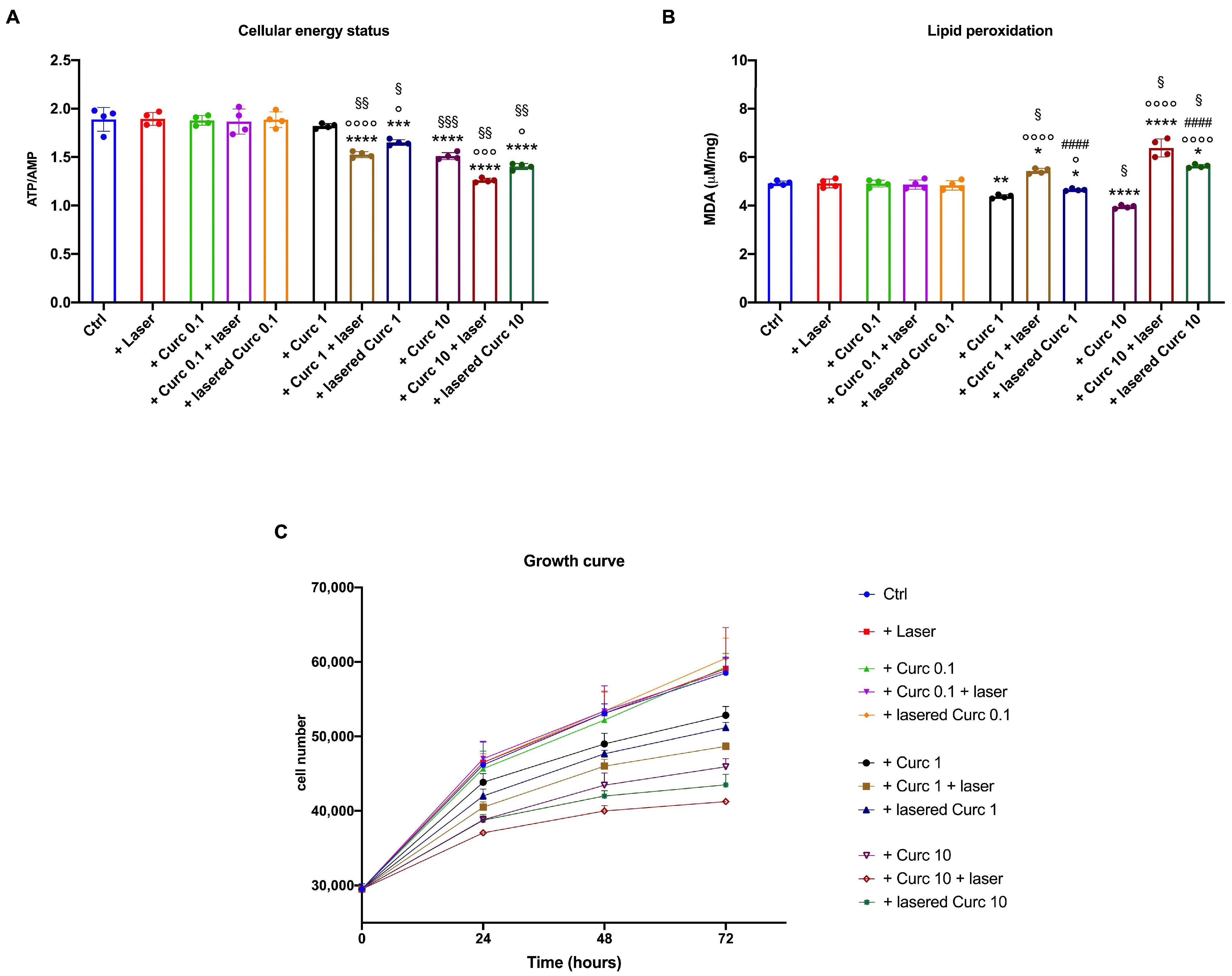
3.4. Curcumin in Higher Doses Alone or with Laser Light Irradiation Slows Cell Growth, Decreases Cell Viability, and Blocks the Cell Cycle at the S-G2M Phase Compared to Curcumin in the Dark
3.5. Curcumin in Higher Doses Alone or with Laser Light Irradiation Causes a Decrease in Energy Status and Growth and an Increase in Lipid Peroxidation Accumulation in Primary Human Fibroblasts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Inchingolo, F.; Santacroce, L.; Ballini, A.; Topi, S.; Dipalma, G.; Haxhirexha, K.; Bottalico, L.; Charitos, I.A. Oral Cancer: A Historical Review. Int. J. Environ. Res. Public Health 2020, 17, 3168. [Google Scholar] [CrossRef] [PubMed]
- van Straten, D.; Mashayekhi, V.; de Bruijn, H.S.; Oliveira, S.; Robinson, D.J. Oncologic Photodynamic Therapy: Basic Principles, Current Clinical Status and Future Directions. Cancers 2017, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Beyer, K.; Nikfarjam, F.; Butting, M.; Meissner, M.; König, A.; Bosca, A.R.; Kaufmann, R.; Heidemann, D.; Bernd, A.; Kippenberger, S.; et al. Photodynamic Treatment of Oral Squamous Cell Carcinoma Cells with Low Curcumin Concentrations. J. Cancer 2017, 8, 1271–1283. [Google Scholar] [CrossRef]
- Ibarra, A.M.C.; Cecatto, R.B.; Motta, L.J.; dos Santos Franco, A.L.; de Fátima Teixeira da Silva, D.; Nunes, F.D.; Hamblin, M.R.; Rodrigues, M.F.S.D. Photodynamic Therapy for Squamous Cell Carcinoma of the Head and Neck: Narrative Review Focusing on Photosensitizers. Lasers Med. Sci. 2022, 37, 1441–1470. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Liu, J.; He, L.; Liu, L.; Cheng, B.; Zhou, F.L.; Cao, D.L.; He, Y.C. A Comprehensive Review on the Benefits and Problems of Curcumin with Respect to Human Health. Molecules 2022, 27, 4400. [Google Scholar] [CrossRef] [PubMed]
- Bernd, A. Visible Light and/or UVA Offer a Strong Amplification of the Anti-Tumor Effect of Curcumin. Phytochem. Rev. 2014, 13, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Ji, X.; Zhang, Q.; Wei, Y. Curcumin Combined with Photodynamic Therapy, Promising Therapies for the Treatment of Cancer. Biomed. Pharmacother. 2022, 146, 112567. [Google Scholar] [CrossRef] [PubMed]
- Kah, G.; Chandran, R.; Abrahamse, H. Curcumin a Natural Phenol and Its Therapeutic Role in Cancer and Photodynamic Therapy: A Review. Pharmaceutics 2023, 15, 639. [Google Scholar] [CrossRef]
- Shehzad, A.; Shahzad, R.; Lee, Y.S. Curcumin: A Potent Modulator of Multiple Enzymes in Multiple Cancers. Enzymes 2014, 36, 149–174. [Google Scholar] [CrossRef]
- Shen, L.; Ji, H.F. Theoretical Study on Physicochemical Properties of Curcumin. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2007, 67, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Ravera, S.; Traverso, C.; Amaro, A.; Piaggio, F.; Emionite, L.; Bachetti, T.; Pfeffer, U.; Raffaghello, L. Curcumin Induces a Fatal Energetic Impairment in Tumor Cells in Vitro and in Vivo by Inhibiting ATP-Synthase Activity. Carcinogenesis 2018, 39, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, U.; Bianchi, G.; Ravera, S.; Traverso, C.; Amaro, A.; Piaggio, F.; Emionite, L.; Bacchetti, T.; Raffaghello, L. Abstract 3513: Curcumin Induces a Fatal Energetic Impairment by Inhibiting ATP-Synthase Activity and Decreasing ATP Generation and Oxygen Consumption in in Vitro and in Vivo Tumor Models. Cancer Res. 2018, 78, 3513. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, S.; Kumar, P.; Jain, G.K.; Aggarwal, G.; Almalki, W.H.; Kesharwani, P. Nanomaterials for Photodynamic Therapy, 1st ed.; Woodhead Publishing (Elsevier): Cambridge, UK, 2023; pp. 41–54. [Google Scholar] [CrossRef]
- Juarranz, Á.; Jaén, P.; Sanz-Rodríguez, F.; Cuevas, J.; González, S. Photodynamic Therapy of Cancer. Basic Principles and Applications. Clin. Transl. Oncol. 2008, 10, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, M. Photophysical and Photobiological Processes in the Photodynamic Therapy of Tumours. J. Photochem. Photobiol. B Biol. 1997, 39, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bienkowska, K.J.; Hanley, C.J.; Thomas, G.J. Cancer-Associated Fibroblasts in Oral Cancer: A Current Perspective on Function and Potential for Therapeutic Targeting. Front. Oral. Health 2021, 2, 686337. [Google Scholar] [CrossRef] [PubMed]
- Datar, U.V.; Kale, A.D.; Angadi, P.V.; Hallikerimath, S.; Deepa, M.; Desai, K.M. Role of Cancer-Associated Fibroblasts in Oral Squamous Cell Carcinomas, Surgical Margins, and Verrucous Carcinomas: An Immunohistochemical Study. J. Clin. Transl. Res. 2022, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Ravera, S.; Bertola, N.; Pasquale, C.; Bruno, S.; Benedicenti, S.; Ferrando, S.; Zekiy, A.; Arany, P.; Amaroli, A. 808-Nm Photobiomodulation Affects the Viability of a Head and Neck Squamous Carcinoma Cellular Model, Acting on Energy Metabolism and Oxidative Stress Production. Biomedicines 2021, 9, 1717. [Google Scholar] [CrossRef] [PubMed]
- Bertola, N.; Degan, P.; Cappelli, E.; Ravera, S. Mutated FANCA Gene Role in the Modulation of Energy Metabolism and Mitochondrial Dynamics in Head and Neck Squamous Cell Carcinoma. Cells 2022, 11, 2353. [Google Scholar] [CrossRef]
- Amaroli, A.; Arany, P.; Pasquale, C.; Benedicenti, S.; Bosco, A.; Ravera, S. Improving Consistency of Photobiomodulation Therapy: A Novel Flat-Top Beam Hand-Piece versus Standard Gaussian Probes on Mitochondrial Activity. Int. J. Mol. Sci. 2021, 22, 7788. [Google Scholar] [CrossRef]
- Hanna, R.; Agas, D.; Benedicenti, S.; Ferrando, S.; Laus, F.; Cuteri, V.; Lacava, G.; Sabbieti, M.G.; Amaroli, A. A Comparative Study between the Effectiveness of 980 Nm Photobiomodulation Delivered by Hand-Piece with Gaussian vs. Flat-Top Profiles on Osteoblasts Maturation. Front. Endocrinol. 2019, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Rocca, J.P.; Zhao, M.; Fornaini, C.; Tan, L.; Zhao, Z.; Merigo, E. Effect of Laser Irradiation on Aphthae Pain Management: A Four Different Wavelengths Comparison. J. Photochem. Photobiol. B Biol. 2018, 189, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xu, Y.; Meng, L.; Huang, L.; Sun, H. Curcumin Inhibits Proliferation and Promotes Apoptosis of Breast Cancer Cells. Exp. Ther. Med. 2018, 16, 1266–1272. [Google Scholar] [CrossRef]
- Senft, C.; Polacin, M.; Priester, M.; Seifert, V.; Kögel, D.; Weissenberger, J. The Nontoxic Natural Compound Curcumin Exerts Anti-Proliferative, Anti-Migratory, and Anti-Invasive Properties against Malignant Gliomas. BMC Cancer 2010, 10, 491. [Google Scholar] [CrossRef]
- Dujic, J.; Kippenberger, S.; Hoffmann, S.; Ramirez-Bosca, A.; Miquel, J.; Diaz-Alperi, J.; Bereiter-Hahn, J.; Kaufmann, R.; Bernd, A. Low Concentrations of Curcumin Induce Growth Arrest and Apoptosis in Skin Keratinocytes Only in Combination with UVA or Visible Light. J. Investig. Dermatol. 2007, 127, 1992–2000. [Google Scholar] [CrossRef]
- Hinkle, P.C. P/O Ratios of Mitochondrial Oxidative Phosphorylation. Biochim. Biophys. Acta 2005, 1706, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative Stress and Cancer: An Overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Sathyabhama, M.; Priya Dharshini, L.C.; Karthikeyan, A.; Kalaiselvi, S.; Min, T. The Credible Role of Curcumin in Oxidative Stress-Mediated Mitochondrial Dysfunction in Mammals. Biomolecules 2022, 12, 1405. [Google Scholar] [CrossRef] [PubMed]
- Piper, J.T.; Singhal, S.S.; Salameh, M.S.; Torman, R.T.; Awasthi, Y.C.; Awasthi, S. Mechanisms of Anticarcinogenic Properties of Curcumin: The Effect of Curcumin on Glutathione Linked Detoxification Enzymes in Rat Liver. Int. J. Biochem. Cell Biol. 1998, 30, 445–456. [Google Scholar] [CrossRef]
- Wu, L.; Guo, L.; Liang, Y.; Liu, X.; Jiang, L.; Wang, L. Curcumin Suppresses Stem-like Traits of Lung Cancer Cells via Inhibiting the JAK2/STAT3 Signaling Pathway. Oncol. Rep. 2015, 34, 3311–3317. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.C.; Kang, J.W.; Shin, J.I.; Chung, P.S. Combination Treatment with Photodynamic Therapy and Curcumin Induces Mitochondria-Dependent Apoptosis in AMC-HN3 Cells. Int. J. Oncol. 2012, 41, 2184–2190. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Fang, C.; Zhang, J.; Liu, B.; Wei, Z.; Fan, X.; Sui, Z.; Tan, Q. Catanionic Lipid Nanosystems Improve Pharmacokinetics and Anti-Lung Cancer Activity of Curcumin. Nanomedicine 2016, 12, 1567–1579. [Google Scholar] [CrossRef] [PubMed]
- Borges, G.Á.; Rêgo, D.F.; Assad, D.X.; Coletta, R.D.; De Luca Canto, G.; Guerra, E.N.S. In Vivo and in Vitro Effects of Curcumin on Head and Neck Carcinoma: A Systematic Review. J. Oral. Pathol. Med. 2017, 46, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Van Der Merwe, S.M.; Verhoef, J.C.; Verheijden, J.H.M.; Kotzé, A.F.; Junginger, H.E. Trimethylated Chitosan as Polymeric Absorption Enhancer for Improved Peroral Delivery of Peptide Drugs. Eur. J. Pharm. Biopharm. 2004, 58, 225–235. [Google Scholar] [CrossRef]
- Garcea, G.; Jones, D.J.L.; Singh, R.; Dennison, A.R.; Farmer, P.B.; Sharma, R.A.; Steward, W.P.; Gescher, A.J.; Berry, D.P. Detection of Curcumin and Its Metabolites in Hepatic Tissue and Portal Blood of Patients Following Oral Administration. Br. J. Cancer 2004, 90, 1011–1015. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Morón, E.; Calderón-Montaño, J.M.; Salvador, J.; Robles, A.; López-Lázaro, M. The Dark Side of Curcumin. Int. J. Cancer 2010, 126, 1771–1775. [Google Scholar] [CrossRef] [PubMed]
- Zhongfa, L.; Chiu, M.; Wang, J.; Chen, W.; Yen, W.; Fan-Havard, P.; Yee, L.D.; Chan, K.K. Enhancement of Curcumin Oral Absorption and Pharmacokinetics of Curcuminoids and Curcumin Metabolites in Mice. Cancer Chemother. Pharmacol. 2012, 69, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Chaturvedi, M.; Mishra, S.; Kumar, P.; Somvanshi, P.; Chaturvedi, R. Reductive Metabolites of Curcumin and Their Therapeutic Effects. Heliyon 2020, 6, e05469. [Google Scholar] [CrossRef]
- Appendino, G.; Allegrini, P.; de Combarieu, E.; Novicelli, F.; Ramaschi, G.; Sardone, N. Shedding Light on Curcumin Stability. Fitoterapia 2022, 156, 105084. [Google Scholar] [CrossRef]
- Dujic, J.; Kippenberger, S.; Ramirez-Bosca, A.; Diaz-Alperi, J.; Bereiter-Hahn, J.; Kaufmann, R.; Bernd, A.; Hofmann, M. Curcumin in Combination with Visible Light Inhibits Tumor Growth in a Xenograft Tumor Model. Int. J. Cancer 2009, 124, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Yoshino, F.; Makita, T.; Maehata, Y.; Higashi, K.; Miyamoto, C.; Wada-Takahashi, S.; Takahashi, S.S.; Takahashi, O.; Lee, M.C.-I. Reactive Oxygen Species Production in Mitochondria of Human Gingival Fibroblast Induced by Blue Light Irradiation. J. Photochem. Photobiol. B 2013, 129, 1–5. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravera, S.; Pasquale, C.; Panfoli, I.; Bozzo, M.; Agas, D.; Bruno, S.; Hamblin, M.R.; Amaroli, A. Assessing the Effects of Curcumin and 450 nm Photodynamic Therapy on Oxidative Metabolism and Cell Cycle in Head and Neck Squamous Cell Carcinoma: An In Vitro Study. Cancers 2024, 16, 1642. https://doi.org/10.3390/cancers16091642
Ravera S, Pasquale C, Panfoli I, Bozzo M, Agas D, Bruno S, Hamblin MR, Amaroli A. Assessing the Effects of Curcumin and 450 nm Photodynamic Therapy on Oxidative Metabolism and Cell Cycle in Head and Neck Squamous Cell Carcinoma: An In Vitro Study. Cancers. 2024; 16(9):1642. https://doi.org/10.3390/cancers16091642
Chicago/Turabian StyleRavera, Silvia, Claudio Pasquale, Isabella Panfoli, Matteo Bozzo, Dimitrios Agas, Silvia Bruno, Michael R. Hamblin, and Andrea Amaroli. 2024. "Assessing the Effects of Curcumin and 450 nm Photodynamic Therapy on Oxidative Metabolism and Cell Cycle in Head and Neck Squamous Cell Carcinoma: An In Vitro Study" Cancers 16, no. 9: 1642. https://doi.org/10.3390/cancers16091642
APA StyleRavera, S., Pasquale, C., Panfoli, I., Bozzo, M., Agas, D., Bruno, S., Hamblin, M. R., & Amaroli, A. (2024). Assessing the Effects of Curcumin and 450 nm Photodynamic Therapy on Oxidative Metabolism and Cell Cycle in Head and Neck Squamous Cell Carcinoma: An In Vitro Study. Cancers, 16(9), 1642. https://doi.org/10.3390/cancers16091642