Mucins and Pancreatic Cancer




{kind=link}
{kind=link}
Abstract
:1. Pancreatic Cancer

2. Mucins
2.1. MUC1
2.2. MUC4
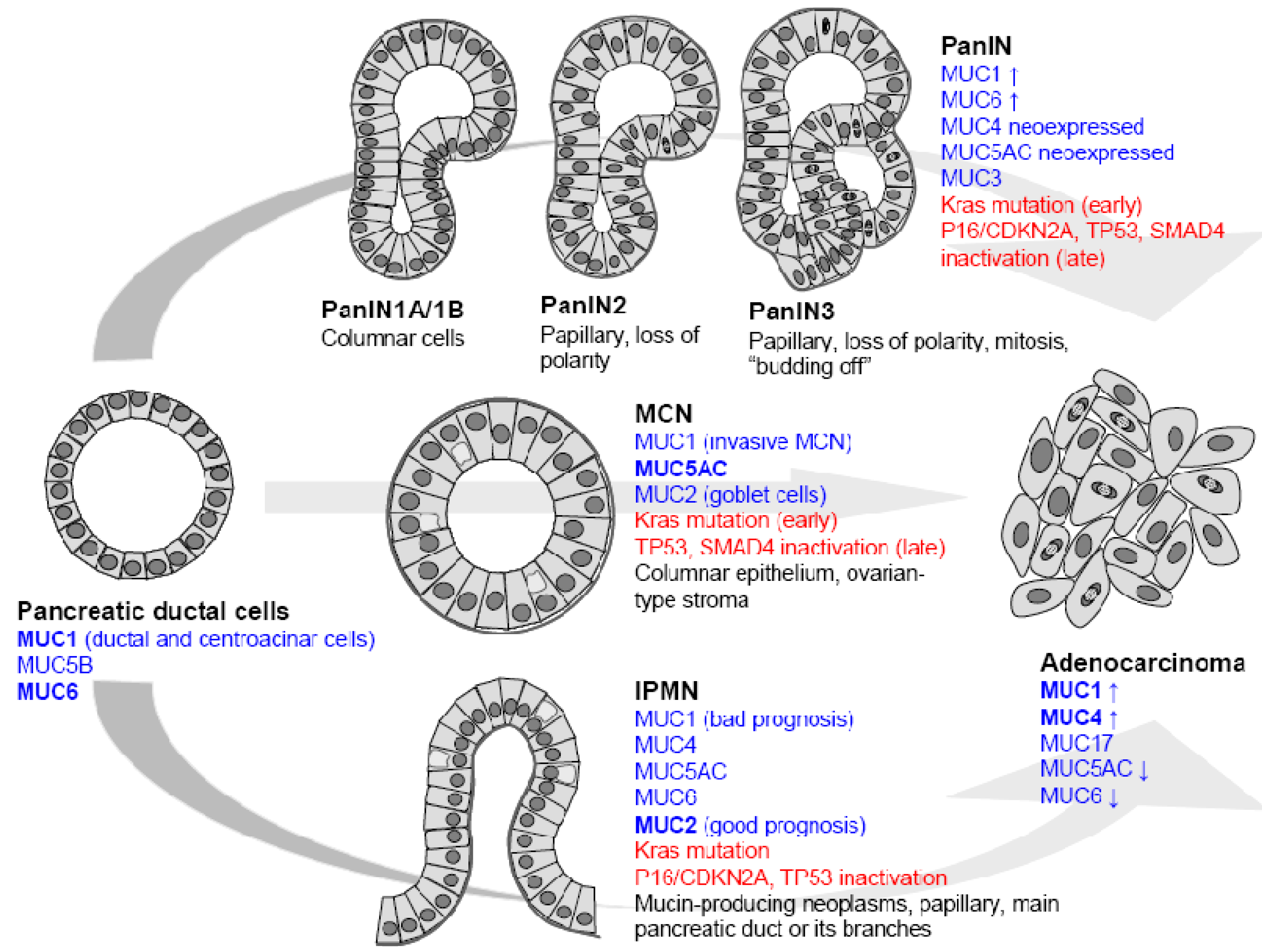
3. Mucins and the Normal Pancreas
4. Mucins and Pancreatic Cancer
5. Mucin Gene Regulation in Pancreatic Cancer
6. Epigenetic Regulation of Mucin Genes in Pancreatic Cancer
7. Mucins as Biomarkers of Pancreatic Cancer
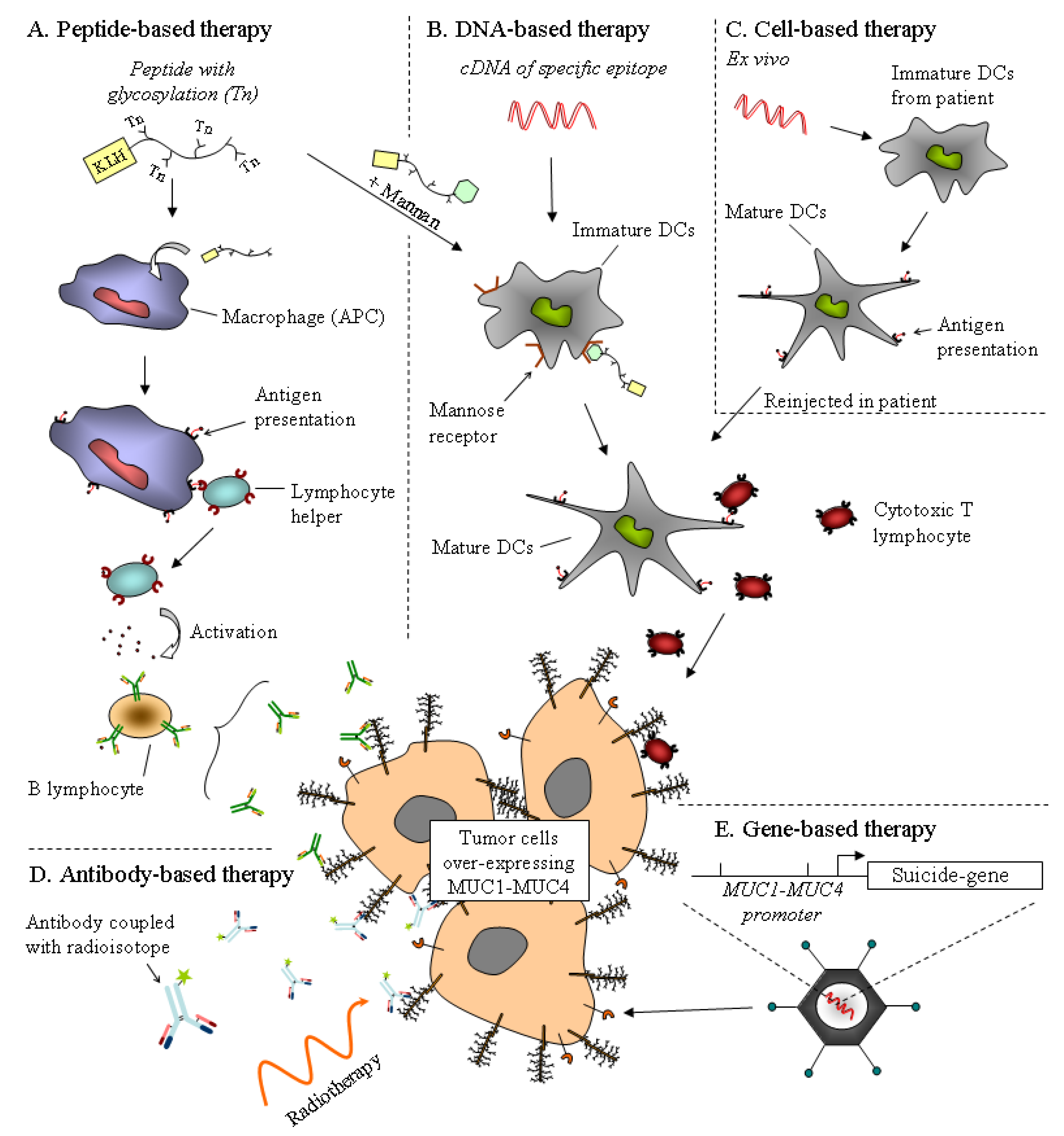
8. Mucins as Therapeutic Tools in Pancreatic Carcinogenesis?
8.1. Immunotherapy
8.2. Gene-Based Therapy

8.3. Chemotherapy
9. Perspectives
Acknowledgments
References
- Bardeesy, N.; DePinho, R.A. Pancreatic cancer biology and genetics. Nat. Rev. Cancer 2002, 2, 897–909. [Google Scholar] [CrossRef]
- Schneider, G.; Siveke, J.T.; Eckel, F.; Schmid, R.M. Pancreatic cancer: basic and clinical aspects. Gastroenterology 2005, 128, 1606–1625. [Google Scholar] [CrossRef]
- El Maalouf, G.; Le Tourneau, C.; Batty, G.N.; Faivre, S.; Raymond, E. Markers involved in resistance to cytotoxics and targeted therapeutics in pancreatic cancer. Cancer Treat. Rev. 2009, 35, 167–174. [Google Scholar] [CrossRef]
- Kang, S.P.; Saif, M.W. Pharmacogenomics and pancreatic cancer treatment. Optimizing current therapy and individualizing future therapy. JOP: J. Pancreas 2008, 9, 251–266. [Google Scholar]
- Hruban, R.H.; Maitra, A.; Kern, S.E.; Goggins, M. Precursors to pancreatic cancer. Gastroenterol. Clin. North. Am. 2007, 36, 831–849. [Google Scholar] [CrossRef]
- Hruban, R.H.; Wilentz, R.E.; Kern, S.E. Genetic progression in the pancreatic ducts. Am. J. Pathol. 2000, 156, 1821–1825. [Google Scholar] [CrossRef]
- Figueiredo, F.A.; Giovannini, M.; Monges, G.; Charfi, S.; Bories, E.; Pesenti, C.; Caillol, F.; Delpero, J.R. Pancreatic endocrine tumors: a large single-center experience. Pancreas 2009, 38, 936–940. [Google Scholar]
- Dekker, J.; Rossen, J.W.; Buller, H.A.; Einerhand, A.W. The MUC family: an obituary. Trends Biochem. Sci. 2002, 27, 126–131. [Google Scholar] [CrossRef]
- Desseyn, J.L.; Gouyer, V.; Tetaert, D. Architecture of the gel-forming mucins. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 1–16. [Google Scholar]
- Yamazoe, S.; Tanaka, H.; Sawada, T.; Amano, R.; Yamada, N.; Ohira, M.; Hirakawa, K. RNA interference suppression of mucin 5AC (MUC5AC) reduces the adhesive and invasive capacity of human pancreatic cancer cells. J. Exp. Clin. Cancer Res. 2010, 29, 53. [Google Scholar] [CrossRef]
- Jonckheere, N.; Van Seuningen, I. The ever growing family of membrane-bound mucins. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 17–38. [Google Scholar]
- Jonckheere, N.; Van Seuningen, I. The membrane-bound mucins: how large O-glycoproteins play key roles in epithelial cancers and hold promise as biological tools for gene-based and immunotherapies. Crit. Rev. Oncog. 2008, 14, 177–196. [Google Scholar] [CrossRef]
- Jonckheere, N.; Van Seuningen, I. The membrane-bound mucins: From cell signalling to transcriptional regulation and expression in epithelial cancers. Biochimie 2010, 92, 1–11. [Google Scholar] [CrossRef]
- Lan, M.S.; Batra, S.K.; Qi, W.N.; Metzgar, R.S.; Hollingsworth, M.A. Cloning and sequencing of a human pancreatic tumor mucin cDNA. J. Biol. Chem. 1990, 265, 15294–15299. [Google Scholar]
- Ligtenberg, M.J.; Vos, H.L.; Gennissen, A.M.; Hilkens, J. Episialin, a carcinoma-associated mucin, is generated by a polymorphic gene encoding splice variants with alternative amino termini. J. Biol. Chem. 1990, 265, 5573–5578. [Google Scholar]
- Swallow, D.M.; Gendler, S.; Griffiths, B.; Corney, G.; Taylor-Papadimitriou, J.; Bramwell, M.E. The human tumour-associated epithelial mucins are coded by an expressed hypervariable gene locus PUM. Nature 1987, 328, 82–84. [Google Scholar] [CrossRef]
- Hollingsworth, M.A.; Swanson, B.J. Mucins in cancer: protection and control of the cell surface. Nat. Rev. Cancer. 2004, 4, 45–60. [Google Scholar] [CrossRef]
- Meerzaman, D.; Shapiro, P.S.; Kim, K.C. Involvement of the MAP kinase ERK2 in MUC1 mucin signaling. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 281, L86–91. [Google Scholar]
- Ho, J.J.; Kim, Y.S. Serological pancreatic tumor markers and the MUC1 apomucin. Pancreas 1994, 9, 674–691. [Google Scholar] [CrossRef]
- Schroeder, J.A.; Adriance, M.C.; Thompson, M.C.; Camenisch, T.D.; Gendler, S.J. MUC1 alters beta-catenin-dependent tumor formation and promotes cellular invasion. Oncogene 2003, 22, 1324–1332. [Google Scholar] [CrossRef]
- Tsutsumida, H.; Swanson, B.J.; Singh, P.K.; Caffrey, T.C.; Kitajima, S.; Goto, M.; Yonezawa, S.; Hollingsworth, M.A. RNA interference suppression of MUC1 reduces the growth rate and metastatic phenotype of human pancreatic cancer cells. Clin. Cancer Res. 2006, 12, 2976–2987. [Google Scholar] [CrossRef]
- Moniaux, N.; Nollet, S.; Porchet, N.; Degand, P.; Laine, A.; Aubert, J.P. Complete sequence of the human mucin MUC4: a putative cell membrane-associated mucin. Biochem. J. 1999, 338 (Pt 2), 325–333. [Google Scholar]
- Nollet, S.; Moniaux, N.; Maury, J.; Petitprez, D.; Degand, P.; Laine, A.; Porchet, N.; Aubert, J.P. Human mucin gene MUC4: organization of its 5'-region and polymorphism of its central tandem repeat array. Biochem. J. 1998, 332 (Pt 3), 739–748. [Google Scholar]
- Porchet, N.; Nguyen, V.C.; Dufosse, J.; Audie, J.P.; Guyonnet-Duperat, V.; Gross, M.S.; Denis, C.; Degand, P.; Bernheim, A.; Aubert, J.P. Molecular cloning and chromosomal localization of a novel human tracheo-bronchial mucin cDNA containing tandemly repeated sequences of 48 base pairs. Biochem. Biophys. Res. Commun. 1991, 175, 414–422. [Google Scholar] [CrossRef]
- Carraway, K.L.; Theodoropoulos, G.; Kozloski, G.A.; Carothers Carraway, C.A. Muc4/MUC4 functions and regulation in cancer. Future Oncol. 2009, 5, 1631–1640. [Google Scholar] [CrossRef]
- Jepson, S.; Komatsu, M.; Haq, B.; Arango, M.E.; Huang, D.; Carraway, C.A.; Carraway, K.L. Muc4/sialomucin complex, the intramembrane ErbB2 ligand, induces specific phosphorylation of ErbB2 and enhances expression of p27(kip), but does not activate mitogen-activated kinase or protein kinaseB/Akt pathways. Oncogene 2002, 21, 7524–7532. [Google Scholar] [CrossRef]
- Carraway, K.L.; Ramsauer, V.P.; Haq, B.; Carothers Carraway, C.A. Cell signaling through membrane mucins. Bioessays 2003, 25, 66–71. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Singh, A.P.; Chakraborty, S.; Chauhan, S.C.; Bafna, S.; Meza, J.L.; Singh, P.K.; Hollingsworth, M.A.; Mehta, P.P.; Batra, S.K. MUC4 mucin interacts with and stabilizes the HER2 oncoprotein in human pancreatic cancer cells. Cancer Res. 2008, 68, 2065–2070. [Google Scholar] [CrossRef]
- Moniaux, N.; Chaturvedi, P.; Varshney, G.C.; Meza, J.L.; Rodriguez-Sierra, J.F.; Aubert, J.P.; Batra, S.K. Human MUC4 mucin induces ultra-structural changes and tumorigenicity in pancreatic cancer cells. Br. J. Cancer 2007, 97, 345–357. [Google Scholar] [CrossRef]
- Balague, C.; Gambus, G.; Carrato, C.; Porchet, N.; Aubert, J.P.; Kim, Y.S.; Real, F.X. Altered expression of MUC2, MUC4, and MUC5 mucin genes in pancreas tissues and cancer cell lines. Gastroenterology 1994, 106, 1054–1061. [Google Scholar]
- Cano, D.A.; Murcia, N.S.; Pazour, G.J.; Hebrok, M. Orpk mouse model of polycystic kidney disease reveals essential role of primary cilia in pancreatic tissue organization. Development 2004, 131, 3457–3467. [Google Scholar] [CrossRef]
- Pierreux, C.E.; Poll, A.V.; Kemp, C.R.; Clotman, F.; Maestro, M.A.; Cordi, S.; Ferrer, J.; Leyns, L.; Rousseau, G.G.; Lemaigre, F.P. The transcription factor hepatocyte nuclear factor-6 controls the development of pancreatic ducts in the mouse. Gastroenterology 2006, 130, 532–541. [Google Scholar] [CrossRef]
- Kim, G.E.; Bae, H.I.; Park, H.U.; Kuan, S.F.; Crawley, S.C.; Ho, J.J.; Kim, Y.S. Aberrant expression of MUC5AC and MUC6 gastric mucins and sialyl Tn antigen in intraepithelial neoplasms of the pancreas. Gastroenterology 2002, 123, 1052–1060. [Google Scholar] [CrossRef]
- Van Seuningen, I.; Leteurtre, E.; Pigny, P. Mucins in pancreas and hepato-biliary tract. Expression, regulation, biomarkers and therapy. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 233–248. [Google Scholar]
- Swartz, M.J.; Batra, S.K.; Varshney, G.C.; Hollingsworth, M.A.; Yeo, C.J.; Cameron, J.L.; Wilentz, R.E.; Hruban, R.H.; Argani, P. MUC4 expression increases progressively in pancreatic intraepithelial neoplasia. Am. J. Clin. Pathol. 2002, 117, 791–796. [Google Scholar]
- Fauquette, V.; Aubert, S.; Groux-Degroote, S.; Hemon, B.; Porchet, N.; Van Seuningen, I.; Pigny, P. Transcription factor AP-2alpha represses both the mucin MUC4 expression and pancreatic cancer cell proliferation. Carcinogenesis 2007, 28, 2305–2312. [Google Scholar] [CrossRef]
- Iacobuzio-Donahue, C.A.; Ashfaq, R.; Maitra, A.; Adsay, N.V.; Shen-Ong, G.L.; Berg, K.; Hollingsworth, M.A.; Cameron, J.L.; Yeo, C.J.; Kern, S.E.; Goggins, M.; Hruban, R.H. Highly expressed genes in pancreatic ductal adenocarcinomas: a comprehensive characterization and comparison of the transcription profiles obtained from three major technologies. Cancer Res. 2003, 63, 8614–8622. [Google Scholar]
- Moniaux, N.; Junker, W.M.; Singh, A.P.; Jones, A.M.; Batra, S.K. Characterization of human mucin MUC17. Complete coding sequence and organization. J. Biol. Chem. 2006, 281, 23676–23685. [Google Scholar] [CrossRef]
- Adsay, N.V. Role of MUC genes and mucins in pancreatic neoplasia. Am. J. Gastroenterol. 2006, 101, 2330–2332. [Google Scholar] [CrossRef]
- Ito, H.; Endo, T.; Oka, T.; Matumoto, T.; Abe, T.; Toyota, M.; Imai, K.; Satoh, M.; Maguchi, H.; Shinohara, T. Mucin expression profile is related to biological and clinical characteristics of intraductal papillary-mucinous tumors of the pancreas. Pancreas 2005, 30, e96–102. [Google Scholar] [CrossRef]
- Adsay, N.V.; Merati, K.; Basturk, O.; Iacobuzio-Donahue, C.; Levi, E.; Cheng, J.D.; Sarkar, F.H.; Hruban, R.H.; Klimstra, D.S. Pathologically and biologically distinct types of epithelium in intraductal papillary mucinous neoplasms: delineation of an "intestinal" pathway of carcinogenesis in the pancreas. Am. J. Surg. Pathol. 2004, 28, 839–848. [Google Scholar] [CrossRef]
- Nakamura, A.; Horinouchi, M.; Goto, M.; Nagata, K.; Sakoda, K.; Takao, S.; Imai, K.; Kim, Y.S.; Sato, E.; Yonezawa, S. New classification of pancreatic intraductal papillary-mucinous tumour by mucin expression: its relationship with potential for malignancy. J. Pathol. 2002, 197, 201–210. [Google Scholar] [CrossRef]
- Luttges, J.; Feyerabend, B.; Buchelt, T.; Pacena, M.; Kloppel, G. The mucin profile of noninvasive and invasive mucinous cystic neoplasms of the pancreas. Am. J. Surg. Pathol. 2002, 26, 466–471. [Google Scholar] [CrossRef]
- Terada, T.; Ohta, T.; Sasaki, M.; Nakanuma, Y.; Kim, Y.S. Expression of MUC apomucins in normal pancreas and pancreatic tumours. J. Pathol. 1996, 180, 160–165. [Google Scholar] [CrossRef]
- Perrais, M.; Pigny, P.; Ducourouble, M.P.; Petitprez, D.; Porchet, N.; Aubert, J.P.; Van Seuningen, I. Characterization of human mucin gene MUC4 promoter: importance of growth factors and proinflammatory cytokines for its regulation in pancreatic cancer cells. J. Biol. Chem. 2001, 276, 30923–30933. [Google Scholar] [CrossRef]
- Jonckheere, N.; Fauquette, V.; Stechly, L.; Saint-Laurent, N.; Aubert, S.; Susini, C.; Huet, G.; Porchet, N.; Van Seuningen, I.; Pigny, P. Tumour growth and resistance to gemcitabine of pancreatic cancer cells are decreased by AP-2alpha overexpression. Br. J. Cancer 2009, 101, 637–644. [Google Scholar] [CrossRef]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Fauquette, V.; Perrais, M.; Cerulis, S.; Jonckheere, N.; Ducourouble, M.P.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. The antagonistic regulation of human MUC4 and ErbB-2 genes by the Ets protein PEA3 in pancreatic cancer cells: implications for the proliferation/differentiation balance in the cells. Biochem. J. 2005, 386, 35–45. [Google Scholar] [CrossRef]
- Andrianifahanana, M.; Singh, A.P.; Nemos, C.; Ponnusamy, M.P.; Moniaux, N.; Mehta, P.P.; Varshney, G.C.; Batra, S.K. IFN-gamma-induced expression of MUC4 in pancreatic cancer cells is mediated by STAT-1 upregulation: a novel mechanism for IFN-gamma response. Oncogene 2007, 26, 7251–7261. [Google Scholar] [CrossRef]
- Akhurst, R.J.; Derynck, R. TGF-beta signaling in cancer--a double-edged sword. Trends Cell. Biol. 2001, 11, S44–51. [Google Scholar]
- Choudhury, A.; Singh, R.K.; Moniaux, N.; El-Metwally, T.H.; Aubert, J.P.; Batra, S.K. Retinoic acid-dependent transforming growth factor-beta 2-mediated induction of MUC4 mucin expression in human pancreatic tumor cells follows retinoic acid receptor-alpha signaling pathway. J. Biol. Chem. 2000, 275, 33929–33936. [Google Scholar]
- Jonckheere, N.; Perrais, M.; Mariette, C.; Batra, S.K.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. A role for human MUC4 mucin gene, the ErbB2 ligand, as a target of TGF-beta in pancreatic carcinogenesis. Oncogene 2004, 23, 5729–5738. [Google Scholar] [CrossRef]
- Andrianifahanana, M.; Agrawal, A.; Singh, A.P.; Moniaux, N.; van Seuningen, I.; Aubert, J.P.; Meza, J.; Batra, S.K. Synergistic induction of the MUC4 mucin gene by interferon-gamma and retinoic acid in human pancreatic tumour cells involves a reprogramming of signalling pathways. Oncogene 2005, 24, 6143–6154. [Google Scholar] [CrossRef]
- McWilliams, R.; Highsmith, W.E.; Rabe, K.G.; de Andrade, M.; Tordsen, L.A.; Holtegaard, L.M.; Petersen, G.M. Cystic fibrosis transmembrane regulator gene carrier status is a risk factor for young onset pancreatic adenocarcinoma. Gut 2005, 54, 1661–1662. [Google Scholar] [CrossRef]
- Singh, A.P.; Chauhan, S.C.; Andrianifahanana, M.; Moniaux, N.; Meza, J.L.; Copin, M.C.; van Seuningen, I.; Hollingsworth, M.A.; Aubert, J.P.; Batra, S.K. MUC4 expression is regulated by cystic fibrosis transmembrane conductance regulator in pancreatic adenocarcinoma cells via transcriptional and post-translational mechanisms. Oncogene 2007, 26, 30–41. [Google Scholar] [CrossRef]
- Kato, S.; Hokari, R.; Crawley, S.; Gum, J.; Ahn, D.H.; Kim, J.W.; Kwon, S.W.; Miura, S.; Basbaum, C.B.; Kim, Y.S. MUC5AC mucin gene regulation in pancreatic cancer cells. Int. J. Oncol. 2006, 29, 33–40. [Google Scholar]
- Ho, J.J.; Crawley, S.; Pan, P.L.; Farrelly, E.R.; Kim, Y.S. Secretion of MUC5AC mucin from pancreatic cancer cells in response to forskolin and VIP. Biochem. Biophys. Res. Commun. 2002, 294, 680–686. [Google Scholar] [CrossRef]
- Prasad, N.B.; Biankin, A.V.; Fukushima, N.; Maitra, A.; Dhara, S.; Elkahloun, A.G.; Hruban, R.H.; Goggins, M.; Leach, S.D. Gene expression profiles in pancreatic intraepithelial neoplasia reflect the effects of Hedgehog signaling on pancreatic ductal epithelial cells. Cancer Res. 2005, 65, 1619–1626. [Google Scholar] [CrossRef]
- Jonckheere, N.; Van Der Sluis, M.; Velghe, A.; Buisine, M.P.; Sutmuller, M.; Ducourouble, M.P.; Pigny, P.; Buller, H.A.; Aubert, J.P.; Einerhand, A.W.; Van Seuningen, I. Transcriptional activation of the murine Muc5ac mucin gene in epithelial cancer cells by TGF-beta/Smad4 signalling pathway is potentiated by Sp1. Biochem. J. 2004, 377, 797–808. [Google Scholar] [CrossRef]
- Jonckheere, N.; Vincent, A.; Perrais, M.; Ducourouble, M.P.; Male, A.K.; Aubert, J.P.; Pigny, P.; Carraway, K.L.; Freund, J.N.; Renes, I.B.; Van Seuningen, I. The human mucin MUC4 is transcriptionally regulated by caudal-related homeobox, hepatocyte nuclear factors, forkhead box A, and GATA endodermal transcription factors in epithelial cancer cells. J. Biol. Chem. 2007, 282, 22638–22650. [Google Scholar] [CrossRef]
- van der Sluis, M.; Melis, M.H.; Jonckheere, N.; Ducourouble, M.P.; Buller, H.A.; Renes, I.; Einerhand, A.W.; Van Seuningen, I. The murine Muc2 mucin gene is transcriptionally regulated by the zinc-finger GATA-4 transcription factor in intestinal cells. Biochem. Biophys. Res. Commun. 2004, 325, 952–960. [Google Scholar] [CrossRef]
- van der Sluis, M.; Vincent, A.; Bouma, J.; Korteland-Van Male, A.; van Goudoever, J.B.; Renes, I.B.; Van Seuningen, I. Forkhead box transcription factors Foxa1 and Foxa2 are important regulators of Muc2 mucin expression in intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2008, 369, 1108–1113. [Google Scholar] [CrossRef]
- Russo, V.E.A.; Martienssen, R.A.; Riggs, A.D. Introduction. In Epigenetic Mechanisms of Gene Regulation; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1996; pp. 1–4. [Google Scholar]
- Van Seuningen, I. What is epigenetics? Strategies for research and application to cancer. Ann. Pathol. 2009, 29 (Spec No 1), S28–30. [Google Scholar] [CrossRef]
- Vincent, A.; Van Seuningen, I. Mucins, epigenetics and cancer. In The Epithelial Mucins: Structure/Function. Roles in Cancer and Inflammatory Diseases; Van Seuningen, I., Ed.; Research Signpost: Kerala, India, 2008; pp. 95–108. [Google Scholar]
- Van Seuningen, I.; Vincent, A. Mucins: A new family of epigenetic biomarkers in epithelial cancers. Expert Opinion on Medical Diagnostics 2009, 3, 411–427. [Google Scholar] [CrossRef]
- Yamada, N.; Nishida, Y.; Tsutsumida, H.; Hamada, T.; Goto, M.; Higashi, M.; Nomoto, M.; Yonezawa, S. MUC1 expression is regulated by DNA methylation and histone H3 lysine 9 modification in cancer cells. Cancer Res. 2008, 68, 2708–2716. [Google Scholar] [CrossRef]
- Vincent, A.; Ducourouble, M.P.; Van Seuningen, I. Epigenetic regulation of the human mucin gene MUC4 in epithelial cancer cell lines involves both DNA methylation and histone modifications mediated by DNA methyltransferases and histone deacetylases. Faseb. J. 2008, 22, 3035–3045. [Google Scholar] [CrossRef]
- Yamada, N.; Nishida, Y.; Tsutsumida, H.; Goto, M.; Higashi, M.; Nomoto, M.; Yonezawa, S. Promoter CpG methylation in cancer cells contributes to the regulation of MUC4. Br. J. Cancer 2009, 100, 344–351. [Google Scholar] [CrossRef]
- de Bustros, A.; Nelkin, B.D.; Silverman, A.; Ehrlich, G.; Poiesz, B.; Baylin, S.B. The short arm of chromosome 11 is a "hot spot" for hypermethylation in human neoplasia. Proc. Natl. Acad. Sci. USA 1988, 85, 5693–5697. [Google Scholar]
- Ho, J.J.; Han, S.W.; Pan, P.L.; Deng, G.; Kuan, S.F.; Kim, Y.S. Methylation status of promoters and expression of MUC2 and MUC5AC mucins in pancreatic cancer cells. Int. J. Oncol. 2003, 22, 273–279. [Google Scholar]
- Vincent, A.; Perrais, M.; Desseyn, J.L.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. Epigenetic regulation (DNA methylation, histone modifications) of the 11p15 mucin genes (MUC2, MUC5AC, MUC5B, MUC6) in epithelial cancer cells. Oncogene 2007, 26, 6566–6576. [Google Scholar] [CrossRef]
- Hamada, T.; Goto, M.; Tsutsumida, H.; Nomoto, M.; Higashi, M.; Sugai, T.; Nakamura, S.; Yonezawa, S. Mapping of the methylation pattern of the MUC2 promoter in pancreatic cancer cell lines, using bisulfite genomic sequencing. Cancer Lett. 2005, 227, 175–184. [Google Scholar] [CrossRef]
- Yamada, N.; Nishida, Y.; Yokoyama, S.; Tsutsumida, H.; Houjou, I.; Kitamoto, S.; Goto, M.; Higashi, M.; Yonezawa, S. Expression of MUC5AC, an early marker of pancreatobiliary cancer, is regulated by DNA methylation in the distal promoter region in cancer cells. J. Hepatobiliary. Pancreat. Sci. 2010, 17, 844–854. [Google Scholar] [CrossRef]
- Yamada, N.; Hamada, T.; Goto, M.; Tsutsumida, H.; Higashi, M.; Nomoto, M.; Yonezawa, S. MUC2 expression is regulated by histone H3 modification and DNA methylation in pancreatic cancer. Int. J. Cancer 2006, 119, 1850–1857. [Google Scholar] [CrossRef]
- Jin, C.; Rajabi, H.; Kufe, D. miR-1226 targets expression of the mucin 1 oncoprotein and induces cell death. Int. J. Oncol. 2010, 37, 61–69. [Google Scholar]
- Rajabi, H.; Jin, C.; Ahmad, R.; McClary, C.; Joshi, M.D.; Kufe, D. Mucin 1 Oncoprotein Expression Is Suppressed by the miR-125b Oncomir. Genes Cancer 2010, 1, 62–68. [Google Scholar] [CrossRef]
- Sachdeva, M.; Mo, Y.Y. MicroRNA-145 suppresses cell invasion and metastasis by directly targeting mucin 1. Cancer Res. 2010, 70, 378–387. [Google Scholar] [CrossRef]
- Fry, L.C.; Monkemuller, K.; Malfertheiner, P. Molecular markers of pancreatic cancer: development and clinical relevance. Langenbecks Arch. Surg. 2008, 393, 883–890. [Google Scholar] [CrossRef]
- Jhala, N.; Jhala, D.; Vickers, S.M.; Eltoum, I.; Batra, S.K.; Manne, U.; Eloubeidi, M.; Jones, J.J.; Grizzle, W.E. Biomarkers in Diagnosis of pancreatic carcinoma in fine-needle aspirates. Am. J. Clin. Pathol. 2006, 126, 572–579. [Google Scholar] [CrossRef]
- Gold, D.V.; Karanjawala, Z.; Modrak, D.E.; Goldenberg, D.M.; Hruban, R.H. PAM4-reactive MUC1 is a biomarker for early pancreatic adenocarcinoma. Clin. Cancer Res. 2007, 13, 7380–7387. [Google Scholar] [CrossRef]
- Haab, B.B.; Porter, A.; Yue, T.; Li, L.; Scheiman, J.; Anderson, M.A.; Barnes, D.; Schmidt, C.M.; Feng, Z.; Simeone, D.M. Glycosylation variants of mucins and CEACAMs as candidate biomarkers for the diagnosis of pancreatic cystic neoplasms. Ann. Surg. 2010, 251, 937–945. [Google Scholar] [CrossRef]
- Tang, C.K.; Apostolopoulos, V. Strategies used for MUC1 immunotherapy: preclinical studies. Expert Rev. Vaccines 2008, 7, 951–962. [Google Scholar] [CrossRef]
- Tang, C.K.; Katsara, M.; Apostolopoulos, V. Strategies used for MUC1 immunotherapy: human clinical studies. Expert Rev. Vaccines 2008, 7, 963–975. [Google Scholar] [CrossRef]
- Bitler, B.G.; Menzl, I.; Huerta, C.L.; Sands, B.; Knowlton, W.; Chang, A.; Schroeder, J.A. Intracellular MUC1 peptides inhibit cancer progression. Clin. Cancer Res. 2009, 15, 100–109. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Q.; Qin, R.; Le, H.; Xia, R.; Li, W.; Kumar, M. Amplification and functional characterization of MUC1 promoter and gene-virotherapy via a targeting adenoviral vector expressing hSSTR2 gene in MUC1-positive Panc-1 pancreatic cancer cells in vitro. Int. J. Mol. Med. 2005, 15, 617–626. [Google Scholar]
- Wei, J.; Gao, W.; Wu, J.; Meng, K.; Zhang, J.; Chen, J.; Miao, Y. Dendritic cells expressing a combined PADRE/MUC4-derived polyepitope DNA vaccine induce multiple cytotoxic T-cell responses. Cancer Biother. Radiopharm. 2008, 23, 121–128. [Google Scholar] [CrossRef]
- Bafna, S.; Kaur, S.; Momi, N.; Batra, S.K. Pancreatic cancer cells resistance to gemcitabine: the role of MUC4 mucin. Br. J. Cancer 2009, 101, 1155–1161. [Google Scholar] [CrossRef]
- Mimeault, M.; Johansson, S.L.; Senapati, S.; Momi, N.; Chakraborty, S.; Batra, S.K. MUC4 down-regulation reverses chemoresistance of pancreatic cancer stem/progenitor cells and their progenies. Cancer Lett. 2010, in press. [Google Scholar]
- Kalra, A.V.; Campbell, R.B. Mucin impedes cytotoxic effect of 5-FU against growth of human pancreatic cancer cells: overcoming cellular barriers for therapeutic gain. Br. J. Cancer 2007, 97, 910–918. [Google Scholar] [CrossRef]
- Kalra, A.V.; Campbell, R.B. Mucin overexpression limits the effectiveness of 5-FU by reducing intracellular drug uptake and antineoplastic drug effects in pancreatic tumours. Eur. J. Cancer 2008, 45, 164–173. [Google Scholar] [CrossRef]
- Tinder, T.L.; Subramani, D.B.; Basu, G.D.; Bradley, J.M.; Schettini, J.; Million, A.; Skaar, T.; Mukherjee, P. MUC1 enhances tumor progression and contributes toward immunosuppression in a mouse model of spontaneous pancreatic adenocarcinoma. J. Immunol. 2008, 181, 3116–3125. [Google Scholar]
- Spicer, A.P.; Duhig, T.; Chilton, B.S.; Gendler, S.J. Analysis of mammalian MUC1 genes reveals potential functionally important domains. Mamm. Genome 1995, 6, 885–888. [Google Scholar] [CrossRef]
- Lalani, E.N.; Berdichevsky, F.; Boshell, M.; Shearer, M.; Wilson, D.; Stauss, H.; Gendler, S.J.; Taylor-Papadimitriou, J. Expression of the gene coding for a human mucin in mouse mammary tumor cells can affect their tumorigenicity. J. Biol. Chem. 1991, 266, 15420–15426. [Google Scholar]
- Aguirre, A.J.; Bardeesy, N.; Sinha, M.; Lopez, L.; Tuveson, D.A.; Horner, J.; Redston, M.S.; DePinho, R.A. Activated Kras and Ink4a/Arf deficiency cooperate to produce metastatic pancreatic ductal adenocarcinoma. Genes Dev. 2003, 17, 3112–3126. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Petricoin, E.F.; Maitra, A.; Rajapakse, V.; King, C.; Jacobetz, M.A.; Ross, S.; Conrads, T.P.; Veenstra, T.D.; Hitt, B.A.; Kawaguchi, Y.; Johann, D.; Liotta, L.A.; Crawford, H.C.; Putt, M.E.; Jacks, T.; Wright, C.V.; Hruban, R.H.; Lowy, A.M.; Tuveson, D.A. Preinvasive and invasive ductal pancreatic cancer and its early detection in the mouse. Cancer Cell 2003, 4, 437–450. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef]
- Izeradjene, K.; Combs, C.; Best, M.; Gopinathan, A.; Wagner, A.; Grady, W.M.; Deng, C.X.; Hruban, R.H.; Adsay, N.V.; Tuveson, D.A.; Hingorani, S.R. Kras(G12D) and Smad4/Dpc4 haploinsufficiency cooperate to induce mucinous cystic neoplasms and invasive adenocarcinoma of the pancreas. Cancer Cell 2007, 11, 229–243. [Google Scholar] [CrossRef]
- Velcich, A.; Yang, W.; Heyer, J.; Fragale, A.; Nicholas, C.; Viani, S.; Kucherlapati, R.; Lipkin, M.; Yang, K.; Augenlicht, L. Colorectal cancer in mice genetically deficient in the mucin Muc2. Science 2002, 295, 1726–1729. [Google Scholar] [CrossRef]
- Cheon, D.J.; Wang, Y.; Deng, J.M.; Lu, Z.; Xiao, L.; Chen, C.M.; Bast, R.C.; Behringer, R.R. CA125/MUC16 is dispensable for mouse development and reproduction. PLoS One 2009, 4, e4675. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jonckheere, N.; Skrypek, N.; Van Seuningen, I. Mucins and Pancreatic Cancer. Cancers 2010, 2, 1794-1812. https://doi.org/10.3390/cancers2041794
Jonckheere N, Skrypek N, Van Seuningen I. Mucins and Pancreatic Cancer. Cancers. 2010; 2(4):1794-1812. https://doi.org/10.3390/cancers2041794
Chicago/Turabian StyleJonckheere, Nicolas, Nicolas Skrypek, and Isabelle Van Seuningen. 2010. "Mucins and Pancreatic Cancer" Cancers 2, no. 4: 1794-1812. https://doi.org/10.3390/cancers2041794
APA StyleJonckheere, N., Skrypek, N., & Van Seuningen, I. (2010). Mucins and Pancreatic Cancer. Cancers, 2(4), 1794-1812. https://doi.org/10.3390/cancers2041794