DNA Methylation in Thyroid Tumorigenesis

Abstract
: Thyroid cancer is the most common endocrine cancer with 1,690 deaths each year. There are four main types of which the papillary and follicular types together account for >90% followed by medullary cancers with 3% to 5% and anaplastic carcinomas making up <3%. Epigenetic events of DNA hypermethylation are emerging as promising molecular targets for cancer detection. Our immediate and long term goal is to identify DNA methylation markers for early detection of thyroid cancer. This pilot study comprised of 21 patients to include 11 papillary thyroid cancers (PTC), 2 follicular thyroid cancers (FTC), 5 normal thyroid cases, and 3 hyperthyroid cases. Aberrant promoter methylation was examined in 24 tumor suppressor genes using the methylation specific multiplex ligation-dependent probe amplification (MS-MLPA) assay and in the NIS gene using methylation-specific PCR (MSP). The frequently methylated genes were CASP8 (17/21), RASSF1 (16/21) and NIS (9/21). In the normal samples, CASP8, RASSF1 and NIS were methylated in 5/5, 4/5 and 1/5 respectively. In the hyperthyroid samples, CASP8, RASSF1 and NIS were methylated in 3/3, 2/3 and 1/3 respectively. In the thyroid cancers, CASP8, RASSF1, and NIS were methylated in 9/13, 10/13, and 7/13 respectively. CASP8, RASSF1 and NIS were also methylated in concurrently present normal thyroid tissue in 3/11, 4/11 and 3/11 matched thyroid cancer cases (matched for presence of both normal thyroid tissue and thyroid cancer), respectively. Our data suggests that aberrant methylation of CASP8, RASSF1, and NIS maybe an early change in thyroid tumorigenesis regardless of cell type.1. Introduction
The National Cancer Institute [1] estimated that in 2010 there would be 44,670 new cases of thyroid cancer (TC) with 1,690 deaths. Compared to other adult cancers, TC tends to occur in younger people between the ages of 20 and 60 years [2]. It is three times more common in women than men [2], has the fastest rising incidence rates in women, and the second fastest in men with an annual percentage change of approximately 5%, making TC the sixth most common cancer in women [3]. Reasons for this trend have been attributed to improvement in imaging (ultrasound technology) that is allowing the identification of ever smaller thyroid nodules. However, with this gain in detection, determining which benign nodules (adenomas) will progress to cancer cannot be determined on the basis of histology alone, underscoring the need for genetic markers of early detection for TC.
There are four main types of thyroid cancers: papillary, follicular, medullary and anaplastic. The papillary and follicular types, also known as differentiated thyroid cancers, account for >90% of thyroid carcinomas, followed by medullary cancers with 3–5% and anaplastic carcinomas with <3% [4]. Papillary thyroid cancers (PTC) are the most common and have the best prognosis. Follicular thyroid carcinomas (FTC) are the next most common type of thyroid cancer and have a slightly worse prognosis than PTC. The cells in follicular carcinomas can accumulate radioactive iodine which can be used in its treatment. Hürthle cell carcinoma, also known as oxyphil cell carcinoma, is a variant of follicular carcinoma but has a worse outcome. It is less likely to concentrate radioactive iodine [4] making it harder to detect and treat. Besides age, other risk factors for developing thyroid cancer include diets low in iodine, radiation exposure and certain hereditary conditions like familial medullary thyroid carcinoma, multiple endocrine neoplasia type 2A (MEN-2A) and multiple endocrine neoplasia type 2B (MEN-2B).
Until recently, thyroid cancer has been largely ignored in the US because the majority of thyroid cancer cases have very good prognosis, with five-year survival rates of approximately 96% when submitted to timely and appropriate treatment [3,5]. However, those diagnosed at late stages and presenting with advanced diseases have a devastating five-year survival rate of under 60% [6]. The recurrence rate of thyroid cancer is high (30%) [7], and only one third of patients with distant metastases respond to radioactive iodine (131I) therapy with complete remission [8]. Thus, despite advances on scientific and clinical fronts, advanced thyroid cancers remain incurable. The present challenge is to obtain a highly accurate diagnostic test for early stage thyroid cancers and effective molecular-directed therapies for advanced thyroid cancer.
Early detection of cancers before metastasis is important for patients and clinicians as it improves prognosis, patient quality of life and provides additional treatment options. One of the best ways for early detection of cancer is through the use biomarkers. These can be DNA, mRNA, proteins, metabolites, or processes such as apoptosis, angiogenesis or proliferation [9]. These markers of early tumor development can be produced by either the tumor or by other tissues, so they are present in all types of body fluids, tissues or cell lines. Tumor markers include hormones, functional subgroups of proteins (like enzymes, glycoproteins, and receptors), molecular markers (such as genetic mutations, amplifications or translocations) [10] and epigenetic markers (like tumor suppressor gene hypermethylation) [11]. Beyond their screening and diagnostic value, biomarkers can be used to estimate tumor volume, evaluate response to treatment, assess disease recurrence, or as prognostic indicators of disease progression [10].
Over the past 15 years, the application of molecular technologies has focused on genetic events. New approaches are necessary to identify potential novel diagnostic and prognostic markers, which would allow accurate early diagnosis, with personalized clinical management and surveillance. Epigenetic silencing through aberrant DNA methylation of tumor suppressor genes has been reported in TC [11]. Methylation of TSHR, ECAD, NIS-L, ATM and DAPK are frequent in PTC [12]. Increased methylation of several tumor suppressor genes (TIMP3, SLC5A8, DAPK, and RARB2) is associated with features of PTC aggressiveness [11,13]. Hypermethylation of RASSF1A, a known tumor suppressor gene, has been described in 75% (9 of 12) of follicular thyroid cancers as well as in a smaller percentage of benign adenomas (44%), and papillary thyroid cancers (20%) [14] indicating that this may be an early step in follicular cell derived thyroid tumorigenesis.
Our immediate and long-term goal is to identify DNA methylation markers for early detection of thyroid cancer. In this exploratory study, we examined promoter hypermethylation in 24 tumor suppressor genes using the methylation-specific multiplex ligation-dependent probe amplification (MS-MLPA) assay and in the NIS gene using methylation-specific PCR (MSP).
2. Results and Discussion
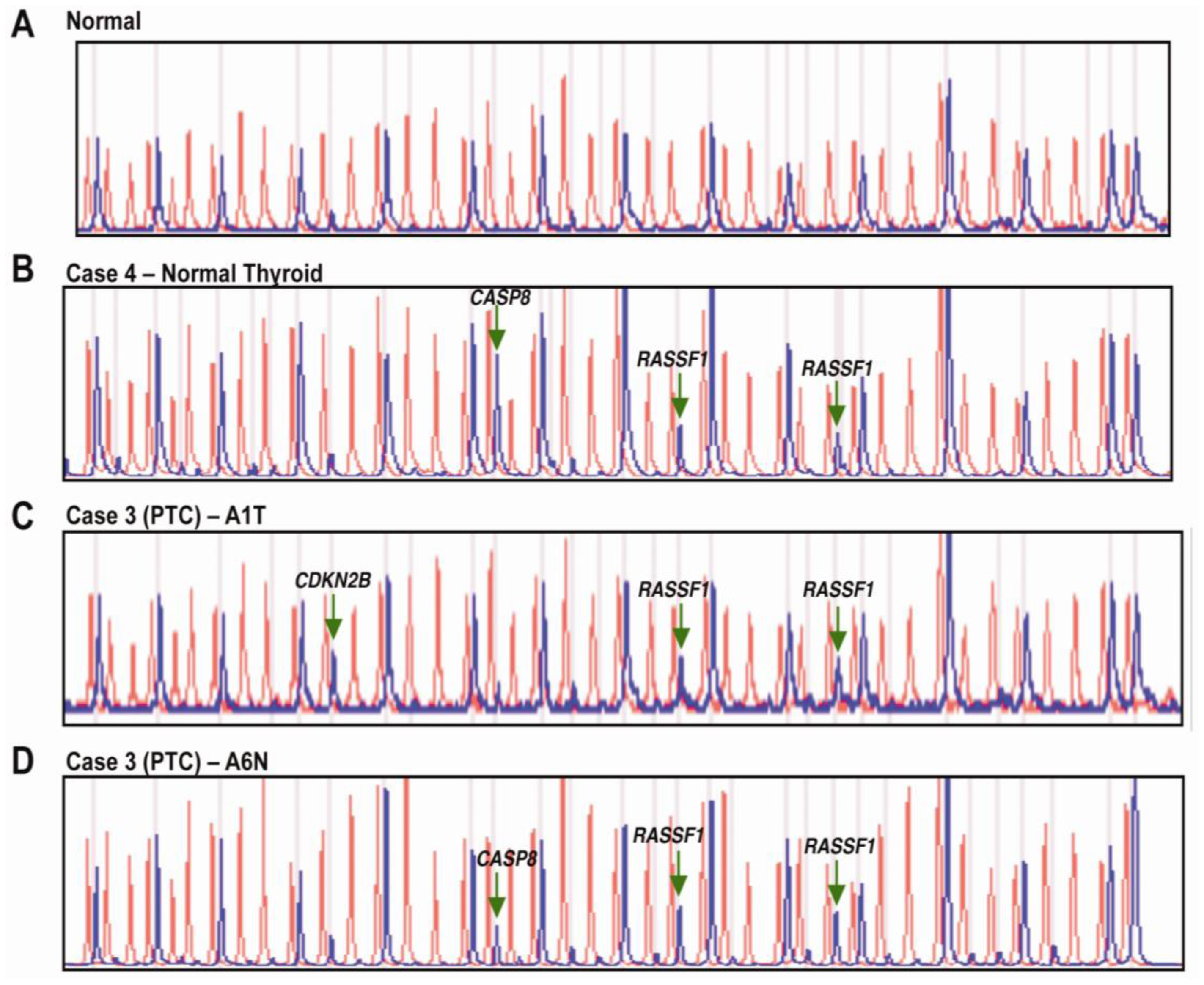
In this study, CASP8 and RASSF1 were the only two genes found to be aberrantly methylated from the MS-MLPA panel of 24 tumor suppressor genes. Additionally, the NIS gene was selected to be examined by MSP as it is not present in the MS-MLPA gene panel, is known to be methylated in PTC [12], and is a thyroid-specific gene that plays a role in iodine uptake and normal thyroid cell function [15-17]. Aberrant methylation of CASP8 was most frequent occurring in 17/21 samples, followed by RASSF1 in 16/21 and NIS in 9/21. Methylation of CASP8 was observed in all 5 normal samples, all 3 hyperthyroid samples and in 3/11 concurrent thyroid cancer with normal thyroid lesions. This was followed by RASSF1 in 4/5 normal samples, 2/3 hyperthyroid samples and in 4/11 concurrent thyroid cancer with normal thyroid lesions. NIS methylation occurred in 1 normal thyroid sample, 1 hyperthyroid sample and in 3/11 concurrent thyroid cancer with normal thyroid lesions.
Proper management of TC depends on accurate pathological diagnosis and although most thyroid nodules are benign, distinguishing thyroid cancer from benign lesions is crucial for appropriate treatment and follow-up. Fine-needle aspiration (FNA) biopsy is the standard diagnostic tool for preoperative diagnosis [18]. The FNA biopsy allows diagnosis of the nature of thyroid nodules in a majority of cases, but it has some limitations, especially in the presence of follicular lesions where it can be difficult to distinguish between a follicular adenoma (FA) and FTC because they are cytologically indistinguishable [19]. About 20% of FNA biopsies performed for nodular thyroid lesions in USA are reported as inconclusive or suspicious for malignancy [19]. A majority of these inconclusive cases (70%) [20] are benign. A single false-negative FNA can delay surgical treatment by 28 months even with clinical evidence suggesting malignancy, leading to higher rates of vascular and capsular invasion and persistent disease at follow up which is in the advanced stage making it incurable [21]. Even amongst PTC, where diagnosis is straightforward on FNA, the tall cell variant (TCV), which has a worse prognosis, can be misdiagnosed [22,23]. FNA has been found to be less sensitive in detecting FTC and Hürthle cell carcinomas when compared to PTC [21]. The diagnosis of PTC, MTC and anaplastic thyroid cancer by FNA biopsy is straightforward in that it is based on nuclear changes that are specific to each of these types of cancers, whereas the diagnosis of FA and FTC is based on the presence of capsular and vascular invasion which cannot be seen on FNA [24]. Only on surgical removal can FA be differentiated from FTC. Hence, there is a need for reliable, novel preoperative markers to help differentiate benign and malignant thyroid nodules.
In this exploratory study methylation of CASP8 is the most frequent (17/21 cases) and appears to be an early change since it was present in all 5 normal samples, all 3 hyperthyroid samples and in 3/11 concurrent thyroid cancer with normal thyroid lesions.
Methylation of CASP8 in thyroid cancer is relatively new. It has been reported in childhood medulloblastoma and neuroblastoma [25,26], neuroendocrine lung tumors [27] and breast cancer [28]. The gene is located on chromosome 2q33, encodes a member of the caspase family, Caspase-8, which is an apical caspase acting in the death receptor–ligand interaction–induced apoptotic process [29]. The absence or downregulation of CASP8 may be attributed to epigenetic changes, such as hypermethylation, or mutations [30,31] The promoter region has binding sites for p53, nuclear factor-κB (NF-κB), AP-1, SP-1, IRF-1, and Ets-like transcription factors [32]. CASP8 functions both as a key molecule for apoptosis as well as a selective signal transducer [33]. As an important initiator of apoptosis [34], loss or reduced expression supports its role in tumorigenesis. The cytotoxic drugs 5-fluorouracil (5-Fu) and methotrexate, used to treat some cancers, have been shown to upregulate CASP8 and induce cell apoptosis in multiple cancers including breast cancer [35,36]. A study by Wu et al. [28] found that breast cancer cells treated with 5-Fu induced cell apoptosis by demethylation of the CASP8 promoter region resulting in a significant increase in its mRNA expression.
RASSF1 was the next most frequently methylated gene, detected in 16 of the 21 cases. Among the 16 cases, 10 cases demonstrated RASSF1 methylation as an early change. It was methylated in 4/5 normal samples, 2/3 hyperthyroid samples and in 4/11 concurrent thyroid cancer with normal thyroid lesions. RASSF1 encodes a signaling protein that functions through a pathway involving Ras which is a component of the phosphatidylinositol 3-kinase (PI3K)/Akt pathway [11]. Mutational inactivation of this gene is very rare (<2%), and the main mechanism of its inactivation is through promoter methylation and LOH [37]. RASSF1 has been found to be methylated in benign thyroid lesions and thyroid cancers, including PTC, FTC, and ATC [14,37].
Methylation of thyroid-specific genes, such as those for sodium/iodide symporter (NIS) and thyroid-stimulating hormone receptor, is a cause for the failure of clinical radioiodine treatment of thyroid cancer [38,39]. Methylation of NIS has severe treatment implications in TC. The function of the thyroid is to produce thyroid hormones, through the uptake and concentration of iodine, to meet the body's normal metabolic needs. Thyroid specific genes, thyroid peroxidase (TPO), thyroglobulin [40], TSH receptor (TSHR) and NIS, all play a role in iodine uptake and normal thyroid cell function [15-17]. The cornerstone of follow-up and treatment of patients with thyroid cancer is by the measurement of iodide uptake by thyroid cancer cells. However, radioiodide uptake is found only in about 67% of patients with persistent or recurrent disease. Several studies have demonstrated a decrease in or a loss of NIS expression in primary human thyroid carcinomas [41,42]. Immunohistochemical studies have confirmed this considerably decreased expression of the NIS protein in thyroid cancer tissues, suggesting that low expression of NIS may represent an early abnormality in the pathway of thyroid cell transformation, rather than being a consequence of cancer progression [43]. NIS methylation has been reported in advanced stage PTC but was not detected in adjacent benign tissue [12]. In this study, NIS methylation occurred in 1 normal thyroid sample, 1 hyperthyroid sample and in 3/11 concurrent thyroid cancer with normal thyroid lesions.
A distinct advantage of MS-MLPA is the ability to examine aberrant promoter methylation in multiple cancer genes in a single assay using formalin-fixed paraffin embedded tissue. This is because probes added to the samples are amplified and quantified instead of target nucleic acids. The probe target sequences are small (50-70 nucleotides), making it ideal for fragmented formalin-fixed paraffin embedded tissue DNA. When compared to MSP, the MS-MLPA assay requires minimal sample preparation and very small quantities of DNA (20 ng). It is extremely easy to perform and is an inexpensive one-tube assay compared to array based technologies. MS-MLPA has been validated in other studies by our group [44,45] with a high degree of sensitivity and specificity as well as by MRC Holland [46], the makers of MS-MLPA. MRC Holland demonstrated the sensitivity of MS-MLPA by analyzing human sample DNA methylated in vitro which resulted in amplification products for all probes. Specificity was demonstrated by noting that all MS-MLPA probes that recognized a HhaI site within a CpG island did not produce amplification products after HhaI digestion. MS-MLPA profiling of multiple genes for aberrantly methylated promoter regions is a valuable screening tool to determine frequency and pattern of promoter methylation in neoplasia. These epigenetic signatures, upon subsequent validation as diagnostic or prognostic biomarkers, can become reduced to a more definitive candidate gene panel of only a few key genes.
CASP8, RASSF1and NIS, were found to be consistently methylated in normal thyroid, hyperthyroid lesions, as well as in those thyroid cancer cases with matched normal and tumor blocks from the same biopsy specimen and likely represent early changes in thyroid tumorigenesis.
3. Experimental Section
3.1. Cohort
The cohort of 21 patients consisted of 11 papillary thyroid cancers (PTC), 2 follicular thyroid cancers (FTC), 5 normal thyroid cases and 3 hyperthyroid cases. Of the 2 FTC, one was a poorly differentiated follicular (insular) carcinoma and the other was a Hürthle cell carcinoma. Thyroid cancer specimens were matched for normal and tumor lesions in 11 of the 13 cases where normal and tumor blocks were taken from the same biopsy specimen. The five histologically normal thyroid tissues were from patients who had their thyroid removed at the time of surgery for laryngeal cancers. The 3 cases of hyperthyroidism were due to Graves' disease.
3.2. DNA Extraction
Whole 5 micron tissue sections or microdissected thyroid lesions and adjacent normal when present were processed for DNA extraction as previously described [47].
3.3. Methylation-Specific Multiplex Ligation-dependent Probe Amplification (MS-MLPA) Assay
MS-MLPA, a modification of the conventional MLPA assay, allows for the simultaneous detection of changes in methylation status as well as copy number changes of approximately 41 different DNA sequences in a single reaction requiring only 20 ng of human DNA [46,48,49].
Briefly, the MS-MLPA panel in the presence of HhaI detects aberrant promoter hypermethylation by taking advantage of a HhaI site in the gene probes of interest. The control gene probes, without a HhaI site, serve as undigested controls. A normal control DNA sample will generate 41 individual peaks for all probes in the absence of HhaI and 15 separate peaks in the presence of HhaI (Figure 1A and Figure 2A). Normal controls for methylation assays are run using DNA from paraffin-embdedded squamous epithelium from individuals with no evidence of cancer.
3.4. Gene Probe Panels
The 41 gene probe panel (ME001B, www.mlpa.com) used in this cohort interrogates 38 unique genes implicated in cancer (24 tumor suppressor genes) for methylation status in two separate reactions (one in the absence of the methyl-sensitive enzyme HhaI and one in the presence of the HhaI enzyme). There are two probes each for MLH1, RASSF1 and BRCA2, and a normal control DNA sample will generate 41 individual peaks in the absence of HhaI and 15 individual peaks in the presence of HhaI.
3.5. Bisulfite Modification and Methylation-Specific Polymerase Chain Reaction (MSP) Assay
Genomic DNA (100 ng) from formalin-fixed paraffin embedded normal and cancerous thyroid tissue, control universal methylated DNA (Chamicon International, Inc) and control unmethylated DNA (normal genomic DNA) were modified using the EZ DNA methylation gold kit (Zymo Research, Orange, CA) during which methylated DNA is protected and unmethylated cytosine is converted to uracil [49]. The modified DNA served as a template using primers specific for the methylated or modified unmethylated sequences (Table 1). MSP amplification was performed using 3 μL of bisulfite modified DNA in a final volume of 25 μL using Promega Hot Start Master Mix (Promega Biosystems, Sunnyvale, CA), 0.5 μM primer followed by 95 °C 5 minutes then 40 cycles at 95 °C 40 seconds, 60 °C 40 seconds, 72 °C 1 min [49]. The resultant PCR products were separated on 2% agarose gel stained with ethidium bromide and visualized under UV illumination (Figure 3).
4. Conclusions
Our data suggests that aberrant methylation of CASP8, RASSF1, and NIS maybe an early change in thyroid tumorigenesis regardless of cell type. Epigenetic events of promoter hypermethylation are emerging as promising molecular targets for cancer detection and represent an important tumor-specific marker in tumorigenesis. Further confirmation and validation of these findings in larger cohorts would support these genes as early makers of thyroid tumorigenesis.




{kind=link}
{kind=link}
{kind=link}
| Methylation Specific Primers | Unmethylation Specific Primers | Product Size | |
|---|---|---|---|
| Forward | 5′-ATAGGGAGGTCGATACGGATATC | 5′-ATAGGGAGGTTGATATGGATATTGA | M-142 bp |
| Reverse | 5′-GAAAAAACAAAACGAAAAAAACG | 5′-AAAAAAACAAAACAAAAAAAACAAA | U-142 bp |
M = methylated product; U = unmethylated product
Acknowledgments
This work was supported by R01 NIH DE 15990 (MJW).
References
- Thyroid cancer. Available online: http://www.cancer.gov/cancertopics/types/thyroid (accessed on 24 February 2011).
- American Cancer Society. Quickfacts. In Thyroid Cancer; American Cancer Society: Atlanta, GA, USA, 2009. [Google Scholar]
- Stearns, V.; Yamauchi, H.; Hayes, D.F. Circulating tumor markers in breast cancer: Accepted utilities and novel prospects. Breast Cancer Res. Treat. 1998, 52, 239–259. [Google Scholar]
- Hu, M.I.; Vassilopoulou-Sellin, R.; Lustig, R.; Lamont, J.P. Thyroid and parathyroid cancers. In Cancer Management: A Multidisciplinary Approach, 11th ed.; Pazdur, R., Wagman, L.D., Camphausen, K.A., Hoskins, W.J., Eds.; Congregational Ministries Publishing: Louisville, KY, USA, 2008. [Google Scholar]
- American Cancer Society. American Cancer Society Cancer Facts and Figures; American Cancer Society: Atlanta, GA, USA, 2005. [Google Scholar]
- Weber, F.; Eng, C. Gene-expression profiling in differentiated thyroid cancer--a viable strategy for the practice of genomic medicine? Future Oncol. 2005, 1, 497–510. [Google Scholar]
- Mazzaferri, E.L.; Kloos, R.T. Clinical review 128: Current approaches to primary therapy for papillary and follicular thyroid cancer. J. Clin. Endocrinol. Metab. 2001, 86, 1447–1463. [Google Scholar]
- Schlumberger, M.J. Papillary and follicular thyroid carcinoma. N. Engl. J. Med. 1998, 338, 297–306. [Google Scholar]
- Hayes, D.F.; Bast, R.C.; Desch, C.E.; Fritsche, H., Jr.; Kemeny, N.E.; Jessup, J.M.; Locker, G.Y.; Macdonald, J.S.; Mennel, R.G.; Norton, L.; Ravdin, P.; Taube, S.; Winn, R.J. Tumor marker utility grading system: A framework to evaluate clinical utility of tumor markers. J. Natl. Cancer Inst. 1996, 88, 1456–1466. [Google Scholar]
- Kulasingam, V.; Diamandis, E.P. Strategies for discovering novel cancer biomarkers through utilization of emerging technologies. Nat. Clin. Pract. Oncol. 2008, 5, 588–599. [Google Scholar]
- Xing, M. Gene methylation in thyroid tumorigenesis. Endocrinology 2007, 148, 948–953. [Google Scholar]
- Smith, J.A.; Fan, C.Y.; Zou, C.; Bodenner, D.; Kokoska, M.S. Methylation status of genes in papillary thyroid carcinoma. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 1006–1011. [Google Scholar]
- Hu, S.; Liu, D.; Tufano, R.P.; Carson, K.A.; Rosenbaum, E.; Cohen, Y.; Holt, E.H.; Kiseljak-Vassiliades, K.; Rhoden, K.J.; Tolaney, S.; Condouris, S.; Tallini, G.; Westra, W.H.; Umbricht, C.B.; Zeiger, M.A.; Califano, J.A.; Vasko, V.; Xing, M. Association of aberrant methylation of tumor suppressor genes with tumor aggressiveness and braf mutation in papillary thyroid cancer. Int. J. Cancer 2006, 119, 2322–2329. [Google Scholar]
- Xing, M.; Cohen, Y.; Mambo, E.; Tallini, G.; Udelsman, R.; Ladenson, P.W.; Sidransky, D. Early occurrence of rassf1a hypermethylation and its mutual exclusion with braf mutation in thyroid tumorigenesis. Cancer Res. 2004, 64, 1664–1668. [Google Scholar]
- Lazar, V.; Bidart, J.M.; Caillou, B.; Mahe, C.; Lacroix, L.; Filetti, S.; Schlumberger, M. Expression of the na+/i- symporter gene in human thyroid tumors: A comparison study with other thyroid-specific genes. J. Clin. Endocrinol. Metab. 1999, 84, 3228–3234. [Google Scholar]
- Xing, M.; Usadel, H.; Cohen, Y.; Tokumaru, Y.; Guo, Z.; Westra, W.B.; Tong, B.C.; Tallini, G.; Udelsman, R.; Califano, J.A.; Ladenson, P.W.; Sidransky, D. Methylation of the thyroid-stimulating hormone receptor gene in epithelial thyroid tumors: A marker of malignancy and a cause of gene silencing. Cancer Res. 2003, 63, 2316–2321. [Google Scholar]
- Ringel, M.D.; Anderson, J.; Souza, S.L.; Burch, H.B.; Tambascia, M.; Shriver, C.D.; Tuttle, R.M. Expression of the sodium iodide symporter and thyroglobulin genes are reduced in papillary thyroid cancer. Mod. Pathol. 2001, 14, 289–296. [Google Scholar]
- Segev, D.L.; Clark, D.P.; Zeiger, M.A.; Umbricht, C. Beyond the suspicious thyroid fine needle aspirate. A review. Acta. Cytol. 2003, 47, 709–722. [Google Scholar]
- Sherman, S.I. Thyroid carcinoma. Lancet 2003, 361, 501–511. [Google Scholar]
- Baloch, Z.W.; Fleisher, S.; LiVolsi, V.A.; Gupta, P.K. Diagnosis of “Follicular neoplasm”: A gray zone in thyroid fine-needle aspiration cytology. Diagn. Cytopathol. 2002, 26, 41–44. [Google Scholar]
- Yeh, M.W.; Demircan, O.; Ituarte, P.; Clark, O.H. False-negative fine-needle aspiration cytology results delay treatment and adversely affect outcome in patients with thyroid carcinoma. Thyroid 2004, 14, 207–215. [Google Scholar]
- Putti, T.C.; Bhuiya, T.A.; Wasserman, P.G. Fine needle aspiration cytology of mixed tall and columnar cell papillary carcinoma of the thyroid. A case report. Acta. Cytol. 1998, 42, 387–390. [Google Scholar]
- Wreesmann, V.B.; Sieczka, E.M.; Socci, N.D.; Hezel, M.; Belbin, T.J.; Childs, G.; Patel, S.G.; Patel, K.N.; Tallini, G.; Prystowsky, M.; Shaha, A.R.; Kraus, D.; Shah, J.P.; Rao, P.H.; Ghossein, R.; Singh, B. Genome-wide profiling of papillary thyroid cancer identifies muc1 as an independent prognostic marker. Cancer Res. 2004, 64, 3780–3789. [Google Scholar]
- Ruggeri, R.M.; Campenni, A.; Baldari, S.; Trimarchi, F.; Trovato, M. What is new on thyroid cancer biomarkers. Biomark. Insights 2008, 3, 237–252. [Google Scholar]
- Pingoud-Meier, C.; Lang, D.; Janss, A.J.; Rorke, L.B.; Phillips, P.C.; Shalaby, T.; Grotzer, M.A. Loss of caspase-8 protein expression correlates with unfavorable survival outcome in childhood medulloblastoma. Clin. Cancer. Res. 2003, 9, 6401–6409. [Google Scholar]
- Yang, Q.; Kiernan, C.M.; Tian, Y.; Salwen, H.R.; Chlenski, A.; Brumback, B.A.; London, W.B.; Cohn, S.L. Methylation of casp8, dcr2, and hin-1 in neuroblastoma is associated with poor outcome. Clin. Cancer. Res. 2007, 13, 3191–3197. [Google Scholar]
- Harada, K.; Toyooka, S.; Shivapurkar, N.; Maitra, A.; Reddy, J.L.; Matta, H.; Miyajima, K.; Timmons, C.F.; Tomlinson, G.E.; Mastrangelo, D.; Hay, R.J.; Chaudhary, P.M.; Gazdar, A.F. Deregulation of caspase 8 and 10 expression in pediatric tumors and cell lines. Cancer Res. 2002, 62, 5897–5901. [Google Scholar]
- Wu, Y.; Alvarez, M.; Slamon, D.J.; Koeffler, P.; Vadgama, J.V. Caspase 8 and maspin are downregulated in breast cancer cells due to cpg site promoter methylation. BMC Cancer 2010, 10, 32. [Google Scholar]
- Ho, P.K.; Hawkins, C.J. Mammalian initiator apoptotic caspases. Febs. J. 2005, 272, 5436–5453. [Google Scholar]
- Teitz, T.; Wei, T.; Valentine, M.B.; Vanin, E.F.; Grenet, J.; Valentine, V.A.; Behm, F.G.; Look, A.T.; Lahti, J.M.; Kidd, V.J. Caspase 8 is deleted or silenced preferentially in childhood neuroblastomas with amplification of mycn. Nat. Med. 2000, 6, 529–535. [Google Scholar]
- Fulda, S.; Debatin, K.M. Ifngamma sensitizes for apoptosis by upregulating caspase-8 expression through the stat1 pathway. Oncogene 2002, 21, 2295–2308. [Google Scholar]
- Liedtke, C.; Groger, N.; Manns, M.P.; Trautwein, C. The human caspase-8 promoter sustains basal activity through sp1 and ets-like transcription factors and can be up-regulated by a p53-dependent mechanism. J. Biol. Chem. 2003, 278, 27593–27604. [Google Scholar]
- Chaudhary, P.M.; Eby, M.T.; Jasmin, A.; Kumar, A.; Liu, L.; Hood, L. Activation of the nf-kappab pathway by caspase 8 and its homologs. Oncogene 2000, 19, 4451–4460. [Google Scholar]
- Salvesen, G.S. Caspase 8: Igniting the death machine. Structure 1999, 7, R225–R229. [Google Scholar]
- Zoli, W.; Ulivi, P.; Tesei, A.; Fabbri, F.; Rosetti, M.; Maltoni, R.; Giunchi, D.C.; Ricotti, L.; Brigliadori, G.; Vannini, I.; Amadori, D. Addition of 5-fluorouracil to doxorubicin-paclitaxel sequence increases caspase-dependent apoptosis in breast cancer cell lines. Breast Cancer Res. 2005, 7, R681–R689. [Google Scholar]
- Ehrhardt, H.; Hacker, S.; Wittmann, S.; Maurer, M.; Borkhardt, A.; Toloczko, A.; Debatin, K.M.; Fulda, S.; Jeremias, I. Cytotoxic drug-induced, p53-mediated upregulation of caspase-8 in tumor cells. Oncogene 2008, 27, 783–793. [Google Scholar]
- Schagdarsurengin, U.; Gimm, O.; Hoang-Vu, C.; Dralle, H.; Pfeifer, G.P.; Dammann, R. Frequent epigenetic silencing of the cpg island promoter of rassf1a in thyroid carcinoma. Cancer Res. 2002, 62, 3698–3701. [Google Scholar]
- Hou, P.; Bojdani, E.; Xing, M. Induction of thyroid gene expression and radioiodine uptake in thyroid cancer cells by targeting major signaling pathways. J. Clin. Endocrinol. Metab. 2010, 95, 820–828. [Google Scholar]
- Venkataraman, G.M.; Yatin, M.; Marcinek, R.; Ain, K.B. Restoration of iodide uptake in dedifferentiated thyroid carcinoma: Relationship to human na+/i-symporter gene methylation status. J. Clin. Endocrinol. Metab. 1999, 84, 2449–2457. [Google Scholar]
- Boltze, C.; Zack, S.; Quednow, C.; Bettge, S.; Roessner, A.; Schneider-Stock, R. Hypermethylation of the cdkn2/p16ink4a promotor in thyroid carcinogenesis. Pathol. Res. Pract. 2003, 199, 399–404. [Google Scholar]
- Smanik, P.A.; Ryu, K.Y.; Theil, K.S.; Mazzaferri, E.L.; Jhiang, S.M. Expression, exon-intron organization, and chromosome mapping of the human sodium iodide symporter. Endocrinology 1997, 138, 3555–3558. [Google Scholar]
- Arturi, F.; Russo, D.; Schlumberger, M.; du Villard, J.A.; Caillou, B.; Vigneri, P.; Wicker, R.; Chiefari, E.; Suarez, H.G.; Filetti, S. Iodide symporter gene expression in human thyroid tumors. J. Clin. Endocrinol. Metab. 1998, 83, 2493–2496. [Google Scholar]
- Filetti, S.; Bidart, J.M.; Arturi, F.; Caillou, B.; Russo, D.; Schlumberger, M. Sodium/iodide symporter: A key transport system in thyroid cancer cell metabolism. Eur. J. Endocrinol. 1999, 141, 443–457. [Google Scholar]
- Stephen, J.K.; Vaught, L.E.; Chen, K.M.; Sethi, S.; Shah, V.; Benninger, M.S.; Gardner, G.M.; Schweitzer, V.G.; Khan, M.; Worsham, M.J. Epigenetic events underlie the pathogenesis of sinonasal papillomas. Mod. Pathol. 2007, 20, 1019–1027. [Google Scholar]
- Stephen, J.K.; Chen, K.M.; Shah, V.; Schweitzer, V.G.; Gardner, G.; Benninger, M.S.; Worsham, M.J. Consistent DNA hypermethylation patterns in laryngeal papillomas. Int. J. Head Neck Surg. 2010, 1, 69–77. [Google Scholar]
- Nygren, A.O.; Ameziane, N.; Duarte, H.M.; Vijzelaar, R.N.; Waisfisz, Q.; Hess, C.J.; Schouten, J.P.; Errami, A. Methylation-specific mlpa (ms-mlpa): Simultaneous detection of cpg methylation and copy number changes of up to 40 sequences. Nucleic Acids Res. 2005, 33, e128. [Google Scholar]
- Stephen, J.K.; Vaught, L.E.; Chen, K.M.; Shah, V.; Schweitzer, V.G.; Gardner, G.; Benninger, M.S.; Worsham, M.J. An epigenetically derived monoclonal origin for recurrent respiratory papillomatosis. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 684–692. [Google Scholar]
- Worsham, M.J.; Chen, K.M.; Meduri, V.; Nygren, A.O.; Errami, A.; Schouten, J.P.; Benninger, M.S. Epigenetic events of disease progression in head and neck squamous cell carcinoma. Arch. Otolaryngol. Head Neck Surg. 2006, 132, 668–677. [Google Scholar]
- Chen, K.; Sawhney, R.; Khan, M.; Benninger, M.S.; Hou, Z.; Sethi, S.; Stephen, J.K.; Worsham, M.J. Methylation of multiple genes as diagnostic and therapeutic markers in primary head and neck squamous cell carcinoma. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 1131–1138. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stephen, J.K.; Chitale, D.; Narra, V.; Chen, K.M.; Sawhney, R.; Worsham, M.J. DNA Methylation in Thyroid Tumorigenesis. Cancers 2011, 3, 1732-1743. https://doi.org/10.3390/cancers3021732
Stephen JK, Chitale D, Narra V, Chen KM, Sawhney R, Worsham MJ. DNA Methylation in Thyroid Tumorigenesis. Cancers. 2011; 3(2):1732-1743. https://doi.org/10.3390/cancers3021732
Chicago/Turabian StyleStephen, Josena K., Dhananjay Chitale, Vinod Narra, Kang Mei Chen, Raja Sawhney, and Maria J. Worsham. 2011. "DNA Methylation in Thyroid Tumorigenesis" Cancers 3, no. 2: 1732-1743. https://doi.org/10.3390/cancers3021732
APA StyleStephen, J. K., Chitale, D., Narra, V., Chen, K. M., Sawhney, R., & Worsham, M. J. (2011). DNA Methylation in Thyroid Tumorigenesis. Cancers, 3(2), 1732-1743. https://doi.org/10.3390/cancers3021732