STAT3: A Novel Molecular Mediator of Resistance to Chemoradiotherapy

Abstract
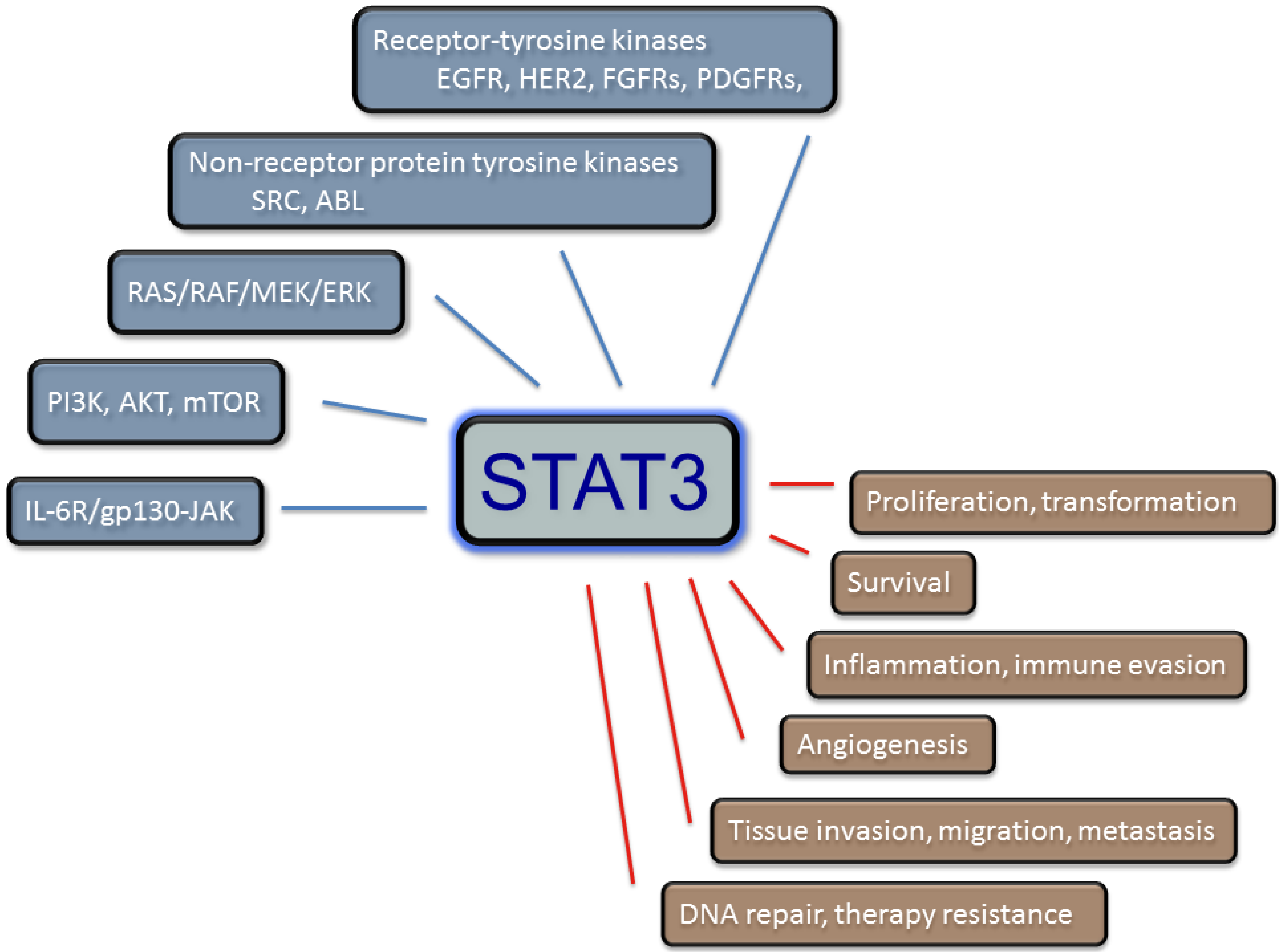
:1. Introduction
1.1. The Clinical Problem of Resistance to Chemoradiotherapy
1.2. The JAK/STAT Signaling Paradigm in Cancer

2. STAT3 as a Molecular Target to Sensitize Tumors to (Chemo-) Radiotherapy
2.1. Brain Tumors
2.2. Breast Cancer
2.3. Colorectal Cancer
2.4. Esophageal Cancer
2.5. Prostate Cancer
2.6. Bladder Cancer
2.7. Cervical Cancer
2.8. Head and Neck Cancer
2.9. Lung Cancer
2.10. Skin Cancer
2.11. Other Cancers
3. Potential Mechanisms of STAT3-Mediated Radio- and Chemoradiotherapy-Resistance
4. STAT3 Inhibitors in Clinical Translation
5. Perspective and Conclusions

{kind=link}
{kind=link}
| Tumor Type | Treatment | Pathway Modulation | Sensitization In Vitro | Sensitization In Vivo | Reference |
|---|---|---|---|---|---|
| Anaplastic thyroid cancer | RT/CRT (cisplatin) | Cucurbitacin I (JSI-124) | yes | yes | [116] |
| Breast cancer | RT | DN-STAT3 | yes | n.a. | [56] |
| RT | Xanthohumol | yes | n.a. | [57] | |
| Cervical cancer | RT | Cepharanthine | yes | yes | [80] |
| Colorectal cancer | CRT (5-FU) | STAT3 shRNA STAT3 siRNA STATTIC | yes yes yes | n.a. n.a. yes | [62] |
| CRT (5-FU) | Selumetinib (AZD6244) | yes | yes | [63] | |
| Esophageal cancer | RT | IL-6 shRNA | yes | yes | [66] |
| Glioblastoma | RT | DN-STAT3 | yes | n.a. | [44] |
| RT | STAT3 siRNA | yes | yes | [45] | |
| RT | Resveratrol STAT3 shRNA AG490 | yes yes yes | yes | [46] | |
| RT | Cucurbitacin I (JSI-124) gp130-blocking antibody | no no | n.a. | [47] | |
| Head and neck cancer | RT | Panitumumab Panitumumab | yes no (SCC-1483) | yes n.a. | [92] |
| RT | IL-6 antibody | yes | n.a. | [90] | |
| RT | STAT3 siRNA | yes | n.a. | [93] | |
| RT | STAT3 shRNA | n.a. | yes | [32] | |
| RT | STAT3 siRNA | yes | n.a. | [94] | |
| RT | STATTIC STATTIC | yes no (UM-SCC-22B) | yes n.a. | [95] | |
| RT | STATTIC | yes | n.a. | [96] | |
| RT | Cucurbitacin I (JSI-124) | yes | yes | [91] | |
| RT | Linifanib (ABT-869) | yes | n.a. | [98] | |
| Hepatocellular carcinoma | RT | STAT3 siRNA Sorafenib | yes yes | n.a. yes | [117] |
| Leukemia | RT | Stat3−/− mouse B-1 cells | yes | n.a. | [25] |
| Lung cancer | RT | Panitumumab | yes | yes | [92] |
| RT | Saracatinib (AZD0530) PP2 | yes no | n.a. | [100] | |
| RT | TG101209 | yes | yes | [101] | |
| RT | TBB TBCA Hematein | yes yes yes | n.a. | [102] | |
| RT | STAT3 shRNA | yes | yes | [103] | |
| RT | Cucurbitacin I (JSI-124) | yes | yes | [104] | |
| Melanoma | RT | Resveratrol | yes | n.a. | [111] |
| RT | STAT3 siRNA STAT3 shRNA | yes yes | n.a. | [112] | |
| Prostate cancer | RT | IL-6 shRNA | yes | yes | [31] |
| Skin cancer | UVB-RT | Stat3−/− mouse keratinocytes | yes | yes | [109] |

Acknowledgments
Conflicts of Interest
References
- Rodel, C.; Liersch, T.; Becker, H.; Fietkau, R.; Hohenberger, W.; Hothorn, T.; Graeven, U.; Arnold, D.; Lang-Welzenbach, M.; Raab, H.R.; et al. Preoperative chemoradiotherapy and postoperative chemotherapy with fluorouracil and oxaliplatin versus fluorouracil alone in locally advanced rectal cancer: Initial results of the German CAO/ARO/AIO-04 randomised phase 3 trial. Lancet Oncol. 2012, 13, 679–687. [Google Scholar] [PubMed]
- Jagsi, R. Progress and controversies: Radiation therapy for invasive breast cancer. CA Cancer J. Clin. 2013, 64, 135–152. [Google Scholar] [PubMed]
- Reck, M.; Heigener, D.F.; Mok, T.; Soria, J.C.; Rabe, K.F. Management of non-small-cell lung cancer: Recent developments. Lancet 2013, 382, 709–719. [Google Scholar] [PubMed]
- Thariat, J.; Hannoun-Levi, J.M.; Sun Myint, A.; Vuong, T.; Gerard, J.P. Past, present, and future of radiotherapy for the benefit of patients. Nat. Rev. Clin. Oncol. 2013, 10, 52–60. [Google Scholar] [PubMed]
- Seiwert, T.Y.; Salama, J.K.; Vokes, E.E. The concurrent chemoradiation paradigm—General principles. Nat. Clin. Pract. Oncol. 2007, 4, 86–100. [Google Scholar] [PubMed]
- Katz, D.; Ito, E.; Liu, F.F. On the path to seeking novel radiosensitizers. Int. J. Radiat. Oncol. Biol. Phys. 2009, 73, 988–996. [Google Scholar] [PubMed]
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to improve radiotherapy with targeted drugs. Nat. Rev. Cancer 2011, 11, 239–253. [Google Scholar] [PubMed]
- Shuai, K.; Stark, G.R.; Kerr, I.M.; Darnell, J.E., Jr. A single phosphotyrosine residue of Stat91 required for gene activation by interferon-gamma. Science 1993, 261, 1744–1746. [Google Scholar] [PubMed]
- Shuai, K.; Ziemiecki, A.; Wilks, A.F.; Harpur, A.G.; Sadowski, H.B.; Gilman, M.Z.; Darnell, J.E. Polypeptide signalling to the nucleus through tyrosine phosphorylation of Jak and Stat proteins. Nature 1993, 366, 580–583. [Google Scholar] [PubMed]
- O'Shea, J.J.; Holland, S.M.; Staudt, L.M. JAKs and STATs in immunity, immunodeficiency, and cancer. N. Engl. J. Med. 2013, 368, 161–170. [Google Scholar] [PubMed]
- Quintas-Cardama, A.; Verstovsek, S. Molecular pathways: Jak/STAT pathway: Mutations, inhibitors, and resistance. Clin. Cancer Res. 2013, 19, 1933–1940. [Google Scholar] [PubMed]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [PubMed]
- Aaronson, D.S.; Horvath, C.M. A road map for those who don’t know JAK-STAT. Science 2002, 296, 1653–1655. [Google Scholar] [PubMed]
- Darnell, J.E., Jr. STATs and gene regulation. Science 1997, 277, 1630–1635. [Google Scholar] [PubMed]
- Leonard, W.J. Role of Jak kinases and STATs in cytokine signal transduction. Int. J. Hematol. 2001, 73, 271–277. [Google Scholar] [PubMed]
- Hebenstreit, D.; Horejs-Hoeck, J.; Duschl, A. JAK/STAT-dependent gene regulation by cytokines. Drug News Perspect. 2005, 18, 243–249. [Google Scholar] [PubMed]
- Kerr, I.M.; Costa-Pereira, A.P.; Lillemeier, B.F.; Strobl, B. Of JAKs, STATs, blind watchmakers, jeeps and trains. FEBS Lett. 2003, 546, 1–5. [Google Scholar] [PubMed]
- Greenhalgh, C.J.; Hilton, D.J. Negative regulation of cytokine signaling. J. Leukoc. Biol. 2001, 70, 348–356. [Google Scholar] [PubMed]
- Krebs, D.L.; Hilton, D.J. SOCS proteins: Negative regulators of cytokine signaling. Stem Cells 2001, 19, 378–387. [Google Scholar] [PubMed]
- Kim, D.J.; Tremblay, M.L.; Digiovanni, J. Protein tyrosine phosphatases, TC-PTP, SHP1, and SHP2, cooperate in rapid dephosphorylation of Stat3 in keratinocytes following UVB irradiation. PLoS One 2010, 5, e10290. [Google Scholar] [PubMed]
- Shuai, K. Regulation of cytokine signaling pathways by PIAS proteins. Cell Res. 2006, 16, 196–202. [Google Scholar] [PubMed]
- Faisst, S.; Meyer, S. Compilation of vertebrate-encoded transcription factors. Nucleic Acids Res. 1992, 20, 3–26. [Google Scholar] [PubMed]
- Levy, D.E.; Lee, C.K. What does Stat3 do? J. Clin. Invest. 2002, 109, 1143–1148. [Google Scholar] [CrossRef]
- Bromberg, J.F.; Wrzeszczynska, M.H.; Devgan, G.; Zhao, Y.; Pestell, R.G.; Albanese, C.; Darnell, J.E., Jr. Stat3 as an oncogene. Cell 1999, 98, 295–303. [Google Scholar] [PubMed]
- Otero, D.C.; Poli, V.; David, M.; Rickert, R.C. Cutting edge: Inherent and acquired resistance to radiation-induced apoptosis in B cells: A pivotal role for STAT3. J. Immunol. 2006, 177, 6593–6597. [Google Scholar] [PubMed]
- Hutchins, A.P.; Diez, D.; Miranda-Saavedra, D. Genomic and computational approaches to dissect the mechanisms of STAT3’s universal and cell type-specific functions. JAKSTAT 2013, 2, e25097. [Google Scholar] [PubMed]
- Chang, Q.; Daly, L.; Bromberg, J. The IL-6 feed-forward loop: A driver of tumorigenesis. Semin. Immunol. 2014, 26, 48–53. [Google Scholar] [PubMed]
- De la Iglesia, N.; Konopka, G.; Puram, S.V.; Chan, J.A.; Bachoo, R.M.; You, M.J.; Levy, D.E.; Depinho, R.A.; Bonni, A. Identification of a PTEN-regulated STAT3 brain tumor suppressor pathway. Genes Dev. 2008, 22, 449–462. [Google Scholar]
- Bromberg, J. Stat proteins and oncogenesis. J. Clin. Invest. 2002, 109, 1139–1142. [Google Scholar] [PubMed]
- Chumsri, S.; Shah, P. Radiation resistance of cancer stem cells as an obstacle in cancer therapy. Mol. Cell. Pharmacol. 2013, 5, 39–49. [Google Scholar]
- Wu, C.T.; Chen, M.F.; Chen, W.C.; Hsieh, C.C. The role of IL-6 in the radiation response of prostate cancer. Radiat. Oncol. 2013, 8, 159. [Google Scholar] [PubMed]
- Li, X.; Wang, H.; Lu, X.; Di, B. Silencing STAT3 with short hairpin RNA enhances radiosensitivity of human laryngeal squamous cell carcinoma xenografts in vivo. Exp. Ther. Med. 2010, 1, 947–953. [Google Scholar] [PubMed]
- Sun, L.; Cabarcas, S.M.; Farrar, W.L. Radioresistance and cancer stem cells: Survival of the fittest. J. Carcinog. Mutagen. 2011, S1, 1–12. [Google Scholar]
- You, S.; Li, R.; Park, D.; Xie, M.; Sica, G.L.; Cao, Y.; Xiao, Z.Q.; Deng, X. Disruption of STAT3 by niclosamide reverses radioresistance of human lung cancer. Mol. Cancer Ther. 2014, 13, 606–616. [Google Scholar] [PubMed]
- Panni, R.Z.; Sanford, D.E.; Belt, B.A.; Mitchem, J.B.; Worley, L.A.; Goetz, B.D.; Mukherjee, P.; Wang-Gillam, A.; Link, D.C.; Denardo, D.G.; et al. Tumor-induced STAT3 activation in monocytic myeloid-derived suppressor cells enhances stemness and mesenchymal properties in human pancreatic cancer. Cancer Immunol. Immunother. 2014, 63, 513–528. [Google Scholar] [PubMed]
- Turkson, J.; Jove, R. STAT proteins: Novel molecular targets for cancer drug discovery. Oncogene 2000, 19, 6613–6626. [Google Scholar] [PubMed]
- Wang, X.; Crowe, P.J.; Goldstein, D.; Yang, J.L. STAT3 inhibition, a novel approach to enhancing targeted therapy in human cancers (review). Int. J. Oncol. 2012, 41, 1181–1191. [Google Scholar] [PubMed]
- Wen, P.Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [PubMed]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [PubMed]
- Mrugala, M.M. Advances and challenges in the treatment of glioblastoma: A clinician’s perspective. Discov. Med. 2013, 15, 221–230. [Google Scholar] [PubMed]
- Tchirkov, A.; Rolhion, C.; Bertrand, S.; Dore, J.F.; Dubost, J.J.; Verrelle, P. IL-6 gene amplification and expression in human glioblastomas. Br. J. Cancer 2001, 85, 518–522. [Google Scholar] [PubMed]
- Birner, P.; Toumangelova-Uzeir, K.; Natchev, S.; Guentchev, M. STAT3 tyrosine phosphorylation influences survival in glioblastoma. J. Neurooncol. 2010, 100, 339–343. [Google Scholar] [PubMed]
- Brantley, E.C.; Benveniste, E.N. Signal transducer and activator of transcription-3: A molecular hub for signaling pathways in gliomas. Mol. Cancer Res. 2008, 6, 675–684. [Google Scholar] [PubMed]
- Zhou, H.; Miki, R.; Eeva, M.; Fike, F.M.; Seligson, D.; Yang, L.; Yoshimura, A.; Teitell, M.A.; Jamieson, C.A.; Cacalano, N.A. Reciprocal regulation of SOCS 1 and SOCS3 enhances resistance to ionizing radiation in glioblastoma multiforme. Clin. Cancer Res. 2007, 13, 2344–2353. [Google Scholar] [PubMed]
- Gao, L.; Li, F.; Dong, B.; Zhang, J.; Rao, Y.; Cong, Y.; Mao, B.; Chen, X. Inhibition of STAT3 and ErbB2 suppresses tumor growth, enhances radiosensitivity, and induces mitochondria-dependent apoptosis in glioma cells. Int. J. Radiat. Oncol. Biol. Phys. 2010, 77, 1223–1231. [Google Scholar] [PubMed]
- Yang, Y.P.; Chang, Y.L.; Huang, P.I.; Chiou, G.Y.; Tseng, L.M.; Chiou, S.H.; Chen, M.H.; Chen, M.T.; Shih, Y.H.; Chang, C.H.; et al. Resveratrol suppresses tumorigenicity and enhances radiosensitivity in primary glioblastoma tumor initiating cells by inhibiting the STAT3 axis. J. Cell Physiol. 2012, 227, 976–993. [Google Scholar] [PubMed]
- Chautard, E.; Loubeau, G.; Tchirkov, A.; Chassagne, J.; Vermot-Desroches, C.; Morel, L.; Verrelle, P. Akt signaling pathway: A target for radiosensitizing human malignant glioma. Neuro-oncology 2010, 12, 434–443. [Google Scholar] [PubMed]
- Patel, J.D.; Krilov, L.; Adams, S.; Aghajanian, C.; Basch, E.; Brose, M.S.; Carroll, W.L.; de Lima, M.; Gilbert, M.R.; Kris, M.G.; et al. Clinical cancer advances 2013: Annual report on progress against cancer from the American society of clinical oncology. J. Clin. Oncol. 2014, 32, 129–160. [Google Scholar] [PubMed]
- Buchholz, T.A. Radiation therapy for early-stage breast cancer after breast-conserving surgery. N. Engl. J. Med. 2009, 360, 63–70. [Google Scholar] [PubMed]
- Hoover, S.; Bloom, E.; Patel, S. Review of breast conservation therapy: Then and now. ISRN Oncol. 2011, 2011, 617593. [Google Scholar] [PubMed]
- Watson, C.J.; Miller, W.R. Elevated levels of members of the STAT family of transcription factors in breast carcinoma nuclear extracts. Br. J. Cancer 1995, 71, 840–844. [Google Scholar] [PubMed]
- Garcia, R.; Yu, C.L.; Hudnall, A.; Catlett, R.; Nelson, K.L.; Smithgall, T.; Fujita, D.J.; Ethier, S.P.; Jove, R. Constitutive activation of Stat3 in fibroblasts transformed by diverse oncoproteins and in breast carcinoma cells. Cell Growth Differ. 1997, 8, 1267–1276. [Google Scholar] [PubMed]
- Bowman, T.; Garcia, R.; Turkson, J.; Jove, R. STATs in oncogenesis. Oncogene 2000, 19, 2474–2488. [Google Scholar] [PubMed]
- Page, C.; Huang, M.; Jin, X.; Cho, K.; Lilja, J.; Reynolds, R.K.; Lin, J. Elevated phosphorylation of AKT and Stat3 in prostate, breast, and cervical cancer cells. Int. J. Oncol. 2000, 17, 23–28. [Google Scholar] [PubMed]
- Burke, W.M.; Jin, X.; Lin, H.J.; Huang, M.; Liu, R.; Reynolds, R.K.; Lin, J. Inhibition of constitutively active Stat3 suppresses growth of human ovarian and breast cancer cells. Oncogene 2001, 20, 7925–7934. [Google Scholar] [PubMed]
- Kim, K.W.; Mutter, R.W.; Cao, C.; Albert, J.M.; Shinohara, E.T.; Sekhar, K.R.; Lu, B. Inhibition of signal transducer and activator of transcription 3 activity results in down-regulation of survivin following irradiation. Mol. Cancer Ther. 2006, 5, 2659–2665. [Google Scholar] [PubMed]
- Kang, Y.; Park, M.A.; Heo, S.W.; Park, S.Y.; Kang, K.W.; Park, P.H.; Kim, J.A. The radio-sensitizing effect of xanthohumol is mediated by STAT3 and EGFR suppression in doxorubicin-resistant MCF-7 human breast cancer cells. Biochim. Biophys. Acta 2013, 1830, 2638–2648. [Google Scholar] [PubMed]
- Cunningham, D.; Atkin, W.; Lenz, H.J.; Lynch, H.T.; Minsky, B.; Nordlinger, B.; Starling, N. Colorectal cancer. Lancet 2010, 375, 1030–1047. [Google Scholar] [PubMed]
- Sauer, R.; Becker, H.; Hohenberger, W.; Rodel, C.; Wittekind, C.; Fietkau, R.; Martus, P.; Tschmelitsch, J.; Hager, E.; Hess, C.F.; et al. Preoperative versus postoperative chemoradiotherapy for rectal cancer. N. Engl. J. Med. 2004, 351, 1731–1740. [Google Scholar] [PubMed]
- Rodel, C.; Hofheinz, R.; Liersch, T. Rectal cancer: State of the art in 2012. Curr. Opin. Oncol. 2012, 24, 441–447. [Google Scholar] [PubMed]
- Spitzner, M.; Emons, G.; Kramer, F.; Gaedcke, J.; Rave-Frank, M.; Scharf, J.G.; Burfeind, P.; Becker, H.; Beissbarth, T.; Ghadimi, B.M.; et al. A gene expression signature for chemoradiosensitivity of colorectal cancer cells. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 1184–1192. [Google Scholar] [PubMed]
- Spitzner, M.; Roesler, B.; Bielfeld, C.; Emons, G.; Gaedcke, J.; Wolff, H.A.; Rave-Frank, M.; Kramer, F.; Beissbarth, T.; Kitz, J.; et al. STAT3 inhibition sensitizes colorectal cancer to chemoradiotherapy in vitro and in vivo. Int. J. Cancer 2014, 134, 997–1007. [Google Scholar]
- Urick, M.E.; Chung, E.J.; Shield, W.P., 3rd; Gerber, N.; White, A.; Sowers, A.; Thetford, A.; Camphausen, K.; Mitchell, J.; Citrin, D.E. Enhancement of 5-fluorouracil-induced in vitro and in vivo radiosensitization with MEK inhibition. Clin. Cancer Res. 2011, 17, 5038–5047. [Google Scholar] [PubMed]
- Enzinger, P.C.; Mayer, R.J. Esophageal cancer. N. Engl. J. Med. 2003, 349, 2241–2252. [Google Scholar] [PubMed]
- Pennathur, A.; Gibson, M.K.; Jobe, B.A.; Luketich, J.D. Oesophageal carcinoma. Lancet 2013, 381, 400–412. [Google Scholar] [PubMed]
- Chen, M.F.; Lu, M.S.; Lin, P.Y.; Chen, P.T.; Chen, W.C.; Lee, K.D. The role of DNA methyltransferase 3b in esophageal squamous cell carcinoma. Cancer 2012, 118, 4074–4089. [Google Scholar] [PubMed]
- Chen, M.F.; Chen, P.T.; Lu, M.S.; Lin, P.Y.; Chen, W.C.; Lee, K.D. IL-6 expression predicts treatment response and outcome in squamous cell carcinoma of the esophagus. Mol. Cancer 2013, 12, 26. [Google Scholar] [PubMed]
- Cooperberg, M.R.; Broering, J.M.; Carroll, P.R. Time trends and local variation in primary treatment of localized prostate cancer. J. Clin. Oncol. 2010, 28, 1117–1123. [Google Scholar] [PubMed]
- Carter, H.B. Management of low (favourable)-risk prostate cancer. BJU Int. 2011, 108, 1684–1695. [Google Scholar] [PubMed]
- D’Amico, A.V.; Moul, J.; Carroll, P.R.; Sun, L.; Lubeck, D.; Chen, M.H. Cancer-specific mortality after surgery or radiation for patients with clinically localized prostate cancer managed during the prostate-specific antigen era. J. Clin. Oncol. 2003, 21, 2163–2172. [Google Scholar] [PubMed]
- Mora, L. B.; Buettner, R.; Seigne, J.; Diaz, J.; Ahmad, N.; Garcia, R.; Bowman, T.; Falcone, R.; Fairclough, R.; Cantor, A.; et al. Constitutive activation of Stat3 in human prostate tumors and cell lines: direct inhibition of Stat3 signaling induces apoptosis of prostate cancer cells. Cancer Res. 2002, 62, 6659–6666. [Google Scholar] [PubMed]
- Smith, Z.L.; Christodouleas, J.P.; Keefe, S.M.; Malkowicz, S.B.; Guzzo, T.J. Bladder preservation in the treatment of muscle-invasive bladder cancer (MIBC): A review of the literature and a practical approach to therapy. BJU Int. 2013, 112, 13–25. [Google Scholar] [PubMed]
- Meeks, J.J.; Bellmunt, J.; Bochner, B.H.; Clarke, N.W.; Daneshmand, S.; Galsky, M.D.; Hahn, N.M.; Lerner, S.P.; Mason, M.; Powles, T.; et al. A systematic review of neoadjuvant and adjuvant chemotherapy for muscle-invasive bladder cancer. Eur. Urol. 2012, 62, 523–533. [Google Scholar] [PubMed]
- Sun, Y.; Cheng, M.K.; Griffiths, T.R.; Mellon, J.K.; Kai, B.; Kriajevska, M.; Manson, M.M. Inhibition of STAT signalling in bladder cancer by diindolylmethane: Relevance to cell adhesion, migration and proliferation. Curr. Cancer Drug Targets 2013, 13, 57–68. [Google Scholar] [PubMed]
- Chen, C.L.; Cen, L.; Kohout, J.; Hutzen, B.; Chan, C.; Hsieh, F.C.; Loy, A.; Huang, V.; Cheng, G.; Lin, J. Signal transducer and activator of transcription 3 activation is associated with bladder cancer cell growth and survival. Mol. Cancer 2008, 7, 78. [Google Scholar] [PubMed]
- Barbera, L.; Thomas, G. Management of early and locally advanced cervical cancer. Semin. Oncol. 2009, 36, 155–169. [Google Scholar] [PubMed]
- Duenas-Gonzalez, A.; Zarba, J.J.; Patel, F.; Alcedo, J.C.; Beslija, S.; Casanova, L.; Pattaranutaporn, P.; Hameed, S.; Blair, J.M.; Barraclough, H.; et al. Phase III, open-label, randomized study comparing concurrent gemcitabine plus cisplatin and radiation followed by adjuvant gemcitabine and cisplatin versus concurrent cisplatin and radiation in patients with stage IIB to IVA carcinoma of the cervix. J. Clin. Oncol. 2011, 29, 1678–1685. [Google Scholar] [PubMed]
- Shukla, S.; Shishodia, G.; Mahata, S.; Hedau, S.; Pandey, A.; Bhambhani, S.; Batra, S.; Basir, S.F.; Das, B.C.; Bharti, A.C. Aberrant expression and constitutive activation of STAT3 in cervical carcinogenesis: Implications in high-risk human papillomavirus infection. Mol. Cancer 2010, 9, 282. [Google Scholar] [PubMed]
- Chen, H.H.; Chou, C.Y.; Wu, Y.H.; Hsueh, W.T.; Hsu, C.H.; Guo, H.R.; Lee, W.Y.; Su, W.C. Constitutive STAT5 activation correlates with better survival in cervical cancer patients treated with radiation therapy. Int. J. Radiat. Oncol. Biol. Phys. 2012, 82, 658–666. [Google Scholar] [PubMed]
- Fang, Z.H.; Li, Y.J.; Chen, Z.; Wang, J.J.; Zhu, L.H. Inhibition of signal transducer and activator of transcription 3 and cyclooxygenase-2 is involved in radiosensitization of cepharanthine in HeLa cells. Int. J. Gynecol. Cancer 2013, 23, 608–614. [Google Scholar] [PubMed]
- Bernier, J.; Domenge, C.; Ozsahin, M.; Matuszewska, K.; Lefebvre, J.L.; Greiner, R.H.; Giralt, J.; Maingon, P.; Rolland, F.; Bolla, M.; et al. Postoperative irradiation with or without concomitant chemotherapy for locally advanced head and neck cancer. N. Engl. J. Med. 2004, 350, 1945–1952. [Google Scholar] [PubMed]
- Cooper, J.S.; Pajak, T.F.; Forastiere, A.A.; Jacobs, J.; Campbell, B.H.; Saxman, S.B.; Kish, J.A.; Kim, H.E.; Cmelak, A.J.; Rotman, M.; et al. Postoperative concurrent radiotherapy and chemotherapy for high-risk squamous-cell carcinoma of the head and neck. N. Engl. J. Med. 2004, 350, 1937–1944. [Google Scholar] [PubMed]
- Bernier, J.; Cooper, J.S. Chemoradiation after surgery for high-risk head and neck cancer patients: how strong is the evidence? Oncologist 2005, 10, 215–224. [Google Scholar] [CrossRef]
- Song, J.I.; Grandis, J.R. STAT signaling in head and neck cancer. Oncogene 2000, 19, 2489–2495. [Google Scholar] [PubMed]
- Markovic, A.; Chung, C.H. Current role of EGF receptor monoclonal antibodies and tyrosine kinase inhibitors in the management of head and neck squamous cell carcinoma. Expert. Rev. Anticancer Ther. 2012, 12, 1149–1159. [Google Scholar] [PubMed]
- Lai, S.Y.; Johnson, F.M. Defining the role of the JAK-STAT pathway in head and neck and thoracic malignancies: Implications for future therapeutic approaches. Drug Resist. Updat. 2010, 13, 67–78. [Google Scholar] [PubMed]
- Hsiao, J.R.; Jin, Y.T.; Tsai, S.T.; Shiau, A.L.; Wu, C.L.; Su, W.C. Constitutive activation of STAT3 and STAT5 is present in the majority of nasopharyngeal carcinoma and correlates with better prognosis. Br. J. Cancer 2003, 89, 344–349. [Google Scholar] [PubMed]
- Kotoula, V.; Lambaki, S.; Televantou, D.; Kalogera-Fountzila, A.; Nikolaou, A.; Markou, K.; Misailidou, D.; Syrigos, K.N.; Fountzilas, G. STAT-Related Profiles are associated with patient response to targeted treatments in locally advanced SCCHN. Transl. Oncol. 2011, 4, 47–58. [Google Scholar] [PubMed]
- Masuda, M.; Suzui, M.; Yasumatu, R.; Nakashima, T.; Kuratomi, Y.; Azuma, K.; Tomita, K.; Komiyama, S.; Weinstein, I.B. Constitutive activation of signal transducers and activators of transcription 3 correlates with cyclin D1 overexpression and may provide a novel prognostic marker in head and neck squamous cell carcinoma. Cancer Res. 2002, 62, 3351–3355. [Google Scholar] [PubMed]
- Chen, C.C.; Chen, W.C.; Lu, C.H.; Wang, W.H.; Lin, P.Y.; Lee, K.D.; Chen, M.F. Significance of interleukin-6 signaling in the resistance of pharyngeal cancer to irradiation and the epidermal growth factor receptor inhibitor. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76, 1214–1224. [Google Scholar] [PubMed]
- Chen, Y.W.; Chen, K.H.; Huang, P.I.; Chen, Y.C.; Chiou, G.Y.; Lo, W.L.; Tseng, L.M.; Hsu, H.S.; Chang, K.W.; Chiou, S.H. Cucurbitacin I suppressed stem-like property and enhanced radiation-induced apoptosis in head and neck squamous carcinoma—Derived CD44(+)ALDH1(+) cells. Mol. Cancer Ther. 2010, 9, 2879–2892. [Google Scholar] [PubMed]
- Kruser, T.J.; Armstrong, E.A.; Ghia, A.J.; Huang, S.; Wheeler, D.L.; Radinsky, R.; Freeman, D.J.; Harari, P.M. Augmentation of radiation response by panitumumab in models of upper aerodigestive tract cancer. Int. J. Radiat. Oncol. Biol. Phys. 2008, 72, 534–542. [Google Scholar] [PubMed]
- Li, X.; Wang, H.; Lu, X.; Di, B. STAT3 blockade with shRNA enhances radiosensitivity in Hep-2 human laryngeal squamous carcinoma cells. Oncol. Rep. 2010, 23, 345–353. [Google Scholar] [PubMed]
- Bonner, J.A.; Yang, E.S.; Trummell, H.Q.; Nowsheen, S.; Willey, C.D.; Raisch, K.P. Inhibition of STAT-3 results in greater cetuximab sensitivity in head and neck squamous cell carcinoma. Radiother. Oncol. 2011, 99, 339–343. [Google Scholar] [PubMed]
- Adachi, M.; Cui, C.; Dodge, C.T.; Bhayani, M.K.; Lai, S.Y. Targeting STAT3 inhibits growth and enhances radiosensitivity in head and neck squamous cell carcinoma. Oral. Oncol. 2012, 48, 1220–1226. [Google Scholar] [PubMed]
- Pan, Y.; Zhou, F.; Zhang, R.; Claret, F.X. Stat3 inhibitor Stattic exhibits potent antitumor activity and induces chemo- and radio-sensitivity in nasopharyngeal carcinoma. PLoS One 2013, 8, e54565. [Google Scholar] [PubMed]
- Goel, A.; Aggarwal, B.B. Curcumin, the golden spice from Indian saffron, is a chemosensitizer and radiosensitizer for tumors and chemoprotector and radioprotector for normal organs. Nutr. Cancer 2010, 62, 919–930. [Google Scholar] [PubMed]
- Hsu, H.W.; Gridley, D.S.; Kim, P.D.; Hu, S.; de Necochea-Campion, R.; Ferris, R.L.; Chen, C.S.; Mirshahidi, S. Linifanib (ABT-869) enhances radiosensitivity of head and neck squamous cell carcinoma cells. Oral. Oncol. 2013, 49, 591–597. [Google Scholar] [PubMed]
- Achcar Rde, O.; Cagle, P.T.; Jagirdar, J. Expression of activated and latent signal transducer and activator of transcription 3 in 303 non-small cell lung carcinomas and 44 malignant mesotheliomas: Possible role for chemotherapeutic intervention. Arch. Pathol. Lab. Med. 2007, 131, 1350–1360. [Google Scholar]
- Purnell, P.R.; Mack, P.C.; Tepper, C.G.; Evans, C.P.; Green, T.P.; Gumerlock, P.H.; Lara, P.N.; Gandara, D.R.; Kung, H.J.; Gautschi, O. The Src inhibitor AZD0530 blocks invasion and may act as a radiosensitizer in lung cancer cells. J. Thorac. Oncol. 2009, 4, 448–454. [Google Scholar] [PubMed]
- Sun, Y.; Moretti, L.; Giacalone, N.J.; Schleicher, S.; Speirs, C.K.; Carbone, D.P.; Lu, B. Inhibition of JAK2 signaling by TG101209 enhances radiotherapy in lung cancer models. J. Thorac. Oncol. 2011, 6, 699–706. [Google Scholar] [PubMed]
- Lin, Y.C.; Hung, M.S.; Lin, C.K.; Li, J.M.; Lee, K.D.; Li, Y.C.; Chen, M.F.; Chen, J.K.; Yang, C.T. CK2 inhibitors enhance the radiosensitivity of human non-small cell lung cancer cells through inhibition of stat3 activation. Cancer Biother. Radiopharm. 2011, 26, 381–388. [Google Scholar] [PubMed]
- Yin, Z.J.; Jin, F.G.; Liu, T.G.; Fu, E.Q.; Xie, Y.H.; Sun, R.L. Overexpression of STAT3 potentiates growth, survival, and radioresistance of non-small-cell lung cancer (NSCLC) cells. J. Surg. Res. 2011, 171, 675–683. [Google Scholar] [PubMed]
- Hsu, H.S.; Huang, P.I.; Chang, Y.L.; Tzao, C.; Chen, Y.W.; Shih, H.C.; Hung, S.C.; Chen, Y.C.; Tseng, L.M.; Chiou, S.H. Cucurbitacin I inhibits tumorigenic ability and enhances radiochemosensitivity in nonsmall cell lung cancer-derived CD133-positive cells. Cancer 2011, 117, 2970–2985. [Google Scholar] [PubMed]
- De Gruijl, F.R.; van Kranen, H.J.; Mullenders, L.H. UV-induced DNA damage, repair, mutations and oncogenic pathways in skin cancer. J. Photochem. Photobiol. B 2001, 63, 19–27. [Google Scholar] [PubMed]
- Sano, S.; Itami, S.; Takeda, K.; Tarutani, M.; Yamaguchi, Y.; Miura, H.; Yoshikawa, K.; Akira, S.; Takeda, J. Keratinocyte-specific ablation of Stat3 exhibits impaired skin remodeling, but does not affect skin morphogenesis. EMBO J. 1999, 18, 4657–4668. [Google Scholar] [PubMed]
- Kim, D.J.; Chan, K.S.; Sano, S.; Digiovanni, J. Signal transducer and activator of transcription 3 (Stat3) in epithelial carcinogenesis. Mol. Carcinog. 2007, 46, 725–731. [Google Scholar] [PubMed]
- Sano, S.; Chan, K.S.; DiGiovanni, J. Impact of Stat3 activation upon skin biology: A dichotomy of its role between homeostasis and diseases. J. Dermatol. Sci. 2008, 50, 1–14. [Google Scholar] [PubMed]
- Sano, S.; Chan, K.S.; Kira, M.; Kataoka, K.; Takagi, S.; Tarutani, M.; Itami, S.; Kiguchi, K.; Yokoi, M.; Sugasawa, K.; et al. Signal transducer and activator of transcription 3 is a key regulator of keratinocyte survival and proliferation following UV irradiation. Cancer Res. 2005, 65, 5720–5729. [Google Scholar] [PubMed]
- Dzwierzynski, W.W. Managing malignant melanoma. Plast. Reconstr. Surg. 2013, 132, 446e–460e. [Google Scholar] [PubMed]
- Johnson, G.E.; Ivanov, V.N.; Hei, T.K. Radiosensitization of melanoma cells through combined inhibition of protein regulators of cell survival. Apoptosis 2008, 13, 790–802. [Google Scholar] [PubMed]
- Bonner, J.A.; Trummell, H.Q.; Willey, C.D.; Plants, B.A.; Raisch, K.P. Inhibition of STAT-3 results in radiosensitization of human squamous cell carcinoma. Radiother. Oncol. 2009, 92, 339–344. [Google Scholar] [PubMed]
- Karras, J.G.; Wang, Z.; Huo, L.; Howard, R.G.; Frank, D.A.; Rothstein, T.L. Signal transducer and activator of transcription-3 (STAT3) is constitutively activated in normal, self-renewing B-1 cells but only inducibly expressed in conventional B lymphocytes. J. Exp. Med. 1997, 185, 1035–1042. [Google Scholar]
- Sugino, K.; Ito, K.; Mimura, T.; Nagahama, M.; Fukunari, N.; Kubo, A.; Iwasaki, H. The important role of operations in the management of anaplastic thyroid carcinoma. Surgery 2002, 131, 245–248. [Google Scholar] [PubMed]
- Zito, G.; Richiusa, P.; Bommarito, A.; Carissimi, E.; Russo, L.; Coppola, A.; Zerilli, M.; Rodolico, V.; Criscimanna, A.; Amato, M.; et al. In vitro identification and characterization of CD133(pos) cancer stem-like cells in anaplastic thyroid carcinoma cell lines. PLoS One 2008, 3, e3544. [Google Scholar] [PubMed]
- Tseng, L.M.; Huang, P.I.; Chen, Y.R.; Chen, Y.C.; Chou, Y.C.; Chen, Y.W.; Chang, Y.L.; Hsu, H.S.; Lan, Y.T.; Chen, K.H.; et al. Targeting signal transducer and activator of transcription 3 pathway by cucurbitacin I diminishes self-renewing and radiochemoresistant abilities in thyroid cancer-derived CD133+ cells. J. Pharmacol. Exp. Ther. 2012, 341, 410–423. [Google Scholar] [PubMed]
- Huang, C.Y.; Lin, C.S.; Tai, W.T.; Hsieh, C.Y.; Shiau, C.W.; Cheng, A.L.; Chen, K.F. Sorafenib enhances radiation-induced apoptosis in hepatocellular carcinoma by inhibiting STAT3. Int. J. Radiat. Oncol. Biol. Phys. 2013, 86, 456–462. [Google Scholar] [PubMed]
- Snyder, M.; Huang, X.Y.; Zhang, J.J. Identification of novel direct Stat3 target genes for control of growth and differentiation. J. Biol. Chem. 2008, 283, 3791–3798. [Google Scholar] [PubMed]
- Haviland, R. Identification of novel STAT3 target genes associated with oncogenesis. PhD Thesis, University of South Florida, Tampa, FL, USA, 2011. [Google Scholar]
- Timofeeva, O.A.; Tarasova, N.I.; Zhang, X.; Chasovskikh, S.; Cheema, A.K.; Wang, H.; Brown, M.L.; Dritschilo, A. STAT3 suppresses transcription of proapoptotic genes in cancer cells with the involvement of its N-terminal domain. Proc. Natl. Acad. Sci. USA 2013, 110, 1267–1272. [Google Scholar] [PubMed]
- Bourillot, P.Y.; Aksoy, I.; Schreiber, V.; Wianny, F.; Schulz, H.; Hummel, O.; Hubner, N.; Savatier, P. Novel STAT3 target genes exert distinct roles in the inhibition of mesoderm and endoderm differentiation in cooperation with Nanog. Stem Cells 2009, 27, 1760–1771. [Google Scholar] [PubMed]
- Courapied, S.; Sellier, H.; de Carne Trecesson, S.; Vigneron, A.; Bernard, A.C.; Gamelin, E.; Barre, B.; Coqueret, O. The cdk5 kinase regulates the STAT3 transcription factor to prevent DNA damage upon topoisomerase I inhibition. J. Biol. Chem. 2010, 285, 26765–26778. [Google Scholar] [PubMed]
- Barry, S.P.; Townsend, P.A.; Knight, R.A.; Scarabelli, T.M.; Latchman, D.S.; Stephanou, A. STAT3 modulates the DNA damage response pathway. Int. J. Exp. Pathol. 2010, 91, 506–514. [Google Scholar] [PubMed]
- Ushijima, R.; Sakaguchi, N.; Kano, A.; Maruyama, A.; Miyamoto, Y.; Sekimoto, T.; Yoneda, Y.; Ogino, K.; Tachibana, T. Extracellular signal-dependent nuclear import of STAT3 is mediated by various importin alphas. Biochem. Biophys. Res. Commun. 2005, 330, 880–886. [Google Scholar] [PubMed]
- Cimica, V.; Chen, H.C.; Iyer, J.K.; Reich, N.C. Dynamics of the STAT3 transcription factor: Nuclear import dependent on Ran and importin-beta1. PLoS One 2011, 6, e20188. [Google Scholar] [PubMed]
- Ma, J.; Cao, X. Regulation of Stat3 nuclear import by importin alpha5 and importin alpha7 via two different functional sequence elements. Cell Signal. 2006, 18, 1117–1126. [Google Scholar] [PubMed]
- Bhattacharya, S.; Schindler, C. Regulation of Stat3 nuclear export. J. Clin. Invest. 2003, 111, 553–559. [Google Scholar] [PubMed]
- Gao, J.; McConnell, M.J.; Yu, B.; Li, J.; Balko, J.M.; Black, E.P.; Johnson, J.O.; Lloyd, M.C.; Altiok, S.; Haura, E.B. MUC1 is a downstream target of STAT3 and regulates lung cancer cell survival and invasion. Int. J. Oncol. 2009, 35, 337–345. [Google Scholar] [PubMed]
- Kamran, M.Z.; Patil, P.; Gude, R.P. Role of STAT3 in cancer metastasis and translational advances. Biomed. Res. Int. 2013, 2013, 421821. [Google Scholar] [PubMed]
- Priester, M.; Copanaki, E.; Vafaizadeh, V.; Hensel, S.; Bernreuther, C.; Glatzel, M.; Seifert, V.; Groner, B.; Kogel, D.; Weissenberger, J. STAT3 silencing inhibits glioma single cell infiltration and tumor growth. Neuro-oncology 2013, 15, 840–852. [Google Scholar] [PubMed]
- Wu, Y.; Diab, I.; Zhang, X.; Izmailova, E.S.; Zehner, Z.E. Stat3 enhances vimentin gene expression by binding to the antisilencer element and interacting with the repressor protein, ZBP-89. Oncogene 2004, 23, 168–178. [Google Scholar] [PubMed]
- Colomiere, M.; Ward, A.C.; Riley, C.; Trenerry, M.K.; Cameron-Smith, D.; Findlay, J.; Ackland, L.; Ahmed, N. Cross talk of signals between EGFR and IL-6R through JAK2/STAT3 mediate epithelial-mesenchymal transition in ovarian carcinomas. Br. J. Cancer 2009, 100, 134–144. [Google Scholar] [PubMed]
- Avtanski, D.B.; Nagalingam, A.; Bonner, M.Y.; Arbiser, J.L.; Saxena, N.K.; Sharma, D. Honokiol inhibits epithelial-mesenchymal transition in breast cancer cells by targeting signal transducer and activator of transcription 3/Zeb1/E-cadherin axis. Mol. Oncol. 2014, 8, 565–580. [Google Scholar] [PubMed]
- Ng, I.H.; Yeap, Y.Y.; Ong, L.S.; Jans, D.A.; Bogoyevitch, M.A. Oxidative stress impairs multiple regulatory events to drive persistent cytokine-stimulated STAT3 phosphorylation. Biochim. Biophys. Acta 2014, 1843, 483–494. [Google Scholar] [PubMed]
- Gough, D.J.; Sehgal, P.; Levy, D.E. Nongenomic functions of STAT3. In Jak-Stat Signaling: From Basics to Disease; Decker, T., Müller, M., Eds.; Springer Vienna: Vienna, Austria, 2012; pp. 91–98. [Google Scholar]
- Zouein, F.A.; Kurdi, M.; Booz, G.W. Dancing rhinos in stilettos: The amazing saga of the genomic and nongenomic actions of STAT3 in the heart. JAKSTAT 2013, 2, e24352. [Google Scholar]
- Wegrzyn, J.; Potla, R.; Chwae, Y.J.; Sepuri, N.B.; Zhang, Q.; Koeck, T.; Derecka, M.; Szczepanek, K.; Szelag, M.; Gornicka, A.; et al. Function of mitochondrial Stat3 in cellular respiration. Science 2009, 323, 793–797. [Google Scholar] [PubMed]
- Meier, J.A.; Larner, A.C. Toward a new STATe: The role of STATs in mitochondrial function. Semin. Immunol. 2014, 26, 20–28. [Google Scholar] [PubMed]
- Elschami, M.; Scherr, M.; Philippens, B.; Gerardy-Schahn, R. Reduction of STAT3 expression induces mitochondrial dysfunction and autophagy in cardiac HL-1 cells. Eur. J. Cell Biol. 2013, 92, 21–29. [Google Scholar] [PubMed]
- Yoon, S.; Woo, S.U.; Kang, J.H.; Kim, K.; Kwon, M.H.; Park, S.; Shin, H.J.; Gwak, H.S.; Chwae, Y.J. STAT3 transcriptional factor activated by reactive oxygen species induces IL6 in starvation-induced autophagy of cancer cells. Autophagy 2010, 6, 1125–1138. [Google Scholar] [PubMed]
- Zhang, Q.; Raje, V.; Yakovlev, V.A.; Yacoub, A.; Szczepanek, K.; Meier, J.; Derecka, M.; Chen, Q.; Hu, Y.; Sisler, J.; et al. Mitochondrial localized Stat3 promotes breast cancer growth via phosphorylation of serine 727. J. Biol. Chem. 2013, 288, 31280–31288. [Google Scholar] [PubMed]
- Mackenzie, G.G.; Huang, L.; Alston, N.; Ouyang, N.; Vrankova, K.; Mattheolabakis, G.; Constantinides, P.P.; Rigas, B. Targeting mitochondrial STAT3 with the novel phospho-valproic acid (MDC-1112) inhibits pancreatic cancer growth in mice. PLoS One 2013, 8, e61532. [Google Scholar] [PubMed]
- Darnell, J.E., Jr. Transcription factors as targets for cancer therapy. Nat. Rev. Cancer 2002, 2, 740–749. [Google Scholar] [PubMed]
- Yue, P.; Turkson, J. Targeting STAT3 in cancer: How successful are we? Expert Opin. Investig. Drugs 2009, 18, 45–56. [Google Scholar] [CrossRef]
- Mankan, A.K.; Greten, F.R. Inhibiting signal transducer and activator of transcription 3: Rationality and rationale design of inhibitors. Expert Opin. Investig. Drugs 2011, 20, 1263–1275. [Google Scholar] [PubMed]
- Nelson, E.A.; Sharma, S.V.; Settleman, J.; Frank, D.A. A chemical biology approach to developing STAT inhibitors: Molecular strategies for accelerating clinical translation. Oncotarget 2011, 2, 518–524. [Google Scholar] [PubMed]
- Debnath, B.; Xu, S.; Neamati, N. Small molecule inhibitors of signal transducer and activator of transcription 3 (Stat3) protein. J. Med. Chem. 2012, 55, 6645–6668. [Google Scholar] [PubMed]
- Reddy, M.M.; Deshpande, A.; Sattler, M. Targeting JAK2 in the therapy of myeloproliferative neoplasms. Expert Opin. Ther. Targets 2012, 16, 313–324. [Google Scholar] [PubMed]
- Harry, B.L.; Eckhardt, S.G.; Jimeno, A. JAK2 inhibition for the treatment of hematologic and solid malignancies. Expert Opin. Investig. Drugs 2012, 21, 637–655. [Google Scholar] [PubMed]
- Sansone, P.; Bromberg, J. Targeting the interleukin-6/Jak/stat pathway in human malignancies. J. Clin. Oncol. 2012, 30, 1005–1014. [Google Scholar] [PubMed]
- Laurie, S.A.; Goss, G.D.; Shepherd, F.A.; Reaume, M.N.; Nicholas, G.; Philip, L.; Wang, L.; Schwock, J.; Hirsh, V.; Oza, A.; et al. A phase II trial of saracatinib, an inhibitor of src kinases, in previously-treated advanced non-small-cell lung cancer: The princess margaret hospital phase II consortium. Clin. Lung Cancer 2014, 15, 52–57. [Google Scholar] [PubMed]
- Johnson, F.M.; Saigal, B.; Talpaz, M.; Donato, N.J. Dasatinib (BMS-354825) tyrosine kinase inhibitor suppresses invasion and induces cell cycle arrest and apoptosis of head and neck squamous cell carcinoma and non-small cell lung cancer cells. Clin. Cancer Res. 2005, 11, 6924–6932. [Google Scholar] [PubMed]
- Guo, Y.; Xu, F.; Lu, T.; Duan, Z.; Zhang, Z. Interleukin-6 signaling pathway in targeted therapy for cancer. Cancer Treat Rev. 2012, 38, 904–910. [Google Scholar] [PubMed]
- NCT01066663, a phase I/II study of pyrimethamine, a STAT3 inhibitor, for the treatment of relapsed chronic lymphocytic Leukemia/small Lymphocytic Lymphoma. Available online: http://clinicaltrials.gov (accessed on 9 February 2010).
- NCT00529113, a randomized phase I/II study with gemcitabine and RTA 402 or gemcitabine and placebo for patients with unresectable pancreatic cancer. Available online: http://clinicaltrials.gov (accessed on 12 September 2007).
- Hong, D.S.; Kurzrock, R.; Supko, J.G.; Lawrence, D.P.; Wheler, J.J.; Meyer, C.J.; Mier, J.W.; Andreeff, M.; Shapiro, G.I.; Dezube, B.J. Phase I trial with a novel oral NF-{kappa}B/STAT3 inhibitor RTA 402 in patients with solid tumors and lymphoid malignancies. J. Clin. Oncol. 2008, 26 15S, 3517. [Google Scholar] [PubMed]
- Meyer, S.C.; Levine, R.L. Molecular pathways: Molecular basis for sensitivity and resistance to JAK kinase inhibitors. Clin. Cancer Res. 2014, 20, 2051–2059. [Google Scholar] [PubMed]
- Guo, X.; Qiu, J.; Tu, T.; Yang, X.; Deng, L.; Anders, R.A.; Zhou, L.; Fu, Y.X. Induction of innate lymphoid cell-derived interleukin-22 by the transcription factor STAT3 mediates protection against intestinal infection. Immunity 2014, 40, 25–39. [Google Scholar] [PubMed]
- Takeda, K.; Noguchi, K.; Shi, W.; Tanaka, T.; Matsumoto, M.; Yoshida, N.; Kishimoto, T.; Akira, S. Targeted disruption of the mouse Stat3 gene leads to early embryonic lethality. Proc. Natl. Acad. Sci. USA 1997, 94, 3801–3804. [Google Scholar] [PubMed]
- Inghirami, G.; Chiarle, R.; Simmons, W.J.; Piva, R.; Schlessinger, K.; Levy, D.E. New and old functions of STAT3: A pivotal target for individualized treatment of cancer. Cell Cycle 2005, 4, 1131–1133. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spitzner, M.; Ebner, R.; Wolff, H.A.; Ghadimi, B.M.; Wienands, J.; Grade, M. STAT3: A Novel Molecular Mediator of Resistance to Chemoradiotherapy. Cancers 2014, 6, 1986-2011. https://doi.org/10.3390/cancers6041986
Spitzner M, Ebner R, Wolff HA, Ghadimi BM, Wienands J, Grade M. STAT3: A Novel Molecular Mediator of Resistance to Chemoradiotherapy. Cancers. 2014; 6(4):1986-2011. https://doi.org/10.3390/cancers6041986
Chicago/Turabian StyleSpitzner, Melanie, Reinhard Ebner, Hendrik A. Wolff, B. Michael Ghadimi, Jürgen Wienands, and Marian Grade. 2014. "STAT3: A Novel Molecular Mediator of Resistance to Chemoradiotherapy" Cancers 6, no. 4: 1986-2011. https://doi.org/10.3390/cancers6041986