
The Hedgehog Signalling Pathway in Cell Migration and Guidance: What We Have Learned from Drosophila melanogaster

Abstract
:1. Introduction
2. Hh Signaling in Border Cell Migration
3. Hh Signaling in Germ Cell Migration
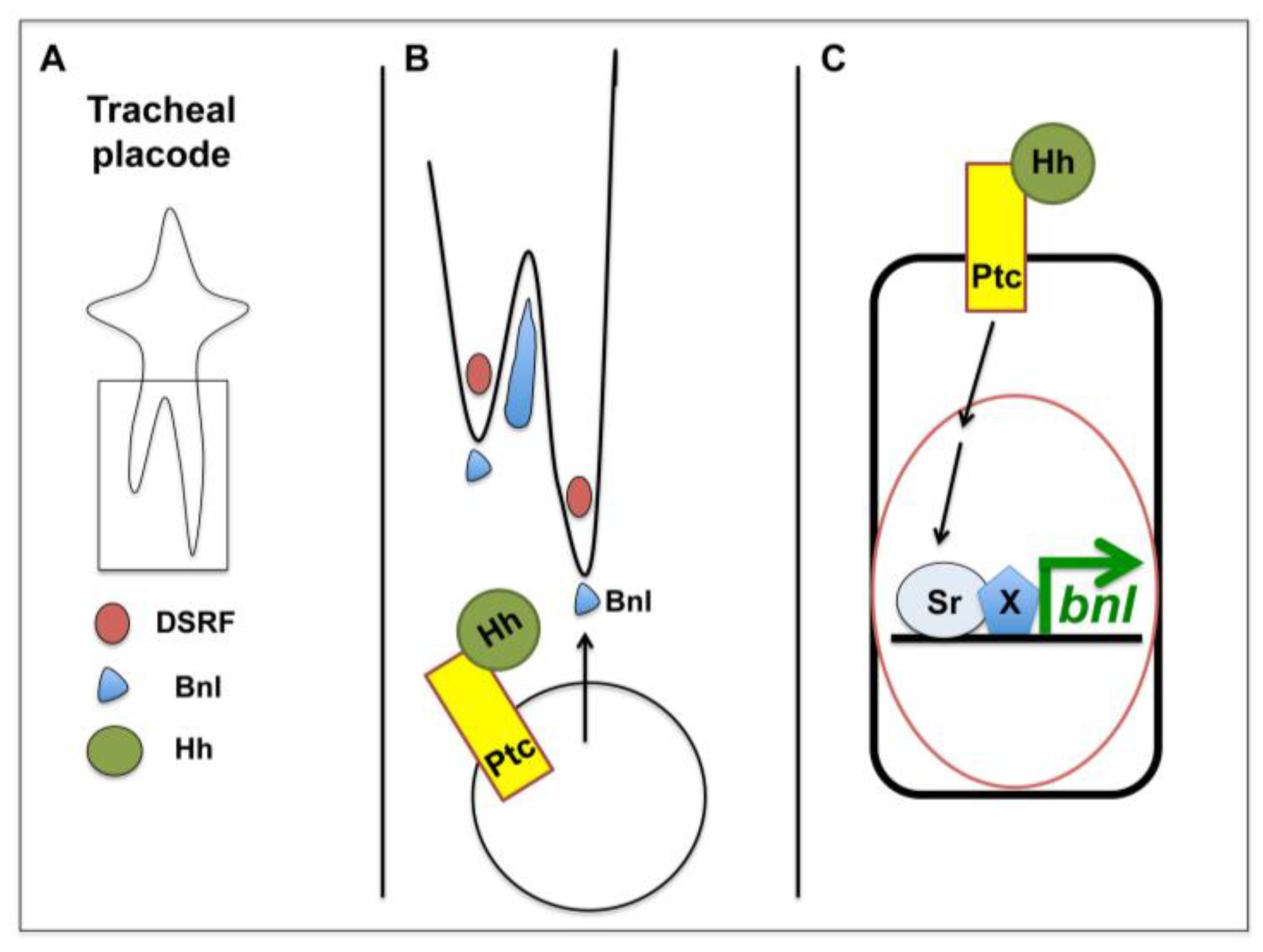
4. Hh Signalling in Tracheal Cell Migration

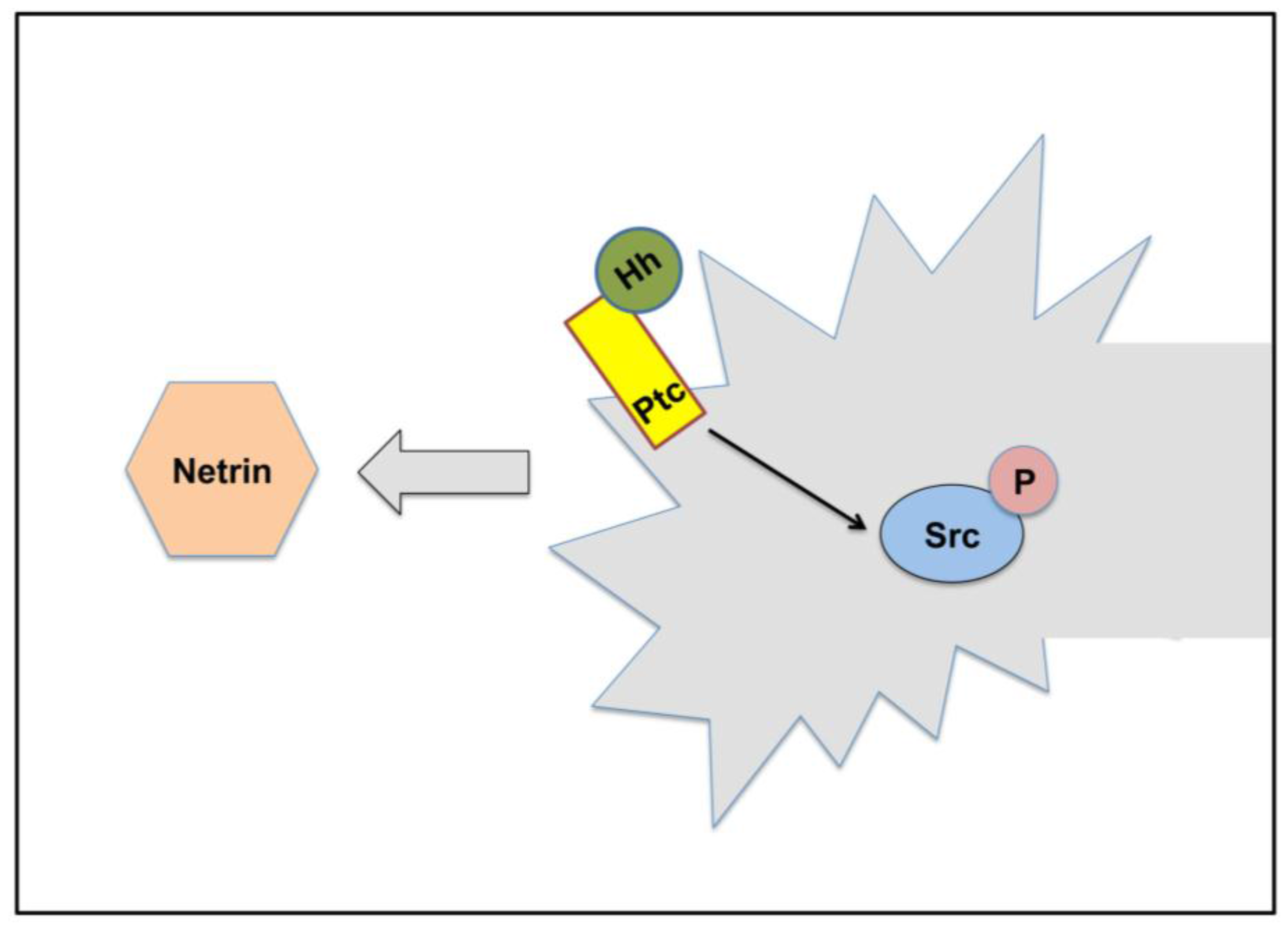
5. Hh signaling in Axonal Guidance

6. Hh signaling in Glial Cell Migration
7. Same Signaling Ligand for Different Cellular Responses

{kind=link}
{kind=link}
| Cell Type | Hh Effect | Hh Signalling | Reference |
|---|---|---|---|
| Border cells | Cell-shape changes | Canonical | [10] |
| Germ cells | Chemoattraction | Non-canonical | [25,29] |
| Tracheal cells | Chemoattraction (via Bnl) | Canonical | [45] |
| Axons | Chemoattraction | Non-canonical | [61] |
| Glial cells | Spatio-temporal control | Non-canonical | [70,71] |
References
- Ingham, P.W.; Nakano, Y.; Seger, C. Mechanisms and functions of hedgehog signalling across the metazoa. Nat. Rev. Genet. 2011, 12, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, J.; Thérond, P.P. The mechanisms of hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Nozawa, Y.I.; Chuang, P.-T. The path to chemotaxis and transcription is smoothened. Sci. Signal. 2012, 5, pe35. [Google Scholar] [CrossRef] [PubMed]
- Choy, S.W.; Cheng, S.H. Hedgehog signaling. Hedgehog Signal. 2012, 88, 1–23. [Google Scholar]
- Robbins, D.J.; Fei, D.L.; Riobo, N.A. The hedgehog signal transduction network. Sci. Signal. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Aza-Blanc, P.; Kornberg, T.B. Ci: A complex transducer of the hedgehog signal. Trends Genet. 1999, 15, 458–462. [Google Scholar] [CrossRef]
- Biehs, B.; Kechris, K.; Liu, S.; Kornberg, T.B. Hedgehog targets in the drosophila embryo and the mechanisms that generate tissue-specific outputs of hedgehog signaling. Development 2010, 137, 3887–3898. [Google Scholar] [CrossRef] [PubMed]
- Torroja, C.; Gorfinkiel, N.; Guerrero, I. Patched controls the hedgehog gradient by endocytosis in a dynamin-dependent manner, but this internalization does not play a major role in signal transduction. Development 2004, 131, 2395–2408. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Struhl, G. Dual roles for patched in sequestering and transducing hedgehog. Cell 1996, 87, 553–563. [Google Scholar] [CrossRef]
- Geisbrecht, E.R.; Sawant, K.; Su, Y.; Liu, Z.C.; Silver, D.L.; Burtscher, A.; Wang, X.; Zhu, A.J.; McDonald, J.A. Genetic interaction screens identify a role for hedgehog signaling in drosophila border cell migration. Dev. Dyn. 2013, 242, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Ruiz i Altaba, A. Gli proteins and hedgehog signaling: Development and cancer. Trends Genet. 1999, 15, 418–425. [Google Scholar] [CrossRef]
- Montell, D.J. Border-cell migration: The race is on. Nat. Rev. Mol. Cell Biol. 2003, 4, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Silver, D.L.; Montell, D.J. Paracrine signaling through the jak/stat pathway activates invasive behavior of ovarian epithelial cells in drosophila. Cell 2001, 107, 831–841. [Google Scholar] [CrossRef]
- Silver, D.L.; Geisbrecht, E.R.; Montell, D.J. Requirement for jak/stat signaling throughout border cell migration in drosophila. Development 2005, 132, 3483–3492. [Google Scholar] [CrossRef] [PubMed]
- Beccari, S.; Teixeira, L.; Rorth, P. The jak/stat pathway is required for border cell migration during drosophila oogenesis. Mech. Dev. 2002, 111, 115–123. [Google Scholar] [CrossRef]
- Jang, A.C.; Chang, Y.C.; Bai, J.; Montell, D. Border-cell migration requires integration of spatial and temporal signals by the btb protein abrupt. Nat. Cell Biol. 2009, 11, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Llense, F.; Martín-Blanco, E. Jnk signaling controls border cell cluster integrity and collective cell migration. Curr. Biol. 2008, 18, 538–544. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.A.; Pinheiro, E.M.; Montell, D.J. Pvf1, a pdgf/vegf homolog, is sufficient to guide border cells and interacts genetically with taiman. Development 2003, 130, 3469–3478. [Google Scholar] [CrossRef] [PubMed]
- Duchek, P.; Rorth, P. Guidance of cell migration by egf receptor signaling during drosophila oogenesis. Science 2001, 291, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.J.; Lin, H.; Ingham, P.W.; Spradling, A.C. Hedgehog is required for the proliferation and specification of ovarian somatic cells prior to egg chamber formation in drosophila. Development 1996, 122, 1125–1135. [Google Scholar] [PubMed]
- Santos, A.C.; Lehmann, R. Germ cell specification and migration in drosophila and beyond. Curr. Biol. 2004, 14, R578–R589. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, P.S.; Sano, H.; Renault, A.D.; Barbosa, V.; Fuse, N.; Lehmann, R. Tre1 gpcr initiates germ cell transepithelial migration by regulating drosophila melanogaster e-cadherin. J. Cell Biol. 2008, 183, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.; DiNardo, S. Specification, migration and assembly of the somatic cells of the drosophila gonad. Development 1995, 121, 1815–1825. [Google Scholar] [PubMed]
- Van Doren, M.; Broihier, H.T.; Moore, L.A.; Lehmann, R. Hmg-coa reductase guides migrating primordial germ cells. Nature 1998, 396, 466–469. [Google Scholar] [PubMed]
- Deshpande, G.; Swanhart, L.; Chiang, P.; Schedl, P. Hedgehog signaling in germ cell migration. Cell 2001, 106, 759–769. [Google Scholar] [CrossRef]
- Deshpande, G.; Sethi, N.; Schedl, P. Toutvelu, a regulator of heparan sulfate proteoglycan biosynthesis, controls guidance cues for germ-cell migration. Genetics 2007, 176, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, G.; Godishala, A.; Schedl, P. Ggamma1, a downstream target for the hmgcr-isoprenoid biosynthetic pathway, is required for releasing the hedgehog ligand and directing germ cell migration. PLoS Genet. 2009, 5, e1000333. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, G.; Schedl, P. Hmgcoa reductase potentiates hedgehog signaling in drosophila melanogaster. Dev. Cell 2005, 9, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, G.; Zhou, K.; Wan, J.Y.; Friedrich, J.; Jourjine, N.; Smith, D.; Schedl, P. The hedgehog pathway gene shifted functions together with the hmgcr-dependent isoprenoid biosynthetic pathway to orchestrate germ cell migration. PLoS Genet. 2013, 9, e1003720. [Google Scholar] [CrossRef] [PubMed]
- Renault, A.D.; Ricardo, S.; Kunwar, P.S.; Santos, A.; Starz-Gaiano, M.; Stein, J.A.; Lehmann, R. Hedgehog does not guide migrating drosophila germ cells. Dev. Biol. 2009, 328, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Affolter, M.; Caussinus, E. Tracheal branching morphogenesis in drosophila: New insights into cell behaviour and organ architecture. Development 2008, 135, 2055–2064. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.; Samakovlis, C.; Krasnow, M.A. Branchless encodes a drosophila fgf homolog that controls tracheal cell migration and the pattern of branching. Cell 1996, 87, 1091–1101. [Google Scholar] [CrossRef]
- Llimargas, M.; Casanova, J. Ventral veinless, a pou domain transcription factor, regulates different transduction pathways required for tracheal branching in drosophila. Development 1997, 124, 3273–3281. [Google Scholar] [PubMed]
- Llimargas, M. Wingless and its signalling pathway have common and separable functions during tracheal development. Development 2000, 127, 4407–4417. [Google Scholar] [PubMed]
- Vincent, S.; Ruberte, E.; Grieder, N.C.; Chen, C.K.; Haerry, T.; Schuh, R.; Affolter, M. Dpp controls tracheal cell migration along the dorsoventral body axis of the drosophila embryo. Development 1997, 124, 2741–2750. [Google Scholar] [PubMed]
- Chihara, T.; Hayashi, S. Control of tracheal tubulogenesis by wingless signaling. Development 2000, 127, 4433–4442. [Google Scholar] [PubMed]
- Wappner, P.; Gabay, L.; Shilo, B.Z. Interactions between the egf receptor and dpp pathways establish distinct cell fates in the tracheal placodes. Development 1997, 124, 4707–4716. [Google Scholar] [PubMed]
- Franch-Marro, X.; Casanova, J. The alternative migratory pathways of the drosophila tracheal cells are associated with distinct subsets of mesodermal cells. Dev. Biol. 2000, 227, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Boube, M.; Martin-Bermudo, M.D.; Brown, N.H.; Casanova, J. Specific tracheal migration is mediated by complementary expression of cell surface proteins. Genes Dev. 2001, 15, 1554–1562. [Google Scholar] [CrossRef] [PubMed]
- Levi, B.P.; Ghabrial, A.S.; Krasnow, M.A. Drosophila talin and integrin genes are required for maintenance of tracheal terminal branches and luminal organization. Development 2006, 133, 2383–2393. [Google Scholar] [CrossRef] [PubMed]
- Urbano, J.M.; Domínguez-Giménez, P.; Estrada, B.; Martín-Bermudo, M.D. Ps integrins and laminins: Key regulators of cell migration during drosophila embryogenesis. PLoS ONE 2011, 6, e23893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klambt, C.; Glazer, L.; Shilo, B. Breathless, a Drosophila FGF receptor homolog, is essential for migration of tracheal and specific midline glial cells. Genes Dev. 1992, 6, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Hacohen, N.; Krasnow, M.; Montell, D.J. Regulated breathless receptor tyrosine kinase activity required to pattern cell migration and branching in the drosophila tracheal system. Genes Dev. 1996, 10, 2912–2921. [Google Scholar] [CrossRef] [PubMed]
- Araújo, S.J.; Casanova, J. Sequoia establishes tip-cell number in drosophila trachea by regulating fgf levels. J. Cell Sci. 2011, 124, 2335–2340. [Google Scholar] [CrossRef] [PubMed]
- Butí, E.; Mesquita, D.; Araújo, S.J. Hedgehog is a positive regulator of fgf signalling during embryonic tracheal cell migration. PLoS ONE 2014, 9, e92682. [Google Scholar] [CrossRef] [PubMed]
- Glazer, L.; Shilo, B.Z. Hedgehog signaling patterns the tracheal branches. Development 2001, 128, 1599–1606. [Google Scholar] [PubMed]
- Matsuda, R.; Hosono, C.; Saigo, K.; Samakovlis, C. The intersection of the extrinsic hedgehog and wnt/wingless signals with the intrinsic hox code underpins branching pattern and tube shape diversity in the drosophila airways. PLoS Genet. 2015, 11, e1004929. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Chihara, T.; Hayashi, S. Hedgehog and decapentaplegic instruct polarized growth of cell extensions in the drosophila trachea. Development 2004, 131, 5253–5261. [Google Scholar] [CrossRef] [PubMed]
- Araújo, S.J.; Tear, G. Axon guidance mechanisms and molecules: Lessons from invertebrates. Nat. Rev. Neurosci. 2003, 4, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.A.; Bashaw, G.J. Axon guidance at the midline: Of mice and flies. Curr. Opin. Neurobiol. 2010, 20, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.J.; Zou, Y. Navigating intermediate targets: The nervous system midline. Cold Spring Harb. Perspect. Biol. 2010, 2, a002055. [Google Scholar] [CrossRef] [PubMed]
- Charron, F.; Tessier-Lavigne, M. Novel brain wiring functions for classical morphogens: A role as graded positional cues in axon guidance. Development 2005, 132, 2251–2262. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Camacho, C.; Bovolenta, P. Emerging mechanisms in morphogen-mediated axon guidance. BioEssays 2009, 31, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; McKinnon, R.D.; Kokel, M.; Thomas, J.B. Wnt-mediated axon guidance via the drosophila derailed receptor. Nature 2003, 422, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Umetsu, D.; Murakami, S.; Yasugi, T.; Tabata, T. Dwnt4 regulates the dorsoventral specificity of retinal projections in the drosophila melanogaster visual system. Nat. Neurosci. 2006, 9, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Bossing, T.; Brand, A.H. Determination of cell fate along the anteroposterior axis of the drosophila ventral midline. Development 2006, 133, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.R. The midline glia of drosophila: A molecular genetic model for the developmental functions of glia. Prog. Neurobiol. 2000, 62, 475–508. [Google Scholar] [CrossRef]
- Watson, J.D.; Wheeler, S.R.; Stagg, S.B.; Crews, S.T. Drosophila hedgehog signaling and engrailed-runt mutual repression direct midline glia to alternative ensheathing and non-ensheathing fates. Development 2011, 138, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.H.; Schafer, B.; Goodman, C.S.; Holmgren, R. The role of segment polarity genes during drosophila neurogenesis. Genes Dev. 1989, 3, 890–904. [Google Scholar] [CrossRef] [PubMed]
- Merianda, T.T.; Botta, V.; Bhat, K.M. Patched regulation of axon guidance is by specifying neural identity in the drosophila nerve cord. Dev. Genes Evol. 2005, 215, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Ricolo, D.; Butí, E.; Araújo, S.J. Drosophila melanogaster hedgehog cooperates with frazzled to guide axons through a non-canonical signalling pathway. Mech. Dev. 2015, 137, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Von Hilchen, C.M.; Beckervordersandforth, R.M.; Rickert, C.; Technau, G.M.; Altenhein, B. Identity, origin, and migration of peripheral glial cells in the drosophila embryo. Mech. Dev. 2008, 125, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Silies, M.; Klämbt, C. Adhesion and signaling between neurons and glial cells in drosophila. Curr. Opin. Neurobiol. 2011, 21, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Silies, M.; Klämbt, C. Apc/c(fzr/cdh1)-dependent regulation of cell adhesion controls glial migration in the drosophila pns. Nat. Neurosci. 2010, 13, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Hummel, T.; Krukkert, K.; Roos, J.; Davis, G.; Klämbt, C. Drosophila futsch/22c10 is a map1b-like protein required for dendritic and axonal development. Neuron 2000, 26, 357–370. [Google Scholar] [CrossRef]
- Hidalgo, A. Neuron-glia interactions during axon guidance in drosophila. Biochem. Soc. Trans. 2003, 31, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Kinrade, E.F.; Brates, T.; Tear, G.; Hidalgo, A. Roundabout signalling, cell contact and trophic support confine longitudinal glia and axons in the drosophila cns. Development 2001, 128, 207–216. [Google Scholar] [PubMed]
- Choi, K.W.; Benzer, S. Migration of glia along photoreceptor axons in the developing drosophila eye. Neuron 1994, 12, 423–431. [Google Scholar] [CrossRef]
- Rangarajan, R.; Gong, Q.; Gaul, U. Migration and function of glia in the developing drosophila eye. Development 1999, 126, 3285–3292. [Google Scholar] [PubMed]
- Hummel, T.; Attix, S.; Gunning, D.; Zipursky, S.L. Temporal control of glial cell migration in the drosophila eye requires gilgamesh, hedgehog, and eye specification genes. Neuron 2002, 33, 193–203. [Google Scholar] [CrossRef]
- Pielage, J.; Kippert, A.; Zhu, M.; Klämbt, C. The drosophila transmembrane protein fear-of-intimacy controls glial cell migration. Dev. Biol. 2004, 275, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Van Doren, M.; Mathews, W.R.; Samuels, M.; Moore, L.A.; Broihier, H.T.; Lehmann, R. Fear of intimacy encodes a novel transmembrane protein required for gonad morphogenesis in drosophila. Development 2003, 130, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, T.; Choi, I.; Banerjee, U. Genetic analysis of fibroblast growth factor signaling in the drosophila eye. G3 2012, 2, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.M.; Lee, T.; Andrews, C.M.; Shilo, B.Z.; Montell, D.J. The breathless fgf receptor homolog, a downstream target of drosophila c/ebp in the developmental control of cell migration. Development 1995, 121, 2255–2263. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araújo, S.J. The Hedgehog Signalling Pathway in Cell Migration and Guidance: What We Have Learned from Drosophila melanogaster. Cancers 2015, 7, 2012-2022. https://doi.org/10.3390/cancers7040873
Araújo SJ. The Hedgehog Signalling Pathway in Cell Migration and Guidance: What We Have Learned from Drosophila melanogaster. Cancers. 2015; 7(4):2012-2022. https://doi.org/10.3390/cancers7040873
Chicago/Turabian StyleAraújo, Sofia J. 2015. "The Hedgehog Signalling Pathway in Cell Migration and Guidance: What We Have Learned from Drosophila melanogaster" Cancers 7, no. 4: 2012-2022. https://doi.org/10.3390/cancers7040873