
The Effect of Stromal Integrin β3-Deficiency on Two Different Tumors in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cell Lines
2.2. Animal Model
2.3. Establishing Primary Tumors
2.4. Interstitial Fluid Pressure
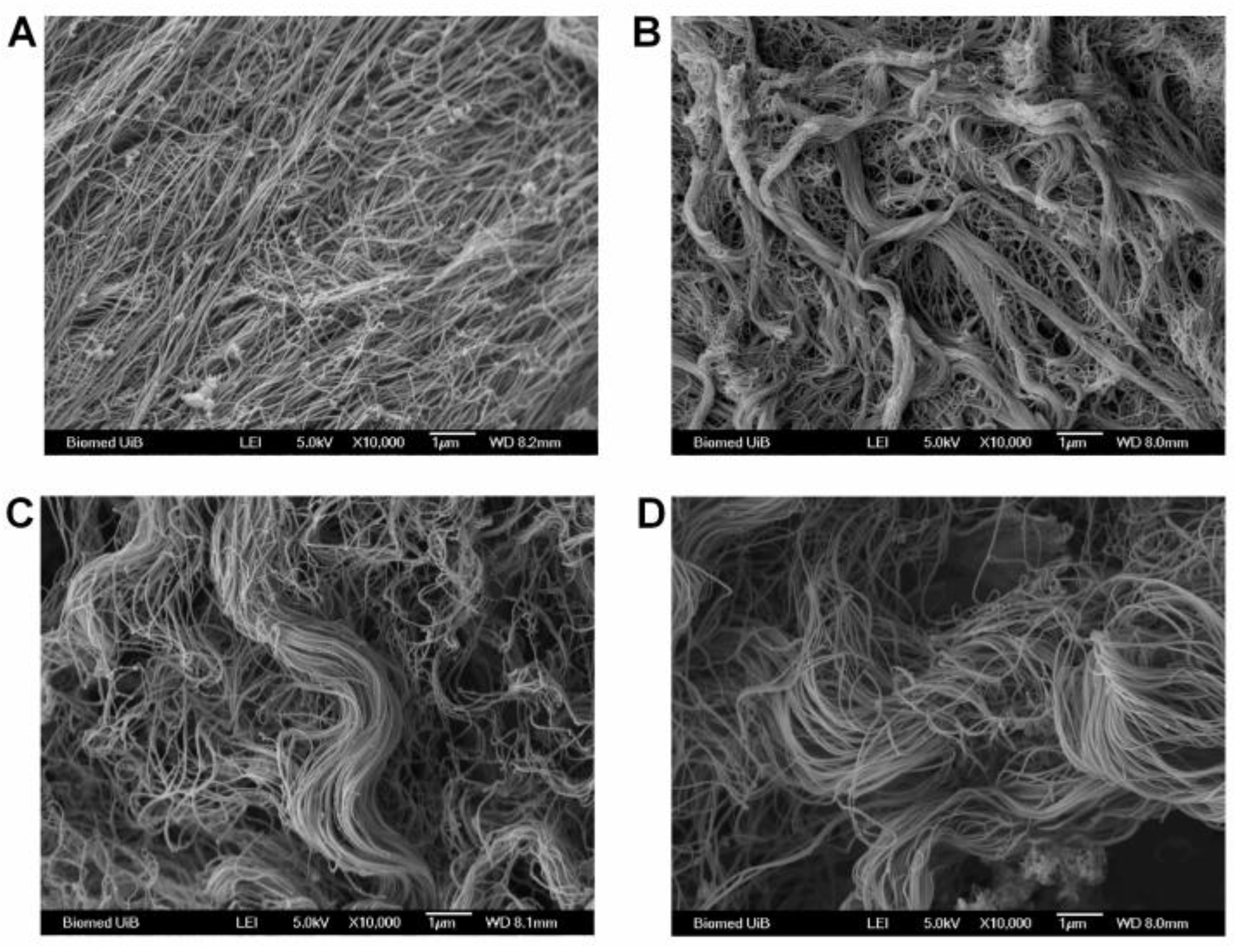
2.5. Electron Microscopy of Collagen Fibrils in the Tumor
2.6. Immunohistochemistry and Immunofluorescence
2.7. Metastasis
2.8. Statistical Methods
3. Results
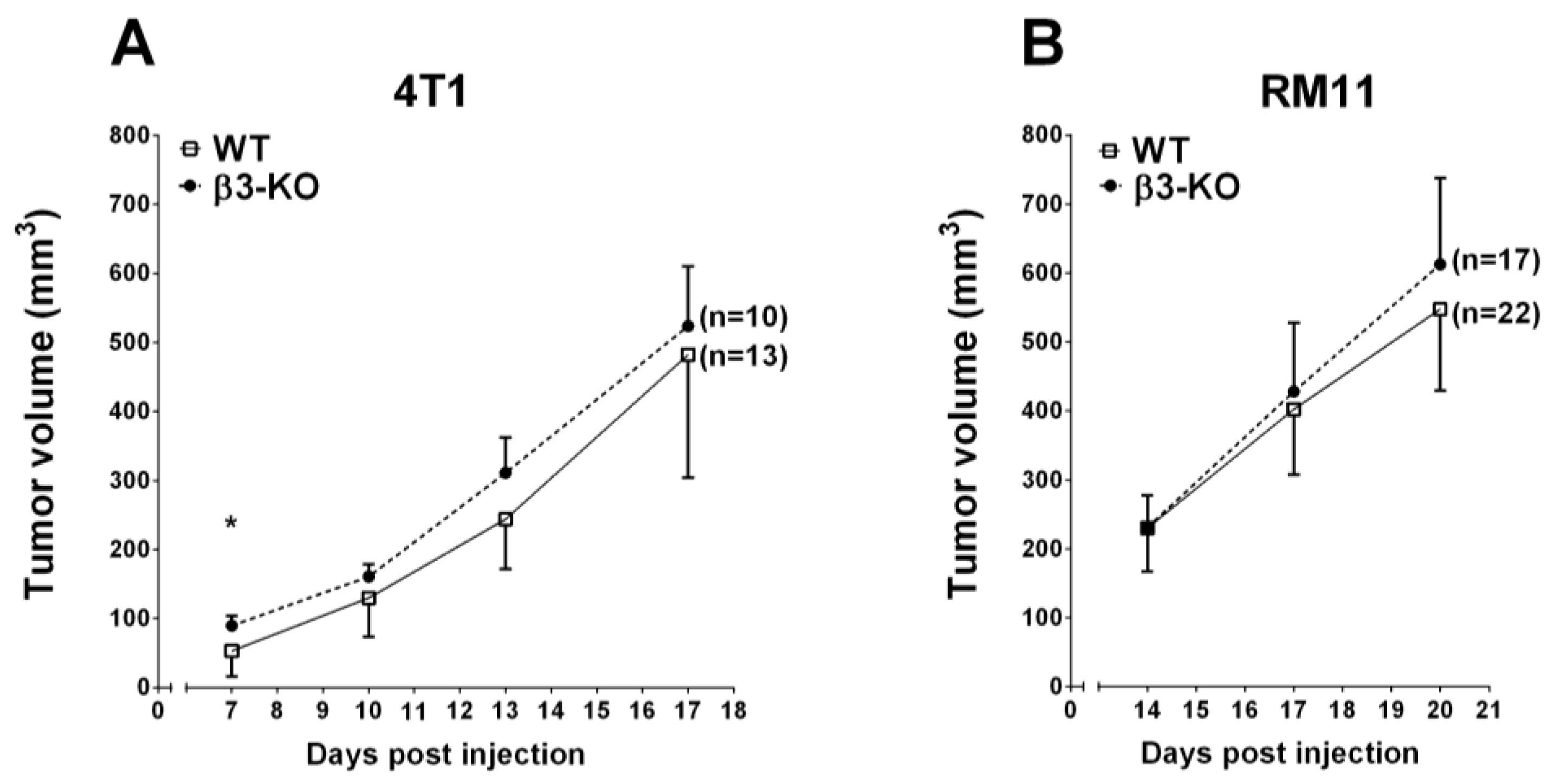
3.1. Stromal Integrin β3-Deficiency and Tumor Growth

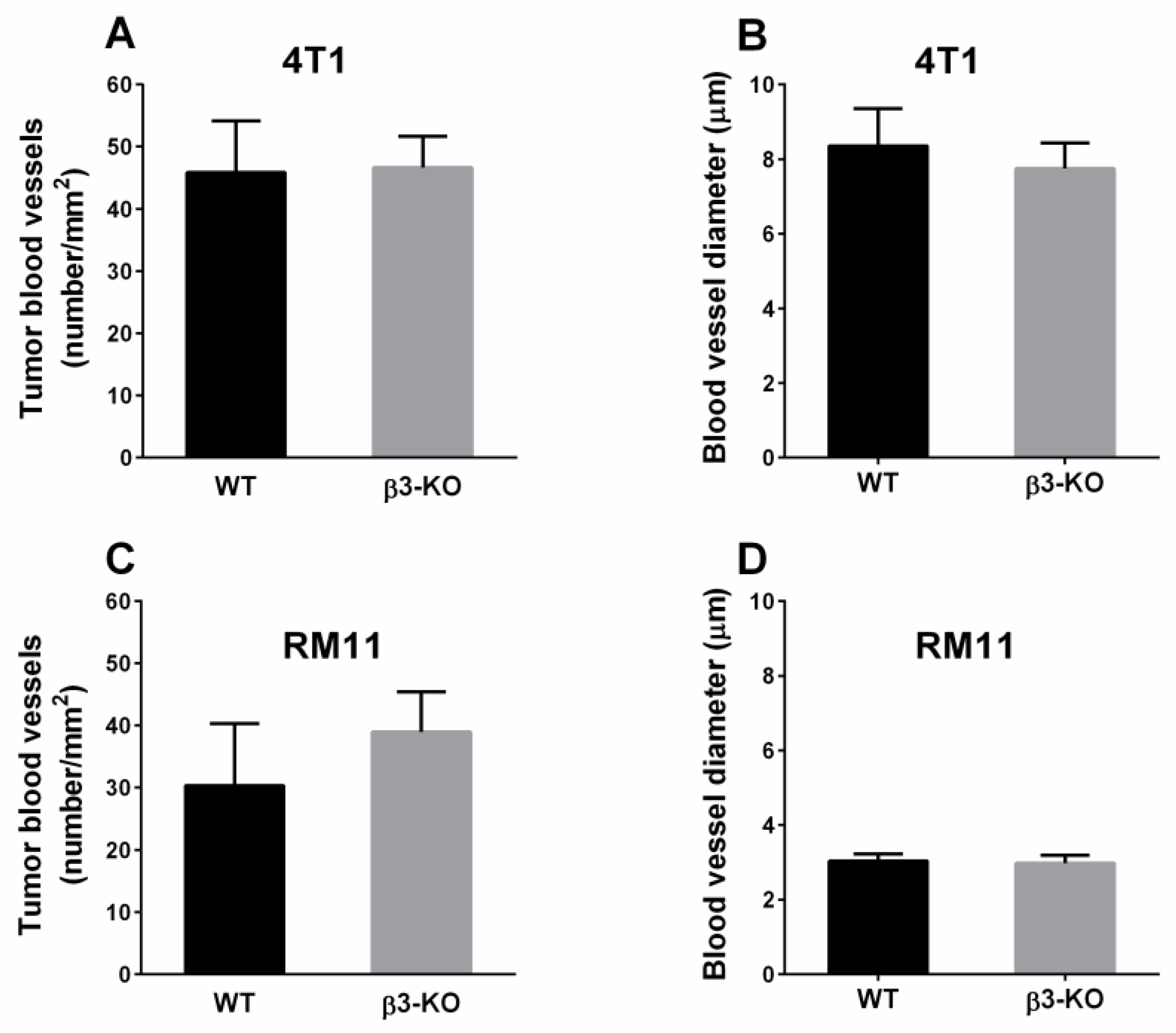
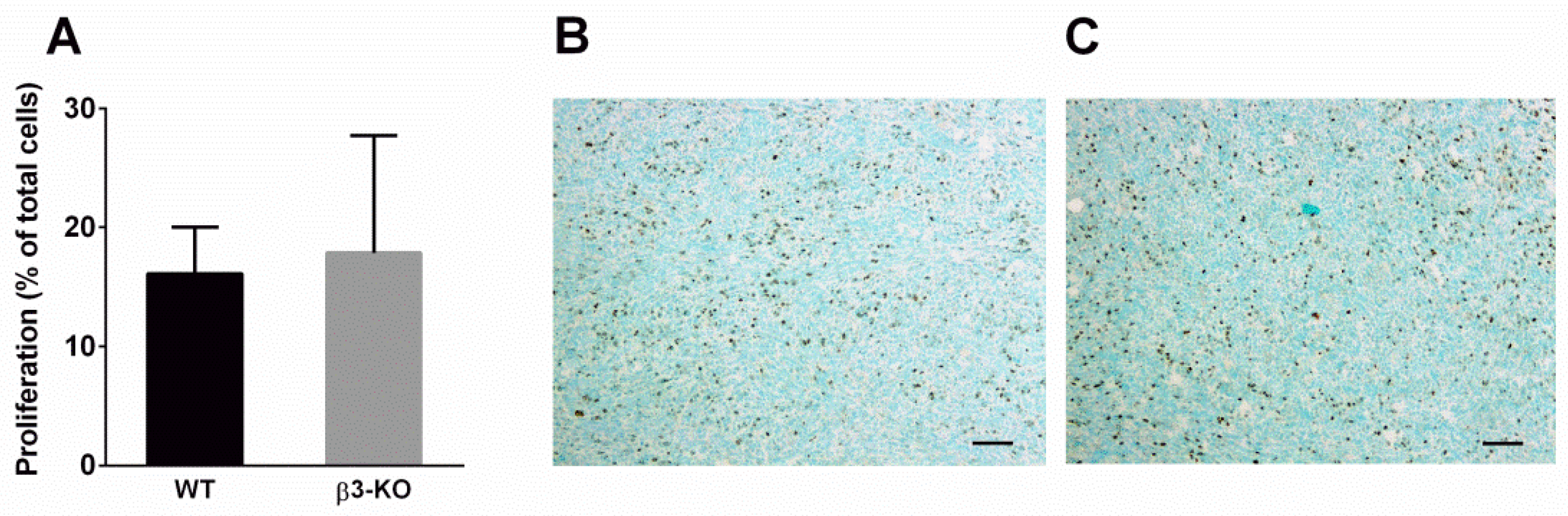
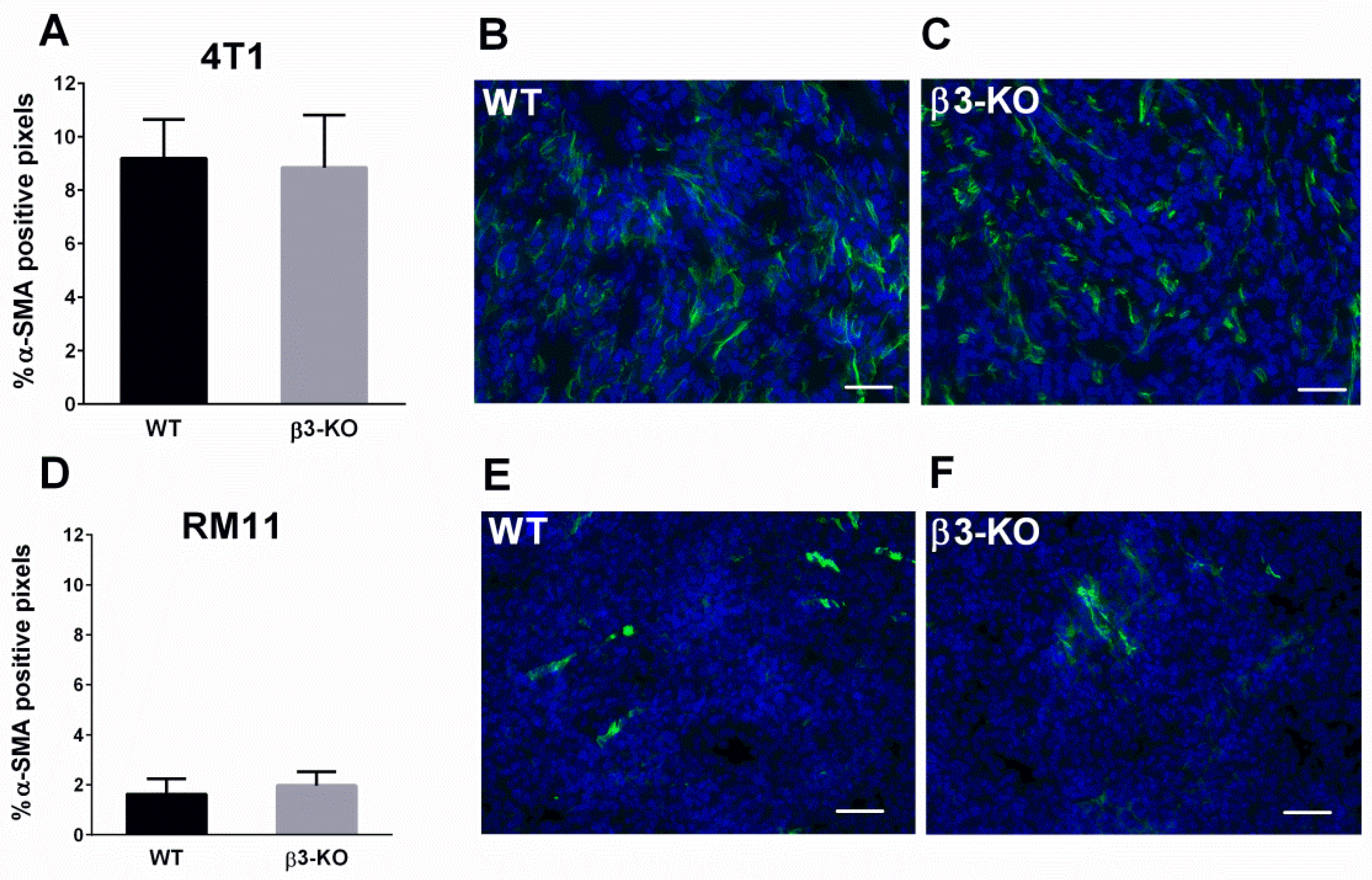
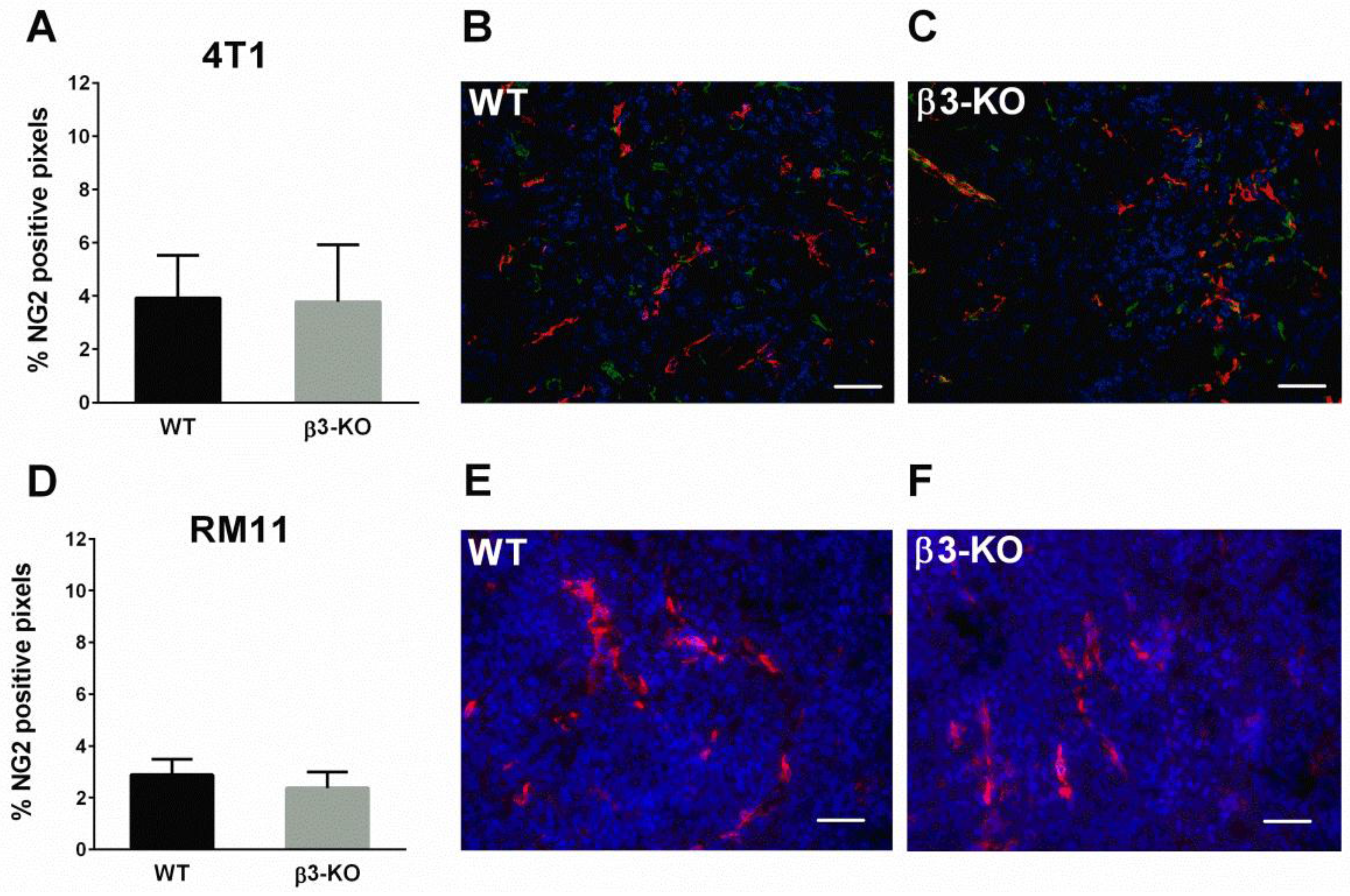
3.2. Integrin β3-Deficiency in Stromal Cells Has No Effect on Blood Vessels, α-SMA or Cell Proliferation




3.3. Integrin β3-Deficiency in Stromal Cells Elevates Interstitial Fluid Pressure (PIF) Only in 4T1 Carcinomas

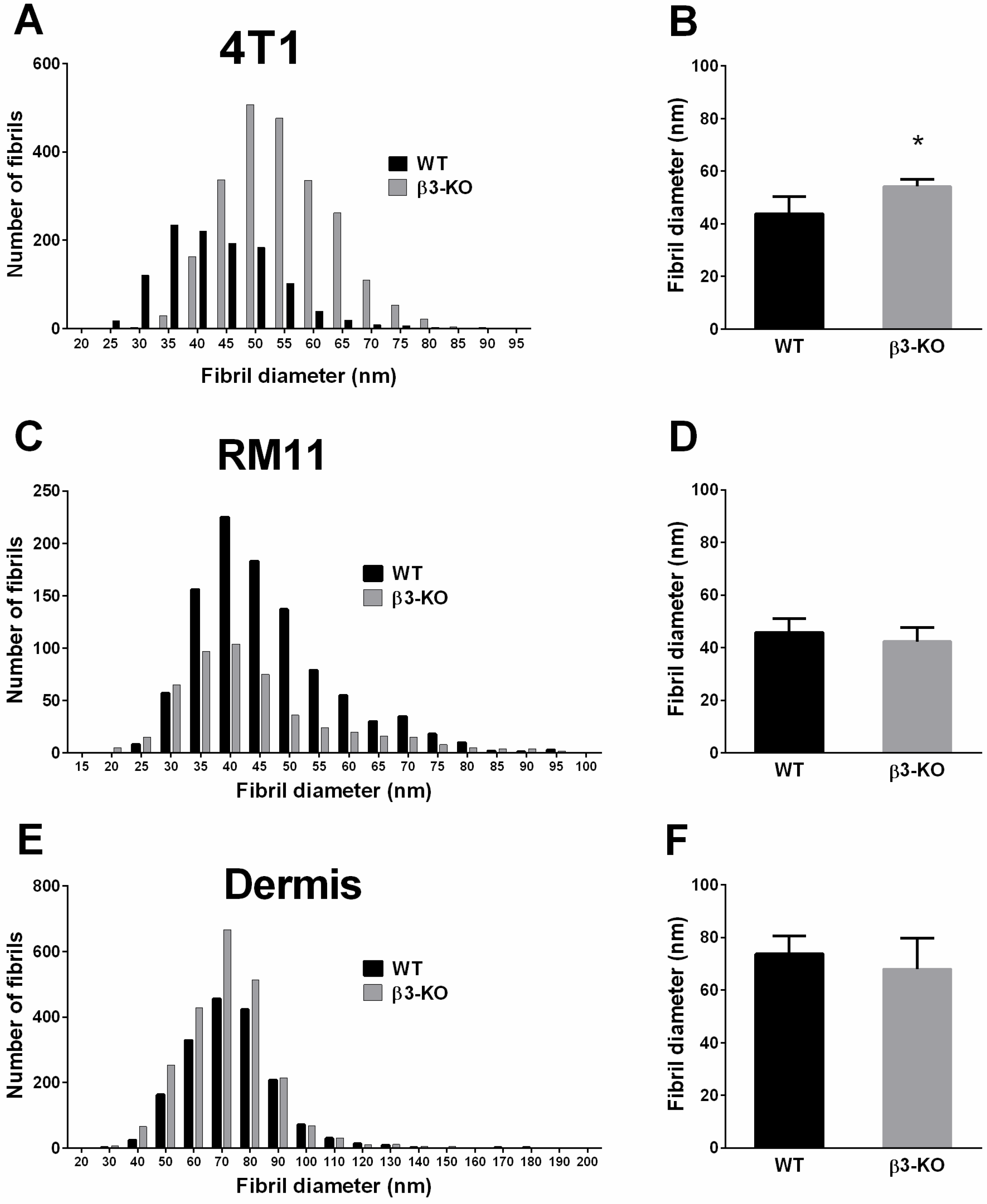
3.4. Integrin β3-Deficiency in Stromal Cells Changes Collagen Architecture in 4T1 Carcinomas


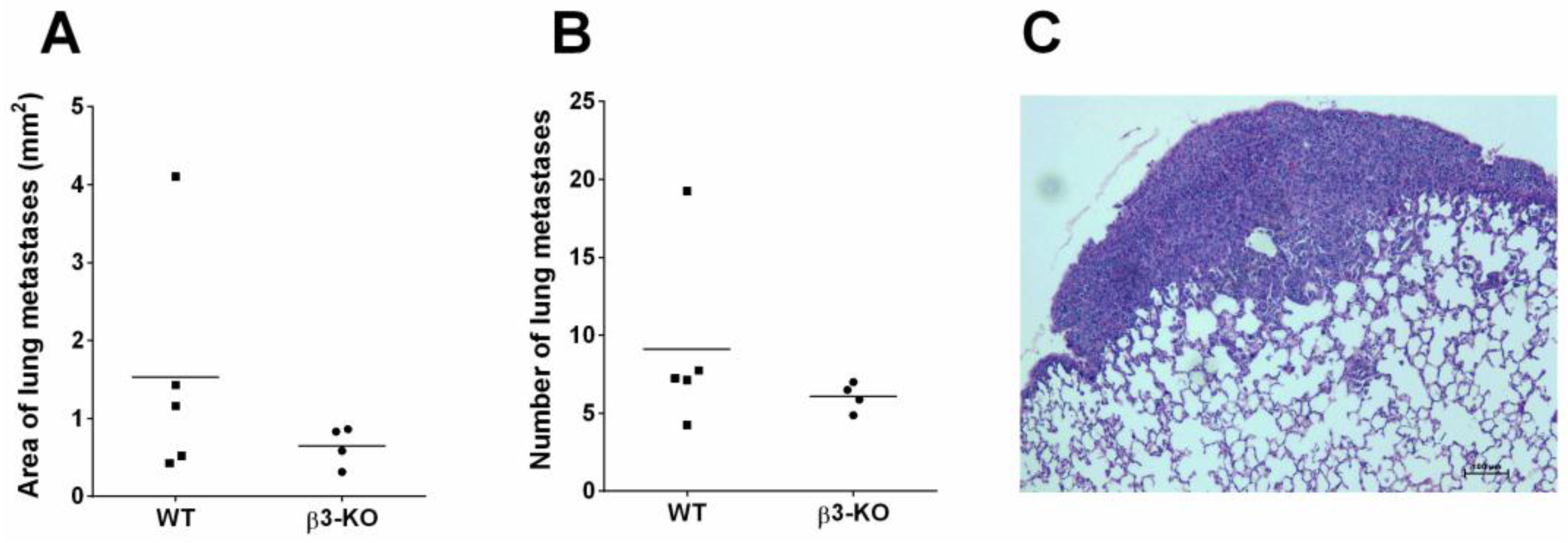
3.5. Integrin β3-Deficiency in Stromal Cells Does not Influence Metastasis

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Kharaishvili, G.; Simkova, D.; Bouchalova, K.; Gachechiladze, M.; Narsia, N.; Bouchal, J. The role of cancer-associated fibroblasts, solid stress and other microenvironmental factors in tumor progression and therapy resistance. Cancer Cell Int. 2014, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H.; Rubin, K.; Pietras, K.; Ostman, A. High interstitial fluid pressure—An obstacle in cancer therapy. Nat. Rev. Cancer 2004, 4, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Le Breton, A.; Preat, V. Rgd-based strategies to target alpha(v) beta(3) integrin in cancer therapy and diagnosis. Mol. Pharm. 2012, 9, 2961–2973. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.C.; Qu, X.J.; Gao, Z.H. Integrins: Players in cancer progression and targets in cancer therapy. Anti-Cancer Drugs 2014, 25, 1107–1121. [Google Scholar] [CrossRef] [PubMed]
- Switala-Jelen, K.; Dabrowska, K.; Opolski, A.; Lipinska, L.; Nowaczyk, M.; Gorski, A. The biological functions of beta3 integrins. Folia Biol. 2004, 50, 143–152. [Google Scholar]
- Hodivala-Dilke, K.M.; McHugh, K.P.; Tsakiris, D.A.; Rayburn, H.; Crowley, D.; Ullman-Cullere, M.; Ross, F.P.; Coller, B.S.; Teitelbaum, S.; Hynes, R.O. Beta3-integrin-deficient mice are a model for glanzmann thrombasthenia showing placental defects and reduced survival. J. Clin. Investig. 1999, 103, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, F.; Chen, X. Integrin alpha(v)beta(3)-targeted cancer therapy. Drug Dev. Res. 2008, 69, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Somanath, P.R.; Malinin, N.L.; Byzova, T.V. Cooperation between integrin alphavbeta3 and vegfr2 in angiogenesis. Angiogenesis 2009, 12, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Borges, E.; Jan, Y.; Ruoslahti, E. Platelet-derived growth factor receptor beta and vascular endothelial growth factor receptor 2 bind to the beta 3 integrin through its extracellular domain. J. Biol. Chem. 2000, 275, 39867–39873. [Google Scholar] [CrossRef] [PubMed]
- Felding-Habermann, B.; O’Toole, T.E.; Smith, J.W.; Fransvea, E.; Ruggeri, Z.M.; Ginsberg, M.H.; Hughes, P.E.; Pampori, N.; Shattil, S.J.; Saven, A.; et al. Integrin activation controls metastasis in human breast cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 1853–1858. [Google Scholar] [CrossRef] [PubMed]
- Sheldrake, H.M.; Patterson, L.H. Function and antagonism of beta3 integrins in the development of cancer therapy. Curr. Cancer Drug Targets 2009, 9, 519–540. [Google Scholar] [CrossRef] [PubMed]
- Trikha, M.; Zhou, Z.; Timar, J.; Raso, E.; Kennel, M.; Emmell, E.; Nakada, M.T. Multiple roles for platelet gpiib/iiia and alphavbeta3 integrins in tumor growth, angiogenesis, and metastasis. Cancer Res. 2002, 62, 2824–2833. [Google Scholar] [PubMed]
- Reinmuth, N.; Liu, W.; Ahmad, S.A.; Fan, F.; Stoeltzing, O.; Parikh, A.A.; Bucana, C.D.; Gallick, G.E.; Nickols, M.A.; Westlin, W.F.; et al. Alphavbeta3 integrin antagonist s247 decreases colon cancer metastasis and angiogenesis and improves survival in mice. Cancer Res. 2003, 63, 2079–2087. [Google Scholar] [PubMed]
- Harms, J.F.; Welch, D.R.; Samant, R.S.; Shevde, L.A.; Miele, M.E.; Babu, G.R.; Goldberg, S.F.; Gilman, V.R.; Sosnowski, D.M.; Campo, D.A.; et al. A small molecule antagonist of the alpha(v)beta3 integrin suppresses mda-mb-435 skeletal metastasis. Clin. Exp. Metastasis 2004, 21, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.E.; Keene, J.L.; Settle, S.L.; Duffin, T.D.; Nickols, M.A.; Westlin, M.; Schroeter, S.; Ruminski, P.G.; Griggs, D.W. Anti-metastatic properties of RGD-peptidomimetic agents S137 and S247. Clin. Exp. Metastasis 2004, 21, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Pickarski, M.; Gleason, A.; Bednar, B.; Duong le, T. Orally active alphavbeta3 integrin inhibitor mk0429 reduces melanoma metastasis. Oncol. Rep. 2015, 33, 2737–2745. [Google Scholar] [PubMed]
- Stupp, R.; Hegi, M.E.; Gorlia, T.; Erridge, S.C.; Perry, J.; Hong, Y.K.; Aldape, K.D.; Lhermitte, B.; Pietsch, T.; Grujicic, D.; et al. Cilengitide combined with standard treatment for patients with newly diagnosed glioblastoma with methylated mgmt promoter (centric eortc 26071–22072 study): A multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2014, 15, 1100–1108. [Google Scholar] [CrossRef]
- Brooks, P.C.; Montgomery, A.M.; Rosenfeld, M.; Reisfeld, R.A.; Hu, T.; Klier, G.; Cheresh, D.A. Integrin alpha v beta 3 antagonists promote tumor regression by inducing apoptosis of angiogenic blood vessels. Cell 1994, 79, 1157–1164. [Google Scholar] [CrossRef]
- Brooks, P.C.; Stromblad, S.; Klemke, R.; Visscher, D.; Sarkar, F.H.; Cheresh, D.A. Antiintegrin alpha v beta 3 blocks human breast cancer growth and angiogenesis in human skin. J. Clin. Investig. 1995, 96, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Bu, X.Y.; Khankaldyyan, V.; Gonzales-Gomez, I.; McComb, J.G.; Laug, W.E. Effect of the angiogenesis inhibitor cilengitide (emd 121974) on glioblastoma growth in nude mice. Neurosurgery 2006, 59, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, T.J.; Taga, T.; Shimada, H.; Tabrizi, P.; Zlokovic, B.V.; Cheresh, D.A.; Laug, W.E. Preferential susceptibility of brain tumors to the antiangiogenic effects of an alpha(v) integrin antagonist. Neurosurgery 2001, 48, 151–157. [Google Scholar] [PubMed]
- Reardon, D.A.; Fink, K.L.; Mikkelsen, T.; Cloughesy, T.F.; O’Neill, A.; Plotkin, S.; Glantz, M.; Ravin, P.; Raizer, J.J.; Rich, K.M.; et al. Randomized phase II study of cilengitide, an integrin-targeting arginine-glycine-aspartic acid peptide, in recurrent glioblastoma multiforme. J. Clin. Oncol. 2008, 26, 5610–5617. [Google Scholar] [CrossRef] [PubMed]
- Manegold, C.; Vansteenkiste, J.; Cardenal, F.; Schuette, W.; Woll, P.J.; Ulsperger, E.; Kerber, A.; Eckmayr, J.; von Pawel, J. Randomized phase ii study of three doses of the integrin inhibitor cilengitide versus docetaxel as second-line treatment for patients with advanced non-small-cell lung cancer. Investig. New Drugs 2013, 31, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.B.; Prieto, V.; Joseph, R.W.; Diwan, A.H.; Gallick, G.E.; Papadopoulos, N.E.; Bedikian, A.Y.; Camacho, L.H.; Hwu, P.; Ng, C.S.; et al. A randomized phase ii study of cilengitide (emd 121974) in patients with metastatic melanoma. Melanoma Res. 2012, 22, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Hersey, P.; Sosman, J.; O'Day, S.; Richards, J.; Bedikian, A.; Gonzalez, R.; Sharfman, W.; Weber, R.; Logan, T.; Buzoianu, M.; et al. A randomized phase 2 study of etaracizumab, a monoclonal antibody against integrin alpha(v)beta(3), + or − dacarbazine in patients with stage iv metastatic melanoma. Cancer 2010, 116, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Friess, H.; Langrehr, J.M.; Oettle, H.; Raedle, J.; Niedergethmann, M.; Dittrich, C.; Hossfeld, D.K.; Stoger, H.; Neyns, B.; Herzog, P.; et al. A randomized multi-center phase ii trial of the angiogenesis inhibitor cilengitide (emd 121974) and gemcitabine compared with gemcitabine alone in advanced unresectable pancreatic cancer. BMC Cancer 2006, 6, 285. [Google Scholar] [CrossRef] [PubMed]
- Alva, A.; Slovin, S.; Daignault, S.; Carducci, M.; Dipaola, R.; Pienta, K.; Agus, D.; Cooney, K.; Chen, A.; Smith, D.C.; et al. Phase ii study of cilengitide (emd 121974, nsc 707544) in patients with non-metastatic castration resistant prostate cancer, nci-6735. A study by the dod/pcf prostate cancer clinical trials consortium. Investig. New Drugs 2012, 30, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Wiig, H.; Reed, R.K.; Aukland, K. Measurement of interstitial fluid pressure: Comparison of methods. Ann. Biomed. Eng. 1986, 14, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Friman, T.; Gustafsson, R.; Stuhr, L.B.; Chidiac, J.; Heldin, N.E.; Reed, R.K.; Oldberg, A.; Rubin, K. Increased fibrosis and interstitial fluid pressure in two different types of syngeneic murine carcinoma grown in integrin beta3-subunit deficient mice. PLoS ONE 2012, 7, e34082. [Google Scholar] [CrossRef] [PubMed]
- Oldberg, A.; Kalamajski, S.; Salnikov, A.V.; Stuhr, L.; Morgelin, M.; Reed, R.K.; Heldin, N.E.; Rubin, K. Collagen-binding proteoglycan fibromodulin can determine stroma matrix structure and fluid balance in experimental carcinoma. Proc. Natl. Acad. Sci. USA 2007, 104, 13966–13971. [Google Scholar] [CrossRef] [PubMed]
- Heppner, G.H.; Miller, F.R.; Shekhar, P.M. Nontransgenic models of breast cancer. Breast Cancer Res. 2000, 2, 331–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, R.Z.; Micocci, K.C.; Natoli, A.; Redvers, R.P.; Paquet-Fifield, S.; Martin, A.C.; Denoyer, D.; Ling, X.; Kim, S.H.; Tomasin, R.; et al. Tumour but not stromal expression of beta3 integrin is essential, and is required early, for spontaneous dissemination of bone-metastatic breast cancer. J. Pathol. 2014, 235, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.E.; Wyder, L.; Lively, J.C.; Taverna, D.; Robinson, S.D.; Huang, X.; Sheppard, D.; Hynes, R.O.; Hodivala-Dilke, K.M. Enhanced pathological angiogenesis in mice lacking beta3 integrin or beta3 and beta5 integrins. Nat. Med. 2002, 8, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Taverna, D.; Moher, H.; Crowley, D.; Borsig, L.; Varki, A.; Hynes, R.O. Increased primary tumor growth in mice null for beta3- or beta3/beta5-integrins or selectins. Proc. Natl. Acad. Sci. USA 2004, 101, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Chen, X. Anti-angiogenic cancer therapy based on integrin alphavbeta3 antagonism. Anticancer Agents Med. Chem. 2006, 6, 407–428. [Google Scholar] [CrossRef] [PubMed]
- Salnikov, A.V.; Iversen, V.V.; Koisti, M.; Sundberg, C.; Johansson, L.; Stuhr, L.B.; Sjoquist, M.; Ahlstrom, H.; Reed, R.K.; Rubin, K. Lowering of tumor interstitial fluid pressure specifically augments efficacy of chemotherapy. FASEB J. 2003, 17, 1756–1758. [Google Scholar] [CrossRef] [PubMed]
- Pietras, K.; Rubin, K.; Sjoblom, T.; Buchdunger, E.; Sjoquist, M.; Heldin, C.H.; Ostman, A. Inhibition of pdgf receptor signaling in tumor stroma enhances antitumor effect of chemotherapy. Cancer Res. 2002, 62, 5476–5484. [Google Scholar] [PubMed]
- Pietras, K.; Stumm, M.; Hubert, M.; Buchdunger, E.; Rubin, K.; Heldin, C.H.; McSheehy, P.; Wartmann, M.; Ostman, A. Sti571 enhances the therapeutic index of epothilone b by a tumor-selective increase of drug uptake. Clin. Cancer Res. 2003, 9, 3779–3787. [Google Scholar] [PubMed]
- Diop-Frimpong, B.; Chauhan, V.P.; Krane, S.; Boucher, Y.; Jain, R.K. Losartan inhibits collagen i synthesis and improves the distribution and efficacy of nanotherapeutics in tumors. Proc. Natl. Acad. Sci. USA 2011, 108, 2909–2914. [Google Scholar] [CrossRef] [PubMed]
- Olive, K.P.; Jacobetz, M.A.; Davidson, C.J.; Gopinathan, A.; McIntyre, D.; Honess, D.; Madhu, B.; Goldgraben, M.A.; Caldwell, M.E.; Allard, D.; et al. Inhibition of hedgehog signaling enhances delivery of chemotherapy in a mouse model of pancreatic cancer. Science 2009, 324, 1457–1461. [Google Scholar] [CrossRef] [PubMed]
- Levick, J.R. Flow through interstitium and other fibrous matrices. Quant. J. Exp. Physiol. 1987, 72, 409–437. [Google Scholar] [CrossRef]
- Taverna, D.; Crowley, D.; Connolly, M.; Bronson, R.T.; Hynes, R.O. A direct test of potential roles for beta3 and beta5 integrins in growth and metastasis of murine mammary carcinomas. Cancer Res. 2005, 65, 10324–10329. [Google Scholar] [CrossRef] [PubMed]
- Bakewell, S.J.; Nestor, P.; Prasad, S.; Tomasson, M.H.; Dowland, N.; Mehrotra, M.; Scarborough, R.; Kanter, J.; Abe, K.; Phillips, D.; et al. Platelet and osteoclast beta3 integrins are critical for bone metastasis. Proc. Natl. Acad. Sci. USA 2003, 100, 14205–14210. [Google Scholar] [CrossRef] [PubMed]
- Gay, L.J.; Felding-Habermann, B. Contribution of platelets to tumour metastasis. Nat. Rev. Cancer 2011, 11, 123–134. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reigstad, I.; Sortland, K.; Skogstrand, T.; Reed, R.K.; Stuhr, L. The Effect of Stromal Integrin β3-Deficiency on Two Different Tumors in Mice. Cancers 2016, 8, 14. https://doi.org/10.3390/cancers8010014
Reigstad I, Sortland K, Skogstrand T, Reed RK, Stuhr L. The Effect of Stromal Integrin β3-Deficiency on Two Different Tumors in Mice. Cancers. 2016; 8(1):14. https://doi.org/10.3390/cancers8010014
Chicago/Turabian StyleReigstad, Inga, Kristina Sortland, Trude Skogstrand, Rolf K. Reed, and Linda Stuhr. 2016. "The Effect of Stromal Integrin β3-Deficiency on Two Different Tumors in Mice" Cancers 8, no. 1: 14. https://doi.org/10.3390/cancers8010014
APA StyleReigstad, I., Sortland, K., Skogstrand, T., Reed, R. K., & Stuhr, L. (2016). The Effect of Stromal Integrin β3-Deficiency on Two Different Tumors in Mice. Cancers, 8(1), 14. https://doi.org/10.3390/cancers8010014