
Epidermal Growth Factor Pathway Signaling in Drosophila Embryogenesis: Tools for Understanding Cancer


{kind=link}
{kind=link}
Abstract
:1. Introduction to EGF Signaling
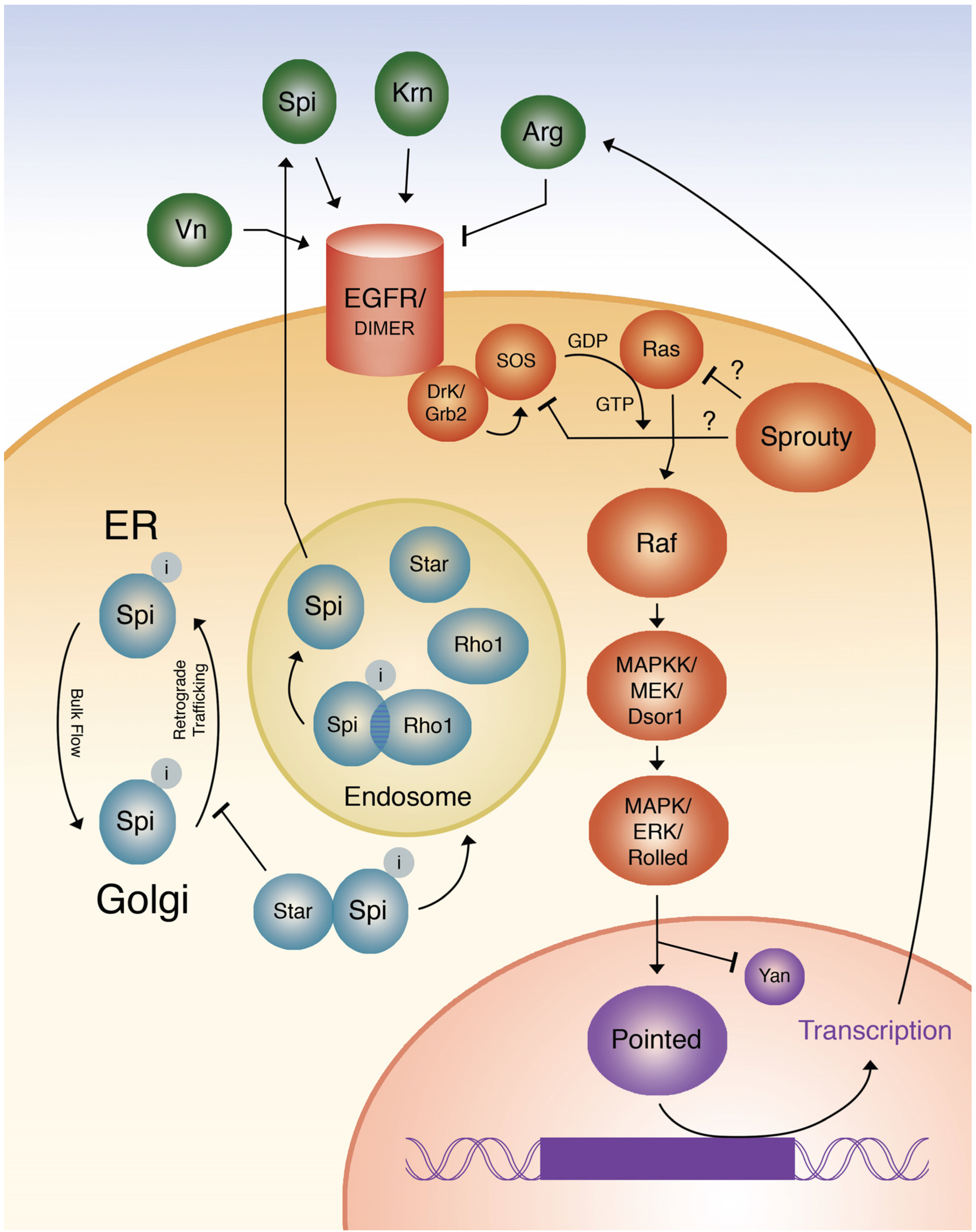
1.1. Function of the EGF Receptor, Its Ligands, and an Antagonist
1.2. Intracellular Regulation of EGF Ligands
1.3. Transduction of the EGF Signal from the Receptor to ras
1.4. Mechanism of RAS Activation of RAF and the Kinase Cascade
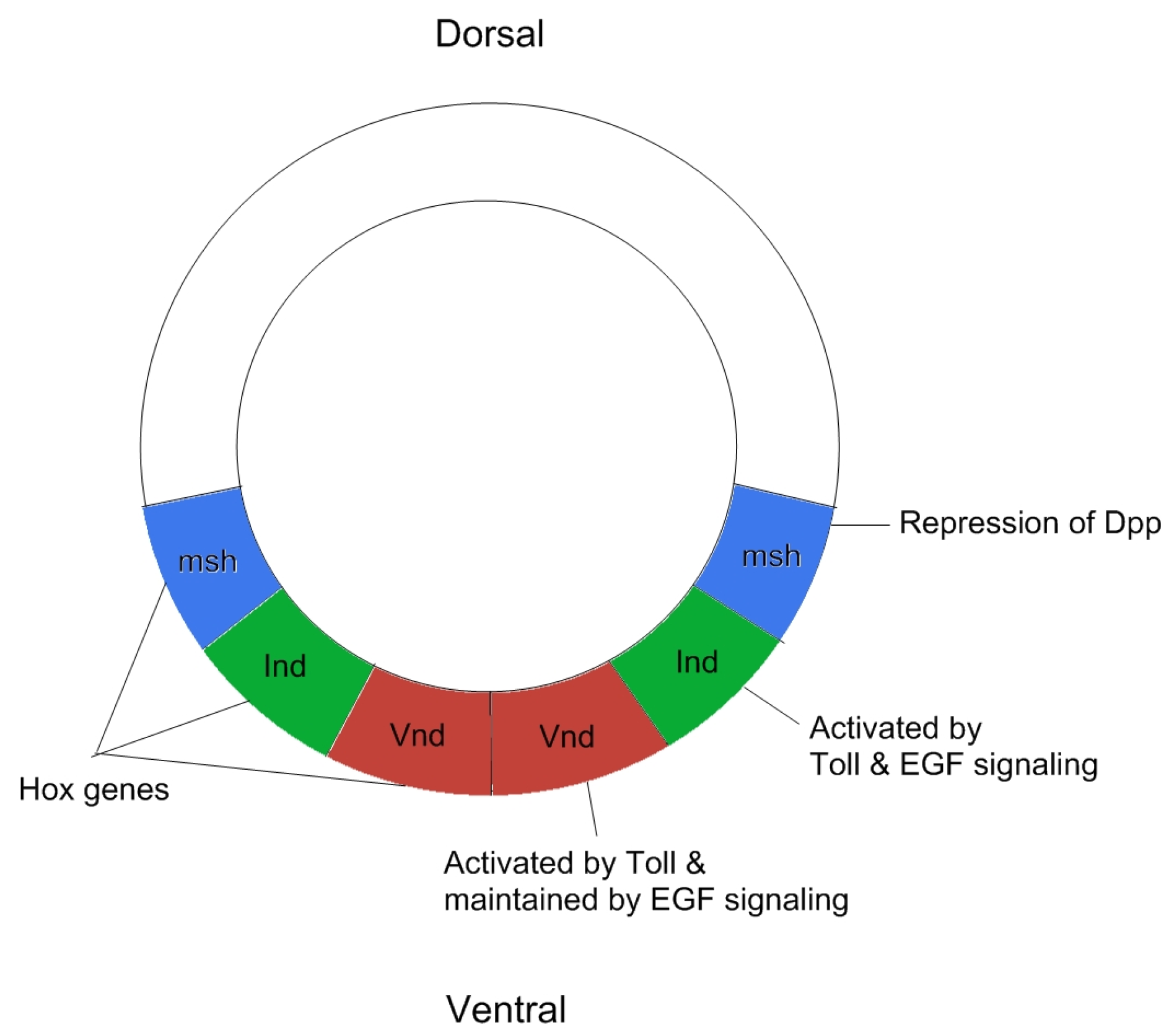
2. Patterning of the Neuroectoderm
3. Non-Neuroectodermal Ventral Patterning, Including Segmentation
4. Specification of Muscle Precursors and Muscle Attachment to Tendons
5. Specification of Tracheal Invagination
6. Cell Recruitment to the Chordotonal Organs
7. Specification of Oenocytes
8. Dorsal Midline Patterning (Including Dorsal Closure and Viability of Dorsal Midline Cells)
9. Other Signaling Pathways
10. Conclusions
10.1. Common Themes
- (1)
- EGF often functions in cell growth and proliferation and regulates apoptosis.
- (2)
- EGF signaling in Drosophila is highly self-regulating, particularly through RHO-1 and STAR mediated processing of EGF ligands and post-transcriptional control of ARGOS expression.
- (3)
- EGF signaling also can differ qualitatively due to expression of different yet functionally similar ligands (i.e., vein vs. spi). The availability of ligands with slightly different functionality allows EGF signaling to be precisely regulated based on ligand availability, rather than attempting to modulate the effects of the pathway after activation by a single ligand.
- (4)
- EGF signaling functions in cell adhesion and polarity. These four characteristics make EGF signaling highly effective at modulating tissue boundaries involving multiple alternative signaling pathways, as EGF signaling can self-regulate expression and interact with many other pathways through both negative and positive regulation.
10.2. Utility of Studying the EGF Signaling Pathway
Acknowledgments
Conflicts of Interest
References
- Price, J.V.; Clifford, R.J.; Schupbach, T. The maternal ventralizing locus torpedo is allelic to faint little ball, an embryonic lethal, and encodes the Drosophila EGFR receptor homolog. Cell 1989, 56, 1085–1092. [Google Scholar] [CrossRef]
- Baker, N.E.; Rubin, G.M. Ellipse mutations in the Drosophila homologue of the EGFR receptor affect pattern formation, cell division, and cell death in eye imaginal discs. Dev. Biol. 1992, 150, 381–396. [Google Scholar] [CrossRef]
- Schejter, E.D.; Shilo, B.Z. The Drosophila EGFR receptor homolog (DER) gene is allelic to faint little ball, a locus essential for embryonic development. Cell 1989, 56, 1093–1104. [Google Scholar] [CrossRef]
- Clifford, R.; Schupbach, T. Molecular analysis of the Drosophila EGFR receptor homolog reveals that several genetically defined classes of alleles cluster in subdomains of the receptor protein. Genetics 1994, 137, 531–550. [Google Scholar] [PubMed]
- Clifford, R.J.; Schupbach, T. Coordinately and differentially mutable activities of torpedo, the Drosophila melanogaster homolog of the vertebrate EGFR receptor gene. Genetics 1989, 123, 771–787. [Google Scholar] [PubMed]
- Bogdan, S.; Klambt, C. Epidermal growth factor receptor signaling. Curr. Biol. 2001, 11, R292–R295. [Google Scholar] [CrossRef]
- Attrill, H.; Falls, K.; Goodman, J.L.; Millburn, G.H.; Antonazzo, G.; Rey, A.J.; Marygold, S.J.; FlyBase, C. Flybase: Establishing a gene group resource for Drosophila melanogaster. Nucleic Acids Res. 2016, 44, D786–D792. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. Uniprot: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar]
- Klein, D.E.; Stayrook, S.E.; Shi, F.; Narayan, K.; Lemmon, M.A. Structural basis for EGFR ligand sequestration by argos. Nature 2008, 453, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.L.; Manivannan, S.N.; Simcox, A. TGF-alpha ligands can substitute for the neuregulin Vein in Drosophila development. Development 2014, 141, 4110–4114. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, J.; Liu, H.H.; Miller, E.; Treisman, J.E. Trafficking of the EGFR ligand Spitz regulates its signaling activity in polarized tissues. J. Cell Sci. 2013, 126, 4469–4478. [Google Scholar] [CrossRef] [PubMed]
- Paul, L.; Wang, S.H.; Manivannan, S.N.; Bonanno, L.; Lewis, S.; Austin, C.L.; Simcox, A. Dpp-induced EGFR signaling triggers postembryonic wing development in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 5058–5063. [Google Scholar] [CrossRef] [PubMed]
- Neuman-Silberberg, F.S.; Schüpbach, T. The Drosophila dorsoventral patterning gene gurken produces a dorsally localized rna and encodes a TGFα-like protein. Cell 1993, 75, 165–174. [Google Scholar] [CrossRef]
- Malartre, M. Regulatory mechanisms of EGFR signalling during Drosophila eye development. Cell. Mol. Life Sci. 2016, 73, 1825–1843. [Google Scholar] [CrossRef] [PubMed]
- Reich, A.; Shilo, B.Z. Keren, a new ligand of the Drosophila epidermal growth factor receptor, undergoes two modes of cleavage. EMBO J. 2002, 21, 4287–4296. [Google Scholar] [CrossRef] [PubMed]
- Urban, S.; Lee, J.R.; Freeman, M. A family of Rhomboid intramembrane proteases activates all Drosophila membrane-tethered EGFR ligands. EMBO J. 2002, 21, 4277–4286. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.; Dsilva, C.J.; Levario, T.J.; Lu, H.; Schupbach, T.; Kevrekidis, I.G.; Shvartsman, S.Y. Dynamics of inductive erk signaling in the Drosophila embryo. Curr. Biol. 2015, 25, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- Schnepp, B.; Grumbling, G.; Donaldson, T.; Simcox, A. Vein is a novel component in the Drosophila epidermal growth factor receptor pathway with similarity to the neuregulins. Genes Dev. 1996, 10, 2302–2313. [Google Scholar] [CrossRef] [PubMed]
- Gabay, L.; Scholz, H.; Golembo, M.; Klaes, A.; Shilo, B.Z.; Klambt, C. EGF receptor signaling induces pointed p1 transcription and inactivates yan protein in the Drosophila embryonic ventral ectoderm. Development 1996, 122, 3355–3362. [Google Scholar] [PubMed]
- Shilo, B.Z. The regulation and functions of MAPK pathways in Drosophila. Methods 2014, 68, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Urban, S.; Lee, J.R.; Freeman, M. Drosophila Rhomboid-1 defines a family of putative intramembrane serine proteases. Cell 2001, 107, 173–182. [Google Scholar] [CrossRef]
- Barberán, S.; Martín-Durán, J.M.; Cebrià, F. Evolution of the EGFR pathway in metazoa and its diversification in the planarian schmidtea mediterranea. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.; Urban, S.; Garvey, C.F.; Freeman, M. Regulated intracellular ligand transport and proteolysis control EGFR signal activation in Drosophila. Cell 2001, 107, 161–171. [Google Scholar] [CrossRef]
- Tsruya, R.; Schlesinger, A.; Reich, A.; Gabay, L.; Sapir, A.; Shilo, B.Z. Intracellular trafficking by star regulates cleavage of the Drosophila EGFR receptor ligand Spitz. Genes Dev. 2002, 16, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Shilo, B.Z. Developmental roles of Rhomboid proteases. Semin. Cell Dev. Biol. 2016, 60, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Kolodkin, A.L.; Pickup, A.T.; Lin, D.M.; Goodman, C.S.; Banerjee, U. Characterization of star and its interactions with sevenless and EGFR receptor during photoreceptor cell development in Drosophila. Development 1994, 120, 1731–1745. [Google Scholar] [PubMed]
- Tsruya, R.; Wojtalla, A.; Carmon, S.; Yogev, S.; Reich, A.; Bibi, E.; Merdes, G.; Schejter, E.; Shilo, B.Z. Rhomboid cleaves star to regulate the levels of secreted Spitz. EMBO J. 2007, 26, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Bang, A.G.; Kintner, C. Rhomboid and star facilitate presentation and processing of the Drosophila TGF-alpha homolog Spitz. Genes Dev. 2000, 14, 177–186. [Google Scholar] [PubMed]
- Bier, E.; Jan, L.Y.; Jan, Y.N. Rhomboid, a gene required for dorsoventral axis establishment and peripheral nervous system development in Drosophila melanogaster. Genes Dev. 1990, 4, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Rebay, I.; Rubin, G.M. Yan functions as a general inhibitor of differentiation and is negatively regulated by activation of the Ras1/MAPK pathway. Cell 1995, 81, 857–866. [Google Scholar] [CrossRef]
- Maixner, A.; Hecker, T.P.; Phan, Q.N.; Wassarman, D.A. A screen for mutations that prevent lethality caused by expression of activated sevenless and Ras1 in the Drosophila embryo. Dev. Genet. 1998, 23, 347–361. [Google Scholar] [CrossRef]
- Egan, S.E.; Giddings, B.W.; Brooks, M.W.; Buday, L.; Sizeland, A.M.; Weinberg, R.A. Association of Sos Ras exchange protein with Grb2 is implicated in tyrosine kinase signal transduction and transformation. Nature 1993, 363, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Kramer, S.; Okabe, M.; Hacohen, N.; Krasnow, M.A.; Hiromi, Y. Sprouty: A common antagonist of FGF and EGFR signaling pathways in Drosophila. Development 1999, 126, 2515–2525. [Google Scholar] [PubMed]
- Casci, T.; Vinos, J.; Freeman, M. Sprouty, an intracellular inhibitor of Ras signaling. Cell 1999, 96, 655–665. [Google Scholar] [CrossRef]
- Marais, R.; Light, Y.; Paterson, H.F.; Marshall, C.J. Ras recruits Raf-1 to the plasma membrane for activation by tyrosine phosphorylation. EMBO J. 1995, 14, 3136–3145. [Google Scholar] [PubMed]
- Stokoe, D.; Macdonald, S.; Cadwallader, K.; Symons, M.; Hancock, J. Activation of Raf as a result of recruitment to the plasma membrane. Science 1994, 264, 1463–1467. [Google Scholar] [CrossRef] [PubMed]
- Radke, K.; Johnson, K.; Guo, R.; Davidson, A.; Ambrosio, L. Drosophila-raf acts to elaborate dorsoventral pattern in the ectoderm of developing embryos. Genetics 2001, 159, 1031–1044. [Google Scholar] [PubMed]
- Tsuda, L.; Inoue, Y.H.; Yoo, M.A.; Mizuno, M.; Hata, M.; Lim, Y.M.; Adachi-Yamada, T.; Ryo, H.; Masamune, Y.; Nishida, Y. A protein kinase similar to MAP kinase activator acts downstream of the raf kinase in Drosophila. Cell 1993, 72, 407–414. [Google Scholar] [CrossRef]
- Gabay, L.; Seger, R.; Shilo, B.Z. In situ activation pattern of Drosophila EGFR receptor pathway during development. Science 1997, 277, 1103–1106. [Google Scholar] [CrossRef] [PubMed]
- Biggs, W.H., 3rd; Zavitz, K.H.; Dickson, B.; van der Straten, A.; Brunner, D.; Hafen, E.; Zipursky, S.L. The Drosophila rolled locus encodes a MAP kinase required in the sevenless signal transduction pathway. EMBO J. 1994, 13, 1628–1635. [Google Scholar] [PubMed]
- Lim, Y.M.; Tsuda, L.; Inoue, Y.H.; Irie, K.; Adachi-Yamada, T.; Hata, M.; Nishi, Y.; Matsumoto, K.; Nishida, Y. Dominant mutations of Drosophila MAP kinase kinase and their activities in Drosophila and yeast MAP kinase cascades. Genetics 1997, 146, 263–273. [Google Scholar] [PubMed]
- Oellers, N.; Hafen, E. Biochemical characterization of rolledsem, an activated form of Drosophila mitogen-activated protein kinase. J. Biol. Chem. 1996, 271, 24939–24944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunner, D.; Oellers, N.; Szabad, J.; Biggs, W.H., 3rd; Zipursky, S.L.; Hafen, E. A gain-of-function mutation in Drosophila MAP kinase activates multiple receptor tyrosine kinase signaling pathways. Cell 1994, 76, 875–888. [Google Scholar] [CrossRef]
- Scholz, H.; Deatrick, J.; Klaes, A.; Klambt, C. Genetic dissection of pointed, a Drosophila gene encoding two ETS-related proteins. Genetics 1993, 135, 455–468. [Google Scholar] [PubMed]
- Samakovlis, C.; Hacohen, N.; Manning, G.; Sutherland, D.C.; Guillemin, K.; Krasnow, M.A. Development of the Drosophila tracheal system occurs by a series of morphologically distinct but genetically coupled branching events. Development 1996, 122, 1395–1407. [Google Scholar] [PubMed]
- Von Ohlen, T.; Doe, C.Q. Convergence of dorsal, Dpp, and EGFR signaling pathways subdivides the Drosophila neuroectoderm into three dorsal-ventral columns. Dev. Biol. 2000, 224, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Skeath, J.B.; Thor, S. Genetic control of Drosophila nerve cord development. Curr. Opin. Neurobiol. 2003, 13, 8–15. [Google Scholar] [CrossRef]
- Skeath, J.B. The Drosophila EGFR receptor controls the formation and specification of neuroblasts along the dorsal-ventral axis of the Drosophila embryo. Development 1998, 125, 3301–3312. [Google Scholar] [PubMed]
- Esteves, P.; Pecqueur, C.; Ransy, C.; Esnous, C.; Lenoir, V.; Bouillaud, F.; Bulteau, A.L.; Lombes, A.; Prip-Buus, C.; Ricquier, D.; et al. Mitochondrial retrograde signaling mediated by UCP2 inhibits cancer cell proliferation and tumorigenesis. Cancer Res. 2014, 74, 3971–3982. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.O.; Kim, I.C.; Kim, S.; Kwon, Y.K.; Han, P.L.; Jeon, S.H.; Kim, S.H. Cns midline cells contribute to maintenance of the initial dorsoventral patterning of the Drosophila ventral neuroectoderm. J. Neurobiol. 2005, 62, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Skeath, J.B. The Sox-domain containing gene Dichaete/fish-hook acts in concert with vnd and ind to regulate cell fate in the Drosophila neuroectoderm. Development 2002, 129, 1165–1174. [Google Scholar] [PubMed]
- Hong, J.W.; Park, K.W.; Levine, M.S. Temporal regulation of single-minded target genes in the ventral midline of the Drosophila central nervous system. Dev. Biol. 2013, 380, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wheeler, S.R.; Skeath, J.B. Genetic control of dorsoventral patterning and neuroblast specification in the Drosophila central nervous system. Int. J. Dev. Biol. 2007, 51, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.T.; Chiba, Y.; Arai, H.; Ishii, M. Discovery of an intermolecular disulfide bond required for the thermostability of a heterodimeric protein from the thermophile hydrogenobacter thermophilus. Biosci. Biotechnol. Biochem. 2016, 80, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jung, K.I.; Kim, S.H.; Jeon, S.H. Dpp represses eagle expression at short-range, but can repress its expression at a long-range via EGFR signal repression. Mol. Cells 2008, 26, 576–582. [Google Scholar] [PubMed]
- Moses, C.; Helman, A.; Paroush, Z.; von Ohlen, T. Phosphorylation of Ind by MAP kinase enhances Ind-dependent transcriptional repression. Dev. Biol. 2011, 360, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Rulifson, E. Serial specification of diverse neuroblast identities from a neurogenic placode by notch and EGFR signaling. Development 2011, 138, 2883–2893. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Shy, D.; Hartenstein, V. Antagonistic relationship between Dpp and EGFR signaling in Drosophila head patterning. Dev. Biol. 2003, 263, 103–113. [Google Scholar] [CrossRef]
- Helman, A.; Lim, B.; Andreu, M.J.; Kim, Y.; Shestkin, T.; Lu, H.; Jimenez, G.; Shvartsman, S.Y.; Paroush, Z. RTK signaling modulates the Dorsal gradient. Development 2012, 139, 3032–3039. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.T.; Verheyen, E.M. Ras-activated Dsor1 promotes wnt signaling in Drosophila development. J. Cell Sci. 2015, 128, 4499–4511. [Google Scholar] [CrossRef] [PubMed]
- Urban, S.; Brown, G.; Freeman, M. EGF receptor signalling protects smooth-cuticle cells from apoptosis during Drosophila ventral epidermis development. Development 2004, 131, 1835–1845. [Google Scholar] [CrossRef] [PubMed]
- Payre, F.; Vincent, A.; Carreno, S. ovo/svb integrates Wingless and DER pathways to control epidermis differentiation. Nature 1999, 400, 271–275. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, L.; Dougan, S.T.; Gabay, L.; Raz, E.; Shilo, B.Z.; DiNardo, S. Spitz and Wingless, emanating from distinct borders, cooperate to establish cell fate across the engrailed domain in the Drosophila epidermis. Development 1997, 124, 4837–4845. [Google Scholar] [PubMed]
- Yarnitzky, T.; Min, L.; Volk, T. An interplay between two EGFR-receptor ligands, Vein and Spitz, is required for the formation of a subset of muscle precursors in Drosophila. Mech. Dev. 1998, 79, 73–82. [Google Scholar] [CrossRef]
- Figeac, N.; Jagla, T.; Aradhya, R.; da Ponte, J.P.; Jagla, K. Specification and behavior of AMPs, muscle-committed transient Drosophila stem cells. Fly 2011, 5, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, X.; Xi, R. EGFR and notch signaling respectively regulate proliferative activity and multiple cell lineage differentiation of Drosophila gastric stem cells. Cell Res. 2014, 24, 610–627. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Jiang, G.; Blume-Jensen, P.; Hunter, T. Epidermal growth factor-induced tumor cell invasion and metastasis initiated by dephosphorylation and downregulation of focal adhesion kinase. Mol. Cell. Biol. 2001, 21, 4016–4031. [Google Scholar] [CrossRef] [PubMed]
- Cela, C.; Llimargas, M. EGFR is essential for maintaining epithelial integrity during tracheal remodelling in Drosophila. Development 2006, 133, 3115–3125. [Google Scholar] [CrossRef] [PubMed]
- Brodu, V.; Casanova, J. The RhoGAP crossveinless-c links trachealess and EGFR signaling to cell shape remodeling in Drosophila tracheal invagination. Genes Dev. 2006, 20, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Inoue, Y.; Hayashi, S. A wave of EGFR signaling determines cell alignment and intercalation in the Drosophila tracheal placode. Development 2007, 134, 4273–4282. [Google Scholar] [CrossRef] [PubMed]
- Letizia, A.; Sotillos, S.; Campuzano, S.; Llimargas, M. Regulated Crb accumulation controls apical constriction and invagination in Drosophila tracheal cells. J. Cell Sci. 2011, 124, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.; Zinn, K. Receptor tyrosine phosphatases control tracheal tube geometries through negative regulation of EGFR signaling. Development 2009, 136, 3121–3129. [Google Scholar] [CrossRef] [PubMed]
- Inbal, A.; Volk, T.; Salzberg, A. Recruitment of ectodermal attachment cells via an EGFR-dependent mechanism during the organogenesis of Drosophila proprioceptors. Dev. Cell 2004, 7, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Brodu, V.; Elstob, P.R.; Gould, A.P. EGF receptor signaling regulates pulses of cell delamination from the Drosophila ectoderm. Dev. Cell 2004, 7, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Chen, X.; Cormier, O.; Cheng, D.C.; Reed, B.; Harden, N. Modulation of morphogenesis by EGFR during dorsal closure in Drosophila. PLoS ONE 2013, 8, e60180. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, A.; Agapite, J.; McCall, K.; Steller, H. The Drosophila gene hid is a direct molecular target of Ras-dependent survival signaling. Cell 1998, 95, 331–341. [Google Scholar] [CrossRef]
- Tamada, M.; Zallen, J.A. Square cell packing in the Drosophila embryo through spatiotemporally regulated EGFR receptor signaling. Dev. Cell 2015, 35, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.D.; Cagan, R.L. Drosophila lung cancer models identify trametinib plus statin as candidate therapeutic. Cell Rep. 2016, 14, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Gruber, J.; Yee, Z.; Tolwinski, N.S. Developmental drift and the role of wnt signaling in aging. Cancers 2016, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Edgar, B.A. EGFR signaling regulates the proliferation of Drosophila adult midgut progenitors. Development 2009, 136, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Kuraishi, T.; Lemaitre, B. Drosophila EGFR pathway coordinates stem cell proliferation and gut remodeling following infection. BMC Biol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Read, R.D. Drosophila melanogaster as a model system for human brain cancers. Glia 2011, 59, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Read, R.D.; Cavenee, W.K.; Furnari, F.B.; Thomas, J.B. A Drosophila model for EGFR-Ras and PI3K-dependent human glioma. PLoS Genet. 2009, 5, e1000374. [Google Scholar] [CrossRef] [PubMed]
- Sonoshita, M.; Cagan, R.L. Modeling human cancers in Drosophila. Curr. Top. Dev. Biol. 2017, 121, 287–309. [Google Scholar] [PubMed]
- Yamamoto, S.; Jaiswal, M.; Charng, W.L.; Gambin, T.; Karaca, E.; Mirzaa, G.; Wiszniewski, W.; Sandoval, H.; Haelterman, N.A.; Xiong, B.; et al. A Drosophila genetic resource of mutants to study mechanisms underlying human genetic diseases. Cell 2014, 159, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Bugaj, L.J.; Spelke, D.P.; Mesuda, C.K.; Varedi, M.; Kane, R.S.; Schaffer, D.V. Regulation of endogenous transmembrane receptors through optogenetic Cry2 clustering. Nat. Commun. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tischer, D.; Weiner, O.D. Illuminating cell signalling with optogenetic tools. Nat. Rev. Mol. Cell Biol. 2014, 15, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.E.; Goyal, Y.; Pannucci, N.L.; Schupbach, T.; Shvartsman, S.Y.; Toettcher, J.E. The spatiotemporal limits of developmental erk signaling. Dev. Cell 2017, 40, 185–192. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lusk, J.B.; Lam, V.Y.M.; Tolwinski, N.S. Epidermal Growth Factor Pathway Signaling in Drosophila Embryogenesis: Tools for Understanding Cancer. Cancers 2017, 9, 16. https://doi.org/10.3390/cancers9020016
Lusk JB, Lam VYM, Tolwinski NS. Epidermal Growth Factor Pathway Signaling in Drosophila Embryogenesis: Tools for Understanding Cancer. Cancers. 2017; 9(2):16. https://doi.org/10.3390/cancers9020016
Chicago/Turabian StyleLusk, Jay B., Vanessa Y. M. Lam, and Nicholas S. Tolwinski. 2017. "Epidermal Growth Factor Pathway Signaling in Drosophila Embryogenesis: Tools for Understanding Cancer" Cancers 9, no. 2: 16. https://doi.org/10.3390/cancers9020016