Production of Structured Phosphatidylcholine with High Content of Myristic Acid by Lipase-Catalyzed Acidolysis and Interesterification


Department of Chemistry, Wroclaw University of Environmental and Life Sciences, Norwida 25, 50-375 Wroclaw, Poland
*
Author to whom correspondence should be addressed.
Catalysts 2018, 8(7), 281; https://doi.org/10.3390/catal8070281
Submission received: 6 June 2018
/
Revised: 4 July 2018
/
Accepted: 12 July 2018
/
Published: 14 July 2018
(This article belongs to the Special Issue Catalyzed Synthesis of Natural Products)
Abstract
:Synthesis of structured phosphatidylcholine (PC) enriched with myristic acid (MA) was conducted by acidolysis and interesterification reactions using immobilized lipases as catalysts and two acyl donors: trimyristin (TMA) isolated from ground nutmeg, and myristic acid obtained by saponification of TMA. Screening experiments indicated that the most effective biocatalyst for interesterification was Rhizomucor miehei lipase (RML), whereas for acidolysis, the most active were Thermomyces lanuginosus lipase (TLL) and RML. The effect of the molar ratio of substrates (egg-yolk PC/acyl donor), enzyme loading, and different solvent on the incorporation of MA into PC and on PC recovery was studied. The maximal incorporation of MA (44 wt%) was achieved after 48 h of RML-catalyzed interesterification in hexane using substrates molar ratio (PC/trimyristin) 1/5 and 30% enzyme load. Comparable results were obtained in toluene with 1/3 substrates molar ratio. Interesterification of PC with trimyristin resulted in significantly higher MA incorporation than acidolysis with myristic acid, particularly in the reactions catalyzed by RML.
1. Introduction
Myristic acid (MA) is a 14-carbon lipid molecule belonging to long-chain saturated fatty acids. It usually accounts for small amounts (approximately 1 wt%) of total fatty acids (FA) in animal tissues, but is more abundant in milk fat (7–12 wt% of total FA) [1]. In copra and palmist oils, it makes up 15–23 wt% and 15–17 wt% of total FA, respectively [2], but the highest content of MA has been found in nutmeg (Myristica fragrans) in the form of trimyristin, which comprises approximately 45% by weight of the nutmeg butter [3].
Epidemiological and clinical studies have shown that dietary fats containing high levels of saturated fatty acids (SFA, usually more than 15% of total energy) induce an increase in plasma total cholesterol and low-density lipoprotein cholesterol (LDL cholesterol) concentrations in humans [4]. Along with other saturated FA, myristic acid has deleterious effects on blood cholesterol level in animals and humans when provided at high levels exceeding 4% of dietary energy [4,5], but these negative effects disappear when its dietary level is in a narrow physiological range (1.0–2.5% of dietary energy) [2,6]. In European countries, myristic acid consumption is 4–8 g per day, which corresponds to 0.5 to 2.0% of total energy. Furthermore, some studies have shown that specific biochemical functions can be assigned to MA; it is known to modify activity of many enzymes and protein functions of both eukaryotic and viral origin through the myristoylation of their N-terminal glycine residues [7,8,9]. The myristoyl residue of proteins increases their affinity to target cell membranes. Myristic acid may be also considered as one of the regulators of cellular bioactive lipid concentration, such as polyunsaturated fatty acids and ceramide [10]. The positive role of MA in the overall conversion of α-linolenic acid to Long Chain Polyunsaturated Fatty Acids (LC-PUFA), like eicosapentaenoic (EPA) and docosahexaenoic (DHA) acids, has been shown in rat and human nutritional experiments [2,11,12,13]. The effect of myristic acid on n-3 PUFA content follows a U-shaped curve, and a beneficial maximum level at around 1.2% of total daily energy is observed [13]. Positive correlation between myristic acid intake and the level of n-3 PUFA is connected with the myristoylation of NADH-cytochrome b5 reductase accounting for the increased Δ6-desaturase activity. In turn, the role of MA in the biosynthesis of ceramide lies in the myristoylation of dihydroceramide ∆4-desaturase. MA also exerted anxiolytic-like effects, producing comparable actions to diazepam in Wistar rats subjected to the elevated plus maze [14].
Phospholipids (PLs) are widely used in the food, cosmetic, and pharmaceutical industries [15]. Interest in the production of structured phospholipids (SPL) containing specific fatty acid residues has grown significantly in recent years. Replacement of existing fatty acids in native PL with desirable fatty acids can improve not only their physical and chemical properties, but also their nutritional, pharmaceutical, and medical functions [16]. Novel SPL were created by incorporating into natural PL functional fatty acids that have multiple biochemical and pharmacological effects on human health and treatment/prevention of some diseases (e.g., docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), and conjugated fatty acids) [17,18,19,20,21,22,23,24].
Compared with chemical methods, enzymatic modification of PL has advantages that are attributed to the specificity of the enzymes. Lipases of microbial origin and phospholipase A1, namely Lecitase Ultra, have been the most commonly used enzymes for the exchange of fatty acids on PL at sn-1 positions, whereas phospholipase A2 have been employed to the modification of sn-2 position [15,17,19,20,25,26]. Regioselectivity of lipases allows the specific removal or/and replacement of the acyl chains at position sn-1 of PL via hydrolysis followed by re-esterification or through direct transesterification with acyl donors (acidolysis with free fatty acids and interesterification with esters of fatty acids).
In our previous study, we reported on a pioneer method for the production of structured phosphatidylcholine (SPC) using plant oils as the source of natural acyl donors in the form of triacylglycerols. Using this methodology, phospholipids were enriched with unsaturated fatty acids from the n-3 or n-6 family in the process of interesterification with corresponding oil, that is, linseed, safflower, sunflower, borage, and primrose evening oil [27]. The same reaction system with pomegranate seed oil as the acyl donor was applied in the incorporation of conjugated linolenic acid (punicic acid) into egg-yolk phosphatidylcholine (PC) [21]. In turn, concentrates of n-3 PUFA and CLA (conjugated linolenic acid) produced from fish oil and sunflower oil, respectively, were applied to the enrichment of egg-yolk PC with desired fatty acids in the process of acidolysis [18,22].
Phospholipids (PLs) can be also an efficient dietary vehicle to deliver MA into the target cells for the myristoylation of proteins. Werbovetz and Englund reported 1-myristoyllysophosphatidylcholine (M-LPC) as myristate donor for the myristoylation of glycosyl phosphatidylinositol (GPI) anchor of the Trypanosoma brucei variant surface glycoprotein (VSG) [28]. M-LPC is first hydrolyzed by plasma membrane-associated phospholipase to release a cell-associated MA that, after uptake by the cell and conversion to mirystoyl-CoA, becomes available for the myristoylation of GPI catalyzed by myristoyltransferase.
Considering the role of myristic acid in the proteins activation and the positive effect of myristic acid on the EPA and DHA levels when supplied at around 1.2% of daily energy, we were encouraged to produce new PC species containing MA in the sn-1 position. As M-LPC, similar to other LPC species, can be easily accessible from 1-myristoyl-2-acyl-PC via the hydrolysis catalyzed by phospholipase A2 [29], we decided to elaborate the novel methods for incorporation of MA into the sn-1 position of egg-yolk PC. In the presented work, continuing our studies on the enzymatic production of PLs using acyl donors of natural origin, we report on the lipase catalyzed-production of MA-enriched PC using two acyl donors: trimyristin (TMA) prepared from nutmeg, and MA obtained from trimyristin by saponification. Using these acyl donors allowed us to compare two reaction systems: interesterification and acidolysis.
2. Results and Discussion
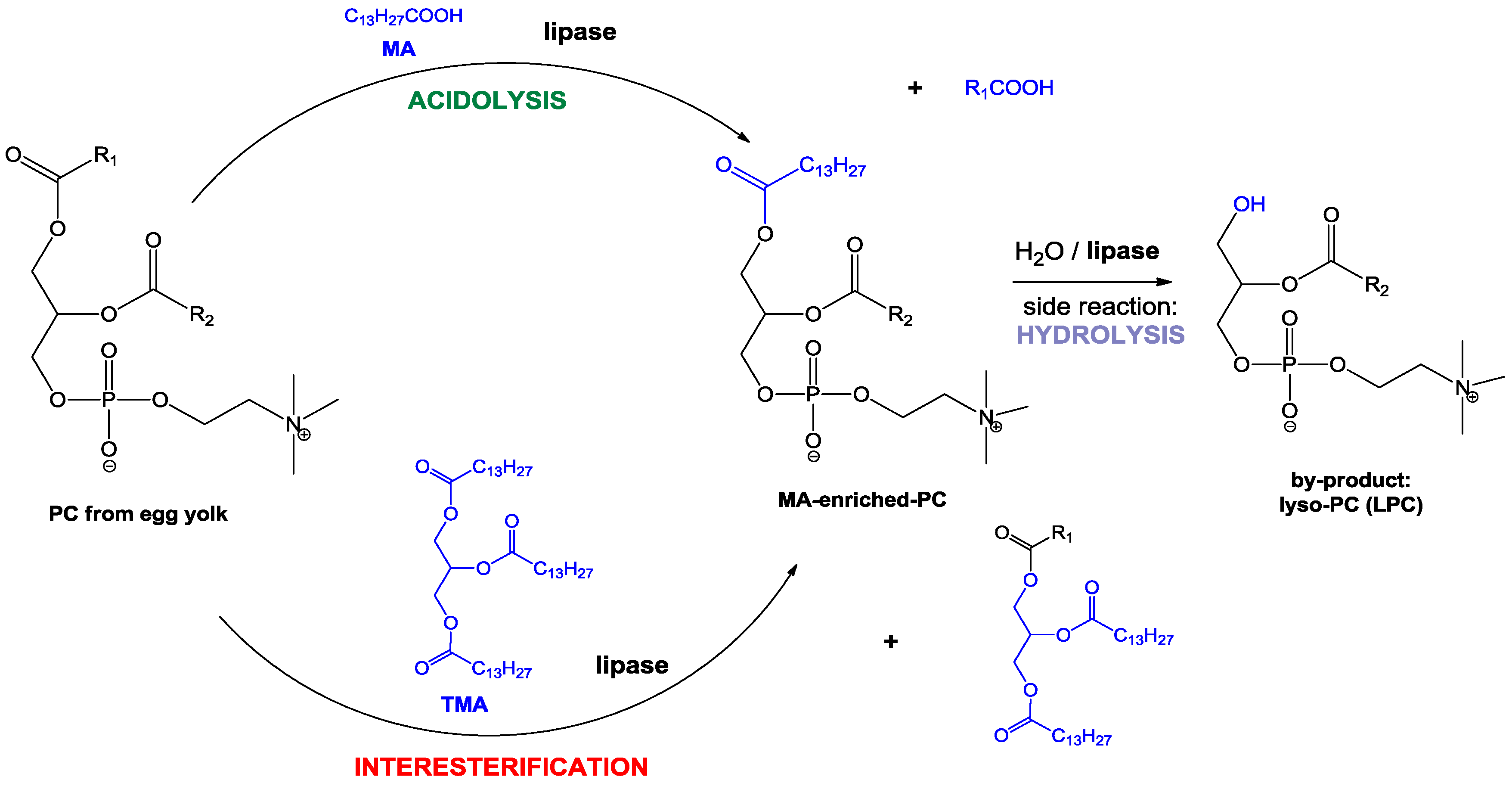
The lipase-catalyzed reaction systems that were applied to obtain MA-riched PC are depicted in Scheme 1.
The raw material for our studies were PC isolated from egg-yolk (purity >99% according to HPLC) and two acyl donors (trimyristin and myristic acid). TMA was isolated from ground nutmeg by continuous Soxhlet extraction with hexane and subsequent crystallization from acetone. Myristic acid was obtained by saponification of TMA isolated previously from nutmeg.
PC, TMA, and MA were subjected to gas chromatography (GC) analysis to determine their fatty acid composition (Table 1). The results for PC showed that palmitic (C16:0) and oleic (C18:1) acids were the predominating ones, and the former, as well as stearic acid (C18:0), are located in the sn-1 position, whereas the sn-2 position is occupied by unsaturated acids from the n-6 (linoleic acid and arachidonic acid) and n-9 family (oleic acid).
Analysis of the FA composition of trimyristin indicated the high content of myristic acid (over 83 wt%) and some small amounts of other acids (about 6 wt%, C16:0 and 5 wt%, C18:1). The purity of myristic acid obtained by saponification of TMA, given as wt% of myristic acid according to GC, was approximately 92%.
2.1. Screening of Enzymes
Four commercially available lipases; Lipozyme® (RML), Lipozyme TL IM (TLL), Candida antarctica lipase B (Novozym 435) and Candida antarctica lipase A (CALA) were evaluated for their ability to catalyze interesterification and acidolysis between egg-yolk PC and TMA or MA, respectively. Lipozyme® and Lipozyme TL IM are classified as 1,3-regiospecific lipases [15] the third one is considered as a non-specific enzyme, but in most lipid modifications it shows high selectivity towards the sn-1 position [27,30,31]. The fourth of the lipases (CALA) is reported to show a sn-2-regioselectivity [32,33], although such preference in the structure of triacylglycerols (TAG) is not sufficient to selective synthesis of 1,3-diglycerides or 2-monoglycerides [34]. Therefore, for practical interesterifications, CALA has been reported rather as a nonselective lipase [35]. All enzymes were used in immobilized form, which made it possible to carry out the reactions in the presence of solvent and at higher temperatures.
The initial conditions applied for both reaction systems (acidolysis and interesterification) were as follows: temperature 50–52 °C; 1/3 PC/acyl donor molar ratio and 30% enzyme dosage. Having in mind a decrease of process costs, the employed lipases were used at the same weight ratios, despite different enzymatic activities declared by the suppliers.
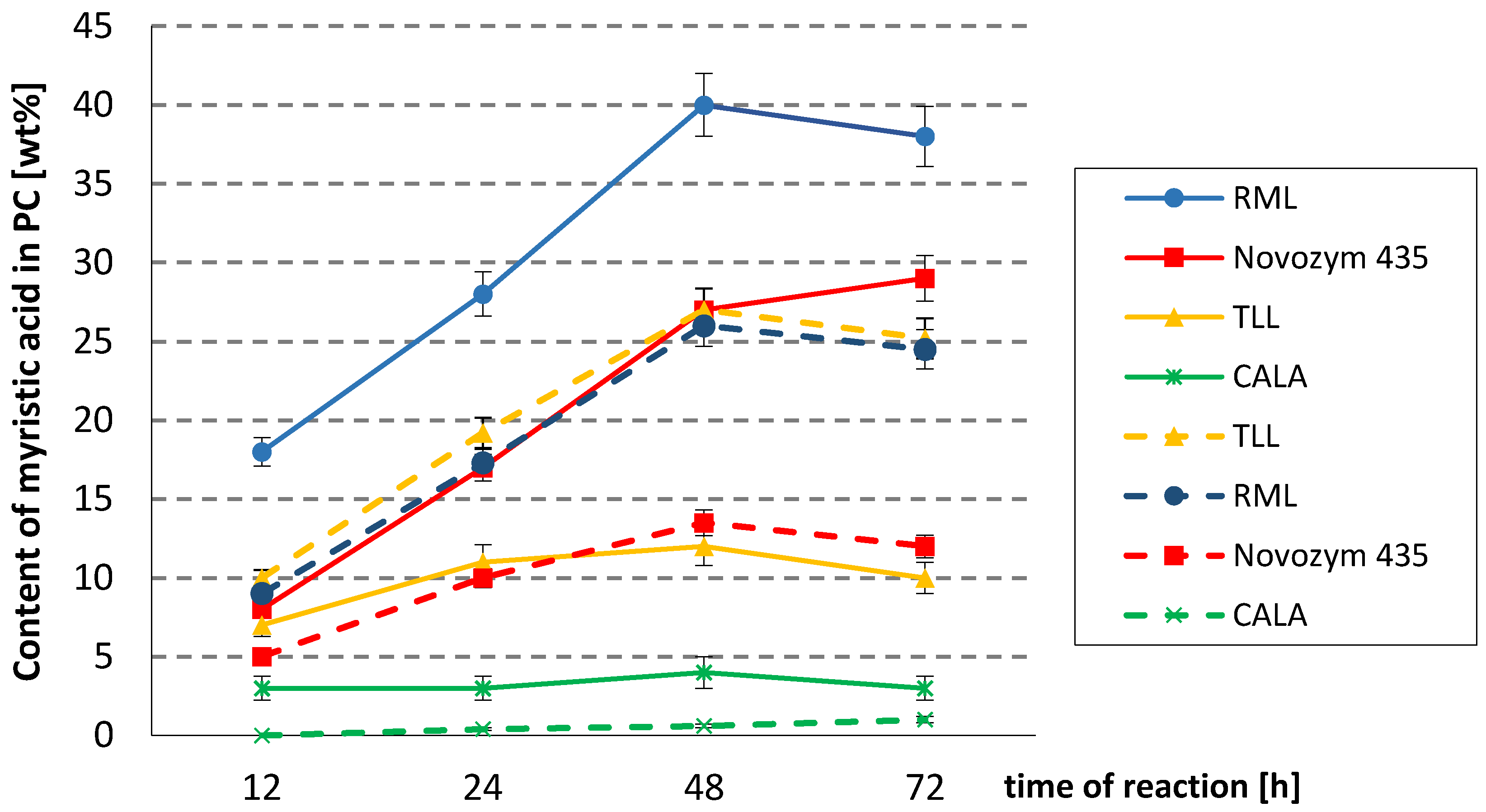
The time course of the incorporation of MA into PC by lipases in both processes is shown in Figure 1.
The screening experiments indicated that immobilized lipases from Rhizomucor miehei (Lipozyme RM IM, RML) and Thermomyces lanuginosus (Lipozyme TL IM, TLL) exhibited the highest activity. The first one was the most efficient in interesterification (40 wt% incorporation of MA into egg-yolk PC after 48h), whereas in acidolysis, both mentioned lipases exhibited satisfactory activity. However, less incorporation degree was observed at the same time. In the reaction catalyzed by TLL, the incorporation of MA reached a maximum (27 wt%) within 48 h, whereas in RLM-catalyzed acidolysis, 26 wt% of incorporation was observed. In this investigation, Novozym 435 showed less activity, giving 27.5 wt% and 13.5 wt% incorporation of MA into PC in interesterification and acidolysis, respectively. In our previous studies, this lipase was the most efficient in incorporation of α-linolenic acid, punicic acid, and PUFA into PC in the interesterification and acidolysis processes [18,21,27,30]. On the other hand, similar to the results presented here, TLL was less active in interesterification of PC with pomegranate seed oil. Comparing acidolysis of PC with concentrates from fish oil and myristic acid, higher selectivity of TLL towards MA was observed.
Lipase A from Candida antarctica was almost inactive in both the interesterification and acidolysis processes; after 72 h of reactions, no more than 4 wt% incorporation of MA was achieved.
Collating efficiency of MA incorporation, about two-fold higher introduction of myristic acid in the process of interesterification than in acidolysis catalyzed by both RML and Novozym 435 can be seen. The reason is the fact, that compared with myristic acid, trimyristin is a more efficient acyl donor, containing two acyl groups in the sn-1 and sn-3 positions, both accessible in interesterification of PC for 1,3-regioselective lipases used as biocatalysts.
Because the highest degree of incorporation of MA into PC in interesterification was achieved for RML, while in acidolysis, comparably high results were achieved for TLL and RML, these enzymes were selected for subsequent experiments.
2.2. Effect of Substrate Molar Ratio
One of the most important factors affecting both esterification and transesterification reactions is an acyl donor concentration [36].
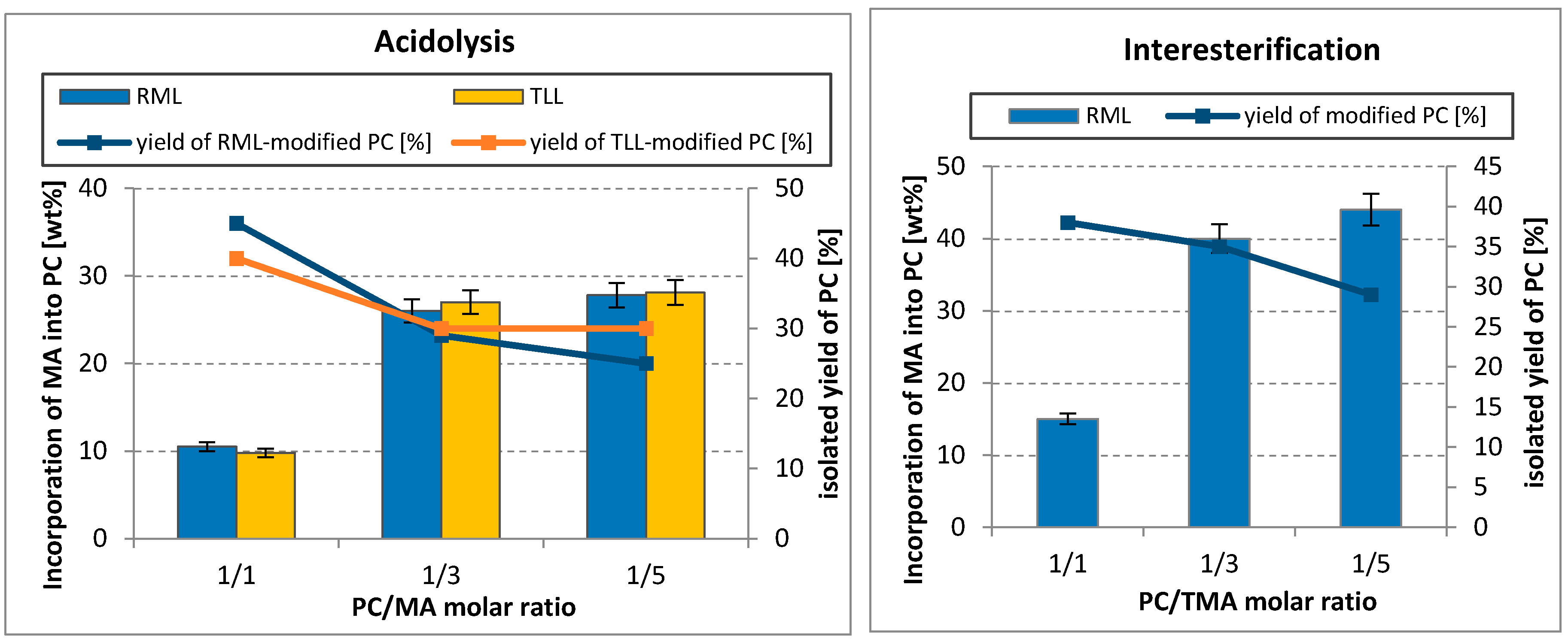
The effect of substrates molar ratio (1/1, 1/3, and 1/5; PC/acyl donor: MA or TMA) on the degree of myristic acid incorporation into PC was evaluated for RML lipase in both interesterification and acidolysis and for TLL in the acidolysis process (Figure 2).
The results showed that for both lipases and both compared processes, the content of myristic acid in modified PC rose with an increasing substrates molar ratio with a simultaneous decrease in isolated yield of PC. In both reaction systems, the highest increase of MA incorporation into PC was observed when the ratio changed from 1/1 to 1/3 (PC/acyl donor). An almost three-fold growth of incorporation (from 9.8 to 27 wt%) can be observed for TLL-catalyzed acidolysis. In the case of the RML, increase of MA incorporation was significantly higher in the process of interesterification (from 15 to 40 wt%) than in acidolysis (from 10.5 to 26 wt%). A further enhancement of PC/acyl donor to 1/5 molar ratio caused an insignificant rise of embedded acid, whose content in modified PC made up about 28 wt% of total FA after acidolysis and 44 wt% after interesterification. It should be noticed that in both reaction systems, equilibrium is moved to the product side with an increasing substrate ratio, which improves acyl incorporation. Simultaneously, the overall PC yield decreased, which may be the result of hydrolysis side-reaction with 2-acyl LPC formation (Scheme 1) and next spontaneous acyl migration resulting in 1-acyl LPC, which may be quickly hydrolyzed to glycerophosphocholine (GPC). A drop of isolated yield of modified PC may be also related with difficulties in separating the unreacted acyl donors and exchanged fatty acids or triacylglycerols from the final products. As a result of that and taking into consideration the economy of the process, 1/3 molar ratio (PC/acyl donor) was chosen for subsequent experiments.
The effect of the substrate molar ratio on the incorporation of other saturated FA into phosholipids was investigated in several studies. Reddy et al. optimized the molar ratio PC/palmitic acid, changing it from 1:2 to 1:10, and established 1/5 molar ratio for maximal incorporation of palmitic acid into PC in the reaction catalyzed by Lipozyme TL IM [37]. Further increase of this parameter from 1/5 to 1/10 did not change the incorporation significantly. During acidolysis of PL with caprylic acid catalyzed by TLL in a packed bed reactor, the increase of incorporation with increasing substrate molar ratio from 1/4 to 1/36 PL/caprylic acid was observed, reaching 35.8% of incorporation [38]. On the other hand, in the solvent-free system, the incorporation of caprylic acid was 13.1 and 24.4% with PC/caprylic acid ratios of 1/6 and 1/8, respectively.
2.3. Effect of Enzyme Dosage
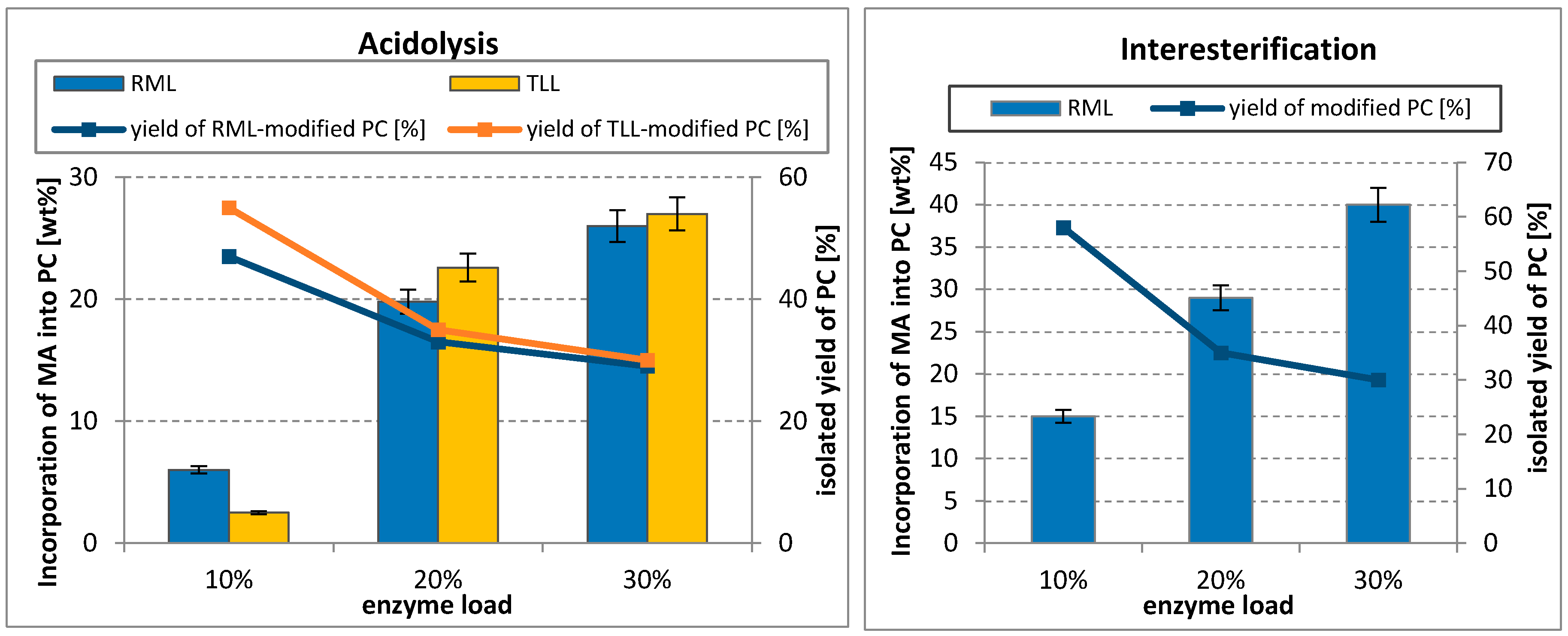
Enzyme dosage was the most significant factor on the lipase-catalyzed acidolysis and interesterification in terms of incorporation of MA into PC and PC recovery.
Incorporation into PC increased with increasing enzyme dosage (Figure 3). The most meaningful rise of MA in PC, over nine-fold for TLL and three-fold for RML-catalyzed acidolysis, was observed when the enzyme load was increased from 10 up to 20 wt%. Simultaneously, a decrease of modified PC yield was appreciable (reduced from 47 to 33 wt% for RML and from 55 to 35 wt% for TLL). A similar effect can be noticed for interesterification process catalyzed with RML. Incorporation of MA raised from 15 to 29 wt%, while the total isolated yield of PC decreased over 1.5-fold. It has been reported earlier that high enzyme dosage is important to achieve effective incorporation of fatty acids into PLs by acidolysis [33,34], but with the increase of the enzyme concentration, usually hydrolysis of PC proceeded lowering the total isolated yield of PC [35].
Increasing the dosage of the enzyme to 30% in the case of acydolysis resulted in a slight increase of myristic acid in PC (26 and 27 wt% incorporation for RML and TLL, respectively), with a little decrease of the total yield of modified PC. A clear increase of MA incorporation (from 29 up to 40 wt%) was observed for RML-catalyzed interesterification, as well as a small reduction of PC recovery.
In summary, with the increase of the enzyme dosage in both reaction systems, higher incorporation of MA can be obtained with simultaneous losses of PC. However, taking into consideration the cost of the enzyme, 20 wt% of enzyme load seems to be enough to achieve high incorporation of MA with a sufficient PC yield. The same enzyme dosage was found to be optimal by Reddy et al., who studied the effect of lipase dosage on the incorporation of palmitic and stearic acid in the TLL-catalyzed acidolysis of PC from egg-yolk [37]. Increasing enzyme dosage from 5 to 20%, the incorporation of palmitic acid and stearic acid reached up to 21.2% and 36.1%, respectively. Further increase of the enzyme load to 25% did not affect the incorporation of both acids.
2.4. Effect of Organic Solvents
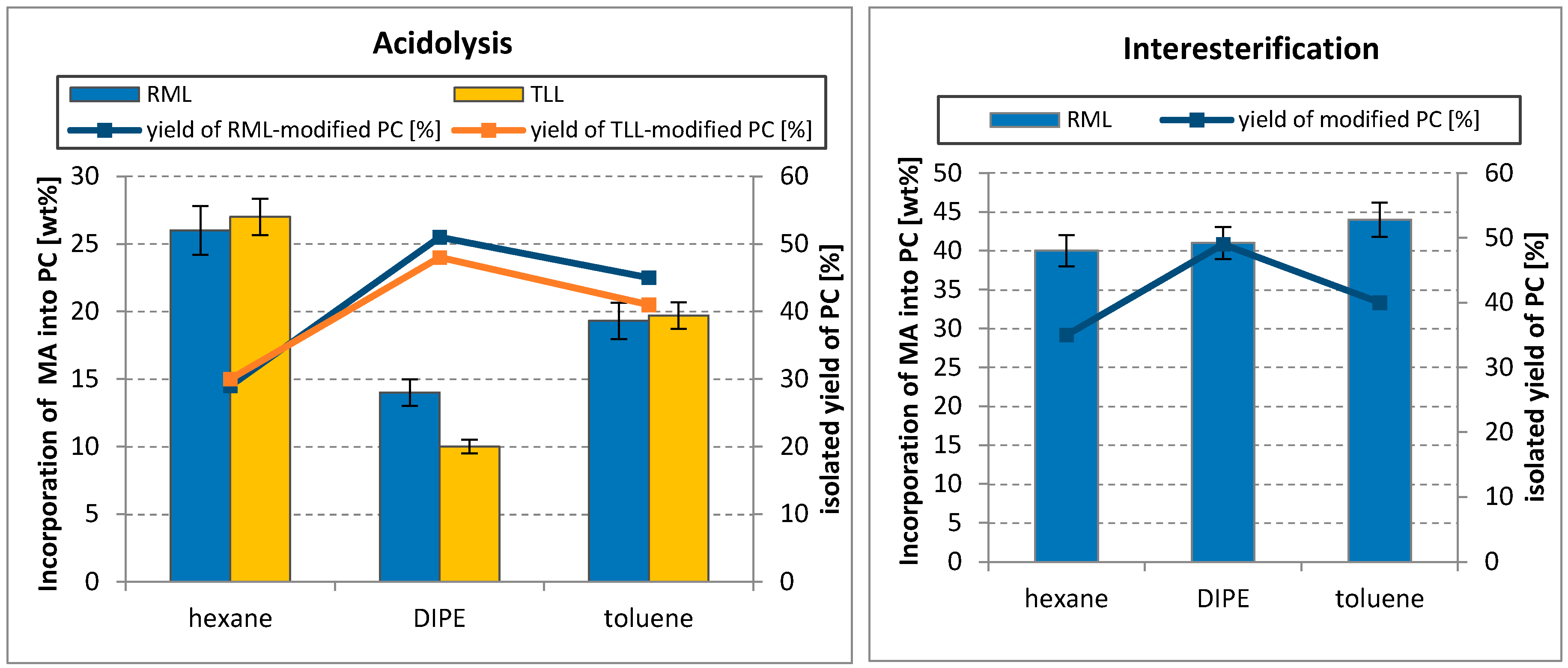
Three organic solvents were tested in both reaction systems. Generally, no significant differences in MA incorporation were observed in the case of interesterification, the content of myristic acid in PC was in the range of 40–45 wt% for all solvents used. In the case of acidolysis, the effect of the solvent was pronounced for both RML and TLL (Figure 4). The highest incorporation (about 26 wt%) was obtained after acidolysis was carried out in hexane. Using toluene resulted in lowering incorporation by 6 wt% and a relevant drop in the content of MA in modified PC (to 10 wt% of total fatty acid) was observed for diisopropyl ether (DIPE), particularly for TLL-catalyzed reaction. In both reaction systems, increase of PC yield was found when DIPE was applied.
Polarity of the solvent is crucial for holding a layer of water around the enzyme, and for this purpose, hydrophobic solvents are the best choice. Logarithm of the partition coefficients (log P) for the solvents used were as follows: 3.5 for hexane, 2.5 for toluene, and 1.52 for diisopropyl ether (DIPE). Solvents with log P in the range of 2–4 are considered as suitable for lipase-catalyzed reactions and the results of acidolysis confirmed the relationship between solvent polarity and enzyme activity. However, using DIPE in the interesterification gave MA incorporation values comparable to those obtained for hexane and toluene. No effect of solvent on the incorporation of caprylic acid into PC was also observed by Kim and Yoon during acidolysis catalyzed by Mucor javanicus lipase [39]. They used three solvents with significantly different polarity indexes: hexane, diethyl ether, and methanol; but in all cases, incorporation of caprylic acid was approximately 36%.
There have been several reports on the introduction of saturated fatty acids into phospholipids using enzymes to improve emulsifying and dispersing properties or heat stability, but only few concern myristic acid. Gan et al. introduced a series of saturated fatty acids (C6:0, C8:0, C10:0, C12:0, C14:0) into the sn-1 position of lecithin (mixture of phospholipids with ≥30% of PC) by Lecitase Ultra-catalyzed acidolysis (showing both lipase and phospholipase A1 activity) [40]. They achieved different molar incorporation values, depending on the chain length; the highest one was observed for capric acid (C10:0, 51%), whereas the lowest was for caproic acid (C6:0, 28.9%). The reactions were carried out in hexane at high substrates molar ratio (10/1 MA/lecithin) and 10% of immobilized enzyme load and incorporation of myristic acid in these conditions was 36.4%. Caprylic acid (C8:0) was also successfully introduced into PC by different lipases: RML, TLL, and Mucor javanicus lipase; incorporation values were 46.3%, 35.4%, and 36.7%, respectively [38,39,41]. TLL was also used for the acidolysis of soybean phosholipids with capric acid (C10:0) and 23.16% of incorporation was achieved after 10 days of reaction at hexane, at 1/4 substrates molar ratio (PL/capric acid) and 10% enzyme load [42]. An interesting method was proposed by Hama et al. [43]. Using immobilized Rhizopus oryzae whole-cells at 30 °C in hexane at 1/8 substrates molar ratio (PC/fatty acid), introduction of lauric acid (C12:0) into egg-yolk PC at the level of 44.2% was achieved. The biocatalyst used was more efficient than different lipase powders. Lipase-catalyzed enrichment of PC with palmitic (C16:0) and stearic acid (C18:0) was performed by Reddy et al.; the best results for egg-yolk PC were obtained using TLL when the content of 16:0 was increased from 37.4% to 58.6%, and stearic acid from 8.6% to 44.7%, whereas for soybean PC, higher incorporation (44.2% for 16:0 and 53.7% for C 18:0) was found for Novozyme 435. In these studies, 20% enzyme load, 1/5 PC/FA molar ratio, and reaction time 24 h were chosen as optimal conditions for maximal incorporation of both incorporated acids into PC [37].
Egger et al. [44] studied the phospholipase A2-catalyzed esterification of LPC with different fatty acids, including myristic acid. The reaction carried out in toluene afforded 1-palmitoyl-2-myristoyl-PC, but the reaction rate was low (below 2 nmol of product per h and mg of enzyme preparation) and no information about the PC yield is given. The only one report concerns the introduction of myristic acid into the sn-1 position of PLs using lipase is the interesterification between soybean PC and methyl myristate catalyzed by RML [45]. Using a solvent free system and 1/5 substrate molar ratio (soy PC/methyl myristate), as well as 10% of the enzyme load, the authors achieved 15.7% MA incorporation in the reaction conducted at 60 °C. Using methyl caprate or methyl laurate as the acyl donors, authors achieved 8.4% and 14.1% of incorporation of capric acid and lauric acid, respectively. No further studies on the effect of reaction condition were presented.
In this paper, for the first time, the complex studies on the lipase-catalyzed incorporation of myristic acid from natural source into egg-yolk PC were presented. Similar to most other investigations concerning incorporation of saturated fatty acids into phospholipids, RML and TLL turned out to be the most active biocatalysts. Comparison of two reaction systems proved higher efficiency of interesterification with trimyristin when compared with acidolysis with myristic acid. The use of 1,3 regioselective lipases allowed as to produce PC enriched with MA mainly in the sn-1 position (over 80 wt%), leaving unsaturated acids (45.5 wt% C18:1, 29.3 wt% C18:2, and 9.4 wt% C20:4) in their original, internal position (Table 1). The results show the characteristic decrease of palmitic and stearic acid (from 64.1 to 12.1 wt% and from 29.2 to 2.5 wt%, respectively) in the external position of PC because of their replacement by myristic acid. These results confirm the regioselectivity of lipases used, a small amount of myristic acid in the sn-2 position may be the result of acyl migration during the processes.
An elaborated method of interesterification with Rhizomucor miehei lipase and trimyristin as the acyl donor led to significantly higher incorporation degree (40–44 wt%) of myristic acid than those reported by Ghosh [45], at the lower substrate molar ratio (1/3, PC/acyl donor) and lower temperature (50–52 °C). The presented results confirm the usefulness of natural triacylglycerols as the effective acyl donors in the interesterifications of phospholipids.
3. Materials and Methods
3.1. Materials and Chemicals
Lohmann Brown hens’ eggs were purchased from the poultry farm “Ovopol” (Nowa Sól, Poland). Lipozyme TL IM (a silica granulated Thermomyces lanuginosus lipase preparation, 250 U/g) was a gift from the Novozymes A/S (Bagsvaerd, Denmark). Lipase B from Candida antarctica immobilized in a macroporous acrylic resin (synonym: Novozym 435, >5000 U/g), lipase A from Candida antarctica (CALA, >500 U/g) immobilized on resin Immobead 150, and lipase from Mucor miehei immobilized in macroporous ion-exchange resin (Lipozyme®, >30 U/g) were purchased from Sigma-Aldrich (St. Louis, MO, USA). A boron trifluoride methanol complex solution (13–15% BF3 × MeOH) was purchased from Sigma-Aldrich (St. Louis, MO, USA). All other chemicals were of analytical grade. Silica gel-coated aluminum plates (Kieselgel 60 F254, 0.2 mm) used in thin layer chromatography (TLC) and the silica gel (Kieselgel 60, 230–400 mesh) used in the column chromatography were purchased from Merck.
3.2. Analysis of Substrates and Products
The purity of the native and modified phosphatidylcholine was determined by HPLC on an Ultimate 3000 DIONEX chromatograph equipped with CoronaTM Charged Aerosol Detector (CAD). A Waters Spherisorb S5W column (150 × 4.6 mm) was used for analysis. The HPLC gradient program was as follows: (flow rate 0.6 mL × min−1); 0 min 0/90/10 (%A/%B/%C) at 2 min, 0/40/60 at 20 min, 1/40/59 at 22 min, 10/40/50 at 38 min, 8/40/52 at 44 min, 1/40/59 at 55 min, 0/90/10 at 56 min, and 0/90/10 hold 10 min (A/B/C, water/0.1% solution of formic acid in hexane/isopropanol).
Fatty acid profiles of starting materials and structured PC were determined after their conversion to the fatty acid methyl esters (FAME) according to the following procedure: samples (50 mg) were heating under reflux (3 min) with 3 mL of BF3 × MeOH complex solution. After cooling, the mixtures were extracted with 2 mL of hexane and the organic layers were washed with a saturated NaCl solution. Hexane extracts were dried over anhydrous magnesium sulphate (MgSO4) and analyzed directly by gas chromatography (GC) on an Agilent 6890 N instrument using a 70% cyanopropyl polysilphenylene-siloxane column (TR FAME, 30 m × 0.25 mm × 0.25 μm) and flame ionization detector. The temperature parameters were as follows: injector 250 °C, detector 280 °C, column: 160 °C (held 3 min), 160–220 °C (rate 5 °C·min−1), 220–260 °C (rate 30 °C·min−1), 260 °C (held 3 min). The FAME were identified by comparing their retention times with those of a standard FAME mixture (Supelco 37 FAME Mix) purchased from Sigma-Aldrich.
The incorporation of myristic acid into PC is refered to the total amount of PC species obtained after enzymatic modification (original and modified PC molecules).
The positional analysis of fatty acids in native and modified PC was based on regiospecific Lipozyme®-catalyzed ethanolysis of PC, which allowed us to obtain fatty acids ethyl esters (FAEE) released from the sn-1 position of PC and 2-acyl LPC. FAEE were analyzed directly by gas chromatography to give the composition of FA in the sn-1 position of PC. After purification, 2-acyl LPC were trans-esterified using ethanolic NaOH solution and boron trifluoride etherate. FAEEs obtained were isolated from the reaction mixture and analyzed by gas chromatography. In this way, FAs composition in the sn-2 position of the PC was determined. The details of the procedure were described in our previous paper [46].
3.3. Isolation of PC from Egg Yolk
The extraction of phospholipids from egg yolk was performed on a semi-technical scale in Wroclaw Technology Park. Eggs were dried in the drying chamber at inlet air temperature 185 ± 5 °C and an outlet air temperature 70 ± 2 °C. In the next step, obtained powder was extracted with ethanol in a tank equipped with a mechanical stir maintaining the yolk/solvent ratio 1:4 (m/v). The process of suspension was carried out for 90 min and then alcohol was removed by filtration. The residue was evaporated in vacuo (0.06 MPa at 50 °C). The pure PL-fraction was obtained from the crude PL by precipitation with cold acetone [47]. The pure PC was separated from this fraction by silica gel column chromatography (chloroform/methanol/water, 65:25:4, v/v/v). The purity of PC fractions was analyzed by TLC on silica gel-coated aluminum plates (chloroform/methanol/water, 65:25:4, v/v/v) and HPLC. Fractions containing more than 99% of PC (according to HPLC) were collected and the solvent was evaporated.
3.4. Preparation of Acyl Donors
3.4.1. Isolation of Trimyristin from Nutmeg
Finely ground nutmeg (25 g) was placed into the extraction thimble, covered with a little cotton wad, and placed in the Soxhlet extractor. Hexane (200 mL) was poured into the flask and heated to reflux. The extraction was continued for 2–3 h (10–15 extraction cycles). The solvent was evaporated from the extract and the residue was crystallized from acetone. After 2 h, crystals of trimyristin were filtered and dried in the desiccator with anhydrous calcium chloride. The fatty acid compositions [wt%] of trimyristin were analyzed by gas chromatography (Table 1).
3.4.2. Production of Myristic Acid from Trimyristin by Saponification
The mixture of TMA isolated previously from nutmeg (10 g) dissolved in 150 mL of ethanol and 100 mL 1 M NaOH ethanolic solution was heated (78 °C) under reflux for 3 h. Then, the mixture was cooled to room temperature, poured into 100 mL of water in a large beaker, and acidified with 20 mL of concentrated hydrochloric acid to pH = 2. After 30 min, white crystals of myristic acid were filtrated on a Büchner funnel, washed with 25 mL of distilled water, and dried in laboratory drier at 40 °C overnight to afford 8.7 g of myristic acid (yield 92%).
The progress of the saponification reaction was monitored by thin layer chromatography (TLC) on a silica gel-coated aluminum plates (developing system—hexane/diethyl ether (3/1, v/v)). After elution, the plates were developed using the 0.05% primuline solution (acetone/ water, 8:2, v/v) and spots were detected under an ultraviolet (UV) lamp (λ = 320 nm).
The purity of myristic acid [wt%] was determined by GC (Table 1).
3.5. The Lipase-Catalyzed Acidolysis of PC with Myristic Acid
The egg-yolk PC (0.325 mmol, 200 mg) and myristic acid (at molar ratio of substrates 1/3, PC/MA) in 5 mL of solvent was mixed at 50 °C until complete dissolution and then 20% of lipase (by weight of substrates) was added. The reactions were carried out using four different lipases at 50–52 °C. The effect of molar ratio of substrates, lipase dosage, and different solvent was tested in another set of experiments for two enzymes with high activity: Lipozyme® and Lipozyme TL IM. Each experiment was carried out in two replications. The reaction mixtures were agitated in a heating plate with a magnetic stirrer at 300 rpm, and stopped at the selected time intervals by enzyme filtration.
After evaporation of solvent in vacuo PC was separated from the other components by column chromatography using silica gel as stationary phase and the mixture of chloroform/methanol/water (65:25:4, v/v/v) as mobile phase (eluent). The procedure was as follows: silica gel was well solvated with chloroform and the prepared slurry was gently poured into the glass column. The sample was dissolved in a minimal (2 mL) amount of chloroform and applied to the top of the column and then eluted with the solvent system mentioned above. The eluted fractions were collected in test tubes and identified by TLC on silica gel-coated aluminum plates (chloroform/methanol/water, 65:25:4, v/v/v) and HPLC. Fractions of PC were combined and dried with anhydrous magnesium (VI) sulfate. After filtration, the solvent was evaporated in vacuo to obtain pure PC (>99% according to HPLC)
Isolated yield was calculated as the total amount (by weight) of PC recovered after the process (which contains modified and unmodified PC) in relation to initial PC.
3.6. The Lipase-Catalyzed Interesterification of PC with Trimyristin
The egg-yolk PC (0.325 mmol, 200 mg) was mixed with TMA (at 1/3 molar ratio of substrates, PC/TMA) in 5 mL of hexane and after dissolving of substrates, lipase (20% by weight of substrates) was added. The reactions were carried out using four different lipases, at 50–52 °C. The effect of molar ratio of substrates, lipase dosage, and different solvent was tested in another set of experiments for Lipozyme®. Each experiment was performed in two replications. The reaction mixtures were agitated in a heating plate with a magnetic stirrer at 300 rpm, and stopped at the selected time intervals by enzyme filtration. Modified PC was separated from the mixtures by silica-gel column chromatography (chloroform/methanol/water, 65:25:4, v/v/v) according to the same procedure described in Section 3.5.
Isolated yield was estimated as the total amount (by weight) of PC recovered after the process (which contains modified and unmodified PC) in relation to initial PC.
Author Contributions
A.C. conceived and designed the experiments; A.C. and W.G performed the experiments, gas chromatography analysis, and HPLC analysis; A.C. analyzed the data and wrote the paper.
Funding
This research received no external funding.
Acknowledgments
Publication supported by Wroclaw Centre of Biotechnology, Programme The Leading National Research Centre (KNOW) for years 2014–2018 (http://know.wroc.pl).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jensen, R.G.; Ferris, A.M.; Lammi-Keefe, C.J.; Henderson, R.A. Lipids of Bovine and Human Milks: A Comparison. J. Dairy Sci. 1990, 73, 223–240. [Google Scholar] [CrossRef]
- Rioux, V.; Catheline, D.; Bouriel, M.; Legrand, P. Dietary myristic acid at physiologically relevant levels increases the tissue content of C20:5 n-3 and C20:3 n-6 in the rat. Reprod. Nutr. Dev. 2005, 45, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Power, F.B.; Salway, A.H. CLXII.—The constituents of the expressed oil of nutmeg. J. Chem. Soc. Trans. 1908, 93, 1653–1659. [Google Scholar] [CrossRef]
- Salter, A.; Mangiapane, E.; Bennett, A.; Bruce, J.; Billett, M.; Anderton, K.; Marenah, C.; Lawson, N.; White, D. The effect of different dietary fatty acids on lipoprotein metabolism: Concentration-dependent effects of diets enriched in oleic, myristic, palmitic and stearic acids. Br. J. Nutr. 1998, 79, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Hayes, K.C.; Khosla, P. Dietary fatty acid thresholds and cholesterolemia. FASEB J. 1992, 6, 2600–2607. [Google Scholar] [CrossRef] [PubMed]
- Dabadie, H.; Motta, C.; Peuchant, E.; LeRuyet, P.; Mendy, F. Variations in daily intakes of myristic and α-linolenic acids in sn-2 position modify lipid profile and red blood cell membrane fluidity. Br. J. Nutr. 2006, 96, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Towler, D.A.; Gordon, J.I.; Adams, S.P.; Glaser, L. The biology and enzymology of eukaryotic protein acylation. Annu. Rev. Biochem. 1988, 57, 69–99. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.R.; Bhatnagar, R.S.; Knoll, L.J.; Gordon, J.I. Genetic and Biochemical Studies of Protein N-Myristoylation. Annu. Rev. Biochem. 1994, 63, 869–914. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, E.; Rioux, V.; Legrand, P. Acide myristique: Nouvelles fonctions de régulation et de signalisation. Medecine/Sciences 2009, 25, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Rioux, V. Fatty acid acylation of proteins: Specific roles for palmitic, myristic and caprylic acids. Oilseeds Fats Crop. Lipids 2016, 23, D304. [Google Scholar] [CrossRef]
- Legrand, P.; Beauchamp, E.; Catheline, D.; Pédrono, F.; Rioux, V. Short chain saturated fatty acids decrease circulating cholesterol and increase tissue pufa content in the rat. Lipids 2010, 45, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Guillou, H.; D’Andrea, S.; Daval, S.; Bouriel, M.; Rioux, V.; Legrand, P. Myristic acid increases Δ6-desaturase activity in cultured rat hepatocytes. Reprod. Nutr. Dev. 2004, 44, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, P.; Rioux, V. The complex and important cellular and metabolic functions of saturated fatty acids. Lipids 2010, 45, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Contreras, C.M.; Rodríguez-Landa, J.F.; García-Ríos, R.I.; Cueto-Escobedo, J.; Guillen-Ruiz, G.; Bernal-Morales, B. Myristic Acid Produces Anxiolytic-Like Effects in Wistar Rats in the Elevated Plus Maze. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Vikbjerg, A.F.; Xu, X. Enzymatic modification of phospholipids for functional applications and human nutrition. Biotechnol. Adv. 2005, 23, 203–259. [Google Scholar] [CrossRef] [PubMed]
- Vikbjerg, A.F.; Mu, H.; Xu, X. Lipase-catalyzed acyl exchange of soybean phosphatidylcholine in n-Hexane: A critical evaluation of both acyl incorporation and product recovery. Biotechnol. Prog. 2005, 21, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; No, D.S.; Kim, B.H.; Garcia, H.S.; Kim, Y.; Kim, I.H. Immobilized phospholipase A1-catalyzed modification of phosphatidylcholine with n-3 polyunsaturated fatty acid. Food Chem. 2014, 157, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, A.; Gładkowski, W.; Grudniewska, A. Lipase-catalyzed transesterification of egg-yolk phophatidylcholine with concentrate of n-3 polyunsaturated fatty acids from cod liver oil. Molecules 2017, 22, 1771. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, J.F.; Yang, B.; Li, D.M.; Wang, Y.H.; Wang, W.F. Production of structured phosphatidylcholine with high content of DHA/EPA by immobilized phospholipase A1-catalyzed transesterification. Int. J. Mol. Sci. 2014, 15, 15244–15258. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.; Feng, X.; Shi, N.; Ma, X.; Lin, H.; Han, Y. Immobilized phospholipase A1-catalyzed acidolysis of phosphatidylcholine from Antarctic krill (Euphausia superba) for docosahexaenoic acid enrichment under supercritical conditions. J. Mol. Catal. B Enzym. 2016, 126, 46–55. [Google Scholar] [CrossRef]
- Chojnacka, A.; Gładkowski, W.; Gliszczyńska, A.; Niezgoda, N.; Kiełbowicz, G.; Wawrzeńczyk, C. Synthesis of structured phosphatidylcholine containing punicic acid by the lipase-catalyzed transesterification with pomegranate seed oil. Catal. Commun. 2016, 75, 60–64. [Google Scholar] [CrossRef]
- Niezgoda, N.; Gliszczyńska, A.; Gładkowski, W.; Chojnacka, A.; Kiełbowicz, G.; Wawrzeńczyk, C. Production of concentrates of CLA obtained from sunflower and safflower and their application to the lipase-catalyzed acidolysis of egg yolk phosphatidylcholine. Eur. J. Lipid Sci. Technol. 2016, 118, 1566–1578. [Google Scholar] [CrossRef]
- Shanker Kaki, S.; Ravinder, T.; Ashwini, B.; Rao, B.V.S.K.; Prasad, R.B.N. Enzymatic modification of phosphatidylcholine with n-3 PUFA from silkworm oil fatty acids. Grasas y Aceites 2014, 65, e021. [Google Scholar] [CrossRef]
- Park, C.W.; Kwon, S.J.; Han, J.J.; Rhee, J.S. Transesterification of phosphatidylcholine with eicosapentaenoic acid ethyl ester using phospholipase A 2 in organic solvent. Biotechnol. Lett. 2000, 22, 147–150. [Google Scholar] [CrossRef]
- Doig, S.D.; Diks, R.M.M. Toolbox for exchanging constituent fatty acids in lecithins. Eur. J. Lipid Sci. Technol. 2003, 105, 359–367. [Google Scholar] [CrossRef]
- Mustranta, A.; Forsell, P.; Aura, A.M.; Suortti, T.; Poutanen, K. Modification of phospholipids with lipases and phospholipases. Biocatal. Biotransform. 1994, 9, 181–194. [Google Scholar] [CrossRef]
- Chojnacka, A.; Gładkowski, W.; Kiełbowicz, G.; Gliszczyńska, A.; Niezgoda, N.; Wawrzeńczyk, C. Lipase-catalyzed interesterification of egg-yolk phosphatidylcholine and plant oils. Grasas y Aceites 2014, 65, e053. [Google Scholar] [CrossRef]
- Werbovetz, K.A.; Englund, P.T. Lipid metabolism in Trypanosoma brucei: Utilization of myristate and myristoyllysophosphatidylcholine for myristoylation of glycosyl phosphatidylinositols. Biochem. J. 1996, 318, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.T.; Tran, K.; Pierce, G.N.; Chan, A.C.; Karmin, O.; Choy, P.C. Lysophosphatidylcholine Stimulates the Release of Arachidonic Acid in Human Endothelial Cells. J. Biol. Chem. 1998, 273, 6830–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chojnacka, A.; Gładkowski, W.; Kiełbowicz, G.; Wawrzeńczyk, C. Enzymatic enrichment of egg-yolk phosphatidylcholine with α-linolenic acid. Biotechnol. Lett. 2009, 31, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Virto, C.; Adlercreutz, P. Lysophosphatidylcholine synthesis with Candida antarctica lipase B (Novozym 435). Enzyme Microb. Technol. 2000, 26, 630–635. [Google Scholar] [CrossRef]
- Anderson, E.M.; Larsson, K.M.; Kirk, O. One biocatalyst—Many applications: The use of Candida antarctica B-lipase in organic synthesis. Biocatal. Biotransform. 1998, 16, 181–204. [Google Scholar] [CrossRef]
- Muralidhar, R.V.; Chirumamilla, R.R.; Marchant, R.; Ramachandran, V.N.; Ward, O.P.; Nigam, P. Understanding lipase stereoselectivity. World J. Microbiol. Biotechnol. 2002, 18, 81–97. [Google Scholar] [CrossRef]
- Kirk, O.; Christensen, M.W. Lipases from Candida antarctica: Unique biocatalysts from a unique origin. Org. Process Res. Dev. 2002, 6, 446–451. [Google Scholar] [CrossRef]
- Heldt-Hansen, H.P.; Ishii, M.; Patkar, S.A.; Hansen, T.T.; Eigtved, P. A New Immobilized Positional Nonspecific Lipase for Fat Modification and Ester Synthesis. In Biocatalysis in Agricultural Biotechnology; Whitaker, J.R., Sonnet, P.E., Eds.; ACS Publications: Washington, DC, USA, 1989; pp. 158–172. ISBN 9780841215719. [Google Scholar]
- Adlercreutz, D.; Budde, H.; Wehtje, E. Synthesis of phosphatidylcholine with defined fatty acid in the sn-1 position by lipase-catalyzed esterification and transesterification reaction. Biotechnol. Bioeng. 2002, 78, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.R.C.; Vijeeta, T.; Karuna, M.S.L.; Rao, B.V.S.K.; Prasad, R.B.N. Lipase-catalyzed preparation of palmitic and stearic acid-rich phosphatidylcholine. J. Am. Oil Chem. Soc. 2005, 82, 727–730. [Google Scholar] [CrossRef]
- Vikbjerg, A.F.; Peng, L.; Mu, H.; Xu, X. Continuous production of structured phospholipids in a packed bed reactor with lipase from Thermomyces lanuginosa. J. Am. Oil Chem. Soc. 2005, 82, 237–242. [Google Scholar] [CrossRef]
- Kim, J.H.; Yoon, S.H. Effects of organic solvents on transesterification of phospholipids using phospholipase A2 and lipase. Food Sci. Biotechnol. 2014, 23, 1207–1211. [Google Scholar] [CrossRef]
- Gan, L.J.; Wang, X.Y.; Yang, D.; Zhang, H.; Shin, J.A.; Hong, S.T.; Park, S.H.; Lee, K.T. Emulsifying properties of lecithin containing different fatty acids obtained by immobilized lecitase ultra-catalyzed reaction. J. Am. Oil Chem. Soc. 2014, 91, 579–590. [Google Scholar] [CrossRef]
- Vikbjerg, A.F.; Mu, H.; Xu, X. Parameters affecting incorporation and by-product formation during the production of structured phospholipids by lipase-catalyzed acidolysis in solvent-free system. J. Mol. Catal. B Enzym. 2005, 36, 14–21. [Google Scholar] [CrossRef]
- Nandi, S. Enzymatic Synthesis and Characterization of Modified Phospholipids using Decanoic Acid. Int. J. Biotechnol. Biochem. 2017, 13, 31–38. [Google Scholar]
- Hama, S.; Miura, K.; Yoshida, A.; Noda, H.; Fukuda, H.; Kondo, A. Transesterification of phosphatidylcholine in sn-1 position through direct use of lipase-producing Rhizopus oryzae cells as whole-cell biocatalyst. Appl. Microbiol. Biotechnol. 2011, 90, 1731–1738. [Google Scholar] [CrossRef] [PubMed]
- Egger, D.; Wehtje, E.; Adlercreutz, P. Characterization and optimization of phospholipase A2 catalyzed synthesis of phosphatidylcholine. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1997, 1343, 76–84. [Google Scholar] [CrossRef]
- Ghosh, M.; Bhattacharyya, D.K. Soy lecithin-monoester interchange reaction by microbial lipase. J. Am. Oil Chem. Soc. 1997, 74, 761–763. [Google Scholar] [CrossRef]
- Kiełbowicz, G.; Gładkowski, W.; Chojnacka, A.; Wawrzeńczyk, C. A simple method for positional analysis of phosphatidylcholine. Food Chem. 2012, 135, 2542–2548. [Google Scholar] [CrossRef] [PubMed]
- Gładkowski, W.; Chojnacka, A.; Kiełbowicz, G.; Trziszka, T.; Wawrzeńczyk, C. Isolation of Pure Phospholipid Fraction from Egg Yolk. J. Am. Oil Chem. Soc. 2012, 89, 179–182. [Google Scholar] [CrossRef]
Scheme 1.
Lipase-catalyzed incorporation of myristic acid into egg-yolk phosphatidylcholine (PC) with partial hydrolysis of modified PC as a side reaction. MA—myristic acid; TMA—trimyristin; LPC—lysophosphatidylcholine.
Scheme 1.
Lipase-catalyzed incorporation of myristic acid into egg-yolk phosphatidylcholine (PC) with partial hydrolysis of modified PC as a side reaction. MA—myristic acid; TMA—trimyristin; LPC—lysophosphatidylcholine.

Figure 1.
Time courses of different lipases-catalyzed acidolysis (- - -) and interesterification (―) between egg-yolk phosphatidylcholine (PC) and myristic acid or trimyristin. Reaction condition: temperature, 50–52 °C; PC/acyl donor molar ratio, 1/3; lipase dosage, 30%; solvent, hexane. RML—Rhizomucor miehei lipase; TLL—Thermomyces lanuginosus lipase; CALA—Candida Antarctica.
Figure 1.
Time courses of different lipases-catalyzed acidolysis (- - -) and interesterification (―) between egg-yolk phosphatidylcholine (PC) and myristic acid or trimyristin. Reaction condition: temperature, 50–52 °C; PC/acyl donor molar ratio, 1/3; lipase dosage, 30%; solvent, hexane. RML—Rhizomucor miehei lipase; TLL—Thermomyces lanuginosus lipase; CALA—Candida Antarctica.

Figure 2.
Effect of substrate molar ratio on the incorporation of myristic acid into egg-yolk PC and isolated yield of modified PC in acidolysis and interesterification (reaction condition: temperature, 50–52 °C; lipase dosage, 30%; solvent, hexane; 48 h). MA—myristic acid; TMA—trimyristin.
Figure 2.
Effect of substrate molar ratio on the incorporation of myristic acid into egg-yolk PC and isolated yield of modified PC in acidolysis and interesterification (reaction condition: temperature, 50–52 °C; lipase dosage, 30%; solvent, hexane; 48 h). MA—myristic acid; TMA—trimyristin.

Figure 3.
Effect of enzyme dosage on the incorporation of myristic acid into egg-yolk PC and isolated yield of modified PC in acidolysis and interesterification (reaction condition: temperature, 50–52 °C; PC/acyl donor molar ratio, 1/3; solvent, hexane; 48 h).
Figure 3.
Effect of enzyme dosage on the incorporation of myristic acid into egg-yolk PC and isolated yield of modified PC in acidolysis and interesterification (reaction condition: temperature, 50–52 °C; PC/acyl donor molar ratio, 1/3; solvent, hexane; 48 h).

Figure 4.
Effect of organic solvent on the incorporation of myristic acid into egg-yolk PC and isolated yield of modified PC in acidolysis and interesterification (reaction condition: temperature, 50–52 °C; PC/acyl donor molar ratio, 1/3; lipase dosage, 30%; 48 h). DIPE—diisopropyl ether.
Figure 4.
Effect of organic solvent on the incorporation of myristic acid into egg-yolk PC and isolated yield of modified PC in acidolysis and interesterification (reaction condition: temperature, 50–52 °C; PC/acyl donor molar ratio, 1/3; lipase dosage, 30%; 48 h). DIPE—diisopropyl ether.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fatty acid (FA) composition (wt%) of substrates used in lipase-catalyzed transesterifications of egg-yolk phosphatidylcholine (PC) with trimyristin (TMA) and myristic acid (MA).
Table 1.
Fatty acid (FA) composition (wt%) of substrates used in lipase-catalyzed transesterifications of egg-yolk phosphatidylcholine (PC) with trimyristin (TMA) and myristic acid (MA).
| FA | TMA | MA | Native PC | Modified PC 2 | ||||
|---|---|---|---|---|---|---|---|---|
| Total | sn-1 | sn-2 | Total | sn-1 | sn-2 | |||
| C14:0 | 83.2 ± 0.21 1 | 92.3 ± 0.31 | - | - | - | 44.0 ± 0.14 | 80.5 ± 0.25 | 7.1 ± 0.11 |
| C16:0 | 6.1 ± 0.45 | 6.2 ± 0.11 | 33.2 ± 0.11 | 64.1 ± 0.22 | 4.4 ± 0.04 | 8.3 ± 0.05 | 12.1 ± 0.11 | 4.3 ± 0.07 |
| C16:1 | - | - | 1.2 ± 0.05 | 1.1 ± 0.12 | 1.5 ± 0.05 | 1.1 ± 0.03 | 1.0 ± 0.07 | 1.7 ± 0.12 |
| C18:0 | - | - | 15.7 ± 0.05 | 29.2 ± 0.55 | 2.7 ± 0.09 | 2.7 ± 0.11 | 2.5 ± 0.05 | 2.7 ± 0.15 |
| C18:1 | 5.3 ± 0.05 | 1.1 ± 0.02 | 29.0 ± 0.04 | 4.6 ± 0.33 | 57.1 ± 0.32 | 23.9 ± 0.04 | 2.6 ± 0.12 | 45.5 ± 0.44 |
| C18:2 | - | - | 15.3 ± 0.02 | 1.0 ± 0.05 | 29.8 ± 0.55 | 14.8 ± 0.21 | 1.0 ± 0.04 | 29.3 ± 0.55 |
| C20:4 | - | - | 3.0 ± 0.01 | - | 4.5 ± 0.06 | 5.2 ± 0.22 | 0.3 ± 0.05 | 9.4 ± 0.05 |
| unidentified | 5.4 ± 0.08 | 0.4 ± 0.03 | - | - | - | - | - | - |
1 Data are presented as mean ± SD of two independent analysis; 2 Reaction conditions: enzyme Rhizomucor miehei lipase (RML)—load 30% (w/w), 1/5 PC/TMA molar ratio, temperature 50–52 °C, hexane, 48 h.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chojnacka, A.; Gładkowski, W. Production of Structured Phosphatidylcholine with High Content of Myristic Acid by Lipase-Catalyzed Acidolysis and Interesterification. Catalysts 2018, 8, 281. https://doi.org/10.3390/catal8070281
AMA Style
Chojnacka A, Gładkowski W. Production of Structured Phosphatidylcholine with High Content of Myristic Acid by Lipase-Catalyzed Acidolysis and Interesterification. Catalysts. 2018; 8(7):281. https://doi.org/10.3390/catal8070281
Chicago/Turabian StyleChojnacka, Anna, and Witold Gładkowski. 2018. "Production of Structured Phosphatidylcholine with High Content of Myristic Acid by Lipase-Catalyzed Acidolysis and Interesterification" Catalysts 8, no. 7: 281. https://doi.org/10.3390/catal8070281
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.