Morphological, Physiological, and Genetic Responses to Salt Stress in Alfalfa: A Review



1
Crop Development Center of Department of Plant Sciences, University of Saskatchewan, 51 Campus Drive, Saskatoon, SK S7N 5A8, Canada
2
Plant Gene Resources of Canada, Saskatoon Research and Development Centre, Agriculture and Agri-Food Canada, 107 Science Place, Saskatoon, SK S9H 3X2, Canada
*
Author to whom correspondence should be addressed.
Agronomy 2020, 10(4), 577; https://doi.org/10.3390/agronomy10040577
Submission received: 9 March 2020
/
Revised: 14 April 2020
/
Accepted: 14 April 2020
/
Published: 17 April 2020
(This article belongs to the Special Issue Breeding and Genetics of Forages for Semi-Arid and Arid Rangelands)
Abstract
:Alfalfa (Medicago sativa L.) is an important legume forage crop. However, its genetic improvement for salt tolerance is challenging, as alfalfa’s response to salt stress is genetically and physiologically complex. A review was made to update the knowledge of morphological, physiological, biochemical, and genetic responses of alfalfa plants to salt stress, and to discuss the potential of applying modern plant technologies to enhance alfalfa salt-resistant breeding, including genomic selection, RNA-Seq analysis, and cutting-edge Synchrotron beamlines. It is clear that alfalfa salt tolerance can be better characterized, genes conditioning salt tolerance be identified, and new marker-based tools be developed to accelerate alfalfa breeding for salt tolerance.
1. Introduction
Alfalfa (Medicago sativa L.) is a perennial forage legume that belongs to the sub-family of Papilionoideae. Though a diploid form exists, cultivated alfalfa is a predominantly cross-pollinated, tetraploid (2n = 4x = 32) species [1], which originated in southwestern Asia with Iran as the geographic center of origin [2,3]. Alfalfa is an important forage source for livestock industries around the world because of its wide adaptability, high yield, good quality, and resistance to frequent cuttings [3,4]. It can be used for pasture, hay, silage, dehydrated products, seed production, and soil improvement [3,4]. Globally, alfalfa is grown on about 30 million ha [5]. In Canada, more than four million ha of alfalfa is produced either in monoculture or in mixture with grasses [6], while approximately seven million ha is grown in the United States [7]. Alfalfa is a moderately saline-tolerant legume [8]. A number of alfalfa cultivars with improved salt tolerance have been developed using conventional breeding approaches [9]. However, the genetic improvement of salt tolerance in alfalfa is challenging, mainly as the response of alfalfa plants to salt stress is physiologically and genetically complex, because salt tolerance is controlled by multiple genes and involves various biochemical and physiological mechanisms [9].
Soil salinity is one of the most influential stressors that limits agricultural production. Saline soil is characterized by an excess concentration of soluble salts (chloride; sulfate; and carbonate of sodium, calcium, magnesium, potassium) in the root zone, making it difficult for plants to extract water and nutrients from the soil and causing plant injury [10]. Specifically, soil with an electrical conductivity (EC) of the saturation extract of more than 4 dS m−1 in the root zone at 25 °C with 15% of exchangeable sodium is considered to be saline [11]. Salinization has long been known as a common environmental phenomenon worldwide [12,13] and is becoming a global issue of land degradation, with more prevalence in arid and semi-arid regions [14] as shown in Figure 1. More than 6% of the world’s total land area is salt-affected, either by salinity (397 million hectares) or by the associated conditions of sodicity (434 million hectares) [15]. For example, salinization has affected the agricultural productivity of more than 10 million ha in the Great Plains of North America [16] with 6 million ha of agricultural land in the Canadian Prairies [17,18]. Due to salt build-up over time, the Food and Agricultural Organization (FAO) has estimated that 0.25–0.50 million ha of irrigated lands are becoming unsuitable for cultivation annually [19]. The development of crop cultivars resistant to soil salinity is one of the most effective strategies for maintaining sustainable crop production.
Understanding the mechanisms conferring salt tolerance and identifying heritable traits for screening for improved salt tolerance are crucial for alfalfa breeding. It is also important to identify and characterize the genes responsible for salt tolerance in alfalfa for the development of molecular markers for precise screening in breeding and genetic improvement. Thus, an attempt is made here to review research on the morphological, physiological, biochemical, and genetic responses of alfalfa plants to salt stress. We also discuss the potential of applying modern technologies to enhance alfalfa salt-resistant breeding, including genomic selection, RNA-Seq analysis, and cutting-edge Synchrotron beamlines.
2. Effect of Salt Stress on Morphology, Growth, Forage Yield and Nutritive Value
Salt stress reduces plant growth by influencing turgor, photosynthesis, and the activity of specific enzymes [21]. The influence can occur in two phases; the first phase is governed by the osmotic effect due to high salt concentration in root zones, whereas the second phase is governed by toxic effects due to high salt accumulation in leaf tissues [22] (Figure 2). The presence of high root zone salt concentration causes higher osmotic pressure in soil solution than in plant cells, reducing the ability of plants to uptake water and essential minerals like potassium and calcium [23]. In severe salt stress, the soil solution becomes hyper-osmotic, causing the root cells to lose water, which results in severe wilting or plant senescence [24]. Osmotic stress initially reduces leaf growth and eventually causes a reduction in shoot development and reproductive growth [25], primarily due to the water deficit in plant tissues [24]. Salt stress induces decreased photosynthetic rate due to osmotic stress-induced partial stomatal closure [25]. Sodium ions absorbed by plant roots, if present at a high concentration in cytosol, can be harmful to the plant [26]. Since sodium and potassium ions are both monovalent cations, they compete for uptake by the plant under fully hydrated saline conditions [27] resulting in a deficiency in potassium, an essential macronutrient required for normal metabolic function [23,28]. Increased concentrations of sodium and chloride ions in the cytoplasm can disrupt cellular processes, causing damage to photosynthetic apparatus as well as the dehydration of cells [25,29]. Therefore, limiting excess amounts of sodium in cytosol and increasing the cellular potassium to sodium ratio are crucial for salt tolerance [30,31]. This means the maintenance of a regular photosynthetic rate and stable K+/Na+ ratio are important traits for salt-tolerant alfalfa cultivars. The development of molecular markers should target the specific traits mentioned above to provide informative markers.
Smith [32] described three different growth stages at which alfalfa plants may be affected by salinity: germination, seedling growth, and mature plant growth. The seed germination and seedling stages of alfalfa are highly sensitive to salt stress [33]. Though many soluble salts exist in the natural environment, the majority of research studies on alfalfa salt tolerance used a NaCl solution, with few studies evaluating the response of alfalfa biomass production under a mixture of different salts [34,35]. Alfalfa seed germination is more susceptible to CaCl2 and NaCl salts and more resistant to KCl [36]. Salt stress can significantly reduce the germination and seed vigor of alfalfa [36,37]. The effect of Na2SO4 solution on alfalfa plants from emergence to maturity was examined and it was found that the relative emergence (%) decreased dramatically at 12.7 dS m−1, with no surviving plants at 30 dS m−1 [38]. In another study, a mixture of CaCl2, NaCl, MgSO4, and Na2SO4 was used to obtain the required EC levels to evaluate alfalfa seedling emergence [16]. In this study, alfalfa seedlings emerged equally under the 1.5 dS m−1 and 8.0 dS m−1 treatments, but the emergence decreased by 3–30% at 15.6 dS m−1, except for a few tolerant cultivars [16]. In the past, many breeding efforts have been targeted to improve the seed germination of alfalfa under salt stress, but the selection for salt tolerance should focus on the whole life cycle of alfalfa rather than focusing on a particular growth stage, as there is no clear correlation between seed germination to post germination performance [39,40].
The shoot growth of alfalfa had been found to be more adversely affected by salinity than root growth [41]. A study on 15 alfalfa populations under salt stress imposed by a mixture of MgSO4, Na2SO4, CaCl2, NaCl, and KCl revealed that root mass per plant at 18.3 dS m−1 and 24.5 dS m−1 EC was reduced by 18% and 49%, while the shoot mass was reduced by 50% and 73%, respectively [35]. The average shoot mass of alfalfa during the first, first + second, and first + second + third cuts decreased by 50%, 44%, and 38% at 8.0 dS m−1 (chloride and sulfate salt solution) and by 80%, 73%, and 67% at 15.6 dS m−1, respectively [16]. Salt stress in alfalfa caused a reduction in the relative growth rate [42,43]. Moderate NaCl salt stress (9 ± 0.2 dS m−1) reduced the plant height, leaf, and stem masses of alfalfa by 32%, 34%, and 35%, respectively [44]. Sibole et al. [45] reported that stem and petiole growth was sensitive to NaCl salt stress in two different salt-tolerant Mediterranean Medicago species. As compared to the salt stress imposed by NaCl, other saline solutions containing Ca2+ and K+ ions not only have less toxic effects on plants, but also they can mitigate the negative effects on a plant under salt stress [46,47].
Though a reduction in growth rate and shoot mass were common, high genetic diversity existed among alfalfa populations under salt stress [16,35,48], suggesting an adequate genetic variation for further selection for improved salt tolerance. Lei et al. [49] compared the performance of seven alfalfa cultivars under NaCl stress and found that the growth rate of salt-tolerant cultivars was less affected by a high salt stress of ~50 dS m−1. However, in another study, no significant variation in both shoot and root biomasses was observed between salt-tolerant and intolerant alfalfa cultivars under NaCl stress [50]. Differences among studies might be because of the genetic backgrounds of the alfalfa, as the studies used different salt-tolerant cultivars.
The forage nutritive value was affected by the salinity and varied among the cultivars [51]. Salt stress up to 20 dS m−1 increased the leaf-to-stem ratio of alfalfa and its crude protein content [52] and reduced acid detergent fiber (ADF) and neutral detergent fiber (NDF) [51,53]. The reduction in stem height may be the main factor causing changes in alfalfa nutritive value, as it increases the leaf-to-stem ratio.
3. Effect of Salt Stress on Physiological Responses in Alfalfa
The mechanisms underlying the ability of alfalfa seedlings to tolerate salt stress are complex processes involving photosynthesis, detoxifying, antioxidant, secondary metabolism, and ion transport [54]. Salinity alters photosynthetic pigments (i.e., chlorophyll content), therefore it reduces the maximum photochemical efficiency of alfalfa genotypes [55,56] and increases the dark respiration rate in alfalfa [43]. Salinity reduces membrane stability, relative water content, and growth rates and increases lipid peroxidation, proline, and hydrogen peroxide (H2O2) contents in the leaf tissue of alfalfa [57]. In fact, studies have shown that an increased level of proline content is associated with improved salt tolerance [41,58]. In addition, effective osmoregulation in salt-tolerant alfalfa cultivars is also associated with increased levels of sucrose and pinitol in leaves and a high accumulation of starch in roots [50]. The pinitol accumulation is a characteristic of halophytic plants growing in a saline environment. Anower et al. [59] characterized the physiological traits of two alfalfa half-sib (HS) families, HS-A and HS-B, selected for improved salt tolerance. Under salt treatment, HS-B showed a greater leaf number (72%) and stem length (44%) and HS-A displayed better leaf production (84%) compared to the unselected initial population. This improved growth was associated with a 208% and 78% greater accumulation of chlorophyll content in HS-B and HS-A, respectively. An increase in protein concentration in salt-tolerant alfalfa cultivars had been reported in previous studies [51,53]; it may be associated with an increase in chlorophyll and enzyme activities in the leaves. It is common for plant breeders to measure the plant vigor and shoot mass to determine the performance of alfalfa under salt stress. However, this analysis could be enhanced by the determination of leaf-to-stem ratio and certain physiological traits such as chlorophyll content and protein concentration.
4. Effect of Salt Stress on Oxidative Stress and Anti-Oxidative Activities
Salinity induces oxidative stress in plants at the sub-cellular level [60]. Salt stress increases the accumulation of superoxide radicals (O2−) and hydrogen peroxide (H2O2) in cell compartments including chloroplast and mitochondria [60]. Therefore, lipid peroxidation and protein oxidation occur in the apoplastic space. An increase in peroxidase (POX) reduces H2O2 to water using various substrates as electron donors in salt-tolerant alfalfa [61,62], suggesting that the analysis of antioxidant enzymes could be useful in understanding the salt tolerance of alfalfa. During seed germination and seedling growth, increased salt stress also increases the activities of superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase activity (APX)—and the increase is higher in salt-tolerant alfalfa cultivars [62,63]. Under salt stress, the unsaturated fatty acids of plant membranes are decomposed to malondialdehyde (MDA), and the rate of lipid peroxidation in terms of MDA can be used as a biochemical indicator to evaluate salt-tolerant cultivars [30,64]. The salt tolerance of alfalfa is also improved by melatonin application, which acts as an antioxidant in scavenging H2O2 and enhances antioxidant enzymes’ activities [65]. Under salt stress, tolerant alfalfa shows less severe cell membrane damage and lower accumulation of reactive oxygen species (ROS) than salt sensitive types [66].
5. Effect of Salt Stress on Ion Uptake in Alfalfa Plants
Under salt stress, about 98% of the ions in soil solutions are excluded from the root in most plants, while the rest are transported to shoot tissues [22]. To prevent tissue ion toxicity, the ion exclusion mechanism can restrict excessive ion transport from root to shoot. The ion exclusion mechanism includes: (1) the minimal uptake of particular toxic ions by the root and maximization of ion efflux to the soil, (2) restricting excessive ion loading into the xylem, (3) increasing the ion retrieval from xylem to other tissues like the root and stem, and (4) increasing the ion transport from shoot to root through the phloem [67]. High concentrations of sodium and chloride ions in leaves generally lead to leaf senescence. However, the tissue tolerance to such a stress is genotype-specific [24]. Tolerance to high concentrations of sodium and chloride ions in leaves can be achieved by the intercellular partitioning of ions, thus avoiding their accumulation in the photosynthetic organelles [25]. For example, salt-tolerant species can sequestrate ions in the cell vacuoles in leaves [25,68]. Thus, the comparison of leaf injury scores under high salt stress can be used as a morphological marker for identifying salt-tolerant genotypes in early phases with similar LD50 values (median lethal dose to kill 50% of population) [33].
Salt stress can lead to nutritional imbalances due to competitive absorption and translocation of elements, which may result in reduced physiological activity [69]. This might in part explain the plant growth reduction and low shoot mass of alfalfa at high salt stress, as discussed in the previous section. The high concentrations of sodium and chloride ions in soil solutions may decrease the calcium, potassium, and magnesium activities in alfalfa [70] and also decrease nitrogen accumulation [71,72]. High salt stress increases the concentrations of sodium, total sulfur, chloride, magnesium, and phosphorus, and decreases the concentrations of potassium and calcium in the shoots of alfalfa genotypes [38]. The maintenance of the K+/Na+ ratio is necessary through Na+ exclusion from the root epidermis, but NaCl-induced K+ efflux from the root causes the deficiency of macronutrients (nitrogen and potassium), reducing plant production and productivity.
Knowledge of the biochemical composition of plant tissues is crucial for salt tolerance studies, as cutting-edge synchrotron-based approaches can help plant scientists to quantify the biochemical compounds and image their structures with minimal sample modifications [73]. The synchrotron is a powerful facility that accelerates charged particles such as electrons in a large ring-like trajectory at relativistic (near-light) speed [73], where the energy of the electron generates light ranging from infrared to soft and hard X-rays at high intensities [74]. In recent years, synchrotron techniques have been employed to study a number of biotic and abiotic stresses, such as the identification of heat-tolerant field pea (Pisum sativum L.) genotypes [75,76], drought tolerance traits in spring wheat (Triticum aestivum L.) [77], and Fusarium head blight tolerance in wheat [78,79]. Synchrotron beamlines were also applied to study the accumulation and distribution of organic compounds and salt ions in leaf, stem, and root tissues of alfalfa cultivars with different salt tolerances [80]. A comprehensive study on ion localization in sub-cellular levels using synchrotron beamlines would provide further insights into salt tolerance in alfalfa.
6. Proteome and Transcriptomic Analyses
Several transcriptome-based and proteome studies have been conducted to understand salinity stress in alfalfa [49,54,66,81,82,83,84,85,86,87]. After a three-day salt treatment, a proteomic study on the roots of two-week-old seedlings found 83 differentially expressed proteins in alfalfa cultivars with contrasting tolerances to salinity (Table 1) [81]. These proteins are involved in ion homeostasis, protein turnover and signaling, protein folding, cell wall components, carbohydrate and energy metabolism, reactive oxygen species (ROS) regulation and detoxification, and purine and fatty acid metabolisms. Proteins such as peroxidase, protein disulfide-isomerase, nicotinamide adenine dinucleotide (NAD) synthetase, and isoflavone reductase were significantly up-regulated in the salt-tolerant alfalfa cultivar “nonomu” [81]. The proteomic analysis of 30-day-old alfalfa seedling roots treated with ~30 dS m−1 NaCl for 8 h found 93 and 30 differentially expressed proteins in salt-tolerant alfalfa (“Zhongmu-1”) and salt-sensitive Medicago truncatula (“Jemalong A17”), respectively (Table 1) [82]. These proteins primarily play roles in molecule binding and catalytic activities. Xiong et al. [54] identified 26 (shoot) and 35 (root) differentially abundant proteins in salt-stressed alfalfa compared to alfalfa that had experienced no salt treatment. Similarly, the proteomic analysis of osmo-primed alfalfa seeds that germinated under salinity stress found they contained 94 proteins with different responses to salt treatments [86]. Based on their functions, the proteins were classified as protein destination and storage (seed storage proteins and small heat-shock proteins), cell growth/division (late embryogenesis abundant proteins and seed maturation proteins), metabolism (methionine synthase, cysteine synthase and haem oxygenase), and disease and defense (glutathione S-transferase).
Transcriptomic approaches have been employed for alfalfa to understand the gene expression associated with salt stress (Table 1) [49,66,83,84,85,87]. Eighty-two unique transcripts were found from the salt-stressed seedlings of alfalfa by sampling at different time intervals from 10 min to 24 h, including 24% that were proteins related to plant metabolism and 9% that were related to abiotic stress [87]. Arshad et al. [88] found that the over-expression of microRNA156 in alfalfa resulted in increased biomass production, stem number, concentration of crude protein, and reduced uptake of Na+ under salt stress. In another transcriptome study, there were 876 and 1303 differentially expressed genes (DEGs) under salt stress in the root tissues of one-week-old seedlings in salt-intolerant and tolerant alfalfa genotypes respectively, with 604 DEGs specific to the salt-tolerant type [83]. Similarly, an RNA-Seq analysis displayed 2237 and 1125 DEGs between “Zhongmu-1” (salt-tolerant) and “Xingjiang Daye” (salt-intolerant) in the presence and absence of salt stress, among which were many genes that are involved in stress-related pathways (Table 1) [49]. After a salt treatment, the number of DEGs in “Xingjiang Daye” (19,373 DEGs) compared with the control treatment was about four times that of “Zhongmu-1” (4833 DEGs). Compared with “Xingjiang Daye”, “Zhongmu-1” maintained a more stable expression of genes related to ROS, calcium pathways, phytohormone biosynthesis, and Na+/K+ transport [49]. The transcriptome responses of salt-tolerant (“211609”) and salt-intolerant (“Xinjiang Daye”) alfalfa cultivars revealed significantly higher expression levels of NHX1, ZFG, CBF4, and HSP23 genes in “211609” than in “Xinjiang Daye” [66]. In addition, a transcriptomic analysis of alfalfa roots under ~25 dS m−1 NaCl identified 8861 NaCl-regulated DEGs in alfalfa [84]. These DEGs were categorized into 13 gene ontology categories: oxidoreductase activity, oxidation-reduction process, structural constituent of cytoskeleton, hydrolase activity, carbohydrate metabolic process, negative regulation of catalytic activity, polysaccharide catabolic process, iron ion binding, transmembrane transporter activity, cytoskeleton, trehalose biosynthetic process, protein polymerization, and ion homeostasis [84]. In previous transcriptomic studies, an individual alfalfa genotype was sampled as a replicate for RNA-Seq analysis. As alfalfa is generally seeded as a synthetic population in a field, it may be more suitable to use a group of genotypes as a replicate for a transcriptomic study. Therefore, 100 alfalfa genotypes were divided into four independent replicates to represent an alfalfa population and researchers found that 50% of DEGs were down-regulated in a salt-intolerant population [85]. The functions of the genes down-regulated in the salt-intolerant alfalfa cultivar were grouped into cell wall structural components, lipids, secondary metabolism, auxin and ethylene hormones, transport, signaling, pathogenesis response, and abiotic stress [85]. The transcriptome analysis identified many annotated sequences homologous to genes involved in osmolyte synthesis (beta-amylase, fructose-1, 6-bisphosphate, aldolase, and sucrose synthase) and ion homeostasis (H+-PPase, cation/H+ exchanger3, Ca-related channel, sodium symporter, nitrate and K+ channels, p translocator, and metal transporters) [49,85,87]. In summary, a large number of candidate genes responsible for salt tolerance in alfalfa have been identified in various studies, and these candidate genes can be used as a baseline for further genetic analyses of salt tolerance in alfalfa. However, it is essential to identify key genes associated with important morphological and physiological traits (i.e., chlorophyll content, sucrose synthase or plant height), and validate them in different alfalfa breeding populations to apply them in salt-tolerant breeding.
7. Breeding for Salt Tolerance
Breeding efforts for salt tolerance in alfalfa have focused on high seed germination under stress to improve stand establishment. There are more than 60 registered alfalfa cultivars with improved salinity tolerance in the USA, representing different fall dormancy categories (Table 2) [89]. The majority of the registered cultivars were selected for salinity tolerance at the germination stage, while only 13 of them were selected for salinity tolerance at mature growth stages. Although certain progresses have been made through conventional plant breeding, the genetic improvement of salt tolerance in alfalfa is low due to several factors. Firstly, alfalfa is polyploidy and outcrossing in nature [90] and genetic studies of its salt tolerance can be complicated and less informative. Secondly, the perennial growth form and low heritability of salt-tolerant traits have further complicated the breeding effort [91]. Thirdly, as previously stated, there is no clear relationship for salt tolerance between the germination stage and post-germination performance [39], indicating a need for a whole life cycle selection from germination to the flowering stage.
A number of studies have evaluated the performance of alfalfa germplasm under salt stress [16,92,93,94]. The genetic variability among nine alfalfa populations in response to sulfate salt revealed a high relative shoot mass of 8 dS m−1 in “Halo” alfalfa, which had a lower seed emergence than that of “Rugged” [16]. Benabderrahim et al. [92] studied the genetic diversity of 36 alfalfa populations from Tunisia using 12 agronomic and physiological traits and identified three salt-tolerant types. Jiang et al. [93] have applied random amplified polymorphic DNA (RAPD) markers and clustered 25 salt-tolerant alfalfa populations into nine clusters, suggesting that salt-tolerant alfalfa germplasm possess a certain genetic diversity which can be utilized in salt-tolerant breeding. Azzam et al. [94] identified a highly salt-tolerant alfalfa population by screening 16 alfalfa populations using RAPD and inter simple sequence repeat markers. Therefore, the available genotypic diversity for salt tolerance in alfalfa provides an opportunity for plant breeders to select and develop superior salt-tolerant cultivars. However, there is still a lack of well-characterized genetic materials and common phenotyping protocols for parental selection. Due to the spatial heterogeneity of soil properties and seasonal variation in rainfall, it is difficult to perform plant screening for salinity tolerance in the field [95]. Thus, early phase screening in controlled environments is a feasible option before testing the advanced lines in the saline field with the use of proper salinity induction.
The success of a breeding program lies in the precise phenotyping of the traits of interest (seed germination, ion transport, osmolyte synthesis, signaling, photosynthesis, and protein synthesis) and correlating them with genes or quantitative trait loci for marker-assisted selection [96]. A comprehensive breeding platform, including high-throughput phenotyping and genotyping, is needed for the efficient improvement of salt tolerance in alfalfa. High-throughput phenotyping contributes directly to the genetic gain by evaluating genetic variation more efficiently. [97]. A hyperspectral imaging platform has the potential for the detection of stress responses as it can capture wavelengths within and beyond the visible spectrum [98]. Hyperspectral cameras detect both spectral and spatial information; each spatially located pixel contains full wavelength (~350–2500 nm) information. Imaging within the visible spectrum (~400–700 nm) can measure the morphological and color properties of plants, while hyperspectral imaging can also study the radiative properties of plant leaves to facilitate the early detection of abiotic stress [98,99]. Hyperspectral imaging has been successfully applied to study the vegetation indices of crops under salt stress [100,101,102]. The physiological reflectance index and normalized difference vegetation index are useful for the early detection of salinity stress in plants [100,101]. Sytar et al. [102] reviewed the application of hyperspectral imaging as a fast and reliable technique for the detection of quantitative and qualitative changes in plants to evaluate plant variation under salt stress.
Several studies have focused on the development of molecular markers associated with salt tolerance in alfalfa at different growth stages [93,94,103,104,105]. The most significant salt tolerance markers during the germination stage were identified on chromosomes 1, 2, and 4, while the marker located on chromosome 6 overlapped with drought resistance [104,106]. During alfalfa seed germination, a broad-sense heritability of the germination rate was observed at 0.60 at 0.5% (~8 dS m−1 NaCl) salt concentration, which decreased to 0.24 at 0.75% (~12 dS m−1 NaCl) and 0.27 at 1.0% (~17 dS m−1 NaCl) [104]. The broad-sense heritability of leaf chlorophyll content increased from 0.22 at 0 dS m−1 to 0.34 at 8 dS m−1 NaCl [105]. As salt-tolerant traits are genetically complex and multi-gene controlled, the development of genome-wide markers might be useful for conducting a genomic selection based on the predictive breeding values of genotypes. Genomic selection can enhance the genetic gain and reduce the breeding cycle when it increases the selection accuracy [107,108]. The trait prediction accuracy remains generally low in current genomic selection models, even with the aid of dense, genome-wide single nucleotide polymorphism (SNP) markers [108]. As demonstrated by Fu et al. [108], salt tolerance-associated SNP markers in alfalfa could be explored to achieve a more accurate trait-specific prediction in genomic selection. This is feasible, as the application of the RNA-Seq technique in alfalfa salinity tolerance studies is relatively common and a large amount of genomic information is already available. With the development of modern breeding techniques, such as high-throughput phenotyping and genotyping-by-sequencing platforms, genomic selection can enhance the salt-tolerant breeding of alfalfa at a reasonable cost.
Author Contributions
B.B. and S.B. conceived this review paper; S.B. wrote the original manuscript; D.B. reviewed and contributed to the writing the physiological section; B.B. and Y.-B.F. reviewed and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Sciences and Engineering Research Council of Canada and Beef Cattle Research Council of Canada.
Acknowledgments
We would like to thank Anuja Thapa of the School of Environment and Sustainability at the University of Saskatchewan for her technical help during ‘World Salt Map’ development using ArcMap software.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lesins, K.A.; Lesins, I. Genus Medicago (Leguminosae): A Taxogenetic Study; Dr. W. Junk bv Publishers: The Hague, The Netherlands, 1979; p. 228. [Google Scholar]
- Bolton, J.L. Alfalfa Botany, Cultivation and Utilization; Leonard Hill Ltd.: London, UK; Interscience Publishers, Inc.: New York, NY, USA, 1962. [Google Scholar]
- Goplen, B.P.; Baenziger, H.; Bailey, L.D.; Gross, A.T.H.; Hanna, M.R.; Michaud, R.; Richards, K.W.; Waddington, J. Agriculture Canada: Growing and Managing Alfalfa in Canada; Publication 1705/E; Agriculture Canada: Ottawa, ON, Canada, 1982. [Google Scholar]
- Coburn, F.D. The Book of Alfalfa: History, Cultivation and Merits. Its Uses as a Forage and Fertilizer; Orange Judd Co: New York, NY, USA, 1907. [Google Scholar]
- Yuegao, H.; Cash, D. Global status and development trends of alfalfa. In Alfalfa Management. Guide for Ningxia; Cash, D., Ed.; United Nations Food and Agriculture Organization: Beijing, China, 2009; pp. 1–14. [Google Scholar]
- Statistics Canada. Census of Agriculture, Hay and Field Crops. Table 32-10-0416-01. 2016. Available online: http://www.statcan.gc.ca/eng (accessed on 5 February 2020).
- USDA–NASS. Crop Production 2018 Summary. USDA–NASS. 2018. Available online: http://www.nass.usda.gov (accessed on 9 February 2020).
- Maas, E.V.; Hoffman, G.J. Crop salt tolerance-current assessment. J. Irrig. Drain. Div. 1977, 103, 115–134. [Google Scholar]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Szabolcs, I. Salt-Affected Soils; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassas, M. Seven paths to desertification. Desert. Control Bull. 1987, 15, 24–26. [Google Scholar]
- Thomas, D.S.G.; Middleton, N.J. Salinization: New perspectives on a major desertification issue. J. Arid Environ. 1993, 24, 95–105. [Google Scholar] [CrossRef]
- Tanji, K.K. Nature and extent of agricultural salinity. In Agricultural Salinity Assessment and Management; Tanji, K.K., Ed.; American Society Civil Engineers: New York, NY, USA, 1990; pp. 1–13. [Google Scholar]
- FAO. Global Network on Integrated Soil Management for Sustainable Use of Salt-Affected Soils; FAO Land and Plant Nutrition Management Service: Rome, Italy, 2005; Available online: http://www.fao.org/ag/agl/agll/spush (accessed on 11 February 2020).
- Steppuhn, H.; Acharya, S.N.; Iwaasa, A.D.; Gruber, M.; Miller, D.R. Inherent responses to root-zone salinity in nine alfalfa populations. Can. J. Plant Sci. 2012, 92, 235–248. [Google Scholar] [CrossRef]
- Steppuhn, H. What is soil salinity? In Proceedings Soil Salinity Assessment Workshop; Alberta Agriculture: Lethbridge, AB, Canada, 1996; pp. 1–5. [Google Scholar]
- Wiebe, B.H.; Eilers, R.G.; Eilers, W.D.; Brierley, J.A. Application of a risk indicator for assessing trends in dryland salinization risk on the Canadian Prairies. Can. J. Soil Sci. 2007, 87, 213–224. [Google Scholar] [CrossRef]
- Martinez, J.; Manzur, C.L. Overview of salinity problems in the world and FAO strategies to address the problem. In Proceedings of the International Salinity Forum, Riverside, CA, USA, 25–27 April 2005; pp. 311–313. [Google Scholar]
- FAO; IIASA; ISRIC; ISS-CSA; JRC. Harmonized World Soil Database (Version 1.2); FAO: Rome, Italy; IIASA: Laxenburg, Austria, 2008. [Google Scholar]
- Munns, R. A leaf elongation assay detects an unknown growth inhibitor in xylem sap from wheat and barley. Aust. J. Plant Physiol. 1992, 19, 127–135. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 654–663. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, N. Mechanisms of high salinity tolerance in plants. Methods Enzymol. 2007, 428, 419–438. [Google Scholar]
- Schachtman, D.; Liu, W.H. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends Plant Sci. 1999, 4, 282–287. [Google Scholar] [CrossRef]
- Bhandal, I.S.; Malik, C.P. Potassium estimation, uptake, and its role in the physiology and metabolism of flowering plant. Int. Rev. Cytol. 1998, 110, 205–254. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environment: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum wheat roots adapt to salinity remodeling the cellular content of nitrogen metabolites and sucrose. Front. Plant Sci. 2017, 7, 2035. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphaelb, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Smith, S.E. Salinity and the production of alfalfa (Medicago sativa L.). In Handbook of Crop Stress; Pessarakli, M., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1993; pp. 431–448. [Google Scholar]
- Peel, M.D.; Waldron, B.L.; Jensen, K.B.; Chatterton, N.J.; Horton, H.; Dudley, L.M. Screening for salinity tolerance in alfalfa. Crop Sci. 2004, 44, 2049–2053. [Google Scholar] [CrossRef]
- Soltanpour, P.N.; Ippolito, J.A.; Rodriguez, J.B.; Self, J.; Gillaume, M.; Al-Wardy, M.M.; Mathews, D. Chloride versus sulfate salinity effects on alfalfa shoot growth and ionic balance. Soil Sci. Soc. Am. J. 1999, 63, 111–116. [Google Scholar] [CrossRef]
- Cornacchione, M.V.; Suarez, D.L. Evaluation of alfalfa (Medicago sativa L.) populations’ response to salinity stress. Crop Sci. 2017, 57, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Azhdari, G.; Tavili, A.; Zare, M.A. Effects of various salts on the germination of two cultivars of Medicago sativa. Front. Agric. China 2010, 4, 63–68. [Google Scholar] [CrossRef]
- Soltani, A.; Khodarahmpour, Z.; Jafari, A.A.; Nakhjavan, S. Selection of alfalfa (Medicago sativa L.) cultivars for salt stress tolerance using germination indices. Afr. J. Biotechnol. 2012, 11, 7899–7905. [Google Scholar]
- Cornacchione, M.V.; Suarez, D.L. Emergence, forage production, and ion relations of alfalfa in response to saline waters. Crop Sci. 2015, 55, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Al-Niemi, T.S.; Campbell, W.F.; Rumbaugh, M.D. Response of alfalfa cultivars to salinity during germination and post-germination growth. Crop Sci. 1992, 32, 976–980. [Google Scholar] [CrossRef]
- Johnson, D.W.; Smith, S.E.; Dobrenz, A.K. Selection for increased forage yield in alfalfa at different NaCl levels. Euphytica 1992, 60, 27–35. [Google Scholar]
- Torabi, M.; Halim, M.R.A. Variation of root and shoot growth and free proline accumulation in Iranian alfalfa ecotypes under salt stress. J. Food Agric. Environ. 2010, 8, 323–327. [Google Scholar]
- Shannon, M.C.; Grieve, C.M.; Francois, L.E. Whole-plant response to salinity. In Handbook of Plant-Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 199–244. [Google Scholar]
- Khavarinejad, R.A.; Chaparzadeh, N. The effects of NaCl and CaCl2 on photosynthesis and growth of alfalfa plants. Photosynthetica 1998, 35, 461–466. [Google Scholar] [CrossRef]
- Valizadeh, M.; Moharamnejad, S.; Ahmadi, M.; Jalaly, H.M. Changes in activity profile of some antioxidant enzymes in alfalfa half-sib families under salt stress. J. Agric. Sci. Technol. 2013, 15, 801–809. [Google Scholar]
- Sibole, J.V.; Cabot, C.; Poschenrieder, C.; Barcelo, J. Ion allocation in two different salt-tolerant Mediterranean Medicago species. J. Plant Physiol. 2003, 160, 1361–1365. [Google Scholar] [CrossRef]
- Hanley, M.E.; Sanders, S.K.D.; Stanton, H.M.; Billington, R.A.; Boden, R. A pinch of salt: Response of coastal grassland plants to simulated seawater inundation treatments. Ann. Bot. 2020, 125, 265–275. [Google Scholar] [CrossRef]
- Tootoonchi, M.; Gettys, L.A. Testing salt stress on aquatic plants: Effect of salt source and substrate. Aquat. Ecol. 2019, 53, 325–334. [Google Scholar] [CrossRef]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.S.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [Google Scholar] [CrossRef]
- Lei, Y.; Xu, Y.; Hettenhausen, C.; Lu, C.; Shen, G.; Zhang, C.; Li, J.; Song, J.; Lin, H.; Wu, J. Comparative analysis of alfalfa (Medicago sativa L.) leaf transcriptomes reveals genotype-specific salt tolerance mechanisms. BMC Plant Biol. 2018, 18, 35. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, A.; Dhont, C.; Bipfubusa, M.; Chalifour, F.P.; Drouin, P.; Beauchamp, C.J. Improving salt stress responses of the symbiosis in alfalfa using salt-tolerant cultivar and rhizobial strain. Appl. Soil Ecol. 2015, 87, 108–117. [Google Scholar] [CrossRef]
- Robinson, P.H.; Grattan, S.R.; Getachew, G.; Grieve, C.M.; Poss, J.A.; Suarez, D.L.; Benes, S.E. Biomass accumulation and potential nutritive value of some forages irrigated with saline-sodic drainage water. Anim. Feed Sci. Technol. 2004, 111, 175–189. [Google Scholar] [CrossRef]
- Al-Khatib, M.; McNeilly, T.; Collins, J. The potential of selection and breeding for improved salt tolerance in lucerne (Medicago sativa L.). Euphytica 1992, 65, 43–51. [Google Scholar] [CrossRef]
- Suyama, H.; Benes, S.E.; Robinson, P.H.; Grattan, S.R.; Grieve, C.M.; Getachew, G. Forage yield and quality under irrigation with saline-sodic drainage water: Greenhouse evaluation. Agric. Water. Manag. 2007, 88, 159–172. [Google Scholar] [CrossRef]
- Xiong, J.; Sun, Y.; Yang, Q.; Tian, H.; Zhang, H.; Liu, Y.; Chen, M. Proteomic analysis of early salt stress responsive proteins in alfalfa roots and shoots. Proteome Sci. 2017, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Smethurst, C.F.; Rix, K.; Garnett, T.; Auricht, G.; Bayart, A.; Lane, P.; Wilson, S.J.; Shabala, S. Multiple traits associated with salt tolerance in lucerne: Revealing the underlying cellular mechanisms. Funct. Plant Biol. 2008, 35, 640–650. [Google Scholar] [CrossRef]
- Shone, M.G.T.; Gale, J. Effects of sodium chloride stress and nitrogen source on respiration, growth and photosynthesis in lucerne (Medicaqo sativa L.). J. Exp. Bot. 1983, 34, 1117–1125. [Google Scholar] [CrossRef]
- Chaparzadeh, N.; Mehrnejad, F. Oxidative markers in five Iranian alfalfa (Medicago sativa L.) cultivars under salinity stress. Iran. J. Plant Physiol. 2013, 3, 793–799. [Google Scholar]
- Campanelli, A.; Ruta, C.; Morone-Fortunato, I.; De Mastro, G. Alfalfa (Medicago sativa L.) clones tolerant to salt stress: In vitro selection. Cent. Eur. J. Biol. 2013, 8, 765–776. [Google Scholar] [CrossRef]
- Anower, R.M.; Mott, I.W.; Peel, M.D.; Wu, Y. Characterization of physiological responses of two alfalfa half-sib families with improved salt tolerance. Plant Physiol. Biochem. 2013, 71, 103–111. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Babakhani, B.; Khavari-Nejad, R.A.; Hassan Sajedi, R.; Fahimi, H.; Saadatmand, S. Biochemical responses of Alfalfa (Medicago sativa L.) cultivars subjected to NaCl salinity stress. Afr. J. Biotechnol. 2011, 10, 11433–11441. [Google Scholar]
- Ashrafi, E.; Razmjoo, J.; Zahedi, M.; Pessarakli, M. Screening alfalfa for salt tolerance based on lipid peroxidation and antioxidant enzymes. Agron. J. 2015, 107, 167–173. [Google Scholar] [CrossRef]
- Wang, X.S.; Han, J.G. Changes of proline content, activity, and active isoforms of antioxidative enzymes in two alfalfa cultivars under salt stress. Agric. Sci. China 2009, 8, 431–440. [Google Scholar] [CrossRef]
- Jain, M.; Tiwary, S.; Gadre, R. Sorbitol-induced changes in various growth and biochemical parameters in maize. Plant Soil Environ. 2010, 56, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Cen, H.; Wang, T.; Liu, H.; Tian, D.; Zhang, Y. Melatonin application improves salt tolerance of alfalfa (Medicago sativa L.) by enhancing antioxidant capacity. Plants 2020, 9, 220. [Google Scholar] [CrossRef] [Green Version]
- Quan, W.L.; Liu, X.; Wang, H.Q.; Chan, Z.L. Physiological and transcriptional responses of contrasting alfalfa (Medicago sativa L.) varieties to salt stress. Plant Cell Tissue Organ Cult. 2016, 126, 105–115. [Google Scholar] [CrossRef]
- Tilbrook, J.; Roy, S.J. Salinity tolerance. In Plant Abiotic Stress, 2nd ed.; Jenks, M.A., Hasewaga, P.M., Eds.; Wiley-Blackwell: New York, NY, USA, 2014; pp. 134–178. [Google Scholar]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995. [Google Scholar]
- Ashrafi, E.; Razmjoo, J.; Zahedi, M. Effect of salt stress on Growth and Ion accumulation of alfalfa (Medicago sativa L.) cultivars. J. Plant Nutr. 2018, 41, 818–831. [Google Scholar] [CrossRef]
- Pessarakli, M.; Huber, T.C.; Nakabayashi, K. Growth response of barley and wheat to salt stress. J. Plant Nutr. 1991, 14, 331–340. [Google Scholar] [CrossRef]
- Khan, M.G.; Silberbush, M.; Lips, S.H. Physiological study on salinity and nitrogen interaction in alfalfa II photosystem and transpiration. J. Plant Nutr. 1994, 17, 669–684. [Google Scholar] [CrossRef]
- Vijayan, P.; Willick, I.R.; Lahlali, R.; Karunakaran, C.; Tanino, K.K. Synchrotron radiation sheds fresh light on plant research: The use of powerful techniques to probe structure and composition of plants. Plant Cell Physiol. 2015, 56, 1252–1263. [Google Scholar] [CrossRef]
- Duncan, W.; Williams, G. Infrared synchrotron radiation from electron storage rings. Appl. Opt. 1983, 22, 2914–2923. [Google Scholar] [CrossRef]
- Jiang, Y.; Lahlali, R.; Karunakaran, C.; Kumar, S.; Davis, A.R.; Bueckert, R.A. Seed set, pollen morphology and pollen surface composition response to heat stress in field pea. Plant Cell Environ. 2015, 38, 2387–2397. [Google Scholar] [CrossRef]
- Lahlali, R.; Jiang, Y.; Kumar, S.; Karunakaran, C.; Liu, X.; Borondics, F.; Hallin, E.; Bueckert, R. ATR-FTIR spectroscopy reveals involvement of lipids and proteins of intact pea pollen grains to heat stress tolerance. Front. Plant Sci. 2014, 5, 747. [Google Scholar] [CrossRef] [Green Version]
- Willick, I.R.; Lahlali, R.; Vijayan, P.; Muir, D.; Karunakaran, C.; Tanino, K.K. Wheat flag leaf epicuticular wax morphology and composition in response to moderate drought stress are revealed by SEM, FTIR-ATR and synchrotron X-ray spectroscopy. Physiol. Plant. 2017, 162, 316–332. [Google Scholar] [CrossRef] [Green Version]
- Lahlali, R.; Karunakaran, C.; Wang, L.; Willick, I.; Schmidt, M.; Liu, X.; Borondics, F.; Forseille, L.; Fobert, P.R.; Tanino, K.; et al. Synchrotron based phase contrast X-ray imaging combined with FTIR spectroscopy reveals structural and biomolecular differences in spikelets play a significant role in resistance to Fusarium in wheat. BMC Plant Biol. 2015, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Lahlali, R.; Kumar, S.; Wang, L.; Forseille, L.; Sylvain, N.; Korbas, M.; Muir, D.; Swerhone, G.; Lawrence, J.R.; Fobert, P.R.; et al. Cell wall biomolecular composition plays a potential role in the host type II resistance to Fusarium head blight in wheat. Front. Microbiol. 2016, 7, 910. [Google Scholar] [CrossRef]
- Bhattarai, S.; Karunakaran, C.; Tanino, K.K.; Fu, Y.B.; Coulman, B.; Biligetu, B. Physiological and biochemical responses of alfalfa (Medicago sativa L.) to salt stress. In Proceedings of the Communicating Innovation in Plant Science, Plant Canada 2019, Guelph, ON, Canada, 7–10 July 2019; p. 158. [Google Scholar]
- Rahman, M.A.; Alam, I.; Kim, Y.G.; Ahn, N.Y.; Heo, S.H.; Lee, D.G.; Liu, G.; Lee, B.H. Screening for salt-responsive proteins in two contrasting alfalfa cultivars using a comparative proteome approach. Plant Physiol. Biochem. 2015, 89, 112–122. [Google Scholar] [CrossRef]
- Long, R.; Li, M.; Zhang, T.; Kang, J.; Sun, Y.; Cong, L.; Gao, Y.; Liu, F.; Yang, Q. Comparative proteomic analysis reveals differential root proteins in Medicago sativa and Medicago truncatula in response to salt stress. Front. Plant Sci. 2016, 7, 424. [Google Scholar] [CrossRef] [Green Version]
- Postnikova, O.A.; Shao, J.; Nemchinov, L.G. Analysis of the alfalfa root transcriptome in response to salinity stress. Plant Cell Physiol. 2013, 54, 1041–1055. [Google Scholar] [CrossRef]
- Luo, D.; Zhou, Q.; Wu, Y.G.; Chai, X.T.; Liu, W.X.; Wang, Y.R.; Yang, Q.C.; Wang, Z.Y.; Liu, Z.P. Full length transcript sequencing and comparative transcriptomic analysis to evaluate the contribution of osmotic and ionic stress components towards salinity tolerance in the roots of cultivated alfalfa (Medicago sativa L.). BMC Plant Biol. 2019, 19, 32. [Google Scholar] [CrossRef] [Green Version]
- Gruber, M.; Xia, J.; Yu, M.; Steppuhn, H.; Wall, K.; Messer, D.; Sharpe, A.; Acharya, S.; Wishart, D.; Johnson, D.; et al. Transcript analysis in two alfalfa salt tolerance selected breeding populations relative to a non-tolerant population. Genome 2017, 60, 104–127. [Google Scholar] [CrossRef]
- Yacoubi, R.; Job, C.; Belghazi, M.; Chaibi, W.; Job, D. Proteomic analysis of the enhancement of seed vigour in osmoprimed alfalfa seeds germinated under salinity stress. Seed Sci. Res. 2013, 23, 99–110. [Google Scholar] [CrossRef]
- Jin, H.; Sun, Y.; Yang, Q.; Chao, Y.; Kang, J.; Jin, H.; Li, Y.; Margaret, G. Screening of genes induced by salt stress from alfalfa. Mol. Biol. Rep. 2010, 37, 745–753. [Google Scholar] [CrossRef]
- Arshad, M.; Gruber, M.Y.; Wall, K.; Hannoufa, A. An insight into microRNA156 role in salinity stress responses of alfalfa. Front. Plant Sci. 2017, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- National Alfalfa and Forage Alliance. Alfalfa Variety Ratings. 2020. Available online: https://www.alfalfa.org/pdf/2020_Alfalfa_Variety_Leaflet.pdf (accessed on 15 January 2020).
- Annicchiarico, P.; Barrett, B.; Brummer, E.C.; Julier, B.; Marshall, A.H. Achievements and challenges in improving temperate perennial forage legumes. Crit. Rev. Plant Sci. 2015, 34, 327–380. [Google Scholar] [CrossRef]
- Allen, S.G.; Dobrenz, A.K.; Schonhorst, M.H.; Stoner, J.E. Heritability of NaCl tolerance in germinating alfalfa seeds. Agron. J. 1985, 77, 99–101. [Google Scholar] [CrossRef]
- Benabderrahim, M.A.; Guiza, M.; Haddad, M. Genetic diversity of salt tolerance in tetraploid alfalfa (Medicago sativa L.). Acta Physiol. Plant. 2020, 42, 5. [Google Scholar] [CrossRef]
- Jiang, J.; Yang, B.L.; Xia, T.; Yu, S.M.; Wu, Y.N.; Jin, H.; Li, J.R. Analysis of genetic diversity of salt-tolerant alfalfa germplasms. Genet. Mol. Res. 2015, 14, 4438–4447. [Google Scholar] [CrossRef] [PubMed]
- Azzam, C.R.; Naby, Z.M.A.E.; Mohamed, N.A. Salt tolerance associated with molecular markers in alfalfa. J. Biosci. Appl. Res. 2019, 5, 416–428. [Google Scholar]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil. 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Tiwari, R.; Mamrutha, H.M. Precision phenotyping for mapping of traits for abiotic stress tolerance in crops. In Biotechnology: Prospects and Applications; Salar, R., Gahlawat, S., Siwach, P., Duhan, J., Eds.; Springer: New Delhi, India, 2013. [Google Scholar]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating high-throughput phenotyping into genetic gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [Green Version]
- Fahlgren, N.; Gehan, M.A.; Baxter, I. Lights, camera, action: High-throughput plant phenotyping is ready for a close-up. Curr. Opin. Plant Biol. 2015, 24, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Römer, C.; Wahabzada, M.; Ballvora, A.; Pinto, F.; Rossini, M.; Panigada, C.; Behmann, J.; Léon, J.; Thurau, C.; Bauckhage, C.; et al. Early drought stress detection in cereals: Simplex volume maximisation for hyperspectral image analysis. Funct. Plant Biol. 2012, 39, 878–890. [Google Scholar] [CrossRef]
- Naumann, J.C.; Young, D.R.; Anderson, J.E. Spatial variations in salinity stress across a coastal landscape using vegetation indices derived from hyperspectral imagery. Plant Ecol. 2009, 202, 285–297. [Google Scholar] [CrossRef]
- Behmann, J.; Steinrucken, J.; Plumer, L. Detection of early plant stress responses in hyperspectral images. ISPRS J. Photogramm. Remote Sens. 2014, 93, 98–111. [Google Scholar] [CrossRef]
- Sytar, O.; Brestic, M.; Zivcak, M.; Olsovska, K.; Kovar, M.; Shao, H.B.; He, X.L. Applying hyperspectral imaging to explore natural plant diversity towards improving salt stress tolerance. Sci. Total Environ. 2017, 578, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.P.; Hawkins, C.; Peel, M.D.; Yu, L.X. Genetic loci associated with salt tolerance in advanced breeding populations of tetraploid alfalfa using genome wide association studies. Plant Genome 2019, 12, 180026. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.X.; Liu, X.; Boge, W.; Liu, X.P. Genome-wide association study identifies loci for salt tolerance during germination in autotetraploid alfalfa (Medicago sativa L.) using genotyping-by-sequencing. Front. Plant Sci. 2016, 7, 956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.P.; Yu, L.X. Genome-wide association mapping of loci associated with plant growth and forage production under salt stress in alfalfa (Medicago sativa L.). Front. Plant Sci. 2017, 8, 853. [Google Scholar] [CrossRef]
- Zhang, T.; Yu, L.-X.; Zheng, P.; Li, Y.; Rivera, M.; Main, D.; Greene, S.L. Identification of loci associated with drought resistance traits in heterozygous autotetraploid alfalfa (Medicago sativa L.) using genome-wide association studies with genotyping by sequencing. PLoS ONE 2015, 10, e0138931. [Google Scholar] [CrossRef] [Green Version]
- Meuwissen, T.H.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar]
- Fu, Y.B.; Yang, M.H.; Zeng, F.; Biligetu, B. Searching for an accurate marker-based prediction of an individual quantitative trait in molecular plant breeding. Front. Plant Sci. 2017, 8, 1182. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Global salt-affected regions and their severity levels (drawn based on Harmonized World Soil Database v. 1.2 [20]).
Figure 1.
Global salt-affected regions and their severity levels (drawn based on Harmonized World Soil Database v. 1.2 [20]).

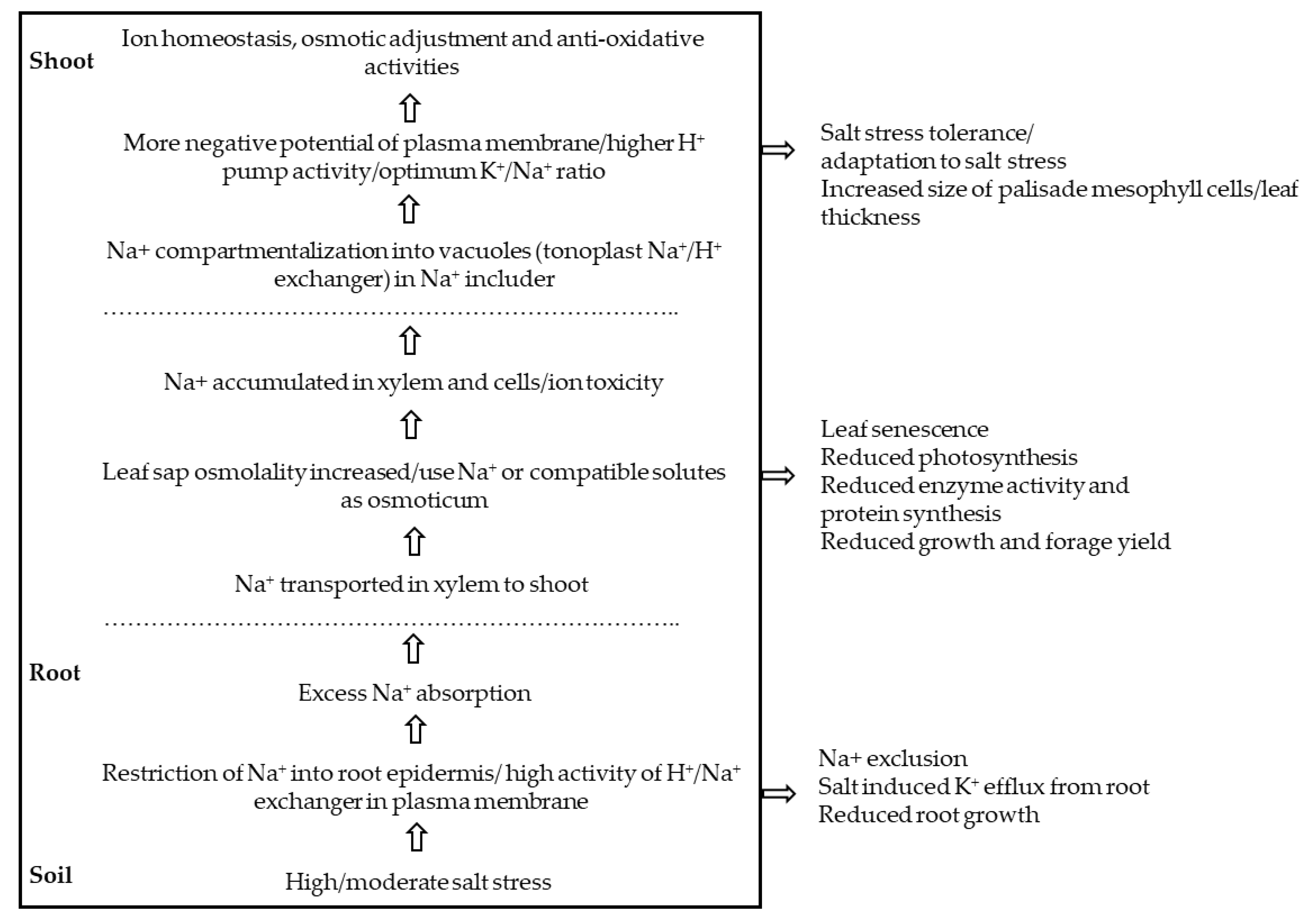
Figure 2.
Summary of salt stress tolerance in alfalfa.


{kind=link}
{kind=link}
Table 1.
Number of differently expressed genes and proteins and their functions in salt-tolerant and intolerant alfalfa cultivars.
Table 1.
Number of differently expressed genes and proteins and their functions in salt-tolerant and intolerant alfalfa cultivars.
| Alfalfa Genotype | Tissue | Salt Stress | Total Number of Differentially Expressed Genes/Proteins | Major Pathway/Function | Reference |
|---|---|---|---|---|---|
| NM-801 (tolerant), Vernal (intolerant) | root | 2-week-old seedlings treated with ~5, ~10 dS m−1 NaCl for 3 days | 83 | Ion homeostasis, protein turnover and signaling, protein folding, cell wall components, carbohydrate and energy metabolism, reactive oxygen species regulation and detoxification, and purine and fatty acid metabolism. | [81] |
| Zhongmu-1 (M. sativa, tolerant), Jemalong A17 (M. truncatula, intolerant) | root | 1-month-old seedlings treated with ~30 dS m−1 NaCl for 8 h | 93 (tolerant) 30 (intolerant) | Molecule binding and catalytic activity. Defense against oxidative stress, metabolism, photosynthesis, protein synthesis and processing, and signal transduction. | [82] |
| AZ-88NDC (intolerant), AZ-GERM SALT-II (tolerant) | root | 1-week-old seedlings treated with ~15 dS m−1 NaCl for 7 days | 288/273 and 468/337 up/down-regulated in intolerant and tolerant, respectively | Response to stress, kinase activity, hydrolase activity, oxidoreductase activity, and extracellular region. | [83] |
| Zhongmu No. 1 (tolerant) | root | 12-day-old seedlings treated with ~25 dS m−1 NaCl for 1, 3, 6, 12, 24 h | 8861 at one or more time points | Iron ion transport, ion homeostasis, antiporter, signal perception, signal transduction, transcriptional regulation, and antioxidative defense. | [84] |
| Zhongmu No. 1 (tolerant) | root, shoot | 1-week-old seedlings treated with ~10, ~20 dS m−1 NaCl for 7 days | 26 (shoot) 35 (root) | Photosynthesis (31%) and stress and defense (20%) in the shoot. Defense (26%); metabolism (17%); and protein translation, processing, and degradation (17%) in the root. | [54] |
| CW064027, Bridgeview (tolerant), Rangelander (intolerant) | shoot | 4th-cut treated with 1.53, 8, 15.6 dS m−1 maintained by sulphate- based sodium, calcium, and magnesium salts | 685/527, 368/139 up/down-regulated in CW064027 and Bridgeview at control, 537/949, 375/1045 up/down-regulated in CW064027 and Bridgeview at 8 dS m−1, 1129/1196, 843/1516 up/down-regulated in CW064027 and Bridgeview at 15.6 dS m−1 | Redox-related genes, B-ZIP transcripts, cell wall structural components, lipids, secondary metabolism, auxin and ethylene hormones, development, transport, signaling, heat shock, proteolysis, pathogenesis-response, abiotic stress, RNA processing, and protein metabolism. | [85] |
| Zhongmu-1 (tolerant), Xingjiang Daye (intolerant) | leaf | 30-day-old plants treated with ~50 dS m−1 NaCl for 7 days | 1125 and 2237 between cultivars at control and stress, respectively | Response to stimulus, reactive oxygen species, responding to stress, and response to hormone and other stress-responsive processes. | [49] |
Table 2.
Salt-tolerant alfalfa cultivars and their fall dormancy (FD) and growth stages (G, germination and/or F, forage production) for salt tolerance (ST) (Source: National Alfalfa & Forage Alliance, [89]).
Table 2.
Salt-tolerant alfalfa cultivars and their fall dormancy (FD) and growth stages (G, germination and/or F, forage production) for salt tolerance (ST) (Source: National Alfalfa & Forage Alliance, [89]).
| Variety | FD | ST | Variety | FD | ST |
|---|---|---|---|---|---|
| Foothold | 2 | G | GUNNER | 5 | G |
| Spredor 5 | 2 | G | MPIII Max Q | 5 | G |
| Hi-Gest 360 | 3 | G | RR NemaStar | 5 | G |
| LegenDairy XHD | 3 | G | RR Tonnica | 5 | G |
| HVX Tundra II | 3 | G | WL 365HQ | 5 | G |
| LegenDairy AA | 3 | G | 6610N | 6 | G |
| RR Presteez | 3 | G | Cisco II | 6 | G/F |
| Rugged | 3 | G | Hi-Gest 660 | 6 | G |
| WL 336HQ.RR | 3 | G | Revolt | 6 | G |
| 6401N | 4 | G | RRALF 6R200 | 6 | G |
| 6472A | 4 | G | WL 454HQ.RR | 6 | G/F |
| 6497R | 4 | G | 6829R | 7 | G |
| AFX 457 | 4 | G | AFX 779 | 7 | G |
| AFX 469 | 4 | G | AmeriStand 715NT RR | 7 | G/F |
| AmeriStand 415NT RR | 4 | G | Sun Titan | 8 | G |
| AmeriStand 427TQ | 4 | G | SW8421S | 8 | F |
| AmeriStand 455TQ RR | 4 | G | WL 535HQ | 8 | G |
| AmeriStand 457TQ RR | 4 | G | WL 552HQ.RR | 8 | G |
| AmeriStand 480 HVXRR | 4 | G | 6906N | 9 | G |
| Barricade SLT | 4 | G/F | AFX 960 | 9 | G |
| DKA40-16 | 4 | G | AmeriStand 901TS | 9 | G |
| DKA44-16RR | 4 | G | LG 9C300 | 9 | G |
| Integra 8444R | 4 | G/F | PGI 908-S | 9 | G/F |
| Magnum Salt | 4 | G/F | RRALF 9R100 | 9 | G |
| Rebound AA | 4 | G | Sun Quest | 9 | G |
| RR Stratica | 4 | G | SW 9215 | 9 | F |
| RR VaMoose | 4 | G | SW 9720 | 9 | F |
| WL 356HQ.RR | 4 | G | SW9215RRS | 9 | G/F |
| 6516R | 5 | G | WL 656HQ | 9 | G |
| 6547R | 5 | G/F | WL 668HQ.RR | 9 | G |
| AFX 579 | 5 | G | 6015R | 10 | G |
| Nimbus | 5 | F | AFX 1060 | 10 | G |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bhattarai, S.; Biswas, D.; Fu, Y.-B.; Biligetu, B. Morphological, Physiological, and Genetic Responses to Salt Stress in Alfalfa: A Review. Agronomy 2020, 10, 577. https://doi.org/10.3390/agronomy10040577
AMA Style
Bhattarai S, Biswas D, Fu Y-B, Biligetu B. Morphological, Physiological, and Genetic Responses to Salt Stress in Alfalfa: A Review. Agronomy. 2020; 10(4):577. https://doi.org/10.3390/agronomy10040577
Chicago/Turabian StyleBhattarai, Surendra, Dilip Biswas, Yong-Bi Fu, and Bill Biligetu. 2020. "Morphological, Physiological, and Genetic Responses to Salt Stress in Alfalfa: A Review" Agronomy 10, no. 4: 577. https://doi.org/10.3390/agronomy10040577
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.