Integration of Extra-Large-Seeded and Double-Podded Traits in Chickpea (Cicer arietinum L.)


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parents in Intraspecific Crosses
2.2. Agronomic Applications
2.3. Soil Properties
2.4. Weather Conditions
2.5. Data Collection
2.6. Data Analyses
3. Results
3.1. F1 Progeny and F2 and F3 Populations
3.2. Transgressive Segregations
3.3. Comparisons of Number of Pods per Node and Leaf Shapes
3.4. Inheritance of Seed Size and Agro-Morphological Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Toker, C.; Yadav, S.S. Legumes cultivars for stress environments. In Climate Change and Management of Cool Season Grain Legume Crops; Yadav, S.S., McNeil, D.L., Redden, R., Patil, S.A., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 351–376. [Google Scholar]
- Devasirvatham, V.; Tan, D.K.Y. Impact of high temperature and drought stresses on chickpea production. Agronomy 2018, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Gaur, P.M.; Samineni, S.; Thudi, M.; Tripathi, S.; Sajja, S.B.; Jayalakshmi, V.; Mannur, D.M.; Vijayakumar, A.G.; Ganga Rao, N.V.; Ojiewo, C.; et al. Integrated breeding approaches for improving drought and heat adaptation in chickpea (Cicer arietinum L.). Plant Breed. 2019, 138, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Naseer, I.; Hussain, A.; Mumtaz, M.Z.; Mustafa, A.; Hilger, T.H.; Zahir, Z.A.; Xu, M. Appraising Endophyte–Plant symbiosis for improved growth, nodulation, nitrogen fixation and abiotic stress tolerance: An experimental investigation with chickpea (Cicer arietinum L.). Agronomy 2019, 9, 621. [Google Scholar] [CrossRef] [Green Version]
- Latati, M.; Dokukin, P.; Aouiche, A.; Rebouh, N.Y.; Takouachet, R.; Hafnaoui, E.; Hamdani, F.Z.; Bacha, F.; Ounane, S.M. Species interactions improve above-ground biomass and land use efficiency in intercropped wheat and chickpea under low soil inputs. Agronomy 2019, 9, 765. [Google Scholar] [CrossRef] [Green Version]
- Soe, K.M.; Htwe, A.Z.; Moe, K.; Tomomi, A.; Yamakawa, T. Diversity and effectivity of indigenous Mesorhizobium strains for chickpea (Cicer arietinum L.) in Myanmar. Agronomy 2020, 10, 287. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization Statistical Databases (FAOSTAT). Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 13 May 2020).
- Van der Maessen, L.J.G. Cicer L. a Monograph of the Genus, with Special Reference to the Chickpea (Cicer arietinum L.), Its Ecology and Cultivation. Ph.D. Thesis, Veenman, Utrecht, The Nederland, 1972. [Google Scholar]
- Muehlbauer, F.J.; Singh, K.B. Genetics of chickpea. In The Chickpea; Saxena, M.C., Singh, K.B., Eds.; CAB Int.: Wallingford, CT, USA, 1987; pp. 99–125. [Google Scholar]
- Regan, K.; MacLeod, B.; Siddique, K. Production packages for kabuli chickpea in Western Australia (No. 117); Farm Note; The University of Western Australia: Perth, Australia, 2006. [Google Scholar]
- Abbo, S.; Grusak, M.A.; Tzuk, T.; Reifen, R. Genetic control of seed weight and calcium concentration in chickpea seed. Plant Breed. 2000, 119, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, H.D.; Kumar, S.; Gowda, C.L.L.; Singh, S. Two major genes for seed size in chickpea (Cicer arietinum L.). Euphytica 2006, 147, 311–315. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, P.; Samineni, S.; Sajja, S.B.; Roy, C.; Singh, S.P.; Joshi, P.; Gaur, P.M. Inheritance and relationships of flowering time and seed size in kabuli chickpea. Euphytica 2019, 215, 144. [Google Scholar] [CrossRef] [Green Version]
- Bicer, B.T.; Sakar, D. Heritability and gene effects for yield and yield components in chickpea. Hereditas 2008, 145, 220–224. [Google Scholar] [CrossRef]
- Bicer, B.T. The effect of seed size on yield and yield components of chickpea and lentil. Afr. J. Biotechnol. 2009, 8. [Google Scholar]
- Singh, K.B. Chickpea breeding. In The Chickpea; Saxena, M.C., Singh, K.B., Eds.; CAB Int.: Wallingford, CT, USA, 1987; pp. 127–162. [Google Scholar]
- Singh, B.D.; Jaiswal, H.K.; Singh, R.M.; Singh, A.K. Isolation of early-flowering recombinants from the interspecific cross between Cicer arietinum and Cicer reticulatum. Int. Chickpea Newslett. 1984, 11, 14–16. [Google Scholar]
- Pundir, R.P.S.; Reddy, K.N.; Mengesha, M.H. ICRISAT Chickpea Germplasm Catalog: Evaluation and Analysis; ICRISAT: Patancheru, India, 1988. [Google Scholar]
- Toker, C. Estimate of heritabilities and genotype by environment interactions for 100-seed weight, days to flowering and plant height in kabuli chick-peas (Cicer arietinum L.). Turk. J. Field Crop. 1998, 3, 16–20. [Google Scholar]
- Koseoglu, K.; Adak, A.; Sari, D.; Sari, H.; Ceylan, F.O.; Toker, C. Transgressive segregations for yield criteria in reciprocal interspecific crosses between Cicer arietinum L. and C. reticulatum Ladiz. Euphytica 2017, 213, 116. [Google Scholar] [CrossRef]
- Dusunceli, F.; Wood, J.A.; Gupta, A.; Yadav, M.; Yadav, S.S. International trade. In Chickpea Breeding and Management; Yadav, S.S., Redden, R.J., Chen, W., Sharma, B., Eds.; CAB Int.: Wallingford, CT, USA, 2007; pp. 555–575. [Google Scholar]
- Gaur, P.M.; Singh, M.K.; Samineni, S.; Sajja, S.B.; Jukanti, A.K.; Kamatam, S.; Varshney, R.K. Inheritance of protein content and its relationships with seed size, grain yield and other traits in chickpea. Euphytica 2016, 209, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Muehlbauer, F.J.; Sarker, A. Economic Importance of Chickpea: Production, Value, and World Trade. In The Chickpea Genome; Varshney, R.K., Thudi, M., Muehlbauer, F.J., Eds.; Springer: Cham, Switzerland, 2017; pp. 5–12. [Google Scholar]
- Toker, C.; Lluch, C.; Tejera, N.A.; Serraj, R.; Siddique, K.H.M. Abiotic stresses. In Chickpea Breeding and Management; Yadav, S.S., Redden, R.J., Chen, W., Sharma, B., Eds.; CAB International: Wallingford, CT, USA, 2007; pp. 474–496. [Google Scholar]
- Balasubrahmanyan, R. The association of size and colour in gram (Cicer arietinum L.). Curr. Sci. 1950, 19, 246–247. [Google Scholar] [PubMed]
- Malhotra, R.S.; Singh, K.B. Detection of epistasis in chickpea. Euphytica 1989, 40, 169–172. [Google Scholar]
- Singh, O.; Gowda, C.L.L.; Sethi, S.C.; Dasgupta, T.; Smithson, J.B. Genetic analysis of agronomic characters in chickpea. Theor. Appl. Genet. 1992, 83, 956–962. [Google Scholar] [CrossRef]
- Singh, O.; Gowda, C.L.L.; Sethi, S.C.; Dasgupta, T.; Kumar, J.; Smithson, J.B. Genetic analysis of agronomic characters in chickpea. II. Estimates of genetic variances from line× tester mating designs. Theor. Appl. Genet. 1993, 85, 1010–1016. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, O. Inheritance of seed size in chickpea. J. Genet. Breed. 1995, 49, 99–103. [Google Scholar]
- Malhotra, R.S.; Bejiga, G.; Singh, K.B. Inheritance of seed size in chickpea. J. Genet. Breed. 1997, 51, 45–50. [Google Scholar]
- Hovav, R.; Upadhyaya, K.C.; Beharav, A.; Abbo, S. Major flowering time gene and polygene effects on chickpea seed weight. Plant Breed. 2003, 122, 539–541. [Google Scholar] [CrossRef]
- Hossain, S.; Ford, R.; McNeil, D.; Pittock, C.; Panozzo, J.F. Inheritance of Seed Size in Chickpea (Cicer arietinum L.) and Identification of QTL Based on 100-seed Weight and Seed Size Index. Aust. J. Crop Sci. 2010, 4, 126. [Google Scholar]
- Sharma, S.; Upadhyaya, H.D.; Gowda, C.L.L.; Kumar, S.; Singh, S. Genetic analysis for seed size in three crosses of chickpea (Cicer arietinum L.). Can. J. Plant Sci. 2013, 93, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Cobos, M.J.; Rubio, J.; Fernández-Romero, M.D.; Garza, R.; Moreno, M.T.; Millán, T.; Gil, J. Genetic analysis of seed size, yield and days to flowering in a chickpea recombinant inbred line population derived from a Kabuli × Desi cross. Ann. Appl. Biol. 2007, 151, 33–42. [Google Scholar] [CrossRef]
- Verma, S.; Gupta, S.; Bandhiwal, N.; Kumar, T.; Bharadwaj, C.; Bhatia, S. High-density linkage map construction and mapping of seed trait QTLs in chickpea (Cicer arietinum L.) using genotyping-by-sequencing (GBS). Sci. Rep. 2015, 5, 17512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Khan, A.W.; Jaganathan, D.; Thudi, M.; Roorkiwal, M.; Takagi, H.; Sutton, T. QTL-seq for rapid identification of candidate genes for 100-seed weight and root/total plant dry weight ratio under rainfed conditions in chickpea. Plant Biotechnol. J. 2016, 14, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Lal, D.; Ravikumar, R.L. Development of Genetic Linkage Map and Identification of QTLs for Agronomic Traits in Chickpea (Cicer arietinum L.). Int. J. Curr. Microbiol. App. Sci. 2018, 7, 66–77. [Google Scholar] [CrossRef]
- Sheldrake, A.R.; Saxena, N.P.; Krishnamurthy, L. The expression and influence on yield of the ‘double-podded ’ character in chickpeas (Cicer arietinum L.). Field Crops Res. 1978, 1, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Srivastava, R.K.; Ganesh, M. Penetrance and expressivity of the gene for double podding in chickpea. J. Hered. 2000, 91, 234–236. [Google Scholar] [CrossRef] [Green Version]
- Yasar, M.; Ceylan, F.O.; Ikten, C.; Toker, C. Comparison of expressivity and penetrance of the double podding trait and yield components based on reciprocal crosses of kabuli and desi chickpeas (Cicer arietinum L.). Euphytica 2014, 196, 331–339. [Google Scholar] [CrossRef]
- Adak, A.; Sari, D.; Sari, H.; Toker, C. Gene effects of Cicer reticulatum on qualitative and quantitative traits in the cultivated chickpea. Plant Breed. 2017, 136, 939–947. [Google Scholar] [CrossRef]
- Rubio, J.; Moreno, M.T.; Cubero, J.I.; Gil, J. Effect of the gene for double pod in chickpea on yield, yield components and stability of yield. Plant Breed. 1998, 117, 585–587. [Google Scholar] [CrossRef]
- Rubio, J.; Flores, F.; Moreno, M.T.; Cubero, J.I.; Gil, J. Effects of the erect/bushy habit, single/double pod and late/early flowering genes on yield and seed size and their stability in chickpea. Field Crops Res. 2004, 90, 255–262. [Google Scholar] [CrossRef]
- Srinivasan, S.; Gaur, P.M.; Chaturvedi, S.K.; Rao, B.V. Allelic relationships of genes controlling number of flowers per axis in chickpea. Euphytica 2006, 152, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Rajesh, P.N.; Tullu, A.; Gil, J.; Gupta, V.; Ranjekar, P.; Muehlbauer, F. Identification of an STMS marker for the double-podding gene in chickpea. Theor. Appl. Genet. 2002, 105, 604–607. [Google Scholar] [CrossRef]
- Cho, S.; Kumar, J.; Shultz, J.L.; Anupama, K.; Tefera, F.; Muehlbauer, F.J. Mapping genes for double podding and other morphological traits in chickpea. Euphytica 2002, 128, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Radhika, P.; Gowda, S.J.M.; Kadoo, N.Y.; Mhase, L.B.; Jamadagni, B.M.; Sainani, M.N.; Gupta, V.S. Development of an integrated intraspecific map of chickpea (Cicer arietinum L.) using two recombinant inbred line populations. Theor. Appl. Genet. 2007, 115, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Taran, B.; Warkentin, T.D.; Vandenberg, A. Fast track genetic improvement of ascochyta blight resistance and double podding in chickpea by marker-assisted backcrossing. Theor. Appl. Genet. 2013, 126, 1639–1647. [Google Scholar] [CrossRef]
- Ali, L.; Deokar, A.; Caballo, C.; Tar’an, B.; Gil, J.; Chen, W.; Millan, L.; Rubio, J. Fine mapping for double podding gene in chickpea. Theor. Appl. Genet. 2016, 129, 77–86. [Google Scholar] [CrossRef]
- Singh, M.; Rani, S.; Malhotra, N.; Katna, G.; Sarker, A. Transgressive segregations for agronomic improvement using interspecific crosses between C. arietinum L. x C. reticulatum Ladiz. and C. arietinum L. x C. echinospermum Davis species. PLoS ONE 2018, 13, e0203082. [Google Scholar] [CrossRef]
- De Vicente, M.C.; Tanksley, S.D. QTL analysis of transgressive segregation in an interspecific tomato cross. Genetics 1993, 134, 585–596. [Google Scholar]
- Gaur, P.M.; Pande, S.; Upadhyaya, H.D.; Rao, B.V. Extra-large kabuli chickpea with high resistance to fusarium wilt. J. Agric. Res. 2006, 2, 1–2. [Google Scholar]
- Bonfil, D.J.; Goren, O.; Mufradi, I.; Lichtenzveig, J.; Abbo, S. Development of early-flowering kabuli chickpea with compound and simple leaves. Plant Breed. 2007, 126, 125–129. [Google Scholar] [CrossRef]
- Gil, J.; Castro, P.; Millan, T.; Madrid, E.; Rubio, J. Development of new kabuli large-seeded chickpea materials with resistance to Ascochyta blight. Crop Pasture Sci. 2017, 68, 967–972. [Google Scholar] [CrossRef]
- Muehlbauer, F.J.; Temple, S.R.; Chen, W. Registration of Sierra chickpea. Crop Sci. 2004, 44, 1864–1865. [Google Scholar] [CrossRef]
- Warkentin, T.; Banniza, S.; Vandenberg, A. CDC Frontier kabuli chickpea. Can. J. Plant Sci. 2005, 85, 909–910. [Google Scholar] [CrossRef]
- Gan, Y.T.; Siddique, K.H.M.; MacLeod, W.J.; Jayakumar, P. Management options for minimizing the damage by ascochyta blight (Ascochyta rabiei) in chickpea (Cicer arietinum L.). Field Crops Res. 2006, 97, 121–134. [Google Scholar] [CrossRef]
- Gowda, C.L.L.; Upadhyaya, H.D.; Dronavalli, N.; Singh, S. Identification of large-seeded high-yielding stable kabuli chickpea germplasm lines for use in crop improvement. Crop Sci. 2011, 51, 198–209. [Google Scholar] [CrossRef] [Green Version]
- Taran, B.; Bandara, M.; Warkentin, T.; Banniza, S.; Vandenberg, A. CDC Orion kabuli chickpea. Can. J. Plant Sci. 2011, 91, 355–356. [Google Scholar] [CrossRef]
- Vandemark, G.; Guy, S.O.; Chen, W.; McPhee, K.; Pfaff, J.; Lauver, M.; Muehlbauer, F.J. Registration of ‘Nash’chickpea. J. Plant Regist. 2015, 9, 275–278. [Google Scholar] [CrossRef]
- Urrea, C.A.; Muehlbauer, F.J.; Harveson, R.M. Registration of ‘New Hope’Chickpea Cultivar with Enhanced Resistance to Ascochyta Blight. J. Plant Regist. 2017, 11, 107–111. [Google Scholar] [CrossRef]
- Vandemark, G.; Nelson, H.; Chen, W.; McPhee, K.; Muehlbauer, F. Registration of ‘Royal’ Chickpea. J. Plant Regist. 2019, 13, 123–127. [Google Scholar] [CrossRef]
- Rubio, J.; Moreno, M.T.; Martinez, C.; Gil, J. Registration of CA2969, an Ascochyta blight resistant and double-podded chickpea germplasm. (Registrations of Germplasm). Crop Sci. 2003, 43, 1567–1569. [Google Scholar] [CrossRef]
- Auckland, A.K.; Van der Maesen, L.J.G. Chickpea. In Hybridization of Crop Plants; Fehr, W.R., Hadley, H.H., Eds.; American Society of Agronomy and Crop Science Society of America: Madison, WI, USA, 1980; pp. 249–259. [Google Scholar]
- MINITAB 2014 Inc. Minitab Statistical Software, Release 17 for Windows; MINITAB 2014 Inc.: State College, PA, USA, 2014. [Google Scholar]
- Poehlman, J.M.; Sleper, D.A. Breeding Field Crops; Iowa State University Press: Ames, IA, USA, 1995; p. 494. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Rao, N.K.; Pundir, R.P.S.; Van Der Maesen, L.J.G. Inheritance of some qualitative characters in chickpea (Cicer arietinum L.). Proc. Plant Sci. 1980, 89, 497–503. [Google Scholar]
- Pundir, R.P.S.; Mengesha, M.H.; Reddy, K.N. Leaf types and their genetics in chickpea (Cicer arietinum L.). Euphytica 1990, 45, 197–200. [Google Scholar]
- Danehloueipour, N.; Clarke, H.J.; Yan, G.; Khan, T.N.; Siddique, K.H.M. Leaf type is not associated with ascochyta blight disease in chickpea (Cicer arietinum L.). Euphytica 2008, 162, 281–289. [Google Scholar] [CrossRef]
- Toker, C.; Ceylan, F.O.; Inci, N.E.; Yildirim, T.; Cagirgan, M.I. Inheritance of leaf shape in the cultivated chickpea (Cicer arietinum L.). Turk Field Crops 2012, 17, 16–18. [Google Scholar]
- Abbo, S.; Goren, O.; Saranga, Y.; Langensiepen, M.; Bonfil, D. Leaf shape × sowing density interaction affects chickpea grain yield. Plant Breed. 2013, 132, 200–204. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Sharma, S.; Gowda, C.L. Major genes with additive effects for seed size in kabuli chickpea (Cicer arietinum L.). J. Genet. 2011, 90, 479–482. [Google Scholar] [CrossRef] [Green Version]
- Ceylan, F.O.; Adak, A.; Sari, D.; Sari, H.; Toker, C. Unveiling of suppressed genes in interspecific and backcross populations derived from mutants of Cicer species. Crop Pasture Sci. 2019, 70, 254–262. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype of F1s | F2 | χ2 | p | ||
|---|---|---|---|---|---|
| Phenotype of F2 Population | Observed | Expected | |||
| Imparipinnate leaf and single pod | Imparipinnate leaf and single pod | 475 | 9:3:3:1 | 8.25 | 0.50–0.10 |
| Imparipinnate leaf and double pods | 181 | ||||
| Unifoliolate leaf and single pod | 189 | ||||
| Unifoliolate leaf and double pods | 70 | ||||
| Traits | Sierra | CA 2969 | Imparipinnate Leaf | Unifoliolate Leaf | F2 Population | h2 | |||
|---|---|---|---|---|---|---|---|---|---|
| Single Pod | Double Pods | Single Pod | Double Pods | ||||||
| Range | |||||||||
| Days to first flowering (days) | 48.0 ± 0.97 | 50.0 ± 0.90 | 41.0 ± 0.29 | 41.5 ± 0.50 | 41.3 ± 0.47 | 40.8 ± 0.70 | 41.2 ± 0.21 | 45–66 | 0.80 |
| Days to 50% flowering (days) | 50.0 ± 0.97 | 52.0 ± 0.90 | 47.6 ± 0.29 | 47.8 ± 0.50 | 48.1 ± 0.46 | 47.5 ± 0.77 | 47.8 ± 0.24 | 45–75 | 0.80 |
| Plant height (cm) | 41.0 ± 0.97 | 34.3 ± 1.17 | 37.7 ± 0.31 | 37.6 ± 0.50 | 38.9 ± 0.43 | 41.7 ± 0.70 | 38.9 ± 0.21 | 17–59 | 0.43 |
| First pod height (cm) | 21.0 ± 0.73 | 19.7 ± 2.35 | 30.5 ± 0.32 | 30.2 ± 0.44 | 32.2 ± 0.49 | 34.0 ± 0.85 | 31.7 ± 0.23 | 11–58 | 0.65 |
| Main stems per plant (No.) | 2.7 ± 0.49 | 2.3 ± 0.21 | 2.0 ± 0.04 | 2.1 ± 0.06 | 2.4 ± 0.07 | 2.7 ± 0.15 | 2.3 ± 0.03 | 1–7 | 0.38 |
| Pods per plant (No.) | 5.2 ± 1.30 | 3.0 ± 2.54 | 3.6 ± 0.16 | 4.5 ± 0.30 | 3.6 ± 0.28 | 4.5 ± 0.55 | 4.1 ± 0.11 | 1–25 | 0.49 |
| Seeds per plant (No.) | 4.2 ± 1.08 | 11.7 ± 1.96 | 3.5 ± 0.14 | 4.2 ± 0.28 | 3.3 ± 0.22 | 4.2 ± 0.44 | 3.8 ± 0.11 | 1–24 | 0.66 |
| Seed yield (g) | 1.8 ± 0.56 | 3.7 ± 0.72 | 1.3 ± 0.05 | 1.5 ± 0.10 | 1.3 ± 0.10 | 1.5 ± 0.16 | 1.4 ± 0.04 | 0.1–6.9 | 0.60 |
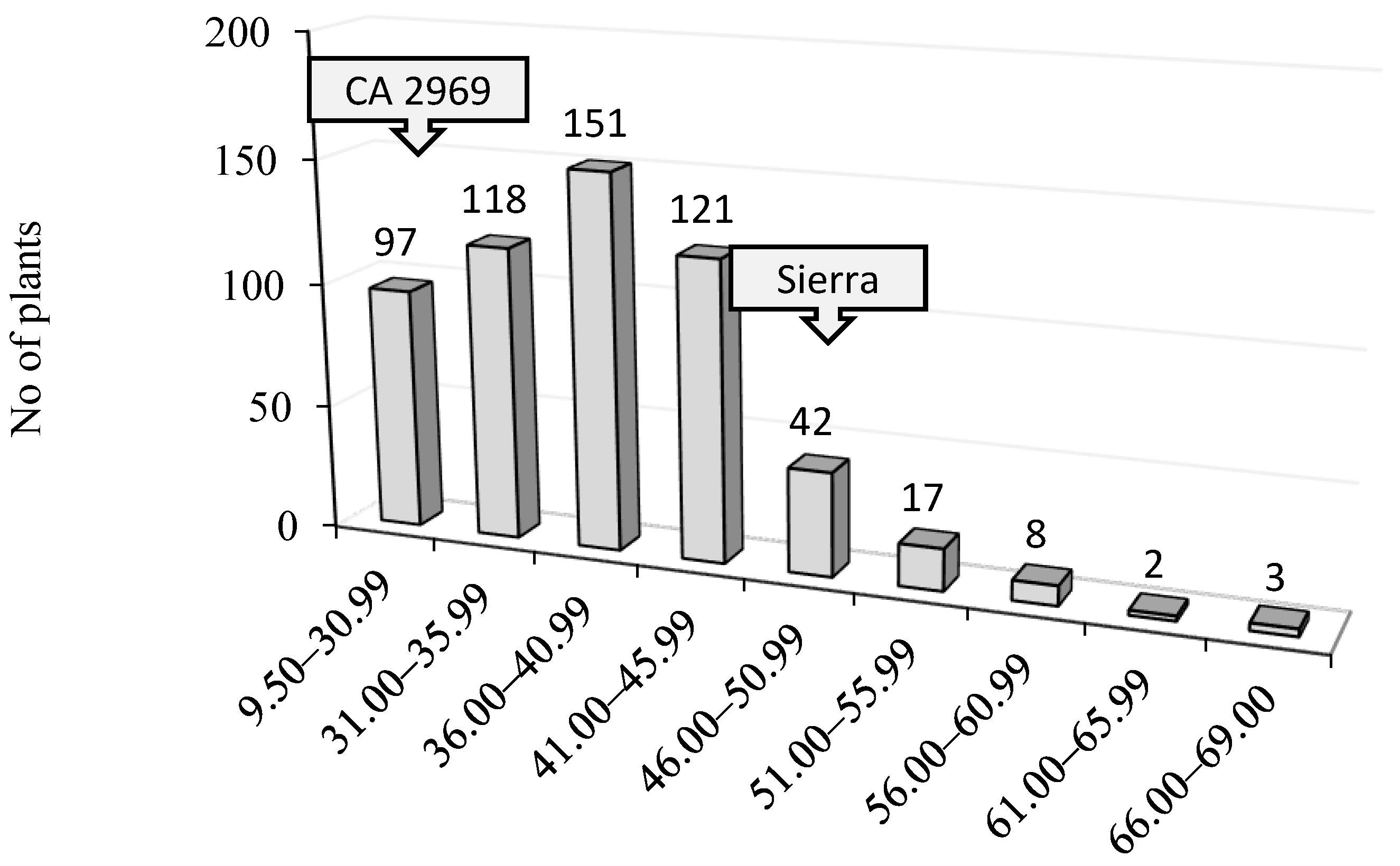
| 100-seed weight (g) | 49.9 ± 1.61 | 31.0 ± 1.26 | 37.2 ± 0.54 | 35.2 ± 0.76 | 37.3 ± 1.70 | 36.0 ± 0.9 | 36.4 ± 0.36 | 9.5–69.0 | 0.45 |
| Traits | Sierra | CA 2969 | Imparipinnate Leaf | Unifoliolate Leaf | F3 Population | |||
|---|---|---|---|---|---|---|---|---|
| Single Pod | Double Pods | Single Pod | Double Pods | |||||
| Range | ||||||||
| Days to first flowering (days) | 48.3 ± 0.08 | 50.0 ± 0.50 | 46.1 ± 0.23 | 46.2 ± 0.19 | 45.0 ± 0.29 | 46.4 ± 0.27 | 46.0 ± 0.12 | 36–75 |
| Days to 50% flowering (days) | 50.3 ± 0.08 | 52.3 ± 0.42 | 48.9 ± 0.20 | 49.2 ± 0.17 | 47.9 ± 0.23 | 49.2 ± 0.21 | 48.9 ± 0.10 | 38–82 |
| Plant height (cm) | 52.3 ± 0.61 | 42.7 ± 0.08 | 43.8 ± 0.27 | 44.5 ± 0.25 | 48.9 ± 0.42 | 50.8 ± 0.40 | 46.3 ± 0.18 | 19–68 |
| First pod height (cm) | 31.3 ± 0.38 | 33.3 ± 0.79 | 29.9 ± 0.26 | 30.7 ± 0.25 | 31.6 ± 0.41 | 32.4 ± 0.37 | 30.9 ± 0.15 | 13–46 |
| Main stems per plant (No.) | 2.3 ± 0.09 | 2.3 ± 0.08 | 2.7 ± 0.04 | 2.8 ± 0.04 | 2.8 ± 0.05 | 2.9 ± 0.04 | 2.8 ± 0.02 | 1–6 |
| Pods per plant (No.) | 36.7 ± 2.05 | 42.7 ± 2.7 | 62.7 ± 1.87 | 69.9 ± 1.85 | 48.5 ± 1.75 | 48.8 ± 1.35 | 59.5 ± 0.96 | 1–254 |
| Seeds per plant (No.) | 32.7 ± 2.02 | 50.7 ± 2.46 | 68.0 ± 2.06 | 77.9 ± 2.09 | 51.4 ± 1.85 | 52.3 ± 1.46 | 64.9 ± 1.07 | 1–267 |
| Seed yield (g) | 15.2 ± 0.86 | 13.9 ± 0.72 | 24.3 ± 0.74 | 26.4 ± 0.69 | 16.3 ± 0.58 | 16.3 ± 0.46 | 21.9 ± 0.37 | 0.1–79.0 |
| 100-seed weight (g) | 46.9 ± 0.31 | 27.4 ± 0.34 | 42.1 ± 0.38 | 41.4 ± 0.38 | 38.9 ± 0.45 | 38.5 ± 0.41 | 40.7 ± 0.21 | 7.0–64.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kivrak, K.G.; Eker, T.; Sari, H.; Sari, D.; Akan, K.; Aydinoglu, B.; Catal, M.; Toker, C. Integration of Extra-Large-Seeded and Double-Podded Traits in Chickpea (Cicer arietinum L.). Agronomy 2020, 10, 901. https://doi.org/10.3390/agronomy10060901
Kivrak KG, Eker T, Sari H, Sari D, Akan K, Aydinoglu B, Catal M, Toker C. Integration of Extra-Large-Seeded and Double-Podded Traits in Chickpea (Cicer arietinum L.). Agronomy. 2020; 10(6):901. https://doi.org/10.3390/agronomy10060901
Chicago/Turabian StyleKivrak, Kamile Gul, Tuba Eker, Hatice Sari, Duygu Sari, Kadir Akan, Bilal Aydinoglu, Mursel Catal, and Cengiz Toker. 2020. "Integration of Extra-Large-Seeded and Double-Podded Traits in Chickpea (Cicer arietinum L.)" Agronomy 10, no. 6: 901. https://doi.org/10.3390/agronomy10060901