
Increased Soil Fertility in Tea Gardens Leads to Declines in Fungal Diversity and Complexity in Subsoils



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Soil Fertility Index Analysis
2.3. DNA Amplicon and Illumina Sequencing of Fungal Communities
2.4. Statistical Analyses
3. Results
3.1. Soil Fertility Index
3.2. Fungal Diversity
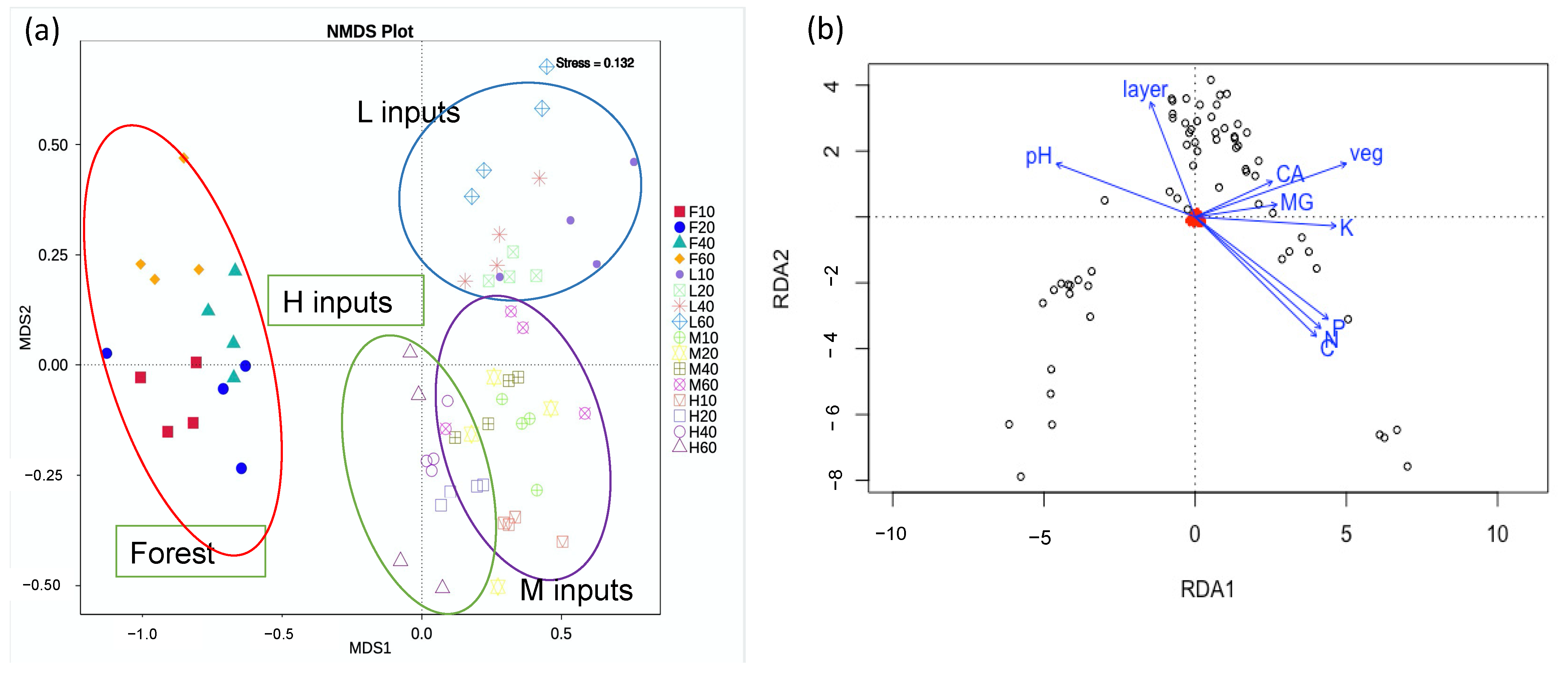
3.3. Fungal Community Composition
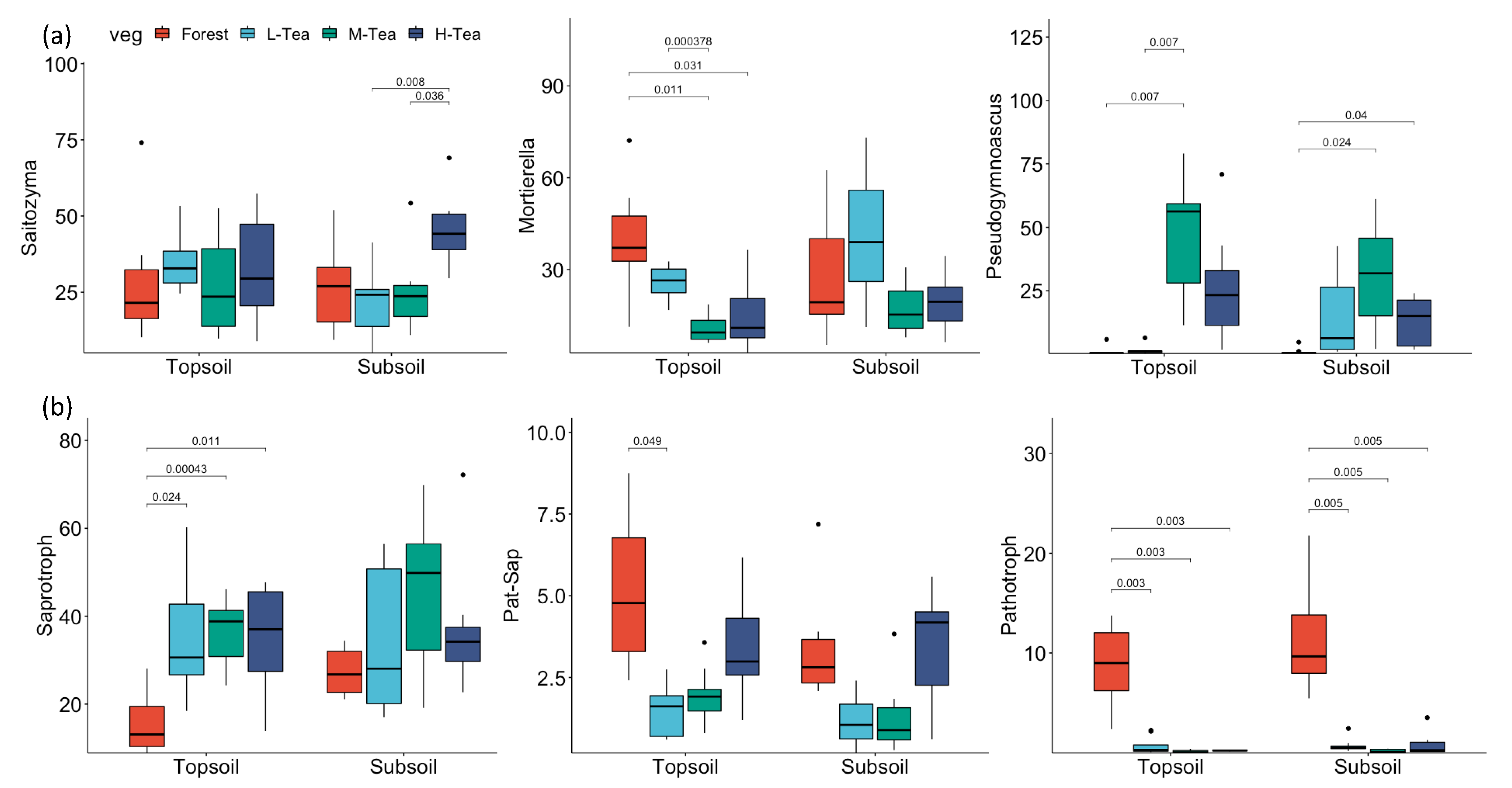
3.4. Functional Guild Composition
3.5. Fungal Co-Occurrence Networks
4. Discussion
4.1. Fertilizer Influences Soil Fungal Diversity and Richness in the Tea Garden
4.2. Fertilizer Influences Soil Fungal Comunitity Stucture and Functional Guilds in the Tea Garden
4.3. Soil Fungal Vertical Networks Complexity Varied along Soil Depths
4.4. Soil Properties Correlated with Soil Fungal Diversity Varied along Soil Depths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Yuan, Y.; Zhang, Y.; Shi, Y.; Hu, G.; Zhu, J.; Rogers, K.M. Geographical traceability of Chinese green tea using stable isotope and multi-element chemometrics. Rapid Commun. Mass Spectrom. 2019, 33, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, Y.; Long, Z.; Hu, S.; Zhang, Y.; Jiang, H. Effects of exotic plantation forests on soil edaphon and organic matter fractions. Sci. Total Environ. 2018, 626, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Haerdter, R.; Gerendás, J. Impact of nitrogen supply on carbon/nitrogen allocation: A case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. Plant Biol. 2010, 12, 724–734. [Google Scholar] [CrossRef]
- Han, B.; Zhang, Q.-H.; Byers, J.A. Attraction of the tea aphid, Toxoptera aurantii, to combinations of volatiles and colors related to tea plants. Èntomol. Exp. Appl. 2012, 144, 258–269. [Google Scholar] [CrossRef]
- Yan, P.; Shen, C.; Fan, L.; Li, X.; Zhang, L.; Zhang, L.; Han, W. Tea planting affects soil acidification and nitrogen and phosphorus distribution in soil. Agric. Ecosyst. Environ. 2018, 254, 20–25. [Google Scholar] [CrossRef]
- Ji, L.; Ni, K.; Wu, Z.; Zhang, J.; Yi, X.; Yang, X.; Ling, N.; You, Z.; Guo, S.; Ruan, J. Effect of organic substitution rates on soil quality and fungal community composition in a tea plantation with long-term fertilization. Biol. Fertil. Soils 2020, 56, 633–646. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.; Kuzyakov, Y.; Wang, H.; Fu, X.; Yang, Y.; Chen, F.; Dungait, J.A.; Green, S.M.; Fang, X. Responses of C-, N- and P-acquiring hydrolases to P and N fertilizers in a subtropical Chinese fir plantation depend on soil depth. Appl. Soil Ecol. 2019, 150, 103465. [Google Scholar] [CrossRef]
- Chen, W.-C.; Ko, C.-H.; Su, Y.-S.; Lai, W.-A.; Shen, F.-T. Metabolic potential and community structure of bacteria in an organic tea plantation. Appl. Soil Ecol. 2020, 157, 103762. [Google Scholar] [CrossRef]
- Van Der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting Soil pH Effects on Fungal and Bacterial Growth Suggest Functional Redundancy in Carbon Mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfante, P. The future has roots in the past: The ideas and scientists that shaped mycorrhizal research. New Phytol. 2018, 220, 982–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenet, B.; Lenhart, K.; Leloup, J.; Giusti-Miller, S.; Pouteau, V.; Mora, P.; Nunan, N.; Abbadie, L. The impact of long-term CO2 enrichment and moisture levels on soil microbial community structure and enzyme activities. Geoderma 2012, 170, 331–336. [Google Scholar] [CrossRef]
- Huang, Y.; Guenet, B.; Ciais, P.; Janssens, I.A.; Soong, J.L.; Wang, Y.; Goll, D.; Blagodatskaya, E.; Huang, Y. ORCHIMIC (v1.0), a microbe-mediated model for soil organic matter decomposition. Geosci. Model Dev. 2018, 11, 2111–2138. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.Y.; Koal, P.; Gerl, G.; Schroll, R.; Joergensen, R.G.; Munch, J.C. Response of water extractable organic matter and its fluorescence fractions to organic farming and tree species in poplar and robinia-based alley cropping agroforestry systems. Geoderma 2016, 290, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Högberg, M.N.; Blaško, R.; Bach, L.H.; Hasselquist, N.J.; Egnell, G.; Näsholm, T.; Högberg, P. The return of an experimentally N-saturated boreal forest to an N-limited state: Observations on the soil microbial community structure, biotic N retention capacity and gross N mineralisation. Plant Soil 2014, 381, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Cira, N.J.; Pearce, M.T.; Quake, S.R. Neutral and selective dynamics in a synthetic microbial community. Proc. Natl. Acad. Sci. USA 2018, 115, E9842–E9848. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, M.A. Everything is everywhere: But the environment selects’: Ubiquitous distribution and ecological determinism in microbial biogeography. Stud. Hist. Philos. Sci. Part C Stud. Hist. Philos. Biol. Biomed. Sci. 2008, 39, 314–325. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Nelson, M.; Helgason, T.; Dytham, C.; Fitter, A.H. Relative roles of niche and neutral processes in structuring a soil microbial community. ISME J. 2009, 4, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Baah-Acheamfour, M.; Carlyle, C.N.; Bissett, A.; Richardson, A.E.; Siddique, T.; Bork, E.W.; Chang, S.X. Determinants of bacterial communities in Canadian agroforestry systems. Environ. Microbiol. 2015, 18, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling Interactions in the Microbiome: A Network Perspective. Trends Microbiol. 2016, 25, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Hu, Q.; Cheng, Y.; Bai, L.; Liu, Z.; Xiao, W.; Gong, Z.; Wu, Y.; Feng, K.; Deng, Y.; et al. Application of organic fertilizer improves microbial community diversity and alters microbial network structure in tea (Camellia sinensis) plantation soils. Soil Tillage Res. 2019, 195, 104356. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Kou, Y.; Yao, M.; He, Z.; Li, X. Distinct mechanisms shape soil bacterial and fungal co-occurrence networks in a mountain ecosystem. FEMS Microbiol. Ecol. 2020, 96, fiaa030. [Google Scholar] [CrossRef]
- Guo, S.; Xu, Y.; He, C.; Wu, S.; Ren, C.; Han, X.; Feng, Y.; Ren, G.; Yang, G. Differential responses of soil quality in revegetation types to precipitation gradients on the Loess Plateau. Agric. For. Meteorol. 2019, 276–277, 107622. [Google Scholar] [CrossRef]
- Fontaine, S.; Henault, C.; Aamor, A.; Bdioui, N.; Bloor, J.; Maire, V.; Mary, B.; Revaillot, S.; Maron, P. Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect. Soil Biol. Biochem. 2011, 43, 86–96. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Chang, C.-H.; Szlavecz, K.; Buyer, J.S. Species-specific effects of earthworms on microbial communities and the fate of litter-derived carbon. Soil Biol. Biochem. 2016, 100, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Tun, H.M.; Jahan, M.; Zhang, Z.; Kumar, A.; Fernando, W.G.D.; Farenhorst, A.; Khafipour, E. Comparison of DNA-, PMA-, and RNA-based 16S rRNA Illumina sequencing for detection of live bacteria in water. Sci. Rep. 2017, 7, 5752. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.S.; Karlen, D.L.; Cambardella, C.A. The soil management assessment framework: A quantitative soil quality evaluation method. Soil Sci. Soc. Am. J. 2004, 68, 1945–1962. [Google Scholar] [CrossRef]
- Shukla, M.K.; Lal, R.; Ebinger, M. Determining soil quality indicators by factor analysis. Soil Till. Res. 2006, 87, 194–204. [Google Scholar] [CrossRef]
- Karlen, D.L.; Ditzler, C.A.; Andrews, S.S. Soil quality: Why and how? Geoderma 2003, 114, 145–156. [Google Scholar] [CrossRef]
- Shang, Q.; Ling, N.; Feng, X.; Yang, X.; Wu, P.; Zou, J.; Shen, Q.; Guo, S. Soil fertility and its significance to crop productivity and sustainability in typical agroecosystem: A summary of long-term fertilizer experiments in China. Plant Soil 2014, 381, 13–23. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.A.; Gelfand, D.H.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal Rna Genes for Phylogenetics. In PCR Protocols; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Reeder, J.; Knight, R. Rapidly denoising pyrosequencing amplicon reads by exploiting rank-abundance distributions. Nat. Methods 2010, 7, 668–669. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruitt, K.D.; Brown, G.R.; Hiatt, S.M.; Thibaud-Nissen, F.; Astashyn, A.; Ermolaeva, O.; Farrell, C.M.; Hart, J.; Landrum, M.J.; McGarvey, K.M.; et al. RefSeq: An update on mammalian reference sequences. Nucleic Acids Res. 2013, 42, D756–D763. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Williams, L.J.; Vincent, J.B.; Stefanski, A.; Cavender-Bares, J.; Messier, C.; Paquette, A.; Gravel, D.; Reich, P.; Kennedy, P.G. Ectomycorrhizal fungal diversity and saprotrophic fungal diversity are linked to different tree community attributes in a field-based tree experiment. Mol. Ecol. 2016, 25, 4032–4046. [Google Scholar] [CrossRef]
- Yang, X.; Yu, H.; Zhang, T.; Guo, J.; Zhang, X. Arbuscular Mycorrhizal Fungi Improve the Antioxidative Response and the Seed Production of Suaedoideae Species Suaeda physophora Pall Under Salt Stress. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Fry, E.L.; Eldridge, D.J.; de Vries, F.T.; Manning, P.; Hamonts, K.; Kattge, J.; Boenisch, G.; Singh, B.K.; Bardgett, R.D. Plant attributes explain the distribution of soil microbial communities in two contrasting regions of the globe. New Phytol. 2018, 219, 574–587. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: http://ieg4.rccc.ou.edu/mena (accessed on 16 June 2022).
- Baldrian, P.; Trögl, J.; Frouz, J.; Šnajdr, J.; Valášková, V.; Merhautová, V.; Cajthaml, T.; Herinková, J. Enzyme activities and microbial biomass in topsoil layer during spontaneous succession in spoil heaps after brown coal mining. Soil Biol. Biochem. 2008, 40, 2107–2115. [Google Scholar] [CrossRef]
- Klaubauf, S.; Inselsbacher, E.; Zechmeister-Boltenstern, S.; Wanek, W.; Gottsberger, R.; Strauss, J.; Gorfer, M. Molecular diversity of fungal communities in agricultural soils from Lower Austria. Fungal Divers. 2010, 44, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa-Hernández, M.A.; Roy, J.; Hempel, S.; Kautz, T.; Köpke, U.; Uksa, M.; Schloter, M.; Caruso, T.; Rillig, M.C. Subsoil arbuscular mycorrhizal fungal communities in arable soil differ from those in topsoil. Soil Biol. Biochem. 2018, 117, 83–86. [Google Scholar] [CrossRef]
- Baumert, V.L.; Vasilyeva, N.A.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root exudates induce soil macroaggregation facilitated by fungi in subsoil. Front. Environ. Sci. 2018, 6, 140. [Google Scholar] [CrossRef]
- Shi, L.-L.; Mortimer, P.E.; Slik, J.W.F.; Zou, X.-M.; Xu, J.; Feng, W.-T.; Qiao, L. Variation in forest soil fungal diversity along a latitudinal gradient. Fungal Divers. 2014, 64, 305–315. [Google Scholar] [CrossRef]
- Lin, X.; Feng, Y.; Zhang, H.; Chen, R.; Wang, J.; Zhang, J.; Chu, H. Long-Term Balanced Fertilization Decreases Arbuscular Mycorrhizal Fungal Diversity in an Arable Soil in North China Revealed by 454 Pyrosequencing. Environ. Sci. Technol. 2012, 46, 5764–5771. [Google Scholar] [CrossRef]
- Dai, M.; Bainard, L.D.; Hamel, C.; Gan, Y.; Lynch, D. Impact of Land Use on Arbuscular Mycorrhizal Fungal Communities in Rural Canada. Appl. Environ. Microbiol. 2013, 79, 6719–6729. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Nadkarni, N.M. Growth measurements of saprotrophic fungi and bacteria reveal differences between canopy and forest floor soils. Soil Biol. Biochem. 2009, 41, 862–865. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, P.; Chi, Y.; Wang, N.; Wang, P.; Liu, N.; Cai, D.; Wu, Z.; Zhong, N. Controlling the Hydrolysis and Loss of Nitrogen Fertilizer (Urea) by using a Nanocomposite Favors Plant Growth. ChemSusChem 2017, 10, 2068–2079. [Google Scholar] [CrossRef]
- Negassa, W.; Baum, C.; Schlichting, A.; Müller, J.; Leinweber, P. Small-Scale Spatial Variability of Soil Chemical and Biochemical Properties in a Rewetted Degraded Peatland. Front. Environ. Sci. 2019, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Feng, W.; Jing, X.; Zang, H.; Mortimer, P.; Zou, X. Contrasting responses of soil fungal communities and soil respiration to the above- and below-ground plant C inputs in a subtropical forest. Eur. J. Soil Sci. 2018, 70, 751–764. [Google Scholar] [CrossRef]
- Rovira, P.; Vallejo, V. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: An acid hydrolysis approach. Geoderma 2002, 107, 109–141. [Google Scholar] [CrossRef]
- Kallenbach, C.; Grandy, A.; Frey, S.; Diefendorf, A. Microbial physiology and necromass regulate agricultural soil carbon accumulation. Soil Biol. Biochem. 2015, 91, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Dannenmann, M.; Simon, J.; Gasche, R.; Holst, J.; Naumann, P.S.; Kögel-Knabner, I.; Knicker, H.; Mayer, H.; Schloter, M.; Pena, R.; et al. Tree girdling provides insight on the role of labile carbon in nitrogen partitioning between soil microorganisms and adult European beech. Soil Biol. Biochem. 2009, 41, 1622–1631. [Google Scholar] [CrossRef]
- Weiher, E.; Freund, D.; Bunton, T.; Stefanski, A.; Lee, T.; Bentivenga, S. Advances, challenges and a developing synthesis of ecological community assembly theory. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2403–2413. [Google Scholar] [CrossRef]
- Ahmed, W.; Jing, H.; Kaillou, L.; Qaswar, M.; Khan, M.N.; Jin, C.; Geng, S.; Qinghai, H.; Yiren, L.; Guangrong, L.; et al. Changes in phosphorus fractions associated with soil chemical properties under long-term organic and inorganic fertilization in paddy soils of southern China. PLoS ONE 2019, 14, e0216881. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Banfield, C.C.; Sanaullah, M.; Gunina, A.; Dippold, M.A. Utilisation of mucilage C by microbial communities under drought. Biol. Fertil. Soils 2017, 54, 83–94. [Google Scholar] [CrossRef]
- Rosado, B.H.P.; Holder, C.D. The significance of leaf water repellency in ecohydrological research: A review. Ecohydrology 2012, 6, 150–161. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Fukasawa, Y.; Osono, T.; Takeda, H. Dynamics of physicochemical properties and occurrence of fungal fruit bodies during decomposition of coarse woody debris of Fagus crenata. J. For. Res. 2009, 14, 20. [Google Scholar] [CrossRef]
- Bartram, A.K.; Jiang, X.; Lynch, M.D.; Masella, A.P.; Nicol, G.; Dushoff, J.; Neufeld, J.D. Exploring links between pH and bacterial community composition in soils from the Craibstone Experimental Farm. FEMS Microbiol. Ecol. 2013, 87, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xu, R.; Hu, T.; Chen, J.; Zhang, Y.; Cao, X.; Wu, D.; Wu, Y.; Shen, Y. Soil-mediated effects of acidification as the major driver of species loss following N enrichment in a semi-arid grassland. Plant Soil 2017, 419, 541–556. [Google Scholar] [CrossRef]
- He, M.; Chen, W.-J.; Tian, L.; Shao, B.; Lin, Y. Plant-microbial synergism: An effective approach for the remediation of shale-gas fracturing flowback and produced water. J. Hazard. Mater. 2018, 363, 170–178. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, P.; Shen, C.; Zou, Z.; Fan, L.; Li, X.; Zhang, L.; Zhang, L.; Dong, C.; Fu, J.; Han, W.; et al. Increased Soil Fertility in Tea Gardens Leads to Declines in Fungal Diversity and Complexity in Subsoils. Agronomy 2022, 12, 1751. https://doi.org/10.3390/agronomy12081751
Yan P, Shen C, Zou Z, Fan L, Li X, Zhang L, Zhang L, Dong C, Fu J, Han W, et al. Increased Soil Fertility in Tea Gardens Leads to Declines in Fungal Diversity and Complexity in Subsoils. Agronomy. 2022; 12(8):1751. https://doi.org/10.3390/agronomy12081751
Chicago/Turabian StyleYan, Peng, Chen Shen, Zhenhao Zou, Lichao Fan, Xin Li, Liping Zhang, Lan Zhang, Chunwang Dong, Jianyu Fu, Wenyan Han, and et al. 2022. "Increased Soil Fertility in Tea Gardens Leads to Declines in Fungal Diversity and Complexity in Subsoils" Agronomy 12, no. 8: 1751. https://doi.org/10.3390/agronomy12081751
APA StyleYan, P., Shen, C., Zou, Z., Fan, L., Li, X., Zhang, L., Zhang, L., Dong, C., Fu, J., Han, W., & Shi, L. (2022). Increased Soil Fertility in Tea Gardens Leads to Declines in Fungal Diversity and Complexity in Subsoils. Agronomy, 12(8), 1751. https://doi.org/10.3390/agronomy12081751