
Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review


1
College of Environmental and Safety Engineering, Fuzhou University, Fuzhou 350108, China
2
Key Laboratory of Urban Environment and Health, Institute of Urban Environment, Chinese Academy of Sciences, Xiamen 361021, China
3
Zhejiang Key Laboratory of Urban Environmental Processes and Pollution Control, CAS Haixi Industrial Technology Innovation Center in Beilun, Ningbo 315830, China
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(8), 1900; https://doi.org/10.3390/agronomy12081900
Submission received: 13 July 2022
/
Revised: 9 August 2022
/
Accepted: 12 August 2022
/
Published: 13 August 2022
Abstract
:Intercropping systems can flexibly use resources such as sunlight, heat, water, and nutrients in time and space, improve crop yield and land utilization rates, effectively reduce continuous cropping obstacles and the occurrence of diseases and insect pests, and control the growth of weeds. Thus, intercropping is a safe and efficient ecological planting mode. The legume–cereal intercropping system is the most common planting combination. Legume crops fix nitrogen from the atmosphere through their symbiotic nitrogen fixation abilities, and the fixed nitrogen can be transferred to and utilized by cereal crops in various ways. The symbiotic nitrogen fixation efficiency of legume crops was improved by reducing the inhibition of soil nitrogen on nitrogenase activity through competitive absorption of soil nitrogen. However, the effects of nitrogen transformation and distribution in intercropping systems and microbial community structure characteristics on nitrogen transfer need to be further explored. In this review, (ⅰ) we present the transformation and distribution of nitrogen in the legume–cereal intercropping system; (ⅱ) we describe the soil microbial community characteristics in intercropping systems; and (ⅲ) we discuss the advantages of using modern biological molecular techniques to study soil microorganisms. We conclude that intercropping can increase the diversity of soil microorganisms, and the interaction between different plants has an important impact on the diversity and composition of the bacterial and fungal communities. The extensive application of modern biological molecular techniques in soil microbial research and the great contribution of intercropping systems to sustainable agriculture are particularly emphasized in this review.
1. Introduction
Nitrogen is the basic substance in the protoplasm composition of plant cells and the basis of plant life activities; it is the most important nutrient element for crop growth [1]. Nitrogen is generally distributed in the most active parts of plants (new leaves, meristem, reproductive organs, etc.). Therefore, nitrogen supply directly affects the growth and development of crops. Nitrogen demands are critical at certain stages of crop growth, such as the tillering stage and spike differentiation stage of cereal crops, flower and boll stage of cotton, rapid growth of cash crops, and formation stages of economic products. Sufficient nitrogen nutrition at these stages can promote crop growth and increase yield.
After the application of chemical fertilizer, crop yield and nitrogen uptake capacity gradually increased, but yield per hectare and marginal benefit gradually decreased. It is evident that this nitrogen, which is not absorbed by crops, enters the environment through ammonia volatilization, nitrification, denitrification, leaching, and runoff [2]. Therefore, the higher the nitrogen application rate is, the higher the nitrogen cycling intensity is. Accordingly, a high crop yield, low efficiency of nitrogen use, and high loss will occur.
The legume–cereal intercropping pattern has a long planting history in China [3] and has been widely used in food crop production systems to obtain ecological and agronomic benefits. In legume–cereal intercropping systems, legumes can effectively fix nitrogen in the atmosphere, and this fixed nitrogen can be used by cereal crops [4]. The symbiotic nitrogen fixation efficiency of legume crops was improved by reducing the inhibition of soil nitrogen on nitrogenase activity through competitive absorption of soil nitrogen [5]. Legumes can provide up to 73% of the nitrogen required by cereals [6], depending on stand age, background soil available nitrogen, cropping system, and crop species [7]. Intercropping intermediate wheatgrass (IWG) with perennial legumes can reduce nitrogen inhibition and increase the soil total nitrogen pool to achieve long-term release, while avoiding negative factors related to fertilizer use [8]. In addition, the legume–cereal intercropping system can increase environmental outcomes, such as increased soil carbon retention and improved nitrogen use efficiency, thereby reducing nitrate leaching. The basis of these benefits may be attributed to complementary nitrogen use and nitrogen transfer between these two crops in intercropping systems. Intercropping systems can effectively reduce the use of chemical nitrogen fertilizer, which is of great significance in solving the environmental problems caused by the irrational application of nitrogen fertilizer. Therefore, it is very important to review the nitrogen fixation rate and nitrogen transfer and distribution of legumes in legume–cereal intercropping systems for future sustainable agricultural development.
In legume–cereal intercropping, microorganisms cross-migrate from the roots of one plant to those of another. In fact, intercropping with legumes can improve soil properties by enhancing the functional diversity of the soil microbial community, thus improving plant growth. Clearly, the soil microbial community is important to soil function because it plays a role in mineral nutrient cycling, organic turnover, nitrification, mineralization, and soil structure formation, which are essential for plant growth.
2. Transformation and Distribution of Nitrogen in the Legume–Cereal Intercropping System
2.1. Fixation of Nitrogen in Legumes
Considerable amounts of fertilizer input have failed to produce good agricultural results and have had low nitrogen utilization rates, resulting in serious negative effects on soil, air, water, and other ecological environments [9]. Biological nitrogen fixation has been implemented in solving agricultural production and environmental problems [10]. In 1886, Helligel found that the nodules on leguminous roots had the ability to assimilate N2 [11]. A large number of studies later confirmed that rhizobia on legume crops can convert N2 in the air into synthetic ammonia without demanding reaction conditions [12,13]. The nitrogen fixed by legumes is not only used for their own growth but also provides nitrogen-containing organic matter to the accompanying plants. Symbiotic nitrogen fixation converts atmospheric N2 into ammonia under the action of nitrogenase [14,15], and this process is the result of complex interactions between host plants and rhizobia. There is a symbiotic relationship between rhizobia and legumes. A large part of the nitrogen and nutrients required for rhizobium growth and physiological metabolic activities come from legume plants, and the amount of cogrowth nitrogen fixation of mature legume plants accounts for 25–66% of the total nitrogen. Symbiotic nitrogen fixation of legume crops can reach 75–150 kg per hectare per year and even 300 kg ha−1 when conditions are favorable [16]. Legume nodules act as a small nitrogen-processing plant, and taking advantage of this symbiotic relationship can reduce the dependence on chemical fertilizers and reduce environmental pollution caused by excessive fertilization.
In the farmland system, bean intercropping with cereal has obvious advantages compared to outputs achieved with monoculture, mainly because during bean and cereal common growth, the roots of the two crops become interwoven with each other, and the competition for root nutrition is relatively intense in cereals. The competitiveness of cereal crops in obtaining soil nitrogen is further strengthened, and the obstacle of “nitrogen repression” of legume root nodules is reduced so that both legume and cereal crops show obvious yield advantages [17]. The formation of an intercropping advantage is closely related to the operation of nitrogen fertilizer in the intercropping system.
Nitrogen application amounts had a substantial impact on the proportion of nitrogen derived from the atmosphere (%Ndfa) and the amount of nitrogen fixed (Ndfa) by intercropped alfalfa [18]. With the addition of nitrogen fertilizer (N1, 200 mg N kg−1 soil), the %Ndfa and Ndfa of intercropped alfalfa were lowered by 41% and 46%, respectively. Cereals compete more aggressively for nutrient absorption than legumes in the legume–cereal intercropping system [19] because of their faster root growth rate and wider root distribution, which reduces the nitrogen content in the rhizosphere and improves the nitrogen fixation capacity of legumes. Pasture–legume intercropping improved the efficiency of biological nitrogen fixation (BNF), and the amount of fixed nitrogen increased by 18% compared with that of legume crops alone [20,21]. The quantity and biomass of soybean root nodules were significantly affected by the nitrogen delivery rate in a soybean–sugarcane intercropping system [22]. In comparison to conventional nitrogen intake, the nodule number increased by 2.1–4.1 times and nodule biomass increased by 1.8–2.0 times when nitrogen input was lowered. The same conclusion showed that the nitrogen fixation ratio of pea under the N1 (225 kg N ha−1) nitrogen level was significantly lower than that under the N0 level because fertilization increased soil mineral nitrogen and then inhibited symbiotic N2 fixation [23]. Hei et al. [24] designed a field experiment of rice–water mimosa intercropping with zero nitrogen (ZN, 0 kg N ha−1), reduced nitrogen (RN, 140 kg N ha−1), and conventional N (CN, 180 kg N ha−1). The study found that mimosa had the highest biological nitrogen fixation efficiency under the RN treatment. In conclusion, the nitrogen fixation capacity, nitrogenase activity, and nitrogen transformation and distribution in intercropping systems of different regions and different crop species were also inconsistent. Therefore, in the process of improving rational nitrogen utilization through legume–cereal intercropping, it is worth studying how to stimulate legume biological nitrogen fixation to the maximum extent and accurately quantify nitrogen distribution in legume–cereal intercropping systems.
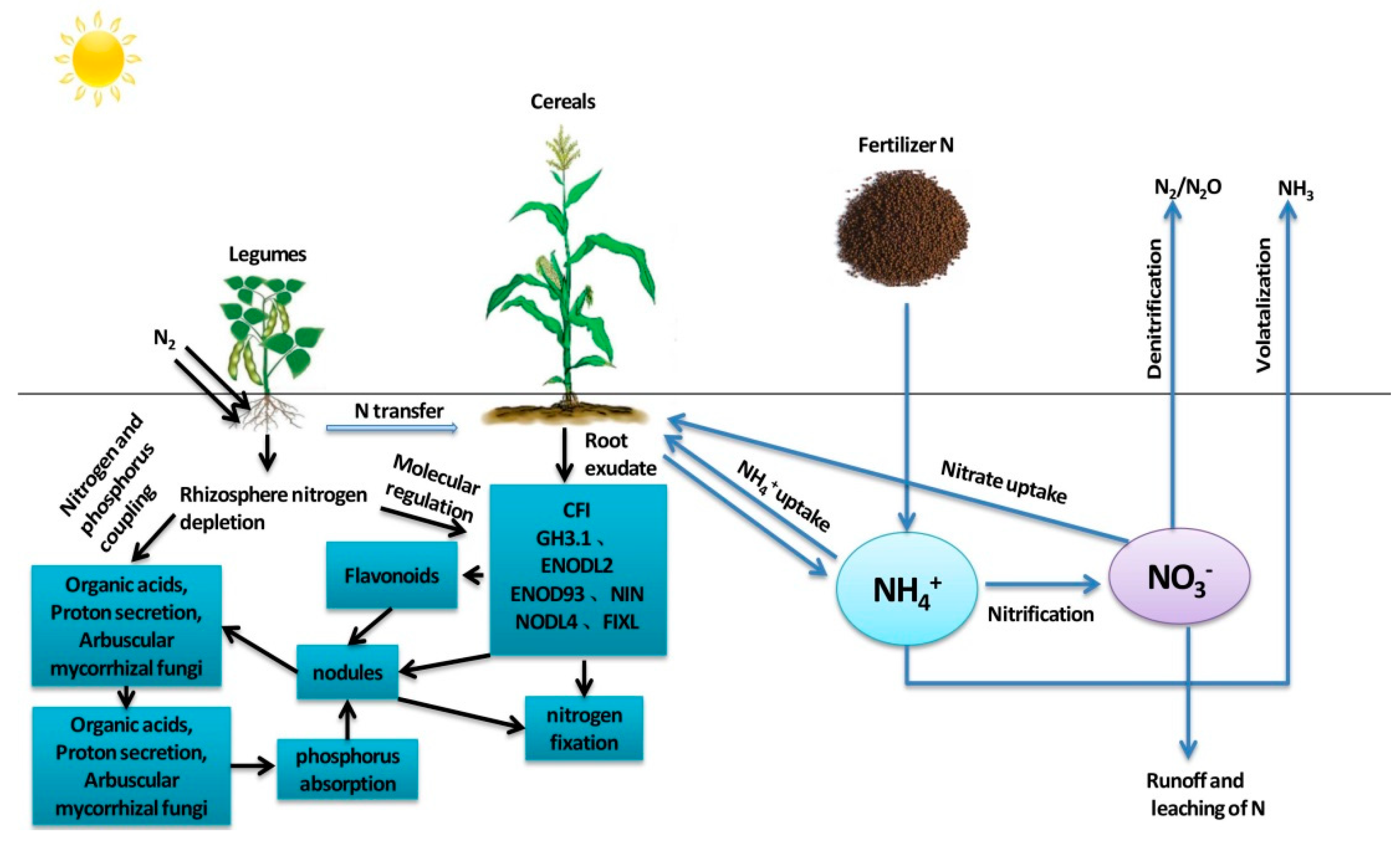
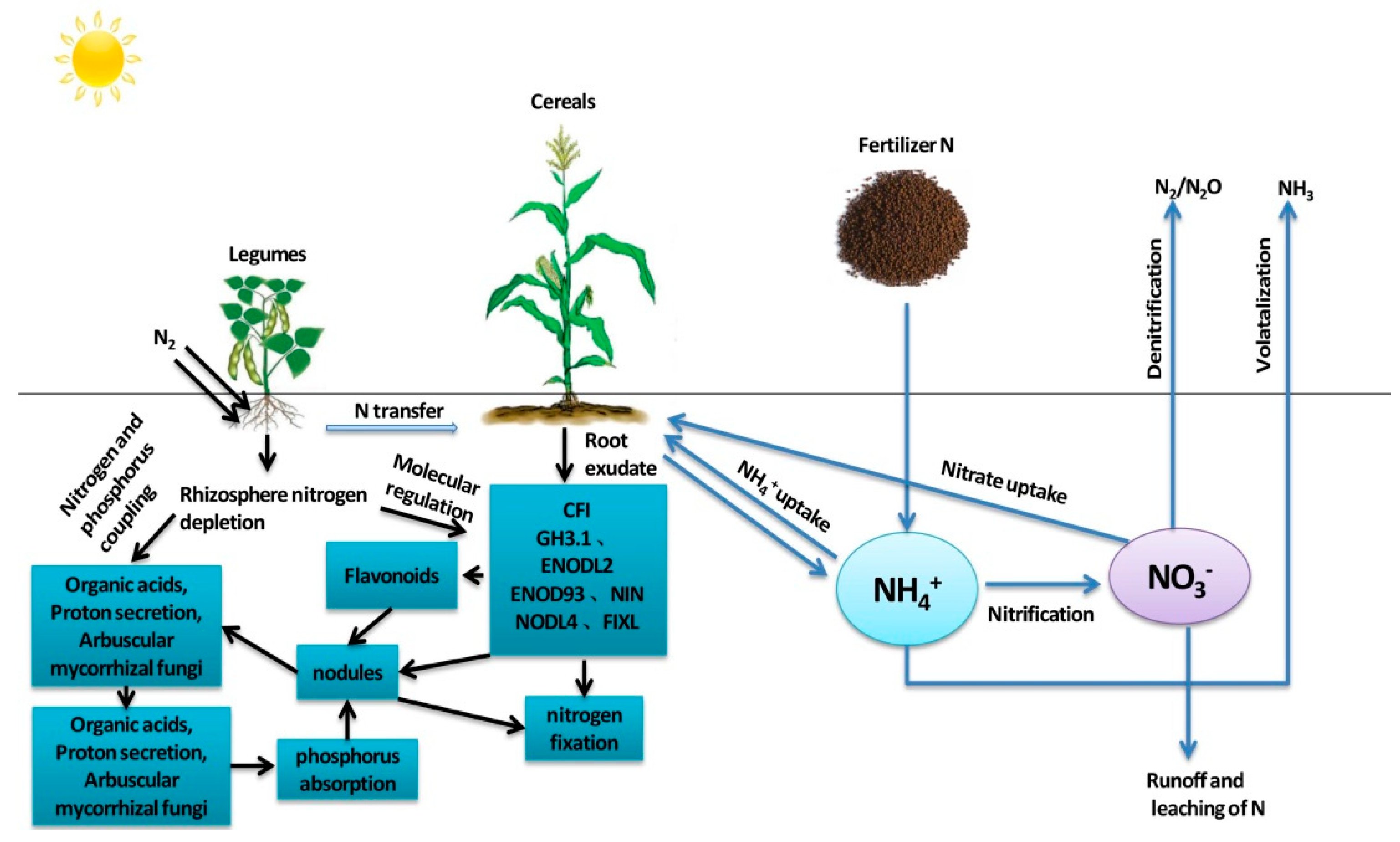
Legumes have a stronger proton releasing capacity than cereals and can significantly acidify the rhizosphere, thus facilitating the activation and absorption of soil insoluble phosphorus [25], meeting the supply of adenosine triphosphate (ATP) required for nitrogen fixation in legumes, and maintaining a stable nitrogen and phosphorus stoichiometric relationship in intercropped legumes (Figure 1). Amarildo et al. [26] found that legumes could replace nitrogen application in the case of low phosphate capacity and no nitrogen application. Seeding and incorporation of phosphorus fertilizer in guinea grass cultivation not only improves pasture yield but also improves nitrogen use efficiency regardless of phosphorus solubility. Intercropping broad bean with maize significantly boosted grain output and aboveground biomass compared to wheat–maize in another field trial at low P rates (75 kg P2O5 ha−1) but not at high P rates (>112.5 kg P2O5 ha−1). Broad beans overproduce because they have a different growing season and rooting depth than maize, so intercropping on low-phosphorus soils can greatly increase yields [27].
2.2. Nitrogen Uptake by Cereals in Intercropping Systems
Legume plants can transfer a certain amount of nitrogen to cereal crops in intercropping systems [28]. The absorption of legume-fixed nitrogen by cereal crops mainly occurs through the decomposition of legume litter and the deposition of root and nodule tissue [29] (Figure 1). Previous studies have shown that the transfer rate of nitrogen from legumes to cereal in intercropping systems ranges from 0 to 73%, depending on the species and planting method of legumes and cereals [6]. Maize + legume–wheat intercropping was more beneficial than monoculture and could save 25% of the nitrogen for subsequent wheat [30]. All intercropping treatments left enough residual fertility to significantly increase the yield of the wheat crop without fertilization. From a cropping system perspective, legume intercropping provides significant nitrogen savings, resulting in lower tillage costs and improved soil health indicators.
Most of the earliest studies on nitrogen transfer and distribution in intercropping systems focused on mixed pasture systems [31]. Legume–cereal intercropping systems transfer nitrogen from legumes to adjacent cereal crops through root and mycorrhizal networks, which improve nitrogen uptake of cereals [32]. Cereal crops can rapidly consume nitrogen in the soil, and their root exudates can induce nodulation in the root system of broad bean, thus increasing the symbiotic nitrogen fixation rate of legumes and ultimately increasing the soil nitrogen pool [33]. Researchers established a basic nitrogen cycling network between crops long ago, but the exact proportions of nitrogen distribution were unclear. The nitrogen distribution in legume–cereal intercropping systems could be analyzed by using 15N isotope-labeled nitrogen and leaf labeling techniques. Yong et al. [34] used 15N isotope dilution labeling and a direct 15N leaf feeding measurement to conduct a pot experiment with a wheat–maize–soybean rotation and discovered that there was bidirectional nitrogen transfer in the symbiotic stage of crops, and the amount of 15N transfer (N transfer) and percentage (%NT) decreased as the soil fertilization level increased. Maize-to-wheat nitrogen transfer was 16.1–163.0% higher than wheat-to-maize nitrogen transfer. The nitrogen transfer from soybean to maize was 1.7–6.0 times that of maize to soybean, and the %NT from soybean to maize was 6.7–22.2 times that of maize to soybean. The researchers concluded that bidirectional nitrogen transfer is the main factor in improving crop nitrogen use efficiency. Ledgard et al. [35] studied nitrogen transfer in a clover–ryegrass intercropping system by using the leaf feeding method of 15N, and the results showed that 2.2% of nitrogen in clover was directly transferred to ryegrass. In a two-year indoor pot experiment, Shao et al. [36] measured the influence of root contact on nitrogen transfer in a maize–alfalfa intercropping system. The majority of nitrogen transfer occurred in one direction, from alfalfa to maize. Nitrogen transfer from alfalfa to maize was 204.56 mg pot−1 (without a root barrier) and 165.13 mg pot−1 (with a nylon net barrier) in the N0 treatment, accounting for 4.72 and 4.48 percent of the total nitrogen accumulation in maize, respectively. Nitrogen transfer from alfalfa to maize was 197.70 mg pot−1 (without root barrier) and 139.04 mg pot−1 (with nylon net barrier) under the N1 (10.2 g N pot−1) treatment, accounting for 3.64% and 2.36% of the total nitrogen accumulation in maize, respectively. Furthermore, regardless of the supplemented nitrogen dosage, the amount of nitrogen transfer without a root barrier was 1.24 to 1.42 times that with a nylon net barrier. The findings emphasize the significance of root interactions in improving nitrogen transfer in maize–alfalfa intercropping systems.
3. Soil Microbial Community Characteristics in the Legume–Cereal Intercropping System
Soil microorganisms are the link and bridge between aboveground and underground ecosystems and play an important role in the regulation of soil ecological functions, the transformation of organic matter in soil, and the promotion of carbon and nitrogen cycles [37]. The intercropping system affected not only the physical and chemical properties of the soil but also the microorganisms in the soil. Due to the different root distributions of different crops, nutrients in different spaces can be fully utilized to change the physiological characteristics of crops and root exudates, thus indirectly affecting the composition of the soil microbial community and soil enzyme activities and improving soil fertility. Plant roots are the best space for microbial communities to gather [38]. Phenotypic traits such as root structure may affect microbial growth and reproduction, and plant roots can secrete primary metabolites such as organic acids or amino acids [39], which play an important role in shaping the rhizosphere environment by changing the chemical composition of soil near plant roots and serving as substrates for the growth of some soil microorganisms [40]. The composition of plant root exudates changes qualitatively and quantitatively according to the plant’s nutritional status, growth stage, and even the root position in time and space [41]. These secretions produce strong selection pressures in the rhizosphere, prompting plants to drive the selection of specific rhizosphere microbial communities [42].
3.1. Soil Microbial Composition and Diversity in the Legume–Cereal Intercropping System
Intercropping systems have a greater diversity of roots and residues, which improves the energy supply of soil microbial biomass by releasing exudates such as amino acids and organic acids [43]. As a result, the effects of these leftovers on soil microbial biomass in intercropping were more noticeable than those in monocropping. The diversity of the rhizosphere soil microbial community structure in the intercropping system was generally higher than that in the monocropping system [44]. Common networks dominated by Acidobacteria, Alphaproteobacteria, and Bacteroidetes were found in wheat–pea intercropping systems, and these findings suggest a more complex rhizosphere bacterial network in intercropping [45]. When the cooccurrence network between populations becomes more complex, the functional expression of the subsurface microbiome increases [46]. Bean–oat intercropping significantly increased the diversity of nitrogen-fixing microbes and changed their community composition [47]. Legume–cereal intercropping significantly altered the composition and structure of functional microbial communities, particularly those microorganisms involved in nitrogen transformation in the rhizosphere soil, such as nitrogen fixation, ammonia oxidation, denitrification, nitrous oxide reduction, and organic nitrogen decomposition [48]. Strip intercropping methods have also been demonstrated to increase soil nitrogen sequestration by improving biological nitrogen fixation and lowering greenhouse gas emissions in recent research [49]. Yu et al. [37] evaluated changes in the microbial composition, structure, and abundance of nitrogen cycling genes in sweet maize intercropped with soybean in tropical and subtropical regions under specific conditions for the first time. It was found that maize–soybean intercropping could promote the soil microbial community. The composition of the arbuscular mycorrhizal fungal community was affected by the planting method and nitrogen application rate [50]. Previous studies showed that intercropping had positive effects on root colonization of arbuscular mycorrhizal fungi (AMF) [43]. The expression of amoA, nirK, nirS, and nosZ was found to be closely connected to the structure of the bacterial community, suggesting that bacterial community structure may have a substantial impact on soil nitrification, denitrification, and nitrogen fixation. AMF of adjacent maize crops could reduce N2O emissions from soybean stubble mainly by changing the abundance and community structure of denitrifying bacteria [51]. The abundance of the nifH gene was significantly positively correlated with planting methods and nitrogen fixation rate. Sugarcane–peanut intercropping significantly increased the Shannon’s diversity index of nitrogen-fixing bacteria in sugarcane crops, and the rhizosphere bacterial richness in sugarcane–peanut intercropping increased by 9.28% compared with monoculture sugarcane [52]. The intercropping roots of the two plants changed the soil physical and chemical properties and microbial community structure and increased the content of soil available nutrients [53]. In conclusion, legume–cereal intercropping, as a composite population formed by the combination of different crop species, has a great impact on the number, biomass, population structure, and diversity of rhizosphere microorganisms (Table 1 and Table 2).
Changes in soil microbial composition and quantity affect crop growth, crop yield, crop nutrient absorption, and pest control. Compared with monoculture, the number of soil fungi, bacteria, and actinomycetes in soybean–sugarcane intercropping increased by 115.5%, 43.6%, and 57.3%, respectively [54]. A field experiment was conducted with a split-pot design to study the changes in rhizosphere microbial diversity driven by interspecific interactions in a maize–soybean intercropping system through Illumina MiSeq sequencing [55]. Intercropping had no significant effect on the α diversity of maize rhizosphere soil and root AMF but significantly increased the α diversity of soybean rhizosphere soil AMF. At the genus level, Glomus_f_Glomeraceae was the dominant strain of AMF in maize rhizosphere soil and roots, and its relative abundance was the highest. In contrast to the α diversity of root AMF, the relative abundance of Glomus_f_Glomeraceae in maize roots increased significantly with the increase in the nitrogen application rate. Intercropping significantly decreased the α diversity of ammonia-oxidizing archaea (AOA) and ammonia-oxidizing bacteria (AOB) in maize rhizosphere soil and significantly increased the α diversity of AOA in soybean rhizosphere soil.
A large number of studies have demonstrated that plant diversity must affect soil biological activity, particularly through the input of more diverse substrates in the soil, possibly affecting decomposer activity in plant growth [56]. Intercropping occurs when two or more crops are grown simultaneously in the same space for a substantial portion of their life cycle [31]. The simultaneous planting of different crops in the same soil space will inevitably lead to the diversity of root and mycorrhizal networks and affect the composition and quantity of microorganisms. Agroecosystem functioning encompasses not only a wide range of spatial interactions between plants and soil but also interactions that occur throughout time as a function of the plants’ and microorganisms’ developmental stages, as well as their association dynamics [57].

{kind=link}
Table 1.
Effects of legume–cereal intercropping on rhizosphere microorganisms of legumes.
| Types of Intercropping Crops | Soil Type | Microorganisms with Increased Proportion | Microorganisms with Reduced Proportion | Microorganisms with No Significant Change | Reference |
|---|---|---|---|---|---|
| Soybean–maize | Brown soil | Actinomycetes, Firmicutes, Corynebacter, Cysts, Halophile bacillus, Phagocytes | Proteobacteria, Bacteroidetes, Burkholderia, Desulphurobacter, Neisseria | [58] | |
| Soybean–sugarcane | Red soil | Streptomyces, Bacillus, Pantomyces, Enterobacter, Arthrobacter, Symbiotic bacteria rhizobia, Chinese rhizobia | [59] | ||
| Peanut–sugarcane | Red soil | Streptomyces, Bacillus, Pantomyces, Enterobacter, Arthrobacter, Symbiotic bacteria rhizobia, Chinese rhizobia | [59,60] | ||
| Pea–wheat | Loamy soil | Acidobacteria, Proteobacteria, Bacteroidetes, Chlorobacteria | [45] | ||
| Soybean–sugarcane | Red soil | Proteobacteria, Acidobacteria, Chlorobacteria, Actinomycetes, Bacteroidetes | [61] | ||
| Peanut–maize | Hydragric anthrosol | Actinobacteria, Acidobacteria | Gammaproteobacteria, Bacteroidetes, Firmicutes | Alphaproteobacteria | [62] |
| Fata beans–maize | Clay loam | Firmicutes, Bacteroidetes, Bacillus, Clostridium, Sporobacteria, Desulphuria, Alicyclobacter | Methylobacter,Sphingomonas | Acidobacteria, Floating mold, Blastomonas phylum | [63] |
| Mung bean–proso millet | Loess-like | Proteobacteria, Chlorobacteria, Blastomonas, Acidobacteria, Helicobacter nitrifying, Firmicutes | Actinomycetes | Phylum fungi, Ascomycetes, Morpita, basidiomycetes | [64] |
| Wild soybean–sorghum | Salined fluvo-aquic soil | Proteobacteria, Bacteroidetes | Firmicutes, Gemmatimonadetes | [65] |
Table 2.
Effects of legume–cereal intercropping on rhizosphere microorganisms of cereals.
| Types of Intercropping Crops | Soil Type | Microorganisms with Increased Proportion | Microorganisms with Reduced Proportion | Microorganisms with No Significant Change | Reference |
|---|---|---|---|---|---|
| Soybean–maize | Brown soil | Actinomycetes, Firmicutes, Desulphurobacter, Erythrococcus, Kinetospora, Bacillus, Bacillus, phagocyte | Proteobacteria, Achromatobacteria, Burkholderia, Pseudomonas, Aikenella | Bacteroidetes | [58] |
| Soybean–sugarcane | Red soil | Streptomyces, Bacillus, Pantomyces, Enterobacter, Arthrobacter, Symbiotic bacteria rhizobia, Chinese rhizobia | [59] | ||
| Peanut–sugarcane | Red soil | Streptomyces, Bacillus, Pantomyces, Enterobacter, Arthrobacter, Symbiotic bacteria rhizobia, Chinese rhizobia, Acidobacteria, Chloroflexi | Proteobacteria, Actinobacteria | [59,60] | |
| Pea–wheat | Loamy Soil | Acidobacteria, Proteobacteria, Bacteroidetes, Chlorobacteria | [45] | ||
| Soybean–sugarcane | Red soil | Proteobacteria, Acidobacteria, Chlorobacteria, Actinomycetes, Bacteroidetes | [61] | ||
| Peanut–maize | Hydragric anthrosol | Deltaproteobacteria, Acidobacteria, Chloroflexi, Gemmatimonadetes | Actinobacteria, | Alphaproteobacteria | [62] |
| Fata beans–maize | Clay loam | Firmicutes, Bacteroidetes, Bacillus, Clostridium, Sporobacteria, Desulphuria, Alicyclobacter | Methylobacter,Sphingomonas | Acidobacteria, Floating mold, Blastomonas phylum | [63] |
| Mung bean–proso millet | Loess-like | Proteobacteria, Chlorobacteria, Blastomonas, Acidobacteria, Helicobacter nitrifying, Firmicutes | Actinomycetes | Phylum fungi, Ascomycetes, Morpita, basidiomycetes | [64] |
| Wild soybean–sorghum | Salined fluvo-aquic soil | Proteobacteria, Bacteroidetes | Firmicutes, Gemmatimonadetes | [65] |
3.2. Soil Microbial Activity in the Legume–Cereal Intercropping System
Soil microbial and enzyme activities are more sensitive than soil physical and chemical properties under changes in the soil environment and can effectively indicate changes in ecosystem function, which is an important indicator of soil fertility [66]. Soil enzymes and microorganisms participate in the transformation of organic matter, the biological cycle of various elements, the formation of humus and organic–inorganic colloids, and have an important influence on the evolution of soil fertility [67]. Soil enzymes are proteins with microbial activity in soil, which is a key factor in soil metabolism and an important indicator of soil productivity and quality. Soil enzyme activity is closely related to soil physical and chemical properties and soil types.
There is a certain correlation between soil microbial activity and soil enzyme activity. Studies have found a significant positive correlation between bacteria and soil enzymes. Soil enzymes power the metabolism of organic matter in soil, and their activity directly reflects the activity of microorganisms, the ability of nutrient transformation and operation, and the strength of soil biochemical reactions. It is one of the important indexes used to evaluate soil organic matter content, quality, and health status [68]. Intercropping has strong interspecific root interactions, which can significantly improve soil microbial activity and promote the decomposition of humus and the transformation of organic matter and nutrients. The effects of intercropping on soil microbial activity were evaluated by total organic carbon, carbon management index, microbial biomass carbon, soil basal respiration, and metabolic quotient. The results showed that cowpea–forage intercropping significantly increased microbial activity compared with monoculture forage [69]. Amit Kumar et al. [70] conducted monoculture and mixed cultivation experiments of broad bean, lupine, and corn on medium fertility soil. Biomass and the kinetic parameters (maximum velocity (Vmax) and Miltonian constant (km)) of leucine aminopeptidase (LAP), β-1,4-n-acetylglucosaminopyrase (NAG), and phosphomonoesterase (PHO) in rhizosphere soil were determined. Maize and lupine are expected to have higher biomass under mixed cultivation than under monoculture. Second, LAP and NAG dynamic parameters were less responsive to monoculture and mixed culture. The Vmax and km of PHO in the faba bean rhizosphere were increased compared with those of maize and lupine. Under the condition of mixed culture, the absorption of phosphorus and nitrogen by Vicia faba seedlings was positively correlated with each other and showed net facilitative interactions. Dehydrogenase, nitrogen enzyme, rhizosphere, and diazotrophic bacteria were significantly affected by the different cropping rotations [59]. The intercropping system significantly increased the number of soil microbes due to the abundance of microbes and the increase in soil enzyme activity. The data results of higher rhizosphere-specific OTUs and higher diversity indexes indicated that the intercropping system had positive effects on soil microbial activity. Peanut intercropping with atractylodes significantly increased the activities of urease and sucrose [71]. Continuous monoculture was harmful to soil enzyme activity. Soil catalase, urease, and sucrase activities were significantly reduced under continuous monoculture [72]. Intercropping changes soil microbial activity through niche complementation. At the same time, the diversity of planting patterns affects the activities of soil microorganisms and enzymes.
3.3. Mechanism of Plant–Soil–Microorganism Interactions in the Legume–Cereal Intercropping System
Agricultural intensification has long-term negative effects, mainly due to the overuse of fertilizers and the simplification of farming systems (e.g., continuous monoculture). However, increasing attention has been given to enhancing root–soil–microbial interactions by selecting crop varieties/genotypes, optimizing crop spatiotemporal combinations, improving nutrient inputs, developing smart fertilizers, and applying soil or microbiome inoculations. Enhancing root–soil–microbial interactions can increase yields and reduce pest and disease stress in farming systems, and it can also reduce greenhouse gas emissions and enhance soil carbon sequestration [73]. Biodiversity can enhance ecosystem services. The mechanism is usually explained as a complementary effect and selection effect [74,75]. Complementary effects refer to the separation of spatial and temporal ecological niches or the promotion of interspecific effects to improve the efficiency of resource utilization and productivity. The selection effect means that the productivity of a community is dominated by more productive species in a community with higher diversity. The selection effect and complementary effect are important mechanisms for increasing yield in intercropping systems. Previous studies have also proven that there is a positive correlation between the diversity effect and efficient resource utilization in intercropping systems [27].
Intercropping between legumes and cereals has close physical roots, and root–soil–microbial interactions can lead to mixing of microbial communities between the two plant species during intercropping. In the rhizosphere, there is close exchange between plants and rhizosphere bacteria. In fact, plants can stimulate rhizosphere bacteria by secreting root exudates and phytochemical metabolites, thus causing the rhizosphere bacterial community to react with the plant [76]. Intercropping broad bean and wheat increased nodule biomass compared to monocropping wheat because of the interaction between the rhizosphere, and wheat promoted the secretion of root exudates containing flavonols, isoflavones, chalcone, and hesperidin [77]. Flavonoids produced by legume roots influence the behavior of related bacteria [78]. It also acts as a chemical attractant to concentrate compatible rhizobia on the root surface [79]. In addition, it stimulates the production of lipid-chitosaccharides in rhizosphere-related rhizobia. The latter induces a signal cascade that regulates the expression of symbiotic genes to transmit rhizobia-mediated signals and promote nodule development [80]. The resulting nodules can then positively affect N levels in the rhizosphere, a fraction of which is absorbed and utilized by the grain. Under soil nutrient deficiency and salt and drought stress, rhizosphere microorganisms dissolve P, immobilize N, and produce hydrogen cyanide, ACC-deaminase, indole-3-acetic acid, and siderophores through rhizosphere interactions to help plants resist stress [81,82]. Currently known salt-tolerant rhizosphere bacteria are Zhinguel-Liuella, Agrobacterium tumefaciens, Brachybacterium saurashtrense, Vibrio, Brevibacterium casei, Bacillus amyloliquefaciens, Haererohalobacter, etc. [83,84].
Complex plant–soil–microbial interactions maintain the balance between the different members of the microbial community. Nitrogen-fixing bacteria can fix nitrogen (N2) in the atmosphere and actively transport ammonium (NH4+) and nitrate (NO3−) into the host cell. Ammoniating bacteria convert organic N2 in soil to NH4+, which is further converted to NO3− by nitrifying bacteria. Legumes and nitrogen-fixing bacteria form nodule symbiosis. Mycorrhizal fungi convert arginine (Arg) to urea, which in turn converts to NH4+. Microbial communities convert minerals into plant usable nutrients such as phosphate (Pi), nitrogen (NH4+), and potassium (K+) by oxidizing, dissolving, or chelating, and release essential elements by producing organic acids and side spores. In addition, mycorrhizal fungi can transport elements directly into the host cytoplasm through specialized structures of mycelium and sporangium over long distances, thus increasing nutrient availability. Microorganisms stimulate plant growth by metabolizing tryptophan and other small molecules in plant secretions and producing plant hormones, including coenzymes, gibberellins, cytokinins, and plant hormone analogs [85,86].
3.4. Application and Development of Modern Biological Detection Technology in Soil Microorganisms
Modern molecular biology has confirmed that culturable microorganisms account for only 0.1~1% of the total number of microorganisms [87]. Since most functional soil microorganisms cannot be cultivated in laboratory conditions, 99% of the species and functions of soil microorganisms are still in the “black box” [88]. Uncultured microorganisms are of inestimable value to human production and play an important role in plant productivity. They are a group with great potential and are a new field for scientists to study because of their huge resources. In recent years, with the development of the independent cultivation of proteomics and bioinformatics, molecular techniques such as DNA stable isotope probing (DNA-SIP), RNA stable isotope probing (RNA-SIP), high-throughput sequencing of functional genes, metagenomics, and carbon and nitrogen functional gene chips have been widely used in the study of rhizosphere microbial community structure, which has allowed more microbes to be found and enhanced the understanding of the diversity of uncultivated microorganisms [89,90,91].
DNA stable isotope probing (SIP) technology can be used to track the flow of elements in different substances and organisms in the environment, thus helping scientists infer various chemical reactions or biological processes in nature [92,93,94,95,96]. The advantage of DNA-SIP technology is that it avoids the deficiency of traditional culture methods and facilitates the identification of target microorganisms by tracking labeled substrates [97,98,99]. However, there are few studies on the application of DNA-SIP technology in legume–cereal intercropping systems. To date, only Gou et al. [100] used DNA-SIP technology to screen and identify functional genes indicating nitrogen cycling at the enrichment position of soil 15N-DNA in the sugarcane/soybean intercropping system. Therefore, it will be a great breakthrough and innovation to apply DNA-SIP technology to the study of legume–cereal intercropping. However, the marker cost involved in DNA-SIP is high, and the operation procedure is cumbersome [101]. If the economy and conditions allow, carefully designed experiments and careful operation are required to avoid repeated work and obtain reliable results.
In recent years, high-throughput sequencing technology has been widely used to study soil microbial species diversity, structural diversity, functional diversity, and genetic diversity. This method overcomes some shortcomings of culture technology and obtains soil microbial information that cannot be isolated and cultured [102,103]. High-throughput sequencing of the nifH gene showed that the intercropping system enriched soil organic carbon and was conducive to the propagation of nitrogen-fixing bacteria at the tillering stage [104]. Legume–cereal intercropping could accumulate more nitrogen-fixing bacteria, which was more closely related to the soil; moreover, the diversity of the rhizosphere bacterial community of different sugarcane varieties intercropping with soybean was significantly different. Using the nifH gene sequencing method will contribute to the design of intercropping systems, selection of optimal varieties, enrichment of bacterial community structure, and creation of an environment conducive to crop growth, thus maximizing crop yield [90].
Metagenomics, the study of the entire genome of soil biota, is currently a powerful tool for assessing the diversity of complex microbial communities, providing access to many new species, genes, or molecules relevant to biotechnology and agricultural applications [105,106]. To improve crop productivity, soil health and sustainable intensification to better understand the interactions between cropping systems and soil microbial diversity, metagenomics can be used to analyze the diversity, abundance, and richness of rhizosphere soil microorganisms [107]. Through metagenomics research on traditional banana planting systems in the change of the microbial community, the results showed that planting systems were very important to support dynamic microbial diversity, and they were particularly useful to bacteria that help small farmers in sub-Saharan Africa (SSA) under the condition of scarce resources to promote collaborative interaction with soil and the total productivity of a plant [108]. The goal of metagenomics is not just to provide a collection of billions of biological projects but also to provide a unique opportunity to explore how microbial communities interact with soils and crops and ultimately harness the power of soil microbial communities to produce healthier, more robust crops or novel biomolecules.
4. Conclusions and Future Prospects
The legume–cereal intercropping system can reduce the application of chemical fertilizer, improve the uptake of nitrogen by cereal plants, promote the nitrogen fixation of legume root nodules, expand the soil nitrogen pool, and reduce the loss of available nitrogen in the soil through leaching, nitrification, and denitrification. Future research may focus on cropping systems rather than single crops. Therefore, the legume–cereal intercropping system has strong prospects for development in future agricultural systems. However, soil is a “black box”, and the specific nitrogen allocation and utilization in legume–cereal intercropping systems is still unclear. The effects of different legume and cereal intercropping systems on microbial community structure, composition, and activity need to be further studied. Using 15N2 isotope and molecular biology technology clearly showed the specific distribution of nitrogen in the legume–cereal intercropping system, and 13C-DNA-SIP, high-throughput sequencing, and metagenomics molecular biology technology were used to explore rhizosphere microbial community characteristics, which helped us understand the mechanism of the underground part. Thus, rhizosphere regulation gives full play to the biological potential of crops and improves nutrient resource use efficiency and crop productivity [109], which is an important way to realize environmentally friendly and sustainable agricultural development.
Author Contributions
Conceptualization, H.Y. and Y.L.; validation, H.L., Y.L., F.G., H.S., P.Z. and H.Y.; investigation, H.L., Y.L., H.Y., P.Z., F.G. and H.S.; resources, Y.L.; writing—original draft preparation, H.L.; writing—review and editing, H.L. and Y.L.; supervision, Y.L. and P.Z.; project administration, Y.L. and H.Y.; funding acquisition, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Number 41877051), and Ningbo Municipal Science and Technology Bureau (202002N3079, 2021Z047).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ladha, J.K.; Peoples, M.B.; Reddy, P.M.; Biswas, J.C.; Bennett, A.; Jat, M.L.; Krupnik, T.J. Biological nitrogen fixation and prospects for ecological intensification in cereal-based cropping systems. Field Crops Res. 2022, 283, 108541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.S.; Wang, J.Q.; Zhang, W.F.; Cui, Z.L.; Ma, W.Q.; Chen, X.P.; Jiang, R.F. Nutrient use efficiencies of major cereal crops in China and measures for improvement. Acta Pedol. Sin. 2008, 45, 915–924. [Google Scholar]
- Zhang, F.S.; Li, L. Using competitive and facilitative interactions in intercropping systems enhances crop productivity and nutrient-use efficiency. Plant Soil 2003, 248, 1–2. [Google Scholar] [CrossRef]
- Li, S.J.; Jensen, E.S.; Liu, N.; Zhang, Y.J.; Martensson, L.M.D. Species Interactions and Nitrogen Use during Early Intercropping of Intermediate Wheatgrass with a White Clover Service Crop. Agronomy 2021, 11, 388. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Gooding, M.; Ambus, P.; Corre-Hellou, G.; Crozat, Y.; Dahlmann, C.; Dibet, A.; von Fragstein, P.; Pristeri, A.; Monti, M.; et al. Pea-barley intercropping for efficient symbiotic N-2-fixation, soil N acquisition and use of other nutrients in European organic cropping systems. Field Crops Res. 2009, 113, 64–71. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; McElroy, M.S.; Chapagain, T.; Papadopoulos, Y.A.; Raizada, M.N. Belowground nitrogen transfer from legumes to non-legumes under managed herbaceous cropping systems. A review. Agron. Sustain. Dev. 2016, 36, 58. [Google Scholar] [CrossRef]
- Reilly, E.C.; Gutknecht, J.L.; Tautges, N.E.; Sheaffer, C.C.; Jungers, J.M. Nitrogen transfer and yield effects of legumes intercropped with the perennial grain crop intermediate wheatgrass. Field Crops Res. 2022, 286, 108627. [Google Scholar] [CrossRef]
- Ashworth, A.J.; West, C.P.; Allen, F.L.; Keyser, P.D.; Weiss, S.A.; Tyler, D.D.; Taylor, A.M.; Warwick, K.L.; Beamer, K.P. Biologically Fixed Nitrogen in Legume Intercropped Systems: Comparison of Nitrogen-Difference and Nitrogen-15 Enrichment Techniques. Agron. J. 2015, 107, 2419–2430. [Google Scholar] [CrossRef]
- Zhou, M.H.; Butterbach-Bahl, K. Assessment of nitrate leaching loss on a yield-scaled basis from maize and wheat cropping systems. Plant Soil 2014, 374, 977–991. [Google Scholar] [CrossRef]
- Mus, F.; Crook, M.B.; Garcia, K.; Costas, A.G.; Geddes, B.A.; Kouri, E.D.; Paramasivan, P.; Ryu, M.H.; Oldroyd, G.E.D.; Poole, P.S.; et al. Symbiotic Nitrogen Fixation and the Challenges to Its Extension to Nonlegumes. Appl. Environ. Microbiol. 2016, 82, 3698–3710. [Google Scholar] [CrossRef]
- Glaze-Corcoran, S.; Hashemi, M.; Sadeghpour, A.; Jahanzad, E.; Afshar, R.K.; Liu, X.B.; Herbert, S.J. Understanding intercropping to improve agricultural resiliency and environmental sustainability. Adv. Agron. 2020, 162, 199–256. [Google Scholar]
- Li, C.J.; Hoffland, E.; Kuyper, T.W.; Yu, Y.; Zhang, C.C.; Li, H.G.; Zhang, F.S.; van der Werf, W. Syndromes of production in intercropping impact yield gains. Nat. Plants 2020, 6, 653–660. [Google Scholar] [CrossRef]
- Li, X.F.; Wang, Z.G.; Bao, X.G.; Sun, J.H.; Yang, S.C.; Wang, P.; Wang, C.B.; Wu, J.P.; Liu, X.R.; Tian, X.L.; et al. Long-term increased grain yield and soil fertility from intercropping. Nat. Sustain. 2021, 4, 943–950. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Murray, J.D.; Poole, P.S.; Downie, J.A. The Rules of Engagement in the Legume-Rhizobial Symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef]
- Udvardi, M.; Poole, P.S. Transport and Metabolism in Legume-Rhizobia Symbioses. Annu. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar] [CrossRef] [PubMed]
- Schipanski, M.E.; Drinkwater, L.E.; Russelle, M.P. Understanding the variability in soybean nitrogen fixation across agroecosystems. Plant Soil 2010, 329, 379–397. [Google Scholar] [CrossRef]
- Li, L.; Yang, S.C.; Li, X.L.; Zhang, F.S.; Christie, P. Interspecific complementary and competitive interactions between intercropped maize and faba bean. Plant Soil 1999, 212, 105–114. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.; Zhang, H.; Shao, Z.; Sun, B.; Gao, Q. Enhancement of rhizosphere citric acid and decrease of NO3−/NH4+ ratio by root interactions facilitate N fixation and transfer. Plant Soil 2019, 447, 169–182. [Google Scholar] [CrossRef]
- Fan, F.L.; Zhang, F.S.; Song, Y.N.; Sun, J.H.; Bao, X.G.; Guo, T.W.; Li, L. Nitrogen fixation of faba bean (Vicia faba L.) interacting with a non-legume in two contrasting intercropping systems. Plant Soil 2006, 283, 275–286. [Google Scholar] [CrossRef]
- Nyfeler, D.; Huguenin-Elie, O.; Suter, M.; Frossard, E.; Lüscher, A. Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources. Agric. Ecosyst. Environ. 2011, 140, 155–163. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Drinkwater, L.E. Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil 2012, 357, 147–159. [Google Scholar] [CrossRef]
- Tian, J.H.; Tang, M.T.; Xu, X.; Luo, S.S.; Condron, L.M.; Lambers, H.; Cai, K.Z.; Wang, J.W. Soybean (Glycine max (L.) Merrill) intercropping with reduced nitrogen input influences rhizosphere phosphorus dynamics and phosphorus acquisition of sugarcane (Saccharum officinarum). Biol. Fertil. Soils 2020, 56, 1063–1075. [Google Scholar] [CrossRef]
- Fan, Z.; Zhao, Y.; Chai, Q.; Zhao, C.; Yu, A.; Coulter, J.A.; Gan, Y.; Cao, W. Synchrony of nitrogen supply and crop demand are driven via high maize density in maize/pea strip intercropping. Sci. Rep. 2019, 9, 10954. [Google Scholar] [CrossRef] [PubMed]
- Hei, Z.W.; Xiang, H.M.; Zhang, J.E.; Liang, K.M.; Zhong, J.W.; Li, M.J.; Lu, Y.Q. Rice intercropping with water mimosa (Neptunia oleracea Lour.) can facilitate soil N utilization and alleviate apparent N loss. Agric. Ecosyst. Environ. 2021, 313, 107378. [Google Scholar] [CrossRef]
- Wang, X.Y.; Gao, Y.Z. Advances in the mechanism of cereal/legume intercropping promotion of symbiotic nitrogen fixation. Chin. Sci. Bull. 2019, 65, 142–149. [Google Scholar] [CrossRef]
- Francisquini, A.; Calonego, J.C.; Rosolem, C.A.; dos Santos, C.H.; Tiritan, C.S. Increase of nitrogen-use efficiency by phosphorus fertilization in grass-legume pastures. Nutr. Cycl. Agroecosystems 2020, 118, 165–175. [Google Scholar] [CrossRef]
- Li, L.; Li, S.M.; Sun, J.H.; Zhou, L.L.; Bao, X.G.; Zhang, H.G.; Zhang, F.S. Diversity enhances agricultural productivity via rhizosphere phosphorus facilitation on phosphorus-deficient soils. Proc. Natl. Acad. Sci. USA 2007, 104, 11192–11196. [Google Scholar] [CrossRef] [PubMed]
- Salgado, G.C.; Ambrosano, E.J.; Rossi, F.; Otsuk, I.P.; Ambrosano, G.M.B.; Santana, C.A.; Muraoka, T.; Trivelin, P.C.O. Biological N Fixation and N Transfer in an Intercropping System between Legumes and Organic Cherry Tomatoes in Succession to Green Corn. Agriculture 2021, 11, 690. [Google Scholar] [CrossRef]
- Boddey, R.M.; Sa, J.C.D.; Alves, B.J.R.; Urquiaga, S. The contribution of biological nitrogen fixation for sustainable agricultural systems in the tropics. Soil Biol. Biochem. 1997, 29, 787–799. [Google Scholar] [CrossRef]
- Tripathi, S.C.; Venkatesh, K.; Meena, R.P.; Chander, S.; Singh, G.P. Sustainable intensification of maize and wheat cropping system through pulse intercropping. Sci. Rep. 2021, 11, 18805. [Google Scholar] [CrossRef]
- Willey, R. Intercropping Its Importance And Research Needs Part 1. Competition And Yield Advantages. Field Crop Abstr. 1979, 32, 1–10. [Google Scholar]
- Wahbi, S.; Maghraoui, T.; Hafidi, M.; Sanguin, H.; Oufdou, K.; Prin, Y.; Duponnois, R.; Galiana, A. Enhanced transfer of biologically fixed N from faba bean to intercropped wheat through mycorrhizal symbiosis. Appl. Soil Ecol. 2016, 107, 91–98. [Google Scholar] [CrossRef]
- Xiao, Y.B.; Li, L.; Zhang, F.S. Effect of root contact on interspecific competition and N transfer between wheat and fababean using direct and indirect N-15 techniques. Plant Soil 2004, 262, 45–54. [Google Scholar] [CrossRef]
- Yong, T.W.; Liu, X.M.; Yang, F.; Song, C.; Wang, X.C.; Liu, W.G.; Su, B.Y.; Zhou, L.; Yang, W.Y. Characteristics of Nitrogen Uptake, Use and Transfer in a Wheat-Maize-Soybean Relay Intercropping System. Plant Prod. Sci. 2015, 18, 388–397. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Freney, J.R.; Simpson, J.R. Assessing nitrogen transfer from legumes to associated grasses. Soil Biol. Biochem. 1985, 17, 575–577. [Google Scholar] [CrossRef]
- Shao, Z.; Wang, X.; Gao, Q.; Zhang, H.; Gao, Y. Root Contact between Maize and Alfalfa Facilitates Nitrogen Transfer and Uptake Using Techniques of Foliar 15N-Labeling. Agronomy 2020, 10, 360. [Google Scholar] [CrossRef]
- Yu, L.; Tang, Y.; Wang, Z.; Gou, Y.; Wang, J. Nitrogen-cycling genes and rhizosphere microbial community with reduced nitrogen application in maize/soybean strip intercropping. Nutr. Cycl. Agroecosyst. 2018, 113, 35–49. [Google Scholar] [CrossRef]
- Turner, T.R.; Ramakrishnan, K.; Walshaw, J.; Heavens, D.; Alston, M.; Swarbreck, D.; Osbourn, A.; Grant, A.; Poole, P.S. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 2013, 7, 2248–2258. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gomez-Cadenas, A.; Perez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Hu, L.F.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.F.; Shen, Q.R.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Zhang, R.Z.; Mu, Y.; Li, X.R.; Li, S.M.; Sang, P.; Wang, X.R.; Wu, H.L.; Xu, N. Response of the arbuscular mycorrhizal fungi diversity and community in maize and soybean rhizosphere soil and roots to intercropping systems with different nitrogen application rates. Sci. Total Environ. 2020, 740, 139810. [Google Scholar] [CrossRef] [PubMed]
- Bainard, L.D.; Koch, A.M.; Gordon, A.M.; Klironomos, J.N. Growth response of crops to soil microbial communities from conventional monocropping and tree-based intercropping systems. Plant Soil 2013, 363, 345–356. [Google Scholar] [CrossRef]
- Pivato, B.; Semblat, A.; Guegan, T.; Jacquiod, S.; Martin, J.; Deau, F.; Moutier, N.; Lecomte, C.; Burstin, J.; Lemanceau, P. Rhizosphere Bacterial Networks, but Not Diversity, Are Impacted by Pea-Wheat Intercropping. Front. Microbiol. 2021, 12, 674556. [Google Scholar] [CrossRef] [PubMed]
- Morrien, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; de Hollander, M.; Soto, R.L.; Bouffaud, M.L.; Buee, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef]
- Li, S.; Wu, F.Z. Diversity and Co-occurrence Patterns of Soil Bacterial and Fungal Communities in Seven Intercropping Systems. Front. Microbiol. 2018, 9, 1521. [Google Scholar] [CrossRef]
- Hai, B.; Diallo, N.H.; Sall, S.; Haesler, F.; Schauss, K.; Bonzi, M.; Assigbetse, K.; Chotte, J.L.; Munch, J.C.; Schloter, M. Quantification of Key Genes Steering the Microbial Nitrogen Cycle in the Rhizosphere of Sorghum Cultivars in Tropical Agroecosystems. Appl. Environ. Microbiol. 2009, 75, 4993–5000. [Google Scholar] [CrossRef]
- Rodriguez, C.; Carlsson, G.; Englund, J.E.; Flohr, A.; Pelzer, E.; Jeuffroy, M.H.; Makowski, D.; Jensen, E.S. Grain legume-cereal intercropping enhances the use of soil-derived and biologically fixed nitrogen in temperate agroecosystems. A meta-analysis. Eur. J. Agron. 2020, 118, 126077. [Google Scholar] [CrossRef]
- Zeng, H.L.; Yu, L.L.; Liu, P.; Wang, Z.G.; Chen, Y.; Wang, J.W. Nitrogen fertilization has a stronger influence than cropping pattern on AMF community in maize/soybean strip intercropping systems. Appl. Soil Ecol. 2021, 167, 104034. [Google Scholar] [CrossRef]
- Zhao, R.T.; Li, X.; Bei, S.K.; Li, D.D.; Li, H.G.; Christie, P.; Bender, S.F.; Zhang, J.L. Enrichment of nosZ-type denitrifiers by arbuscular mycorrhizal fungi mitigates N2O emissions from soybean stubbles. Environ. Microbiol. 2021, 23, 6587–6602. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.Q.; Fallah, N.; Weng, P.Y.; Zhou, Y.M.; Tang, X.M.; Tayyab, M.; Liu, Y.M.; Liu, Q.; Xiao, Y.J.; Hu, C.H.; et al. Sugarcane-Peanut Intercropping System Enhances Bacteria Abundance, Diversity, and Sugarcane Parameters in Rhizospheric and Bulk Soils. Front. Microbiol. 2022, 12, 815129. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.Y.; Huang, L.K.; Liu, Q.Z.; Xu, S.N.; Wen, Z.Y.; Qin, S.; Li, T.Q.; Feng, Y. Positive effects of applying endophytic bacteria in eggplant-Sedum intercropping system on Cd phytoremediation and vegetable production in cadmium polluted greenhouse. J. Environ. Sci. 2022, 115, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Mu, Y.H.; Cheng, Y.B.; Liu, X.G.; Nian, H. Effects of intercropping sugarcane and soybean on growth, rhizosphere soil microbes, nitrogen and phosphorus availability. Acta Physiol. Plant. 2013, 35, 1113–1119. [Google Scholar] [CrossRef]
- Zhang, R.Z. Study on Productivity, Nutrient Uptake and Mechanism of Soil Microbial Activity in Maize/Soybean Intercropping by Nitrogen Fertilizer. Ph.D. Thesis, Northeast Agricultural University, Harbin, China, 2020. [Google Scholar]
- Chung, H.G.; Zak, D.R.; Reich, P.B.; Ellsworth, D.S. Plant species richness, elevated CO2, and atmospheric nitrogen deposition alter soil microbial community composition and function. Glob. Change Biol. 2007, 13, 980–989. [Google Scholar] [CrossRef]
- Duchene, O.; Vian, J.-F.; Celette, F. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric. Ecosyst. Environ. 2017, 240, 148–161. [Google Scholar] [CrossRef]
- Lin, W.W.; Li, N.; Chen, L.S.; Wu, Z.Y.; Lin, W.X.; Shen, L.H. Effects of maize and sobean interspecific interactions on rhizospheric bacteria community structure and diversity. Chin. J. Eco Agric. 2022, 30, 26–37. [Google Scholar]
- Solanki, M.K.; Wang, F.Y.; Wang, Z.; Li, C.N.; Lan, T.J.; Singh, R.K.; Singh, P.; Yang, L.T.; Li, Y.R. Rhizospheric and endospheric diazotrophs mediated soil fertility intensification in sugarcane-legume intercropping systems. J. Soils Sediments 2019, 19, 1911–1927. [Google Scholar] [CrossRef]
- Tang, X.M.; Jiang, J.; Huang, Z.P.; Wu, H.N.; Wang, J.; He, L.Q.; Xiong, F.Q.; Zhong, R.C.; Liu, J.; Han, Z.Q.; et al. Sugarcane/peanut intercropping system improves the soil quality and increases the abundance of beneficial microbes. J. Basic Microbiol. 2021, 61, 165–176. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Liu, Q.; Zhao, X.; Chen, B. Effect of Two Different Sugarcane Cultivars on Rhizosphere Bacterial Communities of Sugarcane and Soybean Upon Intercropping. Front. Microbiol. 2021, 11, 3404. [Google Scholar] [CrossRef]
- Chen, P.; He, W.; Shen, Y.; Zhu, L.Y.; Yao, X.Z.; Sun, R.B.; Dai, C.C.; Sun, B.; Chen, Y. Interspecific Neighbor Stimulates Peanut Growth Through Modulating Root Endophytic Microbial Community Construction. Front. Plant Sci. 2022, 13, 830666. [Google Scholar] [CrossRef]
- Liao, H.; Li, Y.Y.; Yao, H.Y. Biochar Amendment Stimulates Utilization of Plant-Derived Carbon by Soil Bacteria in an Intercropping System. Front. Microbiol. 2019, 10, 1361. [Google Scholar] [CrossRef] [PubMed]
- Xga, B.; Cl, C.; Jing, L.; Yan, L.; Qya, B.; Wza, B.; Pu, Y.; Bfa, B. Responses of rhizosphere soil properties, enzyme activities and microbial diversity to intercropping patterns on the Loess Plateau of China—ScienceDirect. Soil Tillage Res. 2018, 195, 104355. [Google Scholar]
- Zhu, Y.H.; Song, X.L.; Wang, X.F.; Chen, W.F.; Niu, X.C. The yield increase and land improvement effects of different sorghum/wild soybean intercropping patterns on reclaimed coastal salt pans. J. Soils Sediments 2022, 22, 731–744. [Google Scholar] [CrossRef]
- Ma, X.J.; Li, Y.F. Soil Microbial Biomass and Enzyme Activities during Revegetation Process in the Southeastern Fringe of the Tengger Desert. J. Desert Res. 2019, 39, 159–166. [Google Scholar]
- Maynur, Y.K.M.; Zhang, B.C.; Mamtimin, S.L.Y.M. Seaonal Variations of Microbial Biomass and Soil Enzyme Activity in Biological Soil Crusts in the Gurbantunggut Desert. J. Desert Res. 2013, 33, 1091–1097. [Google Scholar]
- Wu, X.S.; Zhou, X.L.; Cao, F.M.; Zhu, B.C.; Zhao, T.K.; Shen, D.L. Effects of different fertilization on the corn yield and soil enzyme activity in corn growth period. Soil Fert. Sci. China 2015, 1, 44–49. [Google Scholar]
- Silva, L.S.; Laroca, J.V.D.; Coelho, A.P.; Gonsalves, E.C.; Gomes, R.P.; Pacheco, L.P.; Carvalho, P.C.D.; Pires, G.C.; Oliveira, R.L.; de Souza, J.M.A.; et al. Does grass-legume intercropping change soil quality and grain yield in integrated crop-livestock systems? Appl. Soil Ecol. 2022, 170, 104257. [Google Scholar] [CrossRef]
- Kumar, A.; Blagodaskaya, E.; Dippold, M.A.; Temperton, V.M. Positive intercropping effects on biomass production are species-specific and involve rhizosphere enzyme activities: Evidence from a field study. Soil Ecol. Lett. 2021, 1–10. [Google Scholar] [CrossRef]
- Dai, C.C.; Chen, Y.; Wang, X.X.; Li, P.D. Effects of intercropping of peanut with the medicinal plant Atractylodes lancea on soil microecology and peanut yield in subtropical China. Agrofor. Syst. 2013, 87, 417–426. [Google Scholar] [CrossRef]
- Yao, H.Y.; Jiao, X.D.; Wu, F.Z. Effects of continuous cucumber cropping and alternative rotations under protected cultivation on soil microbial community diversity. Plant Soil 2006, 284, 195–203. [Google Scholar] [CrossRef]
- Jing, J.; Cong, W.-F.; Bezemer, T.M. Legacies at work: Plant-soil-microbiome interactions underpinning agricultural sustainability. Trends Plant Sci. 2022, 27, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Oram, N.J.; Ravenek, J.M.; Barry, K.E.; Weigelt, A.; Chen, H.M.; Gessler, A.; Gockele, A.; de Kroon, H.; van der Paauw, J.W.; Scherer-Lorenzen, M.; et al. Below-ground complementarity effects in a grassland biodiversity experiment are related to deep-rooting species. J. Ecol. 2018, 106, 265–277. [Google Scholar] [CrossRef]
- Yang, H.; Xu, H.S.; Zhang, W.P.; Li, Z.X.; Fan, H.X.; Lambers, H.; Li, L. Overyielding is accounted for partly by plasticity and dissimilarity of crop root traits in maize/legume intercropping systems. Funct. Ecol. 2022. [Google Scholar] [CrossRef]
- Vora, S.M.; Joshi, P.; Belwalkar, M.; Archana, G. Root exudates influence chemotaxis and colonization of diverse plant growth promoting rhizobacteria in the pigeon pea-maize intercropping system. Rhizosphere 2021, 18, 100331. [Google Scholar] [CrossRef]
- Liu, Y.C.; Qin, X.M.; Xiao, J.X.; Tang, L.; Wei, C.Z.; Wei, J.J.; Zheng, Y. Intercropping influences component and content change of flavonoids in root exudates and nodulation of Faba bean. J. Plant Interact. 2017, 12, 187–192. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.Y.; Wu, H.M.; Zhang, F.F.; Li, C.J.; Li, X.X.; Lambers, H.; Li, L. Root exudates drive interspecific facilitation by enhancing nodulation and N-2 fixation. Proc. Natl. Acad. Sci. USA 2016, 113, 6496–6501. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Lateif, K.; Bogusz, D.; Hocher, V. The role of flavonoids in the establishment of plant roots endosymbioses with arbuscular mycorrhiza fungi, rhizobia and Frankia bacteria. Plant Signal. Behav. 2012, 7, 636–641. [Google Scholar] [CrossRef]
- Concha, C.; Doerner, P. The impact of the rhizobia-legume symbiosis on host root system architecture. J. Exp. Bot. 2020, 71, 3902–3921. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Ramteke, P.W. Plant growth-promoting rhizobacteria: Strategies to improve abiotic stresses under sustainable agriculture. J. Plant Nutr. 2019, 42, 1402–1415. [Google Scholar] [CrossRef]
- Chamkhi, I.; Cheto, S.; Geistlinger, J.; Zeroual, Y.; Kouisni, L.; Bargaz, A.; Ghoulam, C. Legume-based intercropping systems promote beneficial rhizobacterial community and crop yield under stressing conditions. Ind. Crops Prod. 2022, 183, 114958. [Google Scholar] [CrossRef]
- Shukla, P.S.; Agarwal, P.K.; Jha, B. Improved Salinity Tolerance of Arachis hypogaea (L.) by the Interaction of Halotolerant Plant-Growth-Promoting Rhizobacteria. J. Plant Growth Regul. 2012, 31, 195–206. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S.; Seem, K.; Mishra, A.; Sopory, S.K. Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Bioch. 2013, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.M.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Zhang, R.F.; Vivanco, J.M.; Shen, Q.R. The unseen rhizosphere root-soil-microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Volkov, I.; Banavar, J.R.; Maritan, A. Comment on “Computational improvements reveal great bacterial diversity and high metal toxicity in soil”. Science 2006, 313, 918. [Google Scholar] [CrossRef]
- Whiteley, A.S.; Manefield, M.; Lueders, T. Unlocking the ’microbial black box’ using RNA-based stable isotope probing technologies. Curr. Opin. Biotechnol. 2006, 17, 67–71. [Google Scholar] [CrossRef]
- Li, Z.; Yao, Q.M.; Guo, X.; Crits-Christoph, A.; Mayes, M.A.; Hervey, W.J.; Lebeis, S.L.; Banfield, J.F.; Hurst, G.B.; Hettich, R.L.; et al. Genome-Resolved Proteomic Stable Isotope Probing of Soil Microbial Communities Using (CO2)-C-13 and C-13-Methanol. Front. Microbiol. 2019, 10, 2706. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, W.Q.; He, H.L.; Wang, Z.T.; Cao, Y.H. Effects of Sugarcane and Soybean Intercropping on the Nitrogen-Fixing Bacterial Community in the Rhizosphere. Front. Microbiol. 2021, 12, 2846. [Google Scholar] [CrossRef]
- Sertse, D.; You, F.M.; Ravichandran, S.; Cloutier, S. The Complex Genetic Architecture of Early Root and Shoot Traits in Flax Revealed by Genome-Wide Association Analyses. Front. Plant Sci. 2019, 10, 1483. [Google Scholar] [CrossRef]
- Chiewattanakul, M.; McAleer, A.D.A.; Reay, M.K.; Griffiths, R.I.; Buss, H.L.; Evershed, R.P. Compound-specific amino acid 15N-stable isotope probing for the quantification of biological nitrogen fixation in soils. Soil Biol. Biochem. 2022, 169, 108654. [Google Scholar] [CrossRef]
- Ouyang, W.Y.; Su, J.Q.; Richnow, H.H.; Adrian, L. Identification of dominant sulfamethoxazole-degraders in pig farm-impacted soil by DNA and protein stable isotope probing. Environ. Int. 2019, 126, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Koechli, C.; Campbell, A.N.; Pepe-Ranney, C.; Buckley, D.H. Assessing fungal contributions to cellulose degradation in soil by using high-throughput stable isotope probing. Soil Biol. Biochem. 2019, 130, 150–158. [Google Scholar] [CrossRef]
- Meier-Augenstein, W. From stable isotope ecology to forensic isotope ecology—Isotopes’ tales. Forensic Sci. Int. 2019, 300, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.W.; Zhao, J.; Zheng, Y.; Zhang, H.M.; Zhang, J.B.; Chen, R.R.; Lin, X.G.; Jia, Z.J. Active Soil Nitrifying Communities Revealed by In Situ Transcriptomics and Microcosm-Based Stable-Isotope Probing. Appl. Environ. Microbiol. 2020, 86, e01807-20. [Google Scholar] [CrossRef]
- Luo, D.; Meng, X.T.; Zheng, N.G.; Li, Y.Y.; Yao, H.Y.; Chapman, S.J. The anaerobic oxidation of methane in paddy soil by ferric iron and nitrate, and the microbial communities involved. Sci. Total Environ. 2021, 788, 147773. [Google Scholar] [CrossRef]
- Wang, J.; Chapman, S.J.; Ye, Q.F.; Yao, H.Y. Limited effect of planting transgenic rice on the soil microbiome studied by continuous (CO2)-C-13 labeling combined with high-throughput sequencing. Appl. Microbiol. Biotechnol. 2019, 103, 4217–4227. [Google Scholar] [CrossRef]
- Wang, F.; Shi, N.; Jiang, R.F.; Zhang, F.S.; Feng, G. In situ stable isotope probing of phosphate-solubilizing bacteria in the hyphosphere. J. Exp. Bot. 2016, 67, 1689–1701. [Google Scholar] [CrossRef]
- Gou, Y.G.; Yu, L.L.; Xu, X.; Wang, J.W. Identification of 15N-DNA enrichment sites in DNA-SIP to reveal functional genes by qPCR from sugarcane-soybean intercropping soil. J. Agro Environ. Sci. 2019, 38, 140–147. [Google Scholar]
- Wang, J.; Zhang, X.; Yao, H.Y. Optimizing ultracentrifugation conditions for DNA-based stable isotope probing (DNA-SIP). J. Microbiol. Methods 2020, 173, 105938. [Google Scholar] [CrossRef]
- Ding, T.T.; Yan, Z.C.; Zhang, W.Z.; Duan, T.Y. Green Manure Crops Affected Soil Chemical Properties and Fungal Diversity and Community of Apple Orchard in the Loess Plateau of China. J. Soil Sci. Plant Nutr. 2021, 21, 1089–1102. [Google Scholar] [CrossRef]
- Li, H.; Luo, L.Y.; Tang, B.; Guo, H.L.; Cao, Z.Y.; Zeng, Q.; Chen, S.L.; Chen, Z.H. Dynamic changes of rhizosphere soil bacterial community and nutrients in cadmium polluted soils with soybean-corn intercropping. BMC Microbiol. 2022, 22, 57. [Google Scholar] [CrossRef] [PubMed]
- Solanki, M.K.; Wang, Z.; Wang, F.Y.; Li, C.N.; Gupta, C.L.; Singh, R.K.; Malviya, M.K.; Singh, P.; Yang, L.T.; Li, Y.R. Assessment of DiazotrophicProteobacteriain Sugarcane Rhizosphere When Intercropped With Legumes (Peanut and Soybean) in the Field. Front. Microbiol. 2020, 11, 1814. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Naorem, A.K.; Lal, R.; Dalal, R.C.; Sinha, N.K.; Patra, A.K.; Chaudhari, S.K. Disease-Suppressive Soils-Beyond Food Production: A Critical Review. J. Soil Sci. Plant Nutr. 2021, 21, 1437–1465. [Google Scholar] [CrossRef] [PubMed]
- Alawiye, T.; Babalola, O. Metagenomic Insight into the Community Structure and Functional Genes in the Sunflower Rhizosphere Microbiome. Agriculture 2021, 11, 167. [Google Scholar] [CrossRef]
- Iquebal, M.A.; Jagannadham, J.; Jaiswal, S.; Prabha, R.; Rai, A.; Kumar, D. Potential Use of Microbial Community Genomes in Various Dimensions of Agriculture Productivity and Its Management: A Review. Front. Microbiol. 2022, 13, 708335. [Google Scholar] [CrossRef]
- Kaushal, M.; Tumuhairwe, J.B.; Kaingo, J.; Richard, M.; Nakamanya, F.; Taulya, G.; Coyne, D. Compositional Shifts in Microbial Diversity under Traditional Banana Cropping Systems of Sub-Saharan Africa. Biology 2022, 11, 756. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Zeng, Y.; Tang, L.; Xiao, J.X.; Zeng, J.; Zhang, K.X. Rhizosphere Biological Processes of Legume//Cereal Intercropping Systems: A Review. J. Agric. Resour. Environ. 2016, 33, 407–415. [Google Scholar]
Figure 1.
Nitrogen transfer and distribution in the legume–cereal intercropping system.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lai, H.; Gao, F.; Su, H.; Zheng, P.; Li, Y.; Yao, H. Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review. Agronomy 2022, 12, 1900. https://doi.org/10.3390/agronomy12081900
AMA Style
Lai H, Gao F, Su H, Zheng P, Li Y, Yao H. Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review. Agronomy. 2022; 12(8):1900. https://doi.org/10.3390/agronomy12081900
Chicago/Turabian StyleLai, Huiling, Fuyun Gao, Hao Su, Peng Zheng, Yaying Li, and Huaiying Yao. 2022. "Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review" Agronomy 12, no. 8: 1900. https://doi.org/10.3390/agronomy12081900
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.