
Heavy Metals Can Affect Plant Morphology and Limit Plant Growth and Photosynthesis Processes
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Biomass Measurements and Root Growth Analyses of P. divaricata under Cd Stress
2.3. Determination of Photosynthetic Parameters
2.4. Determination of Chlorophyll Fluorescence Parameters
- (1)
- After dark adaptation of the selected leaf for 30 min, a light with a wavelength of 630 nm was flashed on the leaf to measure the initial fluorescence (Fo). Then, light with a wavelength of 630 nm and an intensity of approximately 6000 μmol·m−2·s−1 was used to saturate the reaction centers of photosystem II (Duration 0.8 s, Intensity 7, Blue 10%, Modulation 20 KHz, Filter 50 KHz) to determine the maximum fluorescence (Fm).
- (2)
- The leaves sampled for the measurements were exposed to continuous photochemical active light (constant light in a greenhouse with a light intensity of 80 μmol·m−2·s−1 for 30 min at steady-state fluorescence (Fs)). Then, the light intensity was increased to 6000 μmol·m−2·s−1 to measure the maximum fluorescence (Fm′) after adaptation under saturated light irradiation. Then, the photochemically active light was stopped, and far-red light with a wavelength of 740 nm was applied (Duration 6 s, Intensity 8, Modulation 0.25 KHz, Filter 1 KHz) to measure the minimal fluorescence, Fo′.
- (3)
- Other chlorophyll fluorescence parameters of PSII, such as the maximum photochemical efficiency (Fv/Fm), actual photochemical efficiency of PSII (ΦPSII), the photochemical quenching (qP), non-photochemical quenching (NPQ), and electron transfer rate (ETR) of PSII were calculated according to the following formulas:
2.5. Stomatal Pore Observation by Scanning Electron Microscopy
2.6. Fluorescence CO2 Response Curve
2.7. Statistical Analysis
3. Results
3.1. Effects of Cd Stress on the Biomass, Root Growth, and Metals Content of P. divaricata
3.2. Effects of Cadmium on Leaf Pigment Content of P. divaricata
3.3. Distribution Pattern of Blade Cross-Section
3.4. Effects of Cadmium on the Photochemical Reactions of P. divaricata
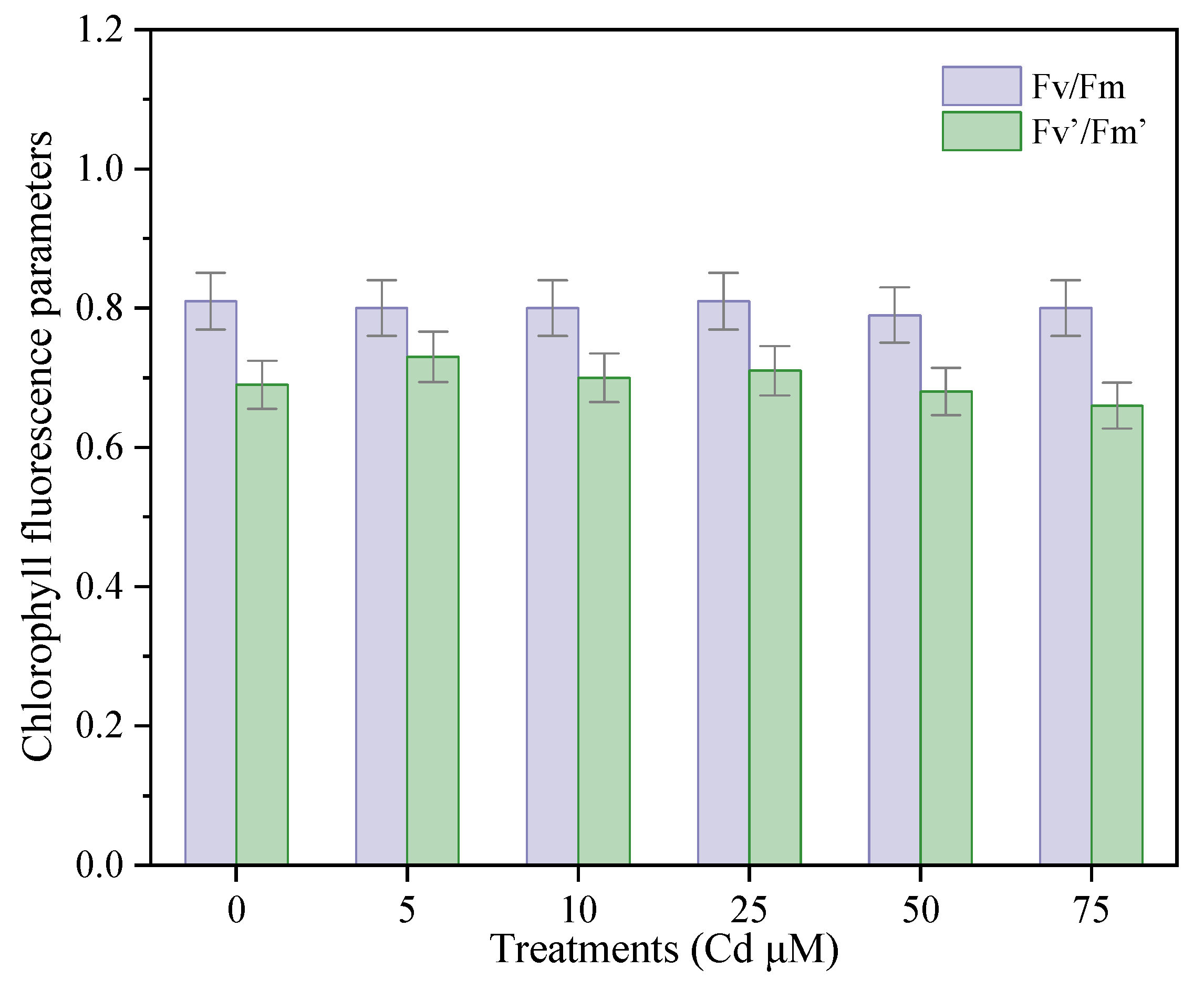
3.4.1. Effects of Cadmium on the Chlorophyll Fluorescence Parameters of P. divaricata
3.4.2. Effects of Cadmium on the Excited-State Energy Distribution and the Photoreaction Quantum Yield in P. divaricata
3.5. Effects of Cadmium on the Photosynthetic Parameters of P. divaricata
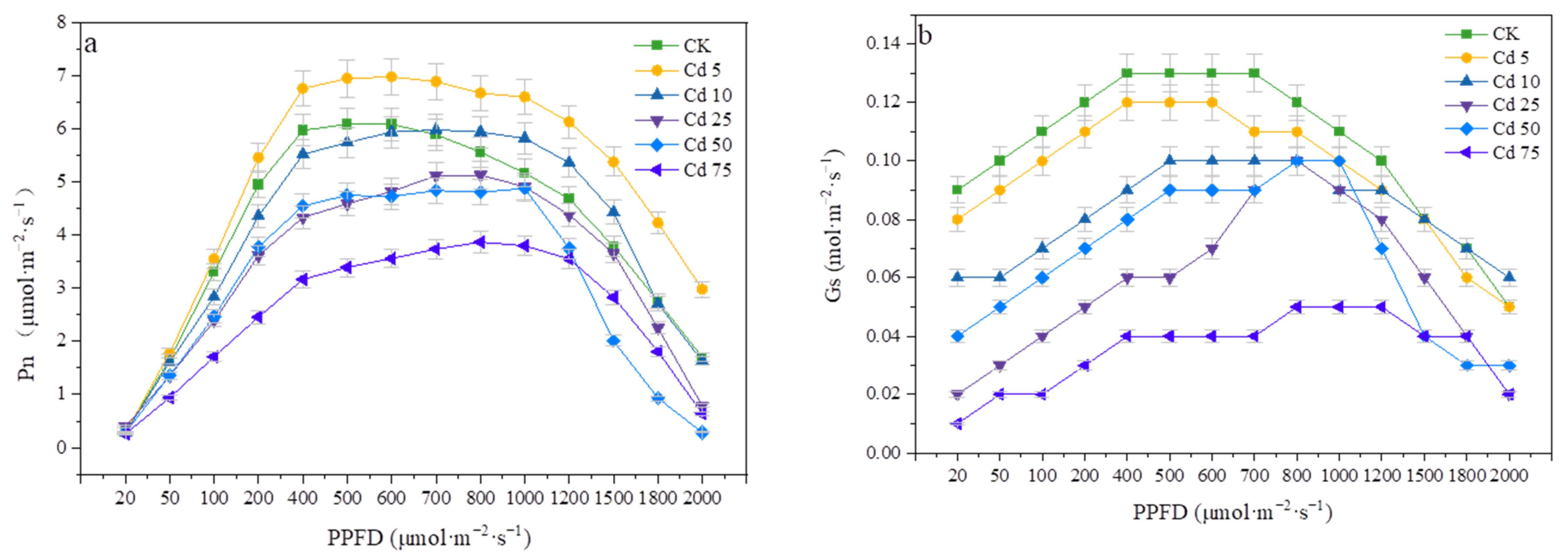
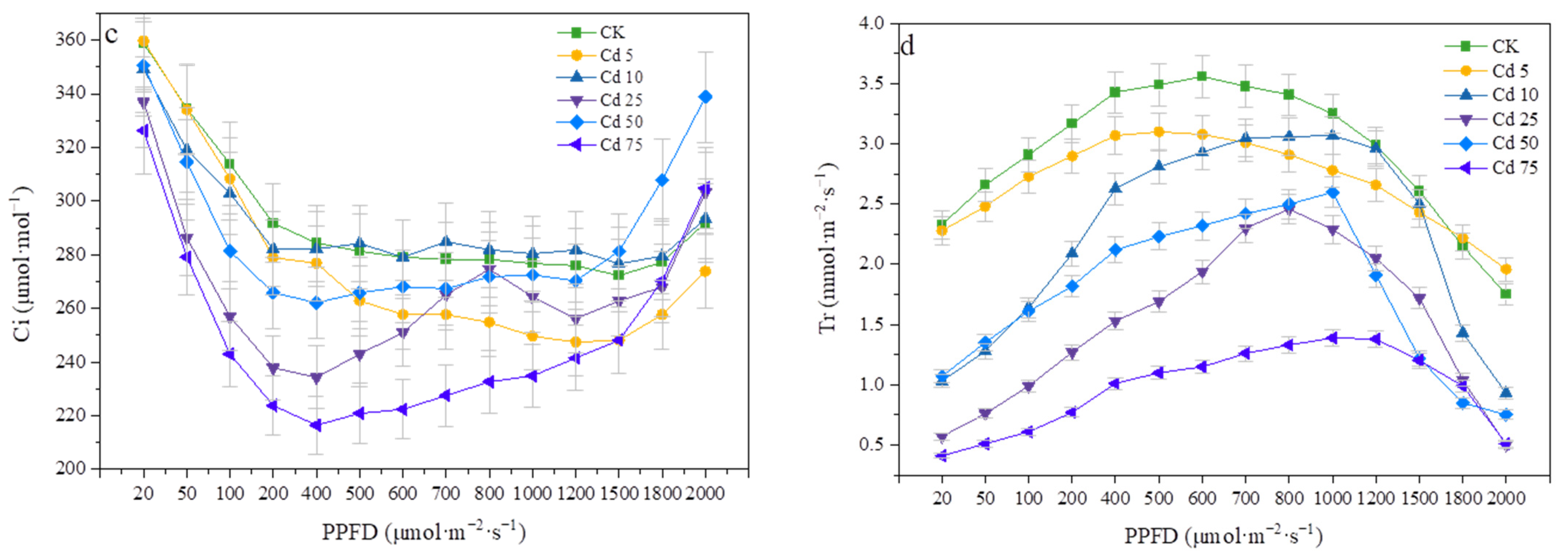
3.5.1. Effects of Cadmium on Photosynthetic Parameters and the Light Response Curve of P. divaricata
3.5.2. Analysis of the Factors Affected by Cadmium That Influence the Photosynthesis of P. divaricata
4. Discussion
4.1. Cadmium Resulted in Growth Promotion in Low Concentrations and Growth Inhibition in High Concentrations in P. divaricata
4.2. Effects of Cadmium on P. divaricata Photosynthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pouresmaieli, M.; Ataei, M.; Forouzandeh, P.; Azizollahi, P.; Mahmoudifard, M. Recent progress on sustainable phytoremediation of heavy metals from soil. J. Environ. Chem. Eng. 2022, 10, 108482. [Google Scholar] [CrossRef]
- Wang, L.; Yang, D.; Ma, F. How a functional soil animal-earthworm affect arbuscular mycorrhizae-assisted phytoremediation in metals contaminated soil? J. Hazard. Mater. 2022, 435, 128991. [Google Scholar] [CrossRef] [PubMed]
- Tavarez, M.; Macri, A.; Sankaran, R.P. Cadmium and zinc partitioning and accumulation during grain filling in two near isogenic lines of durum wheat. Plant Physiol. Biochem. 2015, 97, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Sigfridsson, K.G.V.; Bernat, G.; Mamedov, F.; Styring, S. Molecular interference of Cd2+ with photosystem ii. Biochim. Biophys. Acta-Bioenerg. 2004, 1659, 19–31. [Google Scholar] [CrossRef]
- Bora, M.S.; Sarma, K.P. Anatomical and ultrastructural alterations in Ceratopteris pteridoides under cadmium stress: A mechanism of cadmium tolerance. Ecotoxicol. Environ. Saf. 2021, 218, 112285. [Google Scholar] [CrossRef]
- Waheed, A.; Haxim, Y.; Islam, W.; Ahmad, M.; Ali, S.; Wen, X. Impact of cadmium stress on growth and physio-biochemical attributes of Eruca sativa mill. Plants 2022, 11, 2981. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M.N.V. Iron and manganese-assisted cadmium tolerance in Oryza sativa L.: Lowering of rhizotoxicity next to functional photosynthesis. Planta 2015, 241, 1519–1528. [Google Scholar] [CrossRef]
- Chen, X.J.; Tao, H.F.; Wu, Y.Z.; Xu, X.M. Effects of cadmium on metabolism of photosynthetic pigment and photosynthetic system in Lactuca sativa L. revealed by physiological and proteomics analysis. Sci. Hortic. 2022, 305, 111371. [Google Scholar] [CrossRef]
- Abidli, A.; Huang, Y.; Rejeb, Z.B. Sustainable and efficient technologies for removal and recovery of toxic and valuable metals from wastewater: Recent progress, challenges, and future perspectives. Chemosphere 2022, 292, 133102. [Google Scholar]
- Raheem, A.; Prinsen, P.; Vuppaladadiyam, A.K. A review on sustainable microalgae based biofuel and bioenergy production: Recent developments. J. Clean. Prod. 2018, 181, 42–59. [Google Scholar]
- Seth, C.S. A review on mechanisms of plant tolerance and role of transgenic plants in environmental clean-up. Bot. Rev. 2012, 78, 32–62. [Google Scholar]
- Sharma, P.; Ngo, H.H.; Khanal, S.; Larroche, C.; Kim, S.H.; Pandey, A. Efficiency of transporter genes and proteins in hyperaccumulator plants for metals tolerance in wastewater treatment: Sustainable technique for metal detoxification. Environ. Technol. Innov. 2021, 23, 101725. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Gautam, S.; Landi, M.; Kandhol, N.; Araniti, F. Heavy metal induced regulation of plant biology: Recent insights. Physiol. Plant. 2022, 174, e13688. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, C.L.; Bauchan, G.R.; Murphy, C.A.; Tang, Y.T.; Pooley, C.; Davis, A.P. Accumulation of zinc and cadmium and localization of zinc in Picris divaricata Vant. Environ. Exp. Bot. 2013, 87, 1–9. [Google Scholar] [CrossRef]
- Mohd-Radzman, N.A.; Drapek, C. Compartmentalisation: A strategy for optimising symbiosis and tradeoff management. Plant Cell Environ. 2023, 46, 2998–3011. [Google Scholar] [CrossRef]
- Ru, C.; Hu, X.; Chen, D.; Wang, W.; Song, T. Heat and drought priming induce tolerance to subsequent heat and drought stress by regulating leaf photosynthesis, root morphology, and antioxidant defense in maize seedlings. Environ. Exp. Bot. 2022, 202, 105010. [Google Scholar] [CrossRef]
- Li, Y.; Rahman, S.U.; Qiu, Z.; Shahzad, S.M.; Nawaz, M.F.; Huang, J. Toxic effects of cadmium on the physiological and biochemical attributes of plants, and phytoremediation strategies: A review. Environ. Pollut. 2023, 325, 121433. [Google Scholar]
- Liu, Y.; Wiren, N. Integration of nutrient and water availabilities via auxin into the root developmental program. Curr. Opin. Plant Biol. 2022, 65, 102117. [Google Scholar] [CrossRef]
- Zhu, S.; Luo, L.; Zhang, X.; Zhao, M.; Wang, X.; Zhang, J. Study on the relationship of root morphology and phosphorus absorption efficiency with phosphorus uptake capacity in 235 peanut (Arachis hypogaea L.) germplasms. Front. Environ. Sci. 2022, 10, 855815. [Google Scholar] [CrossRef]
- Abdullah, A.; Abdullah, A.Z.; Ahmed, M. A review on recent developments and progress in sustainable acrolein production through catalytic dehydration of bio-renewable glycerol. J. Clean. Prod. 2022, 341, 130876. [Google Scholar]
- Li, T.; Yang, X.; Lu, L.; Islam, E.; He, Z. Effects of zinc and cadmium interactions on root morphology and metal translocation in a hyperaccumulating species under hydroponic conditions. J. Hazard. Mater. 2009, 169, 734–741. [Google Scholar] [CrossRef]
- Liu, S.; Tang, Y.; Qiu, R.; Ying, R.; Ge, R.; Ji, X. Responses of carbonic anhydrase to cadmium in the zinc/cadmium hyperaccumulator Picris divaricata vant. Pedosphere 2016, 26, 709–716. [Google Scholar] [CrossRef]
- Xu, N.; Sun, Y.; Wang, Y.; Cui, Y.; Jiang, Y.; Zhang, C. Hormesis effects in tomato plant growth and photosynthesis due to acephate exposure based on physiology and transcriptomic analysis. Pest Manag. Sci. 2023, 79, 2029–2039. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis mediates acquired resilience: Using plant-derived chemicals to enhance health. Annu. Rev. Food Sci. Technol. 2021, 12, 355–381. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Nayak, M.; Ghosh, A. A review on co-culturing of microalgae: A greener strategy towards sustainable biofuels production. Sci. Total Environ. 2022, 802, 149765. [Google Scholar] [PubMed]
- Zhou, W.; Xin, J.; Tian, R. Photosynthetic response, antioxidase activity, and cadmium uptake and translocation in monochoria korsakowii with cadmium exposure. Water Sci. Technol. 2022, 86, 2974–2986. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.T.; Li, L.Y.; Duan, Q.X.; Liu, X.L.; Chen, M. Progress in our understanding of plant responses to the stress of heavy metal cadmium. Plant Signal. Behav. 2021, 16, 1836884. [Google Scholar] [CrossRef]
- Parmar, P.; Kumari, N.; Sharma, V. Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress. Bot. Stud. 2013, 54, 45. [Google Scholar] [CrossRef]
- She, W.; Cui, G.X.; Li, X.L.; Su, X.H.; Jie, Y.C.; Yang, R.F. Characterization of cadmium concentration and translocation among ramie cultivars as affected by zinc and iron deficiency. Acta Physiol. Plant. 2018, 40, 104. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef]
- Hoshika, Y.; Fares, S.; Pellegrini, E.; Conte, A.; Paoletti, E. Water use strategy affects avoidance of ozone stress by stomatal closure in mediterranean trees—A modelling analysis. Plant Cell Environ. 2020, 43, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.V.D.; Bundervoet, B.; Verstraete, W. A sustainable, carbon neutral methane oxidation by a partnership of methane oxidizing communities and microalgae. Water Res. 2011, 45, 2845–2854. [Google Scholar] [PubMed]
- Tang, L.; Ying, R.R.; Jiang, D.; Zeng, X.W.; Morel, J.L.; Tang, Y.T. Impaired leaf CO2 diffusion mediates cd-induced inhibition of photosynthesis in the Zn/Cd hyperaccumulator Picris divaricata. Plant Physiol. Biochem. 2013, 73, 70–76. [Google Scholar] [CrossRef]
- Lee, H.J.; Abdula, S.E.; Jang, D.W.; Park, S.H.; Yoon, U.H.; Jung, Y.J. Over expression of the glutamine synthetase gene modulates oxidative stress response in rice after exposure to cadmium stress. Plant Cell Rep. 2013, 32, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.R.; Cai, Q.S. Photosynthetic and anatomic responses of peanut leaves to cadmium stress. Photosynthetica 2008, 46, 627–630. [Google Scholar] [CrossRef]
- Niinemets, U.; Diaz-Espejo, A.; Flexas, J.; Galmes, J.; Warren, C.R. Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field. J. Exp. Bot. 2009, 60, 2249–2270. [Google Scholar]
- Okcu, G.D.; Eustance, E.; Lai, Y.J.S. Evaluation of co-culturing a diatom and a coccolithophore using different silicate concentrations. Sci. Total Environ. 2021, 769, 145217. [Google Scholar] [CrossRef]
- Bunce, J.A. Use of the response of photosynthesis to oxygen to estimate mesophyll conductance to carbon dioxide in water-stressed soybean leaves. Plant Cell Environ. 2009, 32, 875–881. [Google Scholar] [CrossRef]
- Sagardoy, R.; Vazquez, S.; Florez-Sarasa, I.D.; Albacete, A.; Ribas-Carbo, M.; Flexas, J. Stomatal and mesophyll conductances to CO2 are the main limitations to photosynthesis in sugar beet (Beta vulgaris L.) plants grown with excess zinc. New Phytol. 2010, 187, 145–158. [Google Scholar] [CrossRef]
- Harley, P.C.; Loreto, F.; Di Marco, G.; Sharkey, T.D. Theoretical Considerations when Estimating the Mesophyll Conductance to CO2 Flux by Analysis of the Response of Photosynthesis to CO2. Plant Physiol. 1992, 98, 1429–1436. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Portis, A.R.; Nakano, H.; Von Caemmerer, S.; Long, S.P. Temperature Response of Mesophyll Conductance. Implications for the Determination of Rubisco Enzyme Kinetics and for Limitations to Photosynthesis in Vivo. Plant Physiol. 2002, 130, 1992–1998. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Firth, P.M.; Wetselaar, R.; Weir, B. On the Gaseous Exchange of Ammonia between Leaves and the Environment: Determination of the Ammonia Compensation Point. Plant Physiol. 1980, 66, 710–714. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (Cd μM) | Root Length (cm) | Average Root Diameter (mm) | Total Surface Area of Root System (cm2) | Total Volume of Root System (cm3) |
|---|---|---|---|---|
| 0 | 1681.8 ± 154.3 b | 0.30 ± 0.03 a | 156.46 ± 5.80 b | 1.17 ± 0.17 c |
| 5 | 2555.4 ± 356.8 a | 0.31 ± 0.02 a | 252.92 ± 52.91 a | 2.00 ± 0.56 ab |
| 10 | 2429.2 ± 139.2 a | 0.35 ± 0.02 a | 262.90 ± 23.20 a | 2.27 ± 0.33 a |
| 25 | 2741.0 ± 196.8 a | 0.34 ± 0.01 a | 288.28 ± 10.03 a | 2.43 ± 0.07 a |
| 50 | 1501.3 ± 385.8 b | 0.33 ± 0.00 a | 155.79 ± 38.02 b | 1.27 ± 0.30 bc |
| 75 | 1678.0 ± 473.5 b | 0.31 ± 0.03 a | 165.18 ± 57.93 b | 1.30 ± 0.55 bc |
| Treatment (Cd μM) | Fe (mg kg−1) | Mg (g kg−1) | Mn (mg kg−1) | Ca (g kg−1) | Cu (mg kg−1) | Zn (mg kg−1) |
|---|---|---|---|---|---|---|
| 0 | 227.4 ± 52.8 a | 6.58 ± 0.47 a | 234.5 ± 123.1 a | 22.9 ± 2.59 a | 3.11 ± 0.50 a | 37.2 ± 10.2 a |
| 5 | 144.1 ± 13.0 b | 6.57 ± 0.62 a | 307.7 ± 164.9 a | 22.0 ± 1.83 a | 2.09 ± 1.62 a | 30.8 ± 2.57 a |
| 10 | 153.9 ± 45.8 b | 5.77 ± 0.44 a | 222.1 ± 45.6 a | 20.1 ± 0.33 a | 3.27 ± 0.15 a | 35.0 ± 7.04 a |
| 25 | 164.7 ± 14.6 ab | 5.28 ± 1.12 a | 259.1 ± 76.1 a | 17.8 ± 0.73 a | 2.95 ± 1.14 a | 22.9 ± 8.50 a |
| 50 | 173.8 ± 40.6 ab | 5.06 ± 0.26 a | 449.3 ± 174.1 a | 20.2 ± 4.62 a | 4.62 ± 0.85 a | 22.1 ± 14.3 a |
| 75 | 201.8 ± 21.8 ab | 5.81 ± 1.47 a | 281.7 ± 132.1 a | 20.4 ± 5.66 a | 4.40 ± 2.57 a | 26.7 ± 11.4 a |
| Treatment (Cd μM) | SL | MCL | BL | NL | DL | TL |
|---|---|---|---|---|---|---|
| 0 | 9.0 ± 1.3 c | 4.2 ± 3.3 c | 5.1 ± 1.2 a | 9.7 ± 2.7 c | 13.6 ± 2.3 c | 18.7 ± 2.4 c |
| 10 | 12.1 ± 0.3 c | 2.6 ± 0.4 c | 0.6 ± 0.0 c | 3.1 ± 0.4 c | 14.7 ± 0.1 c | 15.3 ± 0.2 c |
| 25 | 17.0 ± 1.9 b | 11.7 ± 2.1 b | 0.5 ± 0.2 c | 12.2 ± 2.0 c | 28.7 ± 1.8 b | 39.2 ± 2.0 b |
| 50 | 23.0 ± 1.5 b | 18.5 ± 3.0 b | 2.9 ± 0.8 b | 21.4 ± 4.0 b | 41.5 ± 3.3 b | 44.4 ± 3.7 b |
| 75 | 28.8 ± 2.0 a | 36.8 ± 7.2 a | 3.1 ± 0.4 b | 40.0 ± 7.2 a | 65.7 ± 6.2 a | 68.8 ± 6.3 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Zhao, C.; Li, Q.; Feng, Y.; Zhang, X.; Lu, Y.; Ying, R.; Yin, A.; Ji, W. Heavy Metals Can Affect Plant Morphology and Limit Plant Growth and Photosynthesis Processes. Agronomy 2023, 13, 2601. https://doi.org/10.3390/agronomy13102601
Hu Z, Zhao C, Li Q, Feng Y, Zhang X, Lu Y, Ying R, Yin A, Ji W. Heavy Metals Can Affect Plant Morphology and Limit Plant Growth and Photosynthesis Processes. Agronomy. 2023; 13(10):2601. https://doi.org/10.3390/agronomy13102601
Chicago/Turabian StyleHu, Zhewei, Caiyi Zhao, Qi Li, Yanhong Feng, Xiaoyu Zhang, Yuanyuan Lu, Rongrong Ying, Aijing Yin, and Wenbing Ji. 2023. "Heavy Metals Can Affect Plant Morphology and Limit Plant Growth and Photosynthesis Processes" Agronomy 13, no. 10: 2601. https://doi.org/10.3390/agronomy13102601