
The Impact of Marasmius tricolor 310b on the Degradation of Cellulose in Rapeseed Straw Composting

1
College of Agriculture and Animal Husbandry, Qinghai University, Xining 810016, China
2
Qinghai Province Grassland Improvement Experimental Station, Gonghe 813000, China
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(12), 3012; https://doi.org/10.3390/agronomy13123012
Submission received: 2 November 2023
/
Revised: 28 November 2023
/
Accepted: 1 December 2023
/
Published: 8 December 2023
Abstract
:Composting, a vital process for organic waste management, hinges on intricate microbial dynamics. The role of specific microbial inoculants, such as fungi, in shaping these dynamics remains an area of keen interest. This study aimed to elucidate the impact of the fungal inoculant Marasmius tricolor 310b on composting dynamics, with a focus on temperature variations and the degradation of lignocellulosic components. We conducted a comprehensive composting experiment with four treatments: pure rapeseed straw, rapeseed straw with Marasmius tricolor 310b, a combination of rapeseed straw and pig manure, and the combination with Marasmius tricolor 310b. Temperature dynamics, lignocellulosic degradation rates, and fungal community structures were meticulously analyzed. While Marasmius tricolor 310b did not significantly alter the temperature profile, its presence markedly enhanced the degradation rates of cellulose and lignin. With the introduction of Marasmius tricolor 310b, the degradation rate of cellulose in the rapeseed straw compost increased from 0.0319 (ST) to 0.0419 (STM). In the combined rapeseed straw and pig manure compost, this rate rose from 0.0422 (STPM) to 0.045 (STPMM). For hemicellulose, the degradation rate in the rapeseed straw compost increased from 0.0103 (ST) to 0.014 (STM). However, in the combined compost, it slightly decreased from 0.0146 (STPM) to 0.013 (STPMM). As for lignin, its degradation rate in the rapeseed straw compost saw an increase from 0.0024 (ST) to 0.00366 (STM). In the combined compost, the rate rose from 0.00302 (STPM) to 0.00402 (STPMM). Fungal community analyses revealed discernible shifts in structure and diversity upon the fungus’s introduction. Network analyses further highlighted a highly interconnected microbial community, suggesting its robustness against perturbations. Temporal dynamics underscored the intricate interplay of microbial interactions, substrate availability, and external inoculants. The introduction of Marasmius tricolor 310b profoundly influences composting dynamics, offering valuable insights into microbial community interactions and presenting potential avenues for optimizing composting strategies.
1. Introduction
Composting stands as a pivotal method for managing organic waste, contributing to both environmental sustainability and agricultural productivity [1,2,3]. While its benefits are well-established, optimizing the composting process, particularly with specific agricultural residues like rapeseed straw, remains a significant scientific challenge [4]. Rapeseed straw, a byproduct of one of the world’s major oilseed crops, possesses substantial potential for applications such as bioenergy production, biorefinery processes, and mushroom cultivation [5,6,7]. Composting rapeseed straw offers a dual advantage—addressing agricultural waste challenges and producing valuable organic amendments for sustainable agriculture [8]. Marasmius tricolor 310b, a filamentous fungus renowned for its enzymatic capabilities, introduces a unique dimension to composting practices [9,10]. Its capacity to produce cellulases and ligninases positions it as a promising candidate for enhancing the breakdown of tough plant polymers, especially in lignocellulosic-rich materials like rapeseed straw [11]. Integrating Marasmius tricolor 310b into composting systems has the potential to expedite decomposition, improve compost quality, and enrich it with essential nutrients [12]. Despite the acknowledged benefits of composting and the potential of Marasmius tricolor 310b, there exists a gap in the literature regarding their combined application, especially in the context of utilizing agricultural residues like rapeseed straw. This study aims to fill this gap by investigating the impact of Marasmius tricolor 310b on the composting process, fungal community structure, and nutrient transformation during composting with rapeseed straw.
Globally, the agricultural sector faces the dual challenge of ensuring food security and managing vast amounts of agricultural residues [13]. Rapeseed, as a major oilseed crop, plays a significant role in the global agricultural landscape. Not only does it contribute to the edible oil market, but its cultivation also results in substantial amounts of residues, primarily in the form of straw [14]. Addressing the sustainable management of these residues is crucial both from an environmental and economic perspective [15]. While various methods exist for the disposal and utilization of rapeseed straw, many of these techniques, such as burning, have environmental implications [14,16]. Biodegradation, especially using specific microbial strains, emerges as a promising alternative [17]. The potential of biotechnological approaches, harnessing the capabilities of fungi like Marasmius tricolor 310b, offers an avenue to not only manage these residues but also derive value-added products [18]. Improper management of agricultural residues can lead to environmental concerns, including greenhouse gas emissions, soil degradation, and water pollution [19]. Embracing sustainable strategies, such as composting with efficient microbial strains, can mitigate these impacts. Such approaches align with global sustainability goals, promoting a circular economy where waste is transformed into valuable resources [20].
A thorough review of previous studies highlights the significance of composting in sustainable waste management. Additionally, research on the potential applications of rapeseed straw and the enzymatic capabilities of Marasmius tricolor 310b underscores the rationale for exploring their synergistic integration in composting. Despite the valuable insights provided by previous research, there is a lack of comprehensive exploration in the literature regarding the synergistic application of Marasmius tricolor 310b in composting systems utilizing agricultural residues, particularly rapeseed straw. This study aims to fill this gap by examining the specific contributions of Marasmius tricolor 310b to the composting of rapeseed straw and its implications for fungal communities and nutrient dynamics.
Through an outdoor composting experiment, we established four treatments: pure rapeseed straw (ST), rapeseed straw with Marasmius tricolor 310b (STM), pig manure mixed with rapeseed straw (STPM), and pig manure mixed with rapeseed straw supplemented with Marasmius tricolor 310b (STPMM). The composting process lasted for nearly two months. This study aims to (i) explore the potential of Marasmius tricolor 310b in enhancing the degradation rate of lignocellulosic components in rapeseed straw during composting, (ii) investigate the impact of adding Marasmius tricolor 310b on the fungal community structure within the composting environment, and (iii) assess how the introduction of Marasmius tricolor 310b might influence the transformation of essential nutrients in the compost and their subsequent availability for plant uptake.
2. Material and Methods
2.1. Inoculants
The test strains used were Marasmius tricolor 310b. This strain was isolated and screened from the alpine meadow soil of the East Qilian Mountains by our research group. This strain mainly possesses the ability to degrade lignocellulose. A culture medium primarily composed of wheat was used to proliferate the strains (100 g wheat, 1 g glucose, 1 g peptone, and 100 mL inorganic salt solution. The inorganic salt solution includes: 1 g KH2PO4, 0.1 g CaCl2·6H2O, 0.3 g MgSO4, 0.1 g NaCl, and 0.01 g FeCl3).
2.2. Inoculation, Composting, and Sampling
The experimental site was located in the Chengbei District of Xining City, Qinghai Province, with a geographical position of 101°76′ E longitude and 36°65′ N latitude, at an altitude of 2314 m (Figure S1). The area covers 138 km2 with an average annual temperature of 4.9 °C and annual precipitation of 379 mm. Rapeseed straw, procured from nearby companies in Xining, and fresh pig manure obtained from nearby farms were used as the raw materials. Marasmius tricolor 310b, previously screened and identified by our research group, was selected as the inoculant. External composting was conducted, employing a compost pile shaped as a cone with a base diameter of 1 m and a height of 0.7 m. The rapeseed straw was cut into 3~5 cm fragments, and pig manure was mixed in a mass ratio of 3:1 (pig manure 125 kg, rapeseed straw 41.5 kg) (Table 1) [21,22]. All treatment groups, except for the control group, received an inoculum of 100 g. The moisture content was carefully controlled to fall within the range of 50% to 60%, and the C/N ratio was maintained between 25:1 and 30:1 [23,24]. To ensure constant environmental conditions, temperature measurements were regularly taken at specific intervals within the compost pile using a model TGU73.100(Beijing Zhuoerhuihua Technology Co., Ltd., Beijing, China). Continuous monitoring and adjustments were made to maintain the desired temperature range throughout the composting process. The detailed physicochemical properties of the raw materials, including moisture content, C/N ratio, and temperature determination methods, are provided to offer a comprehensive understanding of the research process. The SD123-R60(Shandong 123 Internet of Things Technology Co., Ltd., Dezhou, China) was employed for accurate temperature readings, providing a robust basis for our observations and conclusions. Measurement errors were determined through a careful analysis of the data using a validated method based on repeatability and analysis of variance (ANOVA). The precision of measurements was ensured by employing model TGU73.100 and SD123-R60 for data collection. These equipment were known for their accuracy and reliability, and they adhere to international standards.
2.3. Determination of Cellulose, Hemicellulose, and Lignin
Neutral detergent fiber (NDF) determination [25]: (i) Samples were dried and ground to pass through a 40-mesh sieve. (ii) Approximately 0.6 g of the ground sample was placed in a crucible, to which 100 mL of neutral detergent solution and 1 mL of octanol were added. The mixture was then boiled for 50 min, maintaining a gentle boiling state throughout. (iii) After boiling, the samples were washed with distilled water and acetone until the waste liquid was neutral. (iv) The washed samples were dried in an oven at 105 °C for about 4 h and then weighed. (v) Calculation of the neutral detergent fiber (NDF) mass fraction (W) is as follows:
m: sample weight (g), m1: weight of the crucible with sand core (g) and m2: weight of the crucible with sand core and residue after drying (g).
Acid detergent fiber (ADF) determination [26]: (i) The procedure is similar to the NDF determination. However, an acid detergent solution is used instead of a neutral one. (ii) Calculation of acid detergent fiber (ADF) mass fraction (G) is as follows:
m: sample weight (g), m1: weight of the crucible with sand core (g), and m2: weight of the crucible with sand core and residue after drying (g).
Acid detergent lignin (ADL) determination [27]: (i) The acid-washed samples were dried and then treated with 12 mol/L sulfuric acid for 3 h at room temperature. (ii) The acid-treated samples were filtered immediately and washed until neutral. (iii) The samples were then dried in an oven at 105 °C for about 4 h, weighed, placed in a crucible, and shed in a muffle furnace at 510 °C for 2 h. After cooling, the samples were weighed again. (iv) Calculation of acid detergent lignin (ADL) mass fraction (Q) is as follows:
m: sample weight (g), m0: weight of the crucible (g), m1: weight of the crucible and ash after ignition (g), and m2: weight of the crucible with sand core and residue after acid detergent washing (g).
Calculation of cellulose, lignin, and hemicellulose: (i) Hemicellulose (%) = NDF − ADF. (ii) Cellulose (%) = ADF − ADL. (iii) Lignin (%) = ADL − Ash content. The determination of neutral detergent fiber (NDF) and acid detergent fiber (ADF) was carried out using the F800 fiber analyzer(Shandong Haineng Scientific Instrument Co., Ltd, Shandong, China), which provides accurate measurements of these components in plant materials.
2.4. DNA Extraction, PCR Amplification, and Sequencing Analysis
The total DNA from the compost samples was extracted using the Mobio PowerSoil DNA Isolation Kit (MOBIO Laboratories, Carlsbad, CA, USA) following the manufacturer’s instructions. The quality and concentration of the extracted DNA were determined using a Nanodrop2000 spectrophotometer (Thermo Fisher Scientific Inc., Shanghai, China). Only DNA samples with a concentration of 10 ng/μL or higher and an A260/280 ratio between 1.8 and 2.0 were considered suitable for further analysis. If the DNA did not meet these criteria, the extraction process was repeated. The fungal ITS (Internal Transcribed Spacer) region was amplified using ITS1F and ITS2R primers, which are commonly used for fungal community analysis. PCR reactions were performed in a 25 μL mixture containing 12.5 μL of PCR master mix, 1 μL of each primer, 2 μL of template DNA, and 8.5 μL of sterile distilled water. The PCR conditions were as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s, with a final extension at 72 °C for 10 min. The PCR products were purified using a gel extraction kit (Shenggong Biotechnology (Shanghai) Co., Ltd., Shanghai, China.) and quantified using a Qubit fluorometer (Thermo Fisher Scientific Inc., Shanghai, China). Purified amplicons were pooled in equimolar concentrations and paired-end sequenced (2 × 300) on an Illumina MiSeq platform according to the manufacturer’s recommendations.
The total DNA from the compost samples was extracted using the Mobio PowerSoil DNA Isolation Kit (MOBIO Laboratories, Carlsbad, CA, USA) following the manufacturer’s instructions. The quality and concentration of the extracted DNA were determined using a Nanodrop2000 spectrophotometer (Thermo Fisher Scientific Inc., Shanghai, China). Only DNA samples with a concentration of 10 ng/μL or higher and an A260/280 ratio between 1.8 and 2.0 were considered suitable for further analysis. If the DNA did not meet these criteria, the extraction process was repeated. The fungal ITS (Internal Transcribed Spacer) region was amplified using ITS1F and ITS2R primers, which are commonly used for fungal community analysis. PCR reactions were performed in a 25 μL mixture containing 12.5 μL of PCR master mix, 1 μL of each primer, 2 μL of template DNA, and 8.5 μL of sterile distilled water. The PCR conditions were as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s, with a final extension at 72 °C for 10 min. The PCR products were purified using a gel extraction kit (Shenggong Biotechnology (Shanghai) Co., Ltd., Shanghai, China.) and quantified using a Qubit fluorometer (Thermo Fisher Scientific Inc., Shanghai, China). Purified amplicons were pooled in equimolar concentrations and paired-end sequenced (2 × 300) on an Illumina MiSeq platform according to the manufacturer’s recommendations.
2.5. Sequence Processing
The ITS rRNA gene raw reads were processed through a widely used sequence analysis platform available at (http://dmap.denglab.org.cn, accessed on 18 November 2023). This platform integrates several bioinformatics tools designed for quality assurance, read trimming, and taxonomic categorization [28]. Initially, reads were mapped to their respective samples based on their barcode identifiers using the “detected barcodes” function, followed by barcode removal. Subsequently, paired forward and reverse reads were merged using FLASH utility [29], and ambiguous base-containing reads were filtered out with the Btrim software [30]. For chimera identification in the fungal community, we employed the ITS RefSeq database [31]. Considering singletons as potential rare species [32], we clustered sequences into operational taxonomic units (OTUs) at a 97% similarity level using the Unoise algorithm based on Vsearch [33]. For the ITS gene sequences, the ITSx utility helped in pinpointing and extracting ITS regions. Post-OTU table generation, we standardized the reads to 50,000 sequences for the fungal dataset. Lastly, the Ribosomal Database Project (RDP) classifier, in conjunction with the UNITE database [34], was employed to categorize fungal OTUs, maintaining confidence values above 0.8.
2.6. Molecular Ecological Network (MEN) Construction
To develop molecular ecological networks, we employed the random matrix theory (RMT) methodology as outlined by Deng and colleagues [35] to create intra-domain ecological networks, termed MEN. The freely accessible tool at http://dmap.denglab.org.cn, accessed on 18 November 2023 [36] facilitated this network development. From the RMT-derived Spearman correlations for microbial communities, we selected those with threshold values of r ≥ 0.76 specific to the fungal community. Visualization of the networks was achieved using platforms like Gephi (0.9.2) and Cytoscape (3.5.1). To determine the significance of each MEN, the Maslov–Sneppen method was employed to generate 100 random networks [37]. Based on their intra-module (Zi) and inter-module (Pi) connectivity values, nodes were categorized into peripherals, connectors, module hubs, and network hubs [38]. In these molecular ecological networks, module hubs, connectors, and network hubs were deemed keystone species. The cohesion method, indicative of cooperative or competitive interaction levels [39], was also applied to indirectly gauge complexity.
To gauge community resilience, robustness indices were utilized as delineated in prior research [40]. The robustness index was derived in a two-step process. Initially, the abundance-weighted mean interaction strength for each node i (wMISi) was computed using species j’s relative abundance (bj) and the association strength between species i and j (sij) was gauged by the Pearson correlation coefficient.
Subsequently, nodes with wMISi values ≤ 0 were excluded from the network, and the proportion of the remaining nodes was presented as the network’s robustness.
Furthermore, the cohesion index was employed to measure network intricacy, a metric adjusted for the null model used to consider abundance-weighting [39]. This index enabled the evaluation of connectivity levels among community entities. Two cohesion values (positive and negative) were derived based on pairwise correlations. Positive correlations potentially signify facilitation/mutualism among taxa, indicating ecological or functional parallels, whereas negative correlations might indicate competition, pointing to distinct niche requirements among taxa. The cohesion values were calculated as follows:
and
where ai is the abundance of OTU i in the sample j and is the connectedness.
2.7. Data Interpretation and Analysis
We utilized a publicly accessible analytical tool found at http://dmap.denglab.org.cn (accessed on 18 November 2023) for our statistical evaluations. To gauge the alpha diversity within microbial communities, we calculated the richness index, which quantified the observed species from the standardized OTU table. The Mantel test was employed to explore the relationship between soil attributes and microbial compositions. Techniques such as MRPP, ANOSIM, and PERMANOVA were used to assess variations in community composition. Differences in soil physicochemical attributes and diversities across treatment pairs were tested using ANOVA, with subsequent Tukey post hoc and LSD tests pinpointing significant variations. The one-sample Student’s t-test facilitated the identification of distinct features between treatment pairs and the variance between observed and randomized networks. Lastly, the random forest’s (RF) methodology was harnessed to pinpoint key environmental determinants impacting the prevalence of pathogenic fungi.
To further understand the temporal dynamics of microbial communities, we employed the time–decay relationship (TDR) model. This model is designed to assess the stability and succession patterns of microbial communities over time. By plotting the community similarity against time lag, the TDR model provides insights into how rapidly microbial communities change and the potential driving factors behind these shifts. The slope of the TDR curve can be interpreted as the rate of community change, with steeper slopes indicating faster community turnover. This approach has been particularly useful in studies where understanding the temporal stability and resilience of microbial communities is crucial.
3. Results
3.1. Changes in Temperature and Degradation of Lignocellulosic Components
Throughout the composting process, distinct variations were observed in temperature and the content of lignocellulosic components, namely cellulose, hemicellulose, and lignin. As shown in Figure S2, the ambient temperature ranged from 9.5 °C to 21.9 °C. The temperature of the rapeseed straw compost varied between 16.4 °C and 51.2 °C, while the compost comprising both rapeseed straw and pig manure exhibited a temperature range of 21.6 °C to 54.9 °C. Under composting conditions, the addition of Marasmius tricolor 310b did not result in significant temperature differences compared to the batches without the fungus. Furthermore, the temperature in the compost with the combination of pig manure and rapeseed straw was notably higher than that of the compost with rapeseed straw alone. However, as the composting process progressed, there was a gradual decline in temperature, eventually approaching ambient levels (Figure S2).
Across the four composting treatments, the initial content of cellulose, hemicellulose, and lignin was approximately 0.55%, 0.20%, and 0.18%, respectively. With the introduction of Marasmius tricolor 310b, the degradation rate of cellulose in the rapeseed straw compost increased from 0.0319 (ST) to 0.0419 (STM). In the combined rapeseed straw and pig manure compost, this rate rose from 0.0422 (STPM) to 0.045 (STPMM). For hemicellulose, the degradation rate in the rapeseed straw compost increased from 0.0103 (ST) to 0.014 (STM). However, in the combined compost, it slightly decreased from 0.0146 (STPM) to 0.013 (STPMM). As for lignin, its degradation rate in the rapeseed straw compost saw an increase from 0.0024 (ST) to 0.00366 (STM). In the combined compost, the rate rose from 0.00302 (STPM) to 0.00402 (STPMM) (Figure 1).
3.2. Influence of Marasmius Tricolor 310b on Fungal Community Dynamics
The incorporation of Marasmius tricolor 310b into the composting matrix offered a distinctive avenue to probe its effects on the fungal community. This study meticulously examined both the diversity and compositional shifts to elucidate the ecological ramifications of this fungal integration. From the analysis, a total of 3295 zOTUs for fungi were identified. Alpha diversity metrics revealed a statistically significant reduction in observed richness upon the addition of Marasmius tricolor 310b (p < 0.05, ANOVA) when juxtaposed with control treatments (Figure 2a). Beta diversity, visualized via NMDS plots based on the Bray–Curtis distance (stress = 0.195), manifested clear clustering distinctions between treatments with and without the fungal addition (Figure 2b). This distinction was further corroborated by the dissimilarity test, highlighting marked differences between specific treatment pairs (Table S1). A core set of 32 species was consistently observed across all treatments (Figure 2c). At the phylum level, Ascomycota emerged as the predominant group, followed by Basidiomycota, Zygomycota, Chytridiomycota, Neocallimastigomycota, and Glomeromycota (Figure 2d). Genera ubiquitously detected across treatments encompassed Chaetomium, Thermomyces, Coprinellus, and several others, with a notable surge in the genus Marasmius, particularly the species Marasmius tricolor 310b, upon its addition (Figure 2e,f).
The random forest model pinpointed Talaromyces thermophilus as the most influential biomarker, with Marasmius tricolor 310b following closely (Figure 3). The biomarker list also identified several unclassified species, underscoring the intricate microbial diversity within the compost. Other significant biomarkers included Marasmius fiardii, Acremonium asperulatum, and Aspergillus fumigatus, among others. The prevalence of numerous unclassified entities hints at the presence of potentially novel species or strains meriting deeper exploration. In essence, this study sheds light on the pivotal microbial entities within the composting milieu and underscores the profound influence of Marasmius tricolor 310b on shaping these dynamics.
3.3. Inter-Domain Networks Analysis for Soil Microbial Communities under Composting Milieu
We employed molecular ecological networks (MENs) to elucidate the shifts in microbial community inter-relationships during the composting process (Figure 4). For a consistent comparison across treatments, MENs were constructed using a uniform threshold of 0.76 for the fungal community. The topological indices revealed average path lengths (GD) for fungal networks between 3.314 and 4.257, approximating the logarithms of the total node count. This, being higher than corresponding random networks, suggests the small-world network property typical of these microbial communities (Deng et al. 2012). The modularity of fungal networks varied between 0.308 and 0.615, surpassing the modularity of randomized counterparts, indicating a modular topology in all constructed networks (Tables S2 and S3).
Upon introducing Marasmius tricolor 310b, a reduction in network scale was evident (Figure 4 and Table S2). Zi-Pi analysis highlighted a diminished count of keystone species post-incubation in fungal networks (STM: 2 and STPMM: 2 nodes) compared to pre-incubation (ST: 15 and STPM: 5 nodes) (Table S3). Cohesion analysis for rapeseed straw composting revealed post-incubation fungal communities had enhanced positive cohesion, suggesting increased fungal cooperation, and elevated negative cohesion, indicating reduced competition (Figure 5a,b). For composts combining rapeseed straw and pig manure, post-incubation fungal communities also displayed heightened positive cohesion, but with an amplified negative cohesion value, suggesting intensified fungal competition (Figure 5a,b). ANOVA results underscored a significant robustness increase in post-incubation fungal communities within the mixed compost (p < 0.05), hinting at enhanced community stability in such environments (Figure 5c). In summary, Marasmius tricolor 310b’s introduction markedly influenced microbial community dynamics, emphasizing the resilience of these communities to environmental shift.
3.4. Impact of Inoculation on Temporal Stability of Fungal Communities in Compost
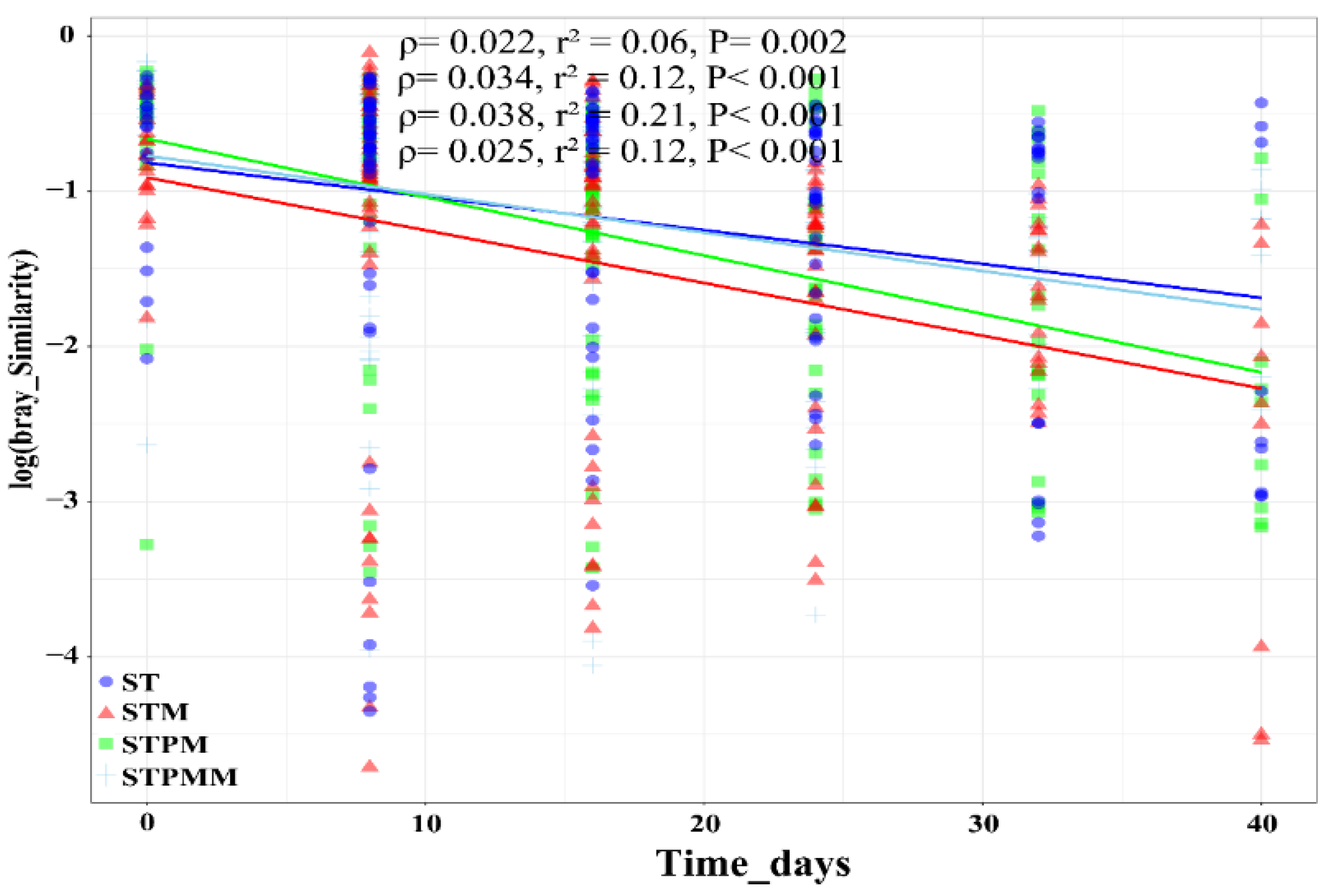
To delve deeper into the repercussions of inoculation on the temporal dynamics of compost microbial communities, we employed the time–decay relationship (TDR) model (Figure 6). This model gauges the temporal stability of fungal communities over the composting duration. The TDR analysis revealed a consistent time-decay trait across all treatments (ST: p = 0.002, STM: p < 0.001, STPM: p < 0.001, STPMM: p < 0.001), suggesting a natural progression in microbial community structure over time. Interestingly, for the rapeseed straw compost, inoculation appeared to heighten the temporal sensitivity of the fungal community. The rate escalated from an initial 0.022 to 0.034 post-inoculation. In contrast, for the combined rapeseed straw and pig manure compost, inoculation seemingly conferred a stabilizing effect, with the temporal sensitivity rate diminishing from 0.038 to 0.025. These findings underscore the nuanced role of inoculation in shaping the temporal dynamics of fungal communities. The differential responses observed between pure straw compost and its combination with pig manure highlight the intricate interplay of microbial interactions, substrate availability, and external inoculants in determining community stability.
4. Discussion
Composting, a biologically driven process, is a testament to the intricate and dynamic interplay of diverse microbial communities [41]. These communities orchestrate the transformation of organic waste into nutrient-rich compost [42], a process that is pivotal for sustainable agriculture and waste management. While the inherent microbial communities in compost play a central role, external factors, especially microbial inoculants, can significantly modulate the composting trajectory [41,43]. Inoculants, by introducing specific microbial strains or consortia, can potentially enhance the efficiency of composting by accelerating organic matter degradation, altering microbial community structures, or improving the overall quality of the end product [44,45]. Our study delves into the role of one such inoculant, Marasmius tricolor 310b, and its influence on composting dynamics. By focusing on the fungal community, which often plays a crucial role in lignocellulosic degradation, we aim to provide a comprehensive understanding of how the introduction of Marasmius tricolor 310b reshapes microbial interactions and composting outcomes.
4.1. Influence on Temperature and Lignocellulosic Degradation
Composting is a thermophilic process, where the rise in temperature is a direct consequence of microbial metabolic activities, particularly during the active phase of decomposition [46]. This temperature elevation not only aids in the breakdown of organic matter but also serves as a natural sanitizing agent, eliminating potential pathogens [47]. In our study, the temperature dynamics observed across different treatments provided a window into the microbial activities within the composting matrix. Despite the introduction of Marasmius tricolor 310b not causing a pronounced shift in the temperature profile, it is crucial to consider the multifaceted roles that fungi play in composting [48]. Fungi, especially those with lignocellulolytic capabilities, often contribute more towards substrate degradation than heat production [49]. This suggests that Marasmius tricolor 310b might be more involved in substrate utilization and community interactions than in rapid metabolic activities that generate heat. The enhanced degradation of cellulose and lignin in treatments with Marasmius tricolor 310b is particularly noteworthy. Lignocellulosic materials, being the primary constituents of plant biomass, are notoriously resistant to decomposition due to their complex structure [50]. The ability of certain fungi to produce a suite of enzymes, including cellulases and ligninases, allows them to effectively break down these materials [51]. Our findings resonate with this understanding, highlighting the potential of Marasmius tricolor 310b in augmenting lignocellulosic breakdown. This could have significant implications for composting practices, especially in scenarios where rapid degradation of plant residues is desired. Furthermore, the synergistic or antagonistic interactions between Marasmius tricolor 310b and native microbial communities in the compost could also play a role in determining the overall degradation efficiency. Such microbial interactions, often mediated by signaling molecules, nutrient competition, or predation, can profoundly influence community dynamics and composting outcomes [52]. Our results demonstrate consistency with prior research, emphasizing the importance of understanding the intricate interplay between temperature, microbial activities, and substrate degradation in composting processes [46,47,49].
4.2. Marasmius Tricolor 310b and Fungal Community Dynamics
The microbial composition within composting systems is inherently dynamic, subject to influences from various factors, including the introduction of specific microbial strains [53,54]. In our study, the inoculation of Marasmius tricolor 310b brought about notable alterations in the fungal community structure. A marked decrease in observed richness post-inoculation suggests a potential competitive advantage or niche specialization of certain fungal taxa, possibly driven by the presence of Marasmius tricolor 310b. Similar shifts in microbial diversity upon the introduction of specific strains have been observed in other studies, underscoring the profound influence of microbial inoculants on community dynamics [55]. Interestingly, despite the reduction in richness, the fungal community exhibited enhanced resilience, especially evident in the compost comprising both rapeseed straw and pig manure. This resilience might be attributed to the establishment of a more functionally redundant community, where multiple species can perform similar ecological roles, thereby ensuring stability even when faced with environmental perturbations [56]. The observed dominance of the phylum Ascomycota in our composting treatments resonates with previous findings [57,58]. Ascomycota, known for its diverse metabolic capabilities and adaptability, often emerges as a dominant player in composting environments, contributing significantly to organic matter decomposition [59,60,61]. Their dominance might also be indicative of the specific stages of composting, with Ascomycota being particularly active during the thermophilic and maturation phases [62]. Furthermore, the increased abundance of the genus Marasmius, especially Marasmius tricolor 310b, post-inoculation, provides insights into its potential interactions with native fungal species. Whether Marasmius tricolor 310b establishes itself through competitive exclusion or through synergistic interactions with other fungi warrants further investigation. Such insights can be pivotal in optimizing composting processes, especially when aiming for specific outcomes like enhanced lignocellulosic degradation or pathogen suppression.
4.3. Network Analysis and Community Interactions
Microbial ecological networks provide a lens through which the intricate web of interactions among community members can be visualized and understood [28,35]. In our study, the observed small-world property of the microbial networks is emblematic of a community characterized by a few highly connected nodes, ensuring efficient communication and a rapid response to environmental changes [63]. Such a property is crucial in dynamic environments like composting systems, where conditions can fluctuate and the community needs to adapt swiftly [64,65]. The modularity observed in our networks is particularly intriguing. High modularity suggests the presence of distinct microbial guilds or clusters [66] that might have specialized functions during the composting process. These microbial modules could be groups that have co-evolved to work synergistically, optimizing the breakdown of specific organic matter fractions or resisting certain antagonistic forces [38]. Upon the introduction of Marasmius tricolor 310b, a reduction in the overall network scale was evident. This could be interpreted in multiple ways. One possibility is that Marasmius tricolor 310b, being a potent decomposer, might streamline microbial interactions by either outcompeting certain taxa or forming synergistic relationships, thereby reducing the overall complexity of the network. Such microbial interactions, especially in environments rich in organic matter like composting systems, play a pivotal role in determining the efficiency of decomposition processes [67]. Another perspective could be that the introduction of Marasmius tricolor 310b creates a more stable environment, where fewer microbial interactions are needed to maintain system equilibrium. Furthermore, the observed changes in network properties upon fungal inoculation underscore the importance of understanding microbial interactions when introducing external agents. Such insights can guide the optimization of composting processes, ensuring that desired outcomes, be it rapid decomposition or pathogen suppression, are achieved.
4.4. Temporal Stability and Community Dynamics
Temporal dynamics in microbial communities are a reflection of the ever-evolving nature of ecosystems, especially in environments as dynamic as composting systems [68,69]. The time–decay trait observed across our treatments underscores a consistent and natural progression in microbial community structure over the composting duration. This progression can be attributed to a myriad of factors, including changing substrate availability, microbial succession, and interactions between community members [70]. Interestingly, the differential temporal sensitivity observed between pure straw compost and its combination with pig manure provides a window into the multifaceted nature of microbial community dynamics. It’s evident that the nature and availability of organic substrates play a pivotal role in shaping these dynamics. Pure straw, being primarily lignocellulosic, offers a different set of challenges and opportunities for microbial communities compared to a mixture that includes nutrient-rich pig manure [71]. The latter not only provides a richer nutrient matrix but also introduces a diverse set of microbial taxa, potentially leading to more complex interactions and competition scenarios [72]. The introduction of external inoculants, such as Marasmius tricolor 310b, further complicates this dynamic. While inoculants can accelerate specific metabolic processes, they can also disrupt established microbial networks, leading to unforeseen consequences on community stability and function [73]. Previous studies have highlighted the importance of substrate availability in driving microbial succession during composting [74]. As organic matter gets degraded, the microbial community shifts to adapt to the changing nutrient landscape [75]. Early colonizers that thrive on easily available substrates give way to more specialized microbes capable of breaking down complex organic compounds [76]. Our findings resonate with this understanding, emphasizing the intricate interplay of microbial interactions, substrate availability, and external factors in determining temporal community dynamics.
Our findings underscore the potential of Marasmius tricolor 310b as a promising inoculant for enhancing composting efficiency, particularly in augmenting lignocellulosic breakdown. However, the broader ecological implications of its introduction, especially its long-term effects on microbial community structure and function, warrant further investigation. Future studies could delve deeper into the metabolic pathways activated upon Marasmius tricolor 310b introduction, offering a mechanistic understanding of its role in composting.
5. Conclusions
Our comprehensive study underscores the multifaceted influence of Marasmius tricolor 310b on composting dynamics, with a particular emphasis on the fungal community. The temperature dynamics observed during the composting process align with established knowledge, suggesting that microbial metabolic activities play a pivotal role in organic matter breakdown. The addition of Marasmius tricolor 310b, while not significantly altering the temperature profile, markedly influenced the degradation rates of lignocellulosic components, highlighting its potential in augmenting lignocellulosic breakdown. With the introduction of Marasmius tricolor 310b, the degradation rate of cellulose in the rapeseed straw compost increased from 0.0319 (ST) to 0.0419 (STM). In the combined rapeseed straw and pig manure compost, this rate rose from 0.0422 (STPM) to 0.045 (STPMM). For hemicellulose, the degradation rate in the rapeseed straw compost increased from 0.0103 (ST) to 0.014 (STM). However, in the combined compost, it slightly decreased from 0.0146 (STPM) to 0.013 (STPMM). As for lignin, its degradation rate in the rapeseed straw compost saw an increase from 0.0024 (ST) to 0.00366 (STM). In the combined compost, the rate rose from 0.00302 (STPM) to 0.00402 (STPMM). Furthermore, the fungal community exhibited discernible shifts in structure and diversity upon the introduction of Marasmius tricolor 310b. Network analyses revealed a highly interconnected microbial community, suggesting robustness against perturbations and a potential role of the fungus in streamlining microbial interactions. Temporal dynamics further emphasized the intricate interplay of microbial interactions, substrate availability, and external inoculants in shaping community progressions over time. In essence, the introduction of Marasmius tricolor 310b offers profound insights into microbial community dynamics, potentially paving the way for optimized composting strategies in the future.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13123012/s1, Table S1: The dissimilarity analysis between treatments; Table S2: The network properties; Table S3: Kernsoorten van het network; Figure S1: The experimental site and sampling method; Figure S2: Changes in composting temperature.
Author Contributions
G.L., designed the experiments. Z.W. and D.S., took samples and performed all data measurements. Z.W., contributed to the data analysis. Z.W., wrote the paper. G.L., revised the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Qinghai Provincial Science and Technology International Cooperation Special Program under the independent number [2023-HZ-803].
Data Availability Statement
The raw sequence data from this study were deposited in the SRA at the NCBI database with accession PRJNA1031796. The relevant data are available from the corresponding author on request.
Acknowledgments
We thank James Walter Voordeckers for carefully editing the grammar of the manuscript and for some valuable suggestions for this manuscript.
Conflicts of Interest
The authors declare no competing financial interests.
References
- Pergola, M.; Persiani, A.; Palese, A.M.; Di Meo, V.; Pastore, V.; D’Adamo, C.; Celano, G. Composting: The way for a sustainable agriculture. Appl. Soil Ecol. 2018, 123, 744–750. [Google Scholar] [CrossRef]
- Ishola, T.M.; Ishola, E.T. Composting and Sustainable Development. In Encyclopedia of Sustainability in Higher Education; Filho, W.L., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 272–279. [Google Scholar]
- Ayilara, M.S.; Olanrewaju, O.S.; Babalola, O.O.; Odeyemi, O. Waste Management through Composting: Challenges and Potentials. Sustainability 2020, 12, 4456. [Google Scholar] [CrossRef]
- Saini, S.; Sharma, K.K. Fungal lignocellulolytic enzymes and lignocellulose: A critical review on their contribution to multiproduct biorefinery and global biofuel research. Int. J. Biol. Macromol. 2021, 193, 2304–2319. [Google Scholar] [CrossRef] [PubMed]
- Mazhari Mousavi, S.M.; Hosseini, S.Z.; Resalati, H.; Mahdavi, S.; Rasooly, E. Garmaroody Papermaking potential of rapeseed straw, a new agricultural-based fiber source. J. Clean. Prod. 2013, 52, 420–424. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, Z.S.; Zhang, X.; Li, L.W.; Li, Y.Y.; Xu, H.M.; Li, X.X.; Yu, X.L.; Zhang, Z.S.; Liang, Y.Y.; et al. Highly selective and active CO2 reduction electrocatalysts based on cobalt phthalocyanine/carbon nanotube hybrid structures. Nat. Commun. 2017, 8, 14675. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Liu, K. Worldwide rapeseed (Brassica napus L.) research: A bibliometric analysis during 2011–2021. Oil Crop Sci. 2022, 7, 157–165. [Google Scholar]
- Yu, Q.G.; Hu, X.; Ma, J.W.; Ye, J.; Sun, W.C.; Wang, Q.; Lin, H. Effects of long-term organic material applications on soil carbon and nitrogen fractions in paddy fields. Soil Tillage Res. 2020, 196, 104483. [Google Scholar] [CrossRef]
- Henrissat, B.; Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 1997, 7, 637–644. [Google Scholar] [CrossRef]
- Grace, C.L. The genus Marasmius (Basidiomycota, Agaricales, Marasmiaceae) from Republic of São Tomé and Príncipe. West Afr. 2019, 414, 8. [Google Scholar] [CrossRef]
- Gupta, P.; Samant, K.; Sahu, A. Isolation of Cellulose-Degrading Bacteria and Determination of Their Cellulolytic Potential. Int. J. Microbiol. 2012, 2012, 578925. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, B.K.; Lee, B.H.; Jo, K.I.; Lee, N.K.; Chung, C.H.; Lee, Y.C.; Lee, J.W. Purification and characterization of cellulase produced by Bacillus amyoliquefaciens DL-3 utilizing rice hull. Bioresour. Technol. 2008, 99, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Hamid, S.; Yaseen, M. Global Agri-Food Sector: Challenges and Opportunities in COVID-19 Pandemic. Front. Sociol. 2021, 6, 647337. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.H.; Jiang, L.Y.; Mason, A.S.; Xiao, M.L.; Zhu, L.R.; Li, L.Z.; Zhou, Q.H.; Shen, C.J.; Huang, C.H. Research progress and strategies for multifunctional rapeseed: A case study of China. J. Integr. Agric. 2016, 15, 1673–1684. [Google Scholar] [CrossRef]
- Beal, T.; Massiot, E.; Arsenault, J.E.; Smith, M.R.; Hijmans, R.J. Global trends in dietary micronutrient supplies and estimated prevalence of inadequate intakes. PLoS ONE 2017, 12, e0175554. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.; Li, W.; Abdalla, N.S.; Ai, P.; Zhang, Y.L.; Abomohra, A.E.F. Innovative approach for rapeseed straw recycling using black solider fly larvae: Towards enhanced energy recovery. Renew. Energy 2022, 188, 211–222. [Google Scholar] [CrossRef]
- Khalid, S.A.; Elsherif, W.M. Types of Microorganisms for Biodegradation. In Handbook of Biodegradable Materials; Ali, G.A.M., Makhlouf, A.S.H., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 195–220. [Google Scholar]
- Bala, S.; Garg, D.; Sridhar, K.; Inbaraj, B.S.; Singh, R.; Kamma, S.; Tripathi, M.; Sharma, M. Transformation of Agro-Waste into Value-Added Bioproducts and Bioactive Compounds: Micro/Nano Formulations and Application in the Agri-Food-Pharma Sector. Bioengineering 2023, 10, 152. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in Agriculture: A Sustainable Approach to Increasing Climate Change Resilience. Front. Sustain. Food Syst. 2021, 5, 667546. [Google Scholar] [CrossRef]
- Garcia-Saravia Ortiz-de-Montellano, C.; Samani, P.; van der Meer, Y. How can the circular economy support the advancement of the Sustainable Development Goals (SDGs)? A comprehensive analysis. Sustain. Prod. Consum. 2023, 40, 352–362. [Google Scholar]
- Yao, S.T.; Lu, G.X.; Li, X.; Dang, N.; Wang, Y.C. Study on the degradation activity of oilseed rape straw by multi-enzymatic bacterial flora. J. Qinghai Univ. 2019, 37, 26–34. [Google Scholar]
- Zhang, X.P. Screening of Compound Bacteria Strains for Rape Straw Composting in Alpine Regions; Qinghai University: Xining, China, 2022. [Google Scholar]
- Wang, J.N. The Preparation and Evaluation of the Mixed Microbial Agents for Pig Manure Composting; Dalian University of Technology: Dalian, China, 2017. [Google Scholar]
- Sun, J.; Zhang, X.P.; Li, S.L.; Wang, Y.Y. Screening of fermentation strains of hog manure and rape straw in Qinghai Province. J. Qinghai Univ. 2022, 40, 55–61. [Google Scholar]
- GB/T 20806-2022; Determination of Neutral Detergent Fiber (NDF) in Feeds. National Administration for Market Regulation State Administration of Standardization Management: Shenzhen, China, 2022.
- NY/T 1459-2007; Determination of Acid Detergent Fiber (ADF) in Feeds. Industry Standard Agriculture: Beijing, China, 2007.
- GB/T 20805-2006; Determination of Acid Detergent Lignin (ADL) in Feeds. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of China: Shenzhen, China, 2006.
- Feng, K.; Zhang, Z.J.; Cai, W.W.; Liu, W.Z.; Xu, M.Y.; Yin, H.Q.; Wang, A.J.; He, Z.L.; Deng, Y. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y. Btrim: A fast, lightweight adapter and quality trimming program for next-generation sequencing technologies. Genomics 2011, 98, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef] [PubMed]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Küsel, K.; Rillig, M.C.; Rivett, D.W.; Salles, J.F.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.E. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016, 081257. [Google Scholar]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.H.; Yang, Y.F.; He, Z.L.; Luo, F.; Zhou, J.Z. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef]
- Feng, K.; Peng, X.; Zhang, Z.; Gu, S.S.; He, Q.; Shen, W.L.; Wang, Z.J.; Wang, D.R.; Hu, Q.L.; Li, Y.; et al. iNAP: An integrated network analysis pipeline for microbiome studies. iMeta 2022, 1, e13. [Google Scholar] [CrossRef]
- Bascompte, J.; Jordano, P.; Melián, C.J.; Olesen, J.M. The nested assembly of plant–animal mutualistic networks. Proc. Natl. Acad. Sci. USA 2003, 100, 9383–9387. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [PubMed]
- Herren, C.M.; McMahon, K.D. Cohesion: A method for quantifying the connectivity of microbial communities. ISME J. 2017, 11, 2426–2438. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. Food-web structure and network theory: The role of connectance and size. Proc. Natl. Acad. Sci. USA 2002, 99, 12917–12922. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, M.; Nandal, M.; Khosla, B. Microbes as vital additives for solid waste composting. Heliyon 2020, 6, e03343. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Sun, H.; Chen, E.; Yang, M.; Wu, C.; Sun, X.; Wang, Q. From waste to wealth: Innovations in organic solid waste composting. Environ. Res. 2023, 229, 115977. [Google Scholar] [CrossRef]
- Bello, A.; Han, Y.; Zhu, H.F.; Deng, L.T.; Yang, W.; Meng, Q.X.; Sun, Y.; Egbeagu, U.U.; Sheng, S.Y.; Wu, X.T.; et al. Microbial community composition, co-occurrence network pattern and nitrogen transformation genera response to biochar addition in cattle manure-maize straw composting. Sci. Total Environ. 2020, 721, 137759. [Google Scholar] [CrossRef]
- Oviedo-Ocaña, E.R.; Soto-Paz, J.; Domínguez, I.; Sanchez-Torres, V.; Komilis, D. A Systematic Review on the Application of Bacterial Inoculants and Microbial Consortia During Green Waste Composting. Waste Biomass Valorization 2022, 13, 3423–3444. [Google Scholar] [CrossRef]
- Jack, C.N.; Petipas, R.H.; Cheeke, T.E.; Rowland, J.R.L.; Friesen, M.L. Microbial Inoculants: Silver Bullet or Microbial Jurassic Park? Trends Microbiol. 2021, 29, 299–308. [Google Scholar] [CrossRef]
- Finore, I.; Feola, A.; Russo, L.; Cattaneo, A.; Di Donato, P.; Nicolaus, B.; Poli, A.; Romano, I. Thermophilic bacteria and their thermozymes in composting processes: A review. Chem. Biol. Technol. Agric. 2023, 10, 7. [Google Scholar] [CrossRef]
- Wang, P.; Han, S.; Lin, Y. Role of microbes and microbial dynamics during composting. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2023; pp. 169–220. [Google Scholar]
- Wright, C.; Gryganskyi, A.P.; Bonito, G. Fungi in Composting. In Fungal Applications in Sustainable Environmental Biotechnology; Purchase, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 3–28. [Google Scholar]
- Karnaouri, A.; Topakas, E.; Antonopoulou, I.; Christakopoulos, P. Genomic insights into the fungal lignocellulolytic system of Myceliophthora thermophila. Front. Microbiol. 2014, 5, 281. [Google Scholar] [CrossRef] [PubMed]
- Zoghlami, A.; Paës, G. Lignocellulosic Biomass: Understanding Recalcitrance and Predicting Hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Fungal laccases—Occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [PubMed]
- De Gannes, V.; Eudoxie, G.; Hickey, W. Insights into fungal communities in composts revealed by 454-pyrosequencing: Implications for human health and safety. Front. Microbiol. 2013, 4, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Chang, Y.; Tao, Y.; Zhang, H.; Lin, Y.; Deng, J.; Ma, T.; Ding, G.; Wei, Y.; Li, J. Insight into the dynamic microbial community and core bacteria in composting from different sources by advanced bioinformatics methods. Environ. Sci. Pollut. Res. 2023, 30, 8956–8966. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Shen, J.; Wang, X.; Xiao, H.; Yaser, A.Z.; Fu, J. Recent advances in research on microbial community in the composting process. Biomass Convers. Biorefinery 2023, 1–15. [Google Scholar] [CrossRef]
- Del Carmen Vargas-García, M.; Suárez-Estrella, F.F.; López, M.J.; Moreno, J. Influence of microbial inoculation and co-composting material on the evolution of humic-like substances during composting of horticultural wastes. Process Biochem. 2006, 41, 1438–1443. [Google Scholar] [CrossRef]
- Amelung, W.; Bossio, D.; de Vries, W.; Kögel-Knabner, I.; Lehmann, J.; Amundson, R.; Bol, R.; Collins, C.; Lal, R.; Leifeld, J.; et al. Towards a global-scale soil climate mitigation strategy. Nat. Commun. 2020, 11, 5427. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, X.; Sun, L.; Men, M.; Wang, B.; Deng, L.; Zhao, L.; Han, Y.; Jong, C.; Bi, R.X.; et al. Microecological insight to fungal structure and key fungal communities regulating nitrogen transformation based on spatial heterogeneity during cow manure composting by multi-angle and multi-aspect analyses. Waste Manag. 2022, 142, 132–142. [Google Scholar] [CrossRef]
- Ji, Y.; Cao, Y.; Wang, Y.; Wang, C.; Qin, Z.; Cai, W.; Yang, Y.; Yan, S.; Guo, X. Effects of adding lignocellulose-degrading microbial agents and biochar on nitrogen metabolism and microbial community succession during pig manure composting. Environ. Res. 2023, 239, 117400. [Google Scholar] [CrossRef]
- Challacombe, J.F.; Hesse, C.N.; Bramer, L.M.; McCue, L.A.; Lipton, M.; Purvine, S.; Nicora, C.; Gallegos-Graves, L.V.; Porras-Alfaro, A.; Kuske, C.R. Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis. BMC Genom. 2019, 20, 976. [Google Scholar] [CrossRef]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Chapter 8—Ascomycota. In Fossil Fungi; Elsevier: Amsterdam, The Netherlands, 2015; pp. 129–171. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef]
- Ryckeboer, J.; Mergaert, J.; Vaes, K.; Klammer, S.H.; Clercq, D.D.; Coosemans, J.; Insam, H.; Swings, J. A survey of bacteria and fungi occurring during composting and self-heating processes. Ann. Microbiol. 2003, 53, 349–410. [Google Scholar]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, H.; Schreiber, F. Small-World Property. In Encyclopedia of Systems Biology; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1957–1959. [Google Scholar]
- Barrat, A.; Weigt, M. On the properties of small-world network models. Eur. Phys. J. B Condens. Matter Complex Syst. 2000, 13, 547–560. [Google Scholar] [CrossRef]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling Interactions in the Microbiome: A Network Perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.B.; Martiny, A.C.; Martiny, J.B.H. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef]
- Ronda, C.; Wang, H.H. Engineering temporal dynamics in microbial communities. Curr. Opin. Microbiol. 2022, 65, 47–55. [Google Scholar] [CrossRef]
- Hassell, N.; Tinker, K.A.; Moore, T.; Ottesen, E.A. Temporal and spatial dynamics in microbial community composition within a temperate stream network. Environ. Microbiol. 2018, 20, 3560–3572. [Google Scholar] [CrossRef]
- Liu, J.W.; Meng, Z.; Liu, X.Y.; Zhang, X.H. Microbial assembly, interaction, functioning, activity and diversification: A review derived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef]
- Kalamdhad, A.S.; Kazmi, A.A. Effects of turning frequency on compost stability and some chemical characteristics in a rotary drum composter. Chemosphere 2009, 74, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Cheng, H.; Han, Z.; Wei, Z.; Song, C. Identification of driving factors of lignocellulose degrading enzyme genes in different microbial communities during rice straw composting. Bioresour. Technol. 2023, 381, 129109. [Google Scholar] [CrossRef] [PubMed]
- Eevers, N.; Hawthorne, J.R.; White, J.C.; Vangronsveld, J.; Weyens, N. Exposure of Cucurbita pepo to DDE-contamination alters the endophytic community: A cultivation dependent vs a cultivation independent approach. Environ. Pollut. 2016, 209, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.T.; Yang, X.D.; Zhang, J.J.; Lin, Z.A.; Zhao, B.Q. Microbial community structure and functional metabolic diversity are associated with organic carbon availability in an agricultural soil. J. Integr. Agric. 2015, 14, 2500–2511. [Google Scholar] [CrossRef]
- Whitman, T.; DeCiucies, S.; Hanley, K.; Enders, A.; Woolet, J.; Lehmann, J. Microbial Community Shifts Reflect Losses of Native Soil Carbon with Pyrogenic and Fresh Organic Matter Additions and Are Greatest in Low-Carbon Soils. Appl. Environ. Microbiol. 2021, 87, e02555-20. [Google Scholar] [CrossRef]
- Kingsley, M.K.; Bhat, B.V. Influence of Microbiome in Shaping the Newborn Immune System: An Overview. In Microbiome, Immunity, Digestive Health and Nutrition; Academic Press: Cambridge, MA, USA, 2022; pp. 11–24. [Google Scholar]
Figure 1.
Lignocellulosic degradation dynamics during composting. (a,b) represent the mean cellulose content; (c,d) represent the mean hemicellulose content; and (e,f) represent the mean lignin content.
Figure 1.
Lignocellulosic degradation dynamics during composting. (a,b) represent the mean cellulose content; (c,d) represent the mean hemicellulose content; and (e,f) represent the mean lignin content.
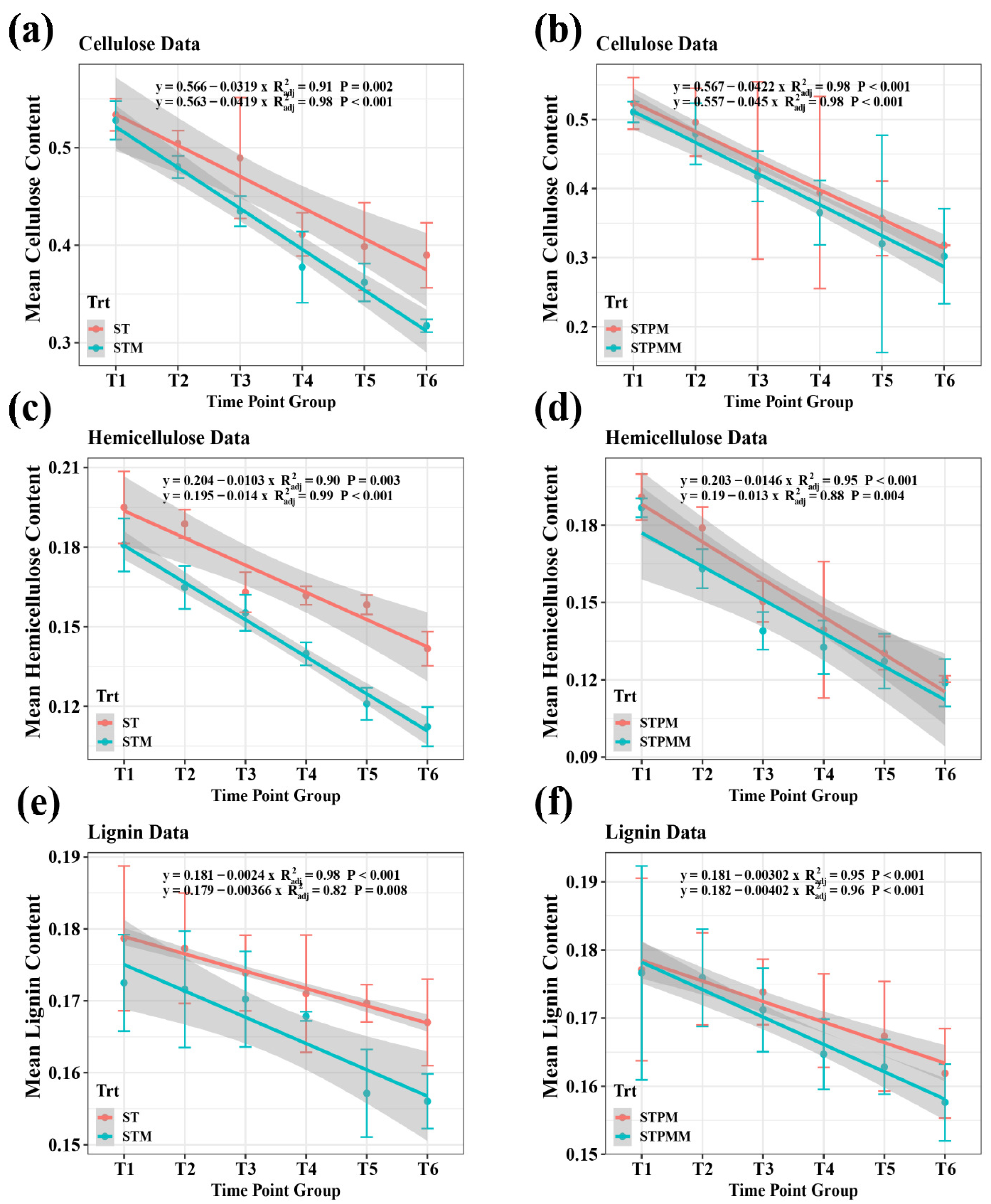
Figure 2.
Fungal community dynamics influenced by Marasmius tricolor 310b. This figure showcases the alpha and beta diversity metrics of the fungal community upon the introduction of Marasmius tricolor 310b. NMDS plots based on the Bray-Curtis distance visualize the clustering distinctions between treatments. The core set of species observed across all treatments and the dominant fungal phyla are also presented. (a) Fungal diversity index; (b) MDS analysis with a Brat-curtis distance matrix of communities; (c) Fungal Venn; (d) Fungal phylum level; (e) Fungal genus level; (f) Fungal species level.
Figure 2.
Fungal community dynamics influenced by Marasmius tricolor 310b. This figure showcases the alpha and beta diversity metrics of the fungal community upon the introduction of Marasmius tricolor 310b. NMDS plots based on the Bray-Curtis distance visualize the clustering distinctions between treatments. The core set of species observed across all treatments and the dominant fungal phyla are also presented. (a) Fungal diversity index; (b) MDS analysis with a Brat-curtis distance matrix of communities; (c) Fungal Venn; (d) Fungal phylum level; (e) Fungal genus level; (f) Fungal species level.
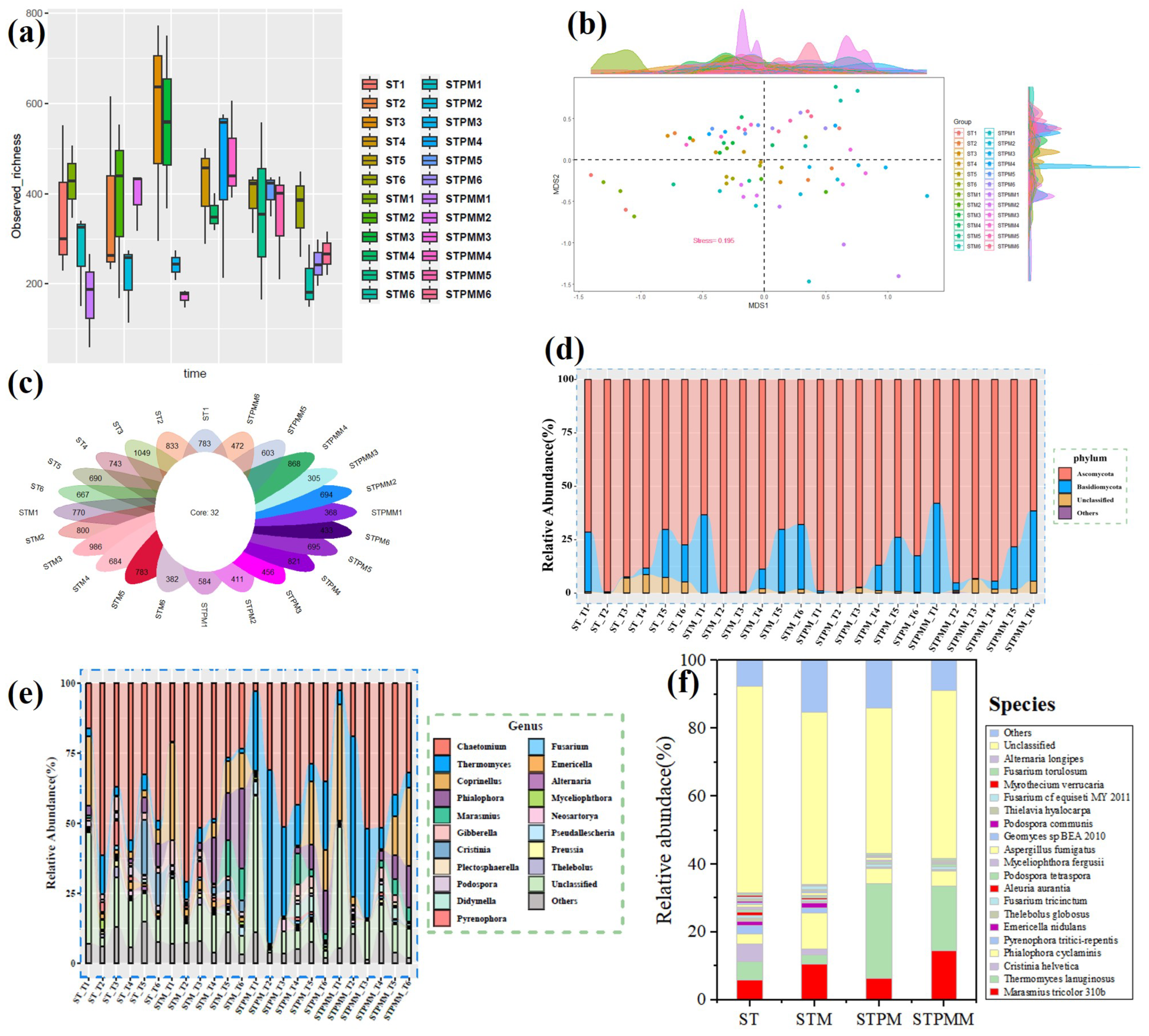
Figure 3.
The top 21 biomarker fungal zOTUs are accurately identified by the random forest model. The biomarkers are ranked in descending order of importance to the accuracy of the model. The inset represents a 10-fold cross-validation error as a function of the number of inputs OTUs used to differentiate all the treatments’ fungal community ranked by variable importance.
Figure 3.
The top 21 biomarker fungal zOTUs are accurately identified by the random forest model. The biomarkers are ranked in descending order of importance to the accuracy of the model. The inset represents a 10-fold cross-validation error as a function of the number of inputs OTUs used to differentiate all the treatments’ fungal community ranked by variable importance.

Figure 4.
The intra-domain network analysis of the fungal community. (a) The fungal network of pure rapeseed straw. (b) The fungal network of rapeseed straw with Marasmius tricolor 310b. (c) The fungal network of pig manure mixed with rapeseed straw. (d) The fungal network of pig manure mixed with rapeseed straw supplemented with Marasmius tricolor 310b. The different colors represent different modules, while modules with five nodes or less are grey. Node size indicates the node degree of node. N, node; L, links.
Figure 4.
The intra-domain network analysis of the fungal community. (a) The fungal network of pure rapeseed straw. (b) The fungal network of rapeseed straw with Marasmius tricolor 310b. (c) The fungal network of pig manure mixed with rapeseed straw. (d) The fungal network of pig manure mixed with rapeseed straw supplemented with Marasmius tricolor 310b. The different colors represent different modules, while modules with five nodes or less are grey. Node size indicates the node degree of node. N, node; L, links.

Figure 5.
The complexity and stability of soil microbial community. (a,b) Cohesion analysis of the microbial communities. (c) Robustness analysis of the microbial communities. The levels of significance are indicated as 0.001 ***. “NS” indicates no significance.
Figure 5.
The complexity and stability of soil microbial community. (a,b) Cohesion analysis of the microbial communities. (c) Robustness analysis of the microbial communities. The levels of significance are indicated as 0.001 ***. “NS” indicates no significance.
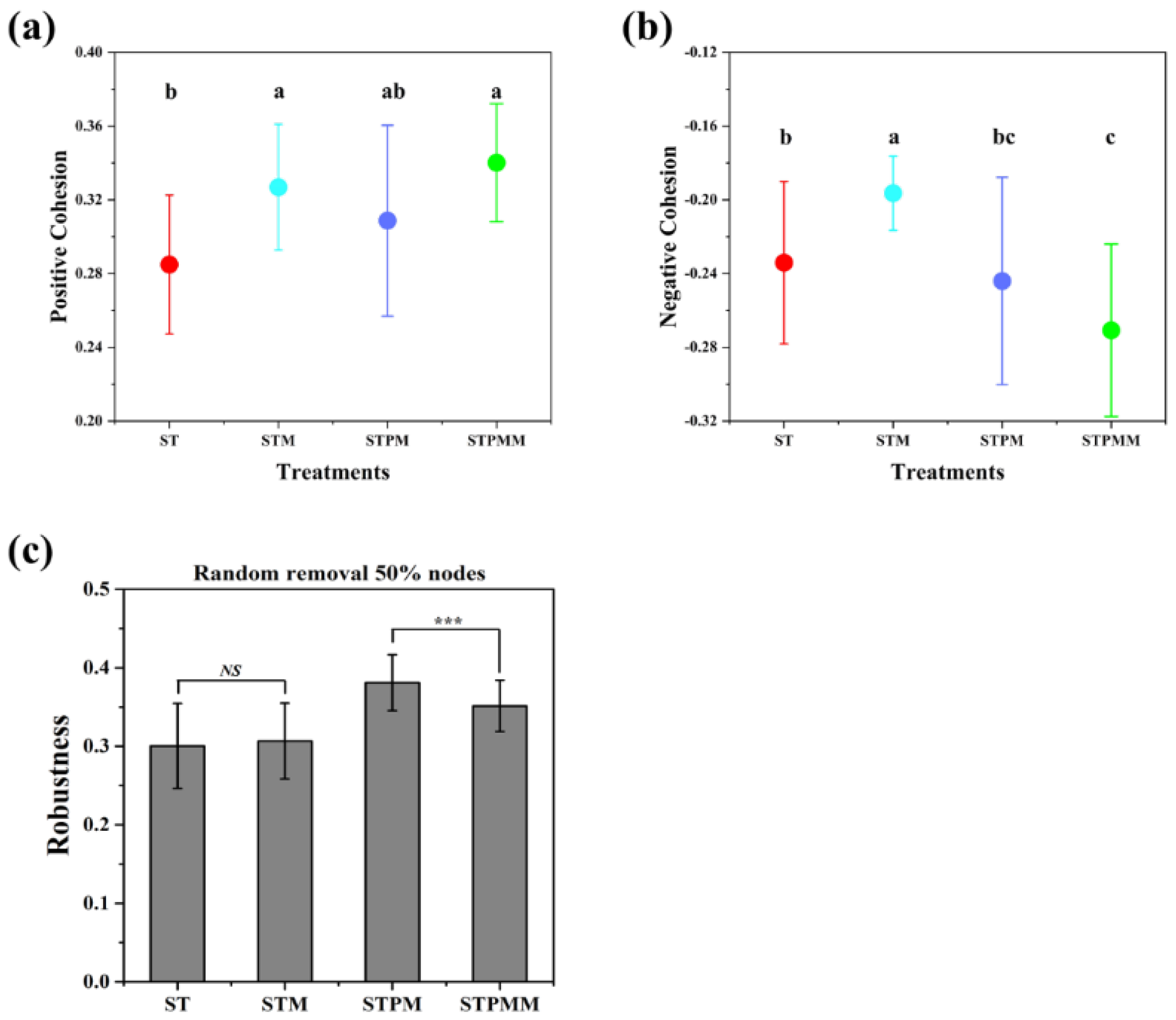
Figure 6.
Temporal stability analysis using the time–decay relationship (TDR) model of fungal community based on taxonomic diversity in rapeseed straw composting conditions. Taxonomic diversity was measured by Sorensen. The TDR values (ρ) are presented as a coefficient in fixed effect ± standard error in random effect. The r2 values were calculated, reflecting variance explained by the whole model. The p value of each TDR was based on a permutation test. The lines show the fixed effects in the model.
Figure 6.
Temporal stability analysis using the time–decay relationship (TDR) model of fungal community based on taxonomic diversity in rapeseed straw composting conditions. Taxonomic diversity was measured by Sorensen. The TDR values (ρ) are presented as a coefficient in fixed effect ± standard error in random effect. The r2 values were calculated, reflecting variance explained by the whole model. The p value of each TDR was based on a permutation test. The lines show the fixed effects in the model.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Physicochemical properties of composting raw materials.
| Raw Material | Pig Manure | Rapeseed Straw |
|---|---|---|
| pH | 7.75 | — |
| Moisture content | 65.59 | 8.86 |
| Organic matter | 72.41 | 67.69 |
| Total N | 1.85 | 0.76 |
| Total P | 2.10 | 0.10 |
| Total K | 1.17 | 1.07 |
| C/N | 22.7:1 | 52.0:1 |
Note: “—” indicates no value.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Z.; Shi, D.; Lu, G. The Impact of Marasmius tricolor 310b on the Degradation of Cellulose in Rapeseed Straw Composting. Agronomy 2023, 13, 3012. https://doi.org/10.3390/agronomy13123012
AMA Style
Wang Z, Shi D, Lu G. The Impact of Marasmius tricolor 310b on the Degradation of Cellulose in Rapeseed Straw Composting. Agronomy. 2023; 13(12):3012. https://doi.org/10.3390/agronomy13123012
Chicago/Turabian StyleWang, Zhihui, Dejun Shi, and Guangxin Lu. 2023. "The Impact of Marasmius tricolor 310b on the Degradation of Cellulose in Rapeseed Straw Composting" Agronomy 13, no. 12: 3012. https://doi.org/10.3390/agronomy13123012
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.