
Inoculation with Plant Growth-Promoting Bacteria Improves the Sustainability of Tropical Pastures with Megathyrsus maximus

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Biological Material
2.2. Plant-Growth-Promotion Evaluation under Greenhouse Conditions
2.2.1. Treatments and Growth Conditions
2.2.2. Evaluation of Root Growth Traits
2.2.3. Biomass Production and Accumulation of Nutrients in Shoots
2.2.4. Statistical Analysis
2.3. Evaluation of Seed and Root Colonization with Scanning Electron Microscopy
2.3.1. Treatments, Plant Growth Conditions
2.3.2. Evaluation of Seed and Root Colonization
3. Results
3.1. Plant Growth Promotion
3.1.1. Root Traits
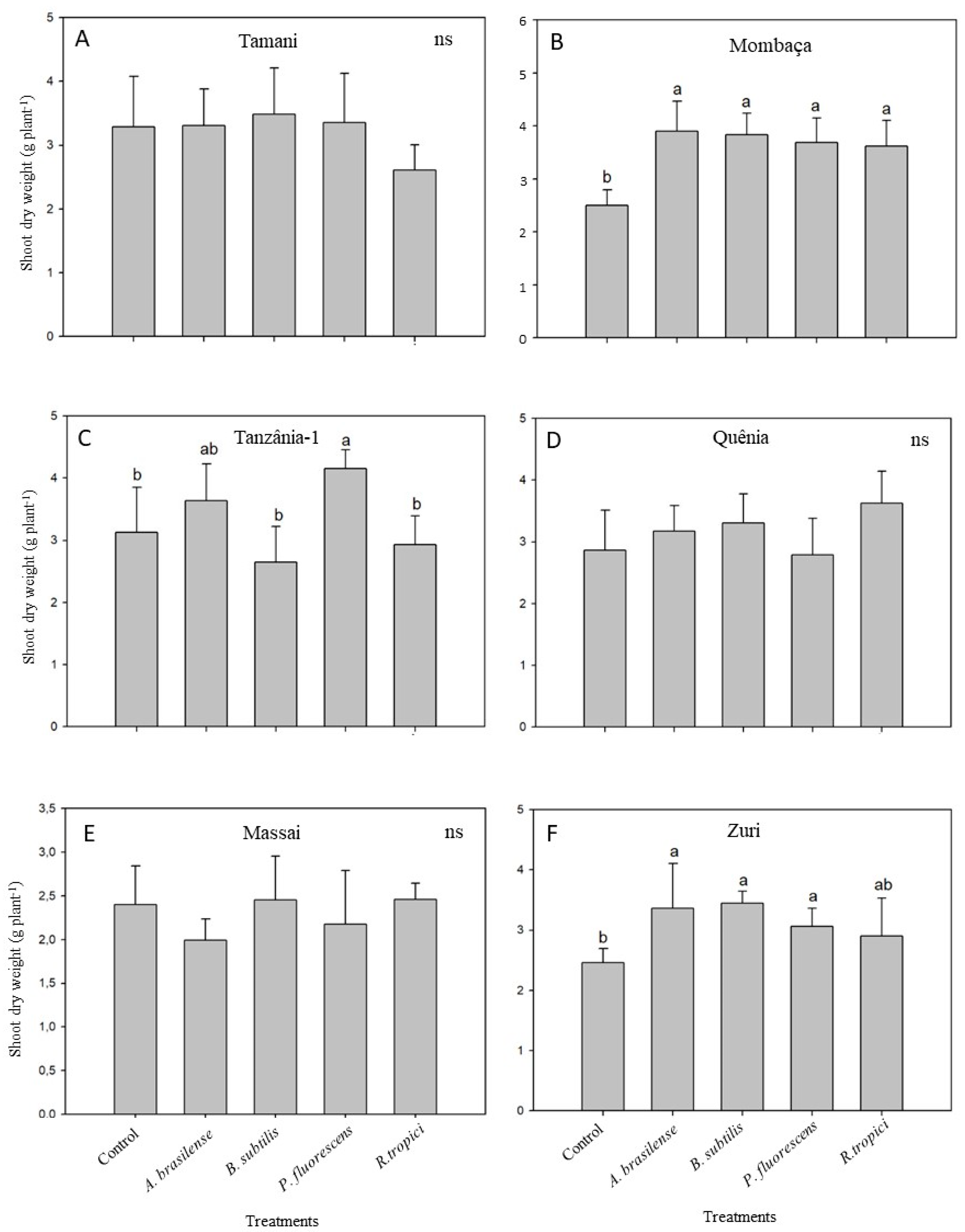
3.1.2. Shoot Biomass Production and Nutrient Accumulation
3.2. Seed and Root Colonization
4. Discussion
5. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MAPA (Ministério da Agricultura, Pecuária e Abastecimento). Agropecuária Brasileira em Números. Secretaria de Política Agrícola. Available online: https://www.gov.br/agricultura/ptbr/assuntos/politica-agricola/todas-publicacoesde-politicaagricola/agropecuaria-brasileira-em-numeros (accessed on 15 November 2022).
- UNIPASTO (Associação para o Fomento à Pesquisa de Melhoramento de Forrageiras). 2022. Available online: https://www.unipasto.com.br (accessed on 14 April 2022).
- Jank, L. A história do Panicum maximum no Brasil. Ver. JC Maschietto. 2003, 1, 14. [Google Scholar]
- Bahlo, C.; Dahlhaus, P.; Thompson, H.; Trotter, M. The role of interoperable data standards in precision livestock farming in extensive livestock systems: A review. Comput. Electron. Agric. 2019, 156, 459–466. [Google Scholar] [CrossRef]
- Terra, A.B.; Florentino, L.A.; Rezende, A.D.; Silva, N.C. Leguminosas forrageiras na recuperação de pastagens no Brasil. Rev. Cienc. Agrícola. 2019, 42, 305–313. [Google Scholar] [CrossRef]
- Macedo, M.C.M.; Araújo, A.R. Sistemas de produção em integração: Alternativa para recuperação de pastagens degradadas. In ILPF: Inovação com Integração de Lavoura, Pecuária e Floresta, 1st ed.; Bungenstab, D.J., de Almeida, R.G., Laura, V.A., Balbino, L.C., Ferreira, A.D., Eds.; Embrapa: Brasília, Brazil, 2019; Volume 1, pp. 296–317. [Google Scholar]
- Peron, A.J.; Evangelista, A.R. Degradação de pastagens em regiões de cerrado. Ciênc. Agrotec. 2004, 28, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Macedo, M.C.M.; Zimmer, A.H.; Kichel, N.A.; Almeida, R.G.; de Araujo, A.R. Degradação de pastagens, alternativas de recuperação e renovação, e formas de mitigação. In Encontro de Adubação de Pastagens da Scot Consultoria-Tec-Fértil; Scot Consultoria: Piracicaba, Brazil, 2013; pp. 158–181. [Google Scholar]
- Hungria, M.; Nogueira, M.A. Nitrogen fixation. In Marschner’s Mineral Nutrition of Plants, 4th ed.; Rengel, Z., Cakmak, I., White, P.J., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2023; pp. 615–650. ISBN 978-0-12-819773-8. [Google Scholar]
- Sharma, S.B.; Sayyed, R.Z.; Trived, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Abramovay, R. Alimentos versus população: Está ressurgindo o fantasma malthusiano? Ciênc. Cult. 2010, 62, 38–42. [Google Scholar]
- Hungria, M.; Campo, R.J.; Souza, E.M.; Pedrosa, F.O. Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil. 2010, 331, 413–425. [Google Scholar] [CrossRef]
- Hungria, M.; Nogueira, M.A.; Araujo, R.S. Inoculation of Brachiaria spp. with the plant growth-promoting bacterium Azospirillum brasilense: An environment-friendly component in the reclamation of degraded pastures in the tropics. Agric. Ecosyst. Environ. 2016, 221, 125–131. [Google Scholar] [CrossRef]
- Hungria, M.; Barbosa, J.Z.; Rondina, A.B.L.; Nogueira, M.A. Improving maize sustainability with partial replacement of N-fertilizers by inoculation with Azospirillum brasilense. Agron. J. 2022, 114, 2969–2980. [Google Scholar] [CrossRef]
- Videira, S.S.; Oliveira, D.M.; Morais, R.F.; Borges, W.L.; Baldani, V.L.D.; Baldani, J.I. Genetic diversity and plant growth promoting traits of diazotrophic bacteria isolated from two Pennisetum purpureum Schum genotypes grown in the field. Plant Soil. 2012, 356, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: Reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 9, 205. [Google Scholar] [CrossRef]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Outstanding impact of Azospirillum brasilense strains Ab-V5 and Ab-V6 on the Brazilian agriculture: Lessons that farmers are receptive to adopt new microbial inoculants. Rev. Bras. Cienc. Solo. 2021, 45, 1–31. [Google Scholar] [CrossRef]
- Guimarães, G.S.; Rondina, A.B.L.; Santos, M.S.; Nogueira, M.A.; Hungria, M. Pointing out opportunities to increase grassland pastures productivity via microbial inoculants: Attending the society’s demands for meat production with sustainability. Agronomy 2022, 12, 1748. [Google Scholar] [CrossRef]
- Ahmed, B.; Shahid, M.; Syed, A.; Rajput, V.D.; Elgorban, A.M.; Minkina, T.; Bahkali, A.H.; Lee, J. Drought tolerant Enterobacter sp. Leclercia adecarboxylata secretes indole-3-acetic scid and other biomolecules and enhances the biological sttributes of Vigna radiata (L.) R. Wilczek in water deficit conditions. Biology 2021, 10, 1149. [Google Scholar] [CrossRef] [PubMed]
- AL-Shwaiman, H.A.; Shahid, M.; Elgorban, A.M.; Siddique, K.H.M.; Syed, A. Beijerinckia fluminensis BFC-33, a novel multi-stress-tolerant soil bacterium: Deciphering the stress amelioration, phytopathogenic inhibition and growth promotion in Triticum aestivum (L.). Chemosphere 2022, 295, 133843. [Google Scholar] [CrossRef] [PubMed]
- Mariano, R.L.R.; Silveira, E.B.; Assis, S.M.P.; Gomes, A.M.A.; Nascimento, A.R.P.; Donato, V.M.T.S. Importância de bactérias promotoras de crescimento e de biocontrole de doenças de plantas para uma agricultura sustentável. An. Acad. Pernamb. Cienc. Agron. 2013, 1, 89–111. [Google Scholar]
- Fukami, J.; Abrantes, J.L.F.; Del Cerro, P.; Nogueira, M.A.; Ollero, F.J.; Megías, M.; Hungria, M. Revealing different strategies of quorum sensing in Azospirillum brasilense strains Ab-V5 and Ab-V6. Arch. Microbiol. 2018, 200, 47–56. [Google Scholar] [CrossRef]
- Msimbira, L.A.; Smith, D.L. The roles of plant growth promoting microbes in enhancing plant tolerance to acidity and alkalinity stresses. Front. Sustain. Food Syst. 2020, 4, 106. [Google Scholar] [CrossRef]
- Lopes, M.J.S.; Dias-Filho, M.B.; Castro, T.H.R.; Gurgel, E.S.C.; da Silva, G.B. Efficiency of biostimulants for alleviating shade effects on forage grass. J. Agric. Stud. 2021, 9, 14–30. [Google Scholar] [CrossRef]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express 2018, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Araujo, F.F.; Hungria, M. Nodulação e rendimento de soja co-infectada com Bacillus subtilis e Bradyrhizobium japonicum/B. elkanii. Pesqui. Agropecuária Bras. 1999, 34, 1633–1643. [Google Scholar] [CrossRef]
- Ormeño-Orrillo, E.; Hungria, M.; Martínez-Romero, E. Dinitrogen-fixing prokaryotes. In The Prokaryotes—Prokaryotic Physiology and Biochemistry, 4th ed.; Rosemberg, E., De Long, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 427–451. [Google Scholar] [CrossRef]
- Sandini, I.E.; Pacentchuk, F.; Hungria, M.; Nogueira, M.A.; Cruz, S.P.; Nakatani, A.S.; Araujo, R.S. Seed inoculation with Pseudomonas fluorescens promotes growth, yield and reduces nitrogen application in maize. Int. J. Agric. Biol. 2019, 22, 1369–1375. [Google Scholar] [CrossRef]
- Hungria, M.; Rondina, A.B.L.; Nunes, A.L.P.; Araujo, R.S.; Nogueira, M.A. Seed and leaf-spray inoculation of PGPR in Brachiaria (Urochloa spp.) as an economic and environmental opportunity to improve plant growth, forage yield and nutrient status. Plant Soil. 2021, 463, 171–186. [Google Scholar] [CrossRef]
- Yates, R.J.; Howieson, J.G.; Hungria, M.; Bala, A.; O’Hara, G.W.; Terpolilli, J. Authentication of rhizobia and assessment of the legume symbiosis in controlled plant growth systems. In Working with Rhizobia, 1st ed.; Howieson, J.G., Dilworth, M.J., Eds.; Australian Center for Interantional Agricultural Research (ACIAR): Canberra, Australia, 2016; pp. 73–108. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agr. Exp. Sta. Cir. 1950, 347, 32. [Google Scholar]
- Tennant, D. A test of modified line intersect method of estimating root length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- Rondina, A.B.L.; Sanzovo, A.W.S.; Guimarães, G.S.; Wendling, J.R.; Nogueira, M.A.; Hungria, M. Changes in root morphological traits in soybean co-inoculated with Bradyrhizobium spp. and Azospirillum brasilense or treated with A. brasilense exudates. Biol. Fertil. Soils 2020, 56, 537–549. [Google Scholar] [CrossRef]
- Zangaro, W.; Nishidate, F.R.; Camargo, F.R.S.; Romagnoli, G.G.; Vandresen, J. Relationships among arbuscular mycorrhizas, root morphology and seedling growth of tropical native woody species in southern Brazil. J. Trop. Ecol. 2005, 21, 529–540. [Google Scholar] [CrossRef]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Princípios, métodos e técnicas de avaliação do estado nutricional. In Avaliação do Estado nutricional das Plantas: Princípios e Aplicações, 2nd ed.; Malavolta, E., Vitti., G.C., Oliveira., S.A., Eds.; Associação Brasileira para a Pesquisa da Potassa e do Fosfato: Piracicaba, Brazil, 1997; pp. 115–230. [Google Scholar]
- Barbosa, J.C.; Maldonado, J.W. AgroEstat: Sistema para análises estatísticas de ensaios agronômicos; FCAV/UNESP: Jaboticabal, Brazil, 2015; 396p. [Google Scholar]
- Freschet, G.T.; Roumet, C.; Comas, L.H.; Weemstra, M.; Bengough, A.G.; Rewald, B.; Bardgett, R.D.; de Deyn, G.B.; Johnson, D.; Klimešová, J.; et al. Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs. New Phytologist. 2021, 232, 1123–1158. [Google Scholar] [CrossRef]
- Přikryl, Z.; Vančura, V.; Wurst, M. Auxin formation by rhizosphere bacteria as a factor of root growth. Biol. Plant. 1985, 27, 159–163. [Google Scholar] [CrossRef]
- Lambrecht, M.; Okon, Y.; Broek, A.V.; Vanderleyden, J. Indole-3-acetic acid: A reciprocal signalling molecule in bacteria–plant interactions. Trends Microbiol. 2000, 8, 298–300. [Google Scholar] [CrossRef]
- Tahir, H.A.S.; Gun, Q.; Wu, H.; Raza, W.; Hanif, A.; Wu, L.; Colman, M.V.C.; Gao, X. Plant growth promotion by volatile organic compounds produced by Bacillus subtilis SYST2. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tullio, L.D.; Nakatani, A.S.; Gomes, D.F.; Ollero, F.J.; Megías, M.; Hungria, M. Revealing the roles of y4wF and tidC genes in Rhizobium tropici CIAT 899: Biosynthesis of indolic compounds and impact on symbiotic properties. Arch. Microbiol. 2019, 201, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Costacurta, A.; Vanderleyden, J. Synthesis of phytohormones by plant-associated bacteria. Crit. Rev. Microbiol. 1995, 21, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.F.D.; Cecato, U.; Biserra, T.T.; Mamédio, D.; Galbeiro, S. Azospirillum spp. in grasses and forages. Review. Rev. Mex. Cienc. Pecu. 2020, 11, 223–240. [Google Scholar] [CrossRef]
- Barbieri, P.; Galli, E. Effect on wheat root development of inoculation with an Azospirillum brasilense mutant with altered indole-3-acetic acid production. Res. Microbiol. 1993, 144, 69–75. [Google Scholar] [CrossRef]
- Ortíz-Castro, R.; Valencia-Cantero, E.; López-Bucio, J. Plant growth promotion by Bacillus megaterium involves cytokinin signaling. Plant Signal. Behav. 2008, 3, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creus, C.M.; Graziano, M.; Casanovas, E.M.; Pereyra, M.A.; Simontacchi, M.; Puntarulo, S.; Barrasi, C.A.; Lamattina, L. Nitric oxide is involved in the Azospirillum brasilense-induced lateral root formation in tomato. Planta 2005, 221, 297–303. [Google Scholar] [CrossRef]
- Ortiz-Castro, R.; Campos-García, J.; López-Bucio, J. Pseudomonas putida and Pseudomonas fluorescens influence Arabidopsis root system architecture through an auxin response mediated by bioactive cyclodipeptides. J. Plant Growth Regul. 2020, 39, 254–265. [Google Scholar] [CrossRef]
- El-Khawas, H.; Adachi, K. Identification and quantification of auxins in culture media of Azospirillum and Klebsiella and their effect on rice roots. Biol. Fertil. Soils. 1999, 28, 377–381. [Google Scholar] [CrossRef]
- Dobbelaere, S.; Croonenborghs, A.; Thys, A.; Broek, A.V.; Vanderleyden, J. Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil 1999, 212, 153–162. [Google Scholar] [CrossRef]
- Garcia-Montiel, D.C.; Neill, C.; Melillo, J.; Thomas, S.; Steudler, P.A.; Cerri, C.C. Soil phosphorus transformations following forest clearing for pasture in the brazilian Amazon. Soil Sci. Soc. Am. J. 2000, 64, 1792–1804. [Google Scholar] [CrossRef]
- Dias-Filho, M.B. Diagnóstico das Pastagens no Brasil; Embrapa Amazônia Oriental: Belém, Brazil, 2014; 38p. [Google Scholar]
- Criollo, P.J.; Obando, M.; Sánchez, M.L.; Bonilla, R. Efecto de bacterias promotoras de crecimiento vegetal (PGPR) asociadas a Pennisetum clandestinum en el altiplano cundiboyacense. Corpoica Cienc. Tecnol. Agropec. 2012, 13, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Telhado, S.F.P.; Capdeville, G. Land-Saving Technologies; Embrapa: Brasília, Brazil, 2021; 161p. [Google Scholar]
- Heinrichs, R.; Meirelles, G.C.; Santos, L.P.M.; Lira, M.C.S.; Lapaz, A.M.; Nogueira, M.A.; Bonini, C.S.B.; Soares Filho, C.V.; Moreira, A. Azospirillum inoculation of ‘Marandu’ palisade grass seeds: Effects on forage production and nutritional status. Semina. Cienc. Agrar. 2020, 41, 465–478. [Google Scholar] [CrossRef]
- Chu, T.N.; Bui, L.V.; Hoang, M.T.T. Pseudomonas PS01 isolated from maize rhizosphere alters root system architecture and promotes plant growth. Microorganisms 2020, 8, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itzigsohn, R.; Burdman, S.; Okon, Y.; Zaady, E.; Yonatan, R.; Perevolotsky, A. Plant-growth promotion in natural pastures by inoculation with Azospirillum brasilense under suboptimal growth conditions. Arid Soil Res. Rehabil. 2000, 13, 151–158. [Google Scholar] [CrossRef]
- Leite, R.D.C.; dos Santos, J.G.; Silva, E.L.; Alves, C.R.; Hungria, M.; Leite, R.D.C.; dos Santos, A.C. Productivity increase, reduction of nitrogen fertiliser use and drought-stress mitigation by inoculation of Marandu grass (Urochloa brizantha) with Azospirillum brasilense. Crop Past. Scienc. 2018, 70, 61–67. [Google Scholar] [CrossRef]
- Braga, G.J.; Ramos, A.K.B.; Carvalho, M.A.; Fonseca, C.E.L.; Fernandes, F.D.; Malaquias, J.V.; Santos, M.F.; Jank, L. Produção de Forragem e Valor Nutritivo de Híbridos de Panicum Maximum Jacq. em Resposta à Adubação; Brasilia, D.F., Ed.; Embrapa Cerrados: Brasília, Brasil, 2019; 19p. [Google Scholar]
- PASTOCERTO. Comparar Cultivares. 2019. Available online: https://www.pastocerto.com (accessed on 15 November 2022).
- Rothballer, M.; Schmid, M.; Hartmann, A. In situ localization and PGPR-effect of Azospirillum brasilense strains colonizing roots of different wheat varieties. Symbiosis 2003, 34, 261–279. [Google Scholar]
- Bhattacharjee, R.B.; Jourand, P.; Chaintreuil, C.; Dreyfus, B.; Singh, A.; Mukhopadhyay, S.N. Indole acetic acid and ACC deaminase-producing Rhizobium leguminosarum bv. trifolii SN10 promote rice growth, and in the process undergo colonization and chemotaxis. Biol. Fertil. Soils 2012, 48, 173–182. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Treatments | Dry Weight (RDW) (g) | Volume (RV) (cm3) | Total Length (TRL) (m) | Specific Length (SRL) (m g−1) | Area (RA) (cm2) | Mean Diameter (RMD) (µm) | Tissue Density (RTD) (g cm−3) | Hair Incidence (RHI) (%) | Hair Length (RHL) (µm) | Total Number of Root Branches (TNB) |
|---|---|---|---|---|---|---|---|---|---|---|
| Tamani | ||||||||||
| Non-inoculated | 0.97 ± 0.12b 1 | 7.75 ± 0.84b | 94 ± 16.6bc | 98 ± 14.7 n.s. | 94 ±126bc | 333 ± 23.3 n.s. | 0.13 ± 0.01 n.s. | 54 ± 6.89 n.s. | 204 ± 17.4 n.s. | 45,762 ± 9587c |
| A. brasilense | 1.17 ± 0.07a | 9.46 ± 0.30a | 115 ± 7.05ab | 99 ± 7.12 | 1167 ± 45.2a | 325 ± 9.67 | 0.12 ± 0.01 | 40 ± 5.82 | 199 ± 22.3 | 53,689 ± 3874bc |
| B. subtilis | 0.93 ± 0.05b | 8.10 ± 0.23ab | 122 ± 6.92a | 133 ± 7.66 | 1117 ± 40.8ab | 290 ± 7.81 | 0.11 ± 0 | 31 ± 6.15 | 189 ± 7.24 | 12,608 ± 16,874a |
| P. fluorescens | 0.79 ± 0.03bc | 7.56 ± 0.39b | 85 ± 6.33bc | 109 ± 6.06 | 902 ± 55.6c | 336 ± 5.69 | 0.10 ± 0 | 37 ± 7.16 | 182 ± 10.9 | 62,497 ± 6155bc |
| R. tropici | 0.68 ± 0.02c | 6.18 ± 0.23c | 72 ± 6.23c | 106 ± 9.1 | 748 ± 34.8c | 332 ± 17.3 | 0.11 ± 0 | 31 ± 6.78 | 186 ± 16.5 | 87,428 ± 19,830ab |
| p value | <0.001 | <0.001 | 0.007 | 0.117 | 0.002 | 0.167 | 0.122 | 0.136 | 0.857 | 0.002 |
| Mombaça | ||||||||||
| Non-inoculated | 0.77 ± 0.08b | 6.58 ± 0.64 | 82 ± 7.65 n.s. | 109 ± 8.11 n.s. | 881 ± 49.9 n.s. | 315 ± 9.26 n.s. | 0.12 ± 0.008b | 34 ± 6.28 n.s. | 205 ± 3.20a | 45,135 ± 5986b |
| A. brasilense | 0.94 ± 0.08ab | 6.48 ± 0.6 | 101 ± 9.23 | 101 ± 1.62 | 1006 ± 42.5 | 294 ± 5.09 | 0.15 ± 0.006a | 51 ± 6.84 | 188 ± 11.3ab | 74,025 ± 3922a |
| B. subtilis | 1.02 ± 0.13ab | 7.63 ± 0.8 | 114 ± 19.2 | 99 ± 8.45 | 928 ± 76.6 | 317 ± 12.3 | 0.13 ± 0.002ab | 32 ± 3.08 | 166 ± 5.77b | 85,098 ± 7690a |
| P. fluorescens | 1.15 ± 0.06a | 8.63 ± 0.69 | 111 ± 7.41 | 106 ± 6.50 | 1093 ± 53.5 | 315 ± 17.7 | 0.14 ± 0.008ab | 48 ± 3.79 | 199 ± 7.07a | 76,837 ± 9638a |
| R. tropici | 1.11 ± 0.09a | 7.98 ± 0.65 | 108 ± 13.5 | 103 ± 7.08 | 1035 ± 93.3 | 311 ± 15.5 | 0.14 ± 0.005ab | 32 ± 4.97 | 182 ± 10.6ab | 69,579 ± 6913a |
| p value | 0.001 | 0.152 | 0.36 | 0.42 | 0.192 | 0.705 | 0.04 | 0.077 | 0.025 | 0.004 |
| Tanzânia-1 | ||||||||||
| Non-inoculated | 0.76 ± 0.08c | 6.99 ± 0.66b | 84 ± 5.19c | 127 ± 16,9 n.s. | 856 ± 3.5c | 324 ± 15.9 n.s. | 0.12 ± 0a n.s. | 33 ± 8.46b | 196 ± 15.9ab | 39,241 ± 13,906b |
| A. brasilense | 1.07 ± 0.11ab | 10.61 ± 1.11a | 135 ± 17.5ab | 117 ± 8.22 | 1342 ± 152ab | 318 ± 1.6 | 0.11 ± 0.01ab | 55 ± 7.70a | 221 ± 14.2a | 82,433 ± 23,123a |
| B. subtilis | 1.00 ± 0.07bc | 9.92 ± 0.53a | 139 ± 10.5ab | 131 ± 7.12 | 1318 ± 80.3ab | 302 ± 7.03 | 0.10 ± 0.01b | 36 ± 6.41b | 161 ± 5.48b | 117,811 ± 5857a |
| P. fluorescens | 1.43 ± 0.16a | 11.46 ± 1.12a | 159 ± 11.1a | 103 ± 6.28 | 150 ± 119a | 302 ± 11.2 | 0.12 ± 0a | 37 ± 5.20b | 181 ± 16.9b | 10,348 ± 3523a |
| R. tropici | 1.06 ± 0.11bc | 9.01 ± 0.73ab | 108 ± 8.9bc | 109 ± 5.78 | 1105 ± 89.6bc | 326 ± 4.20 | 0.12 ± 0a | 27 ± 6.52b | 178 ± 1.95b | 105,178 ± 8801a |
| p value | 0.011 | 0.012 | <0.001 | 0.07 | <0.001 | 0.32 | 0.001 | 0.012 | 0.032 | 0.002 |
| Quênia | ||||||||||
| Non-inoculated | 0.75 ± 0.03b | 5.95 ± 0.17b | 86 ± 2.04bc | 116 ± 6.36ab | 781 ± 32.7b | 309 ± 7.81 n.s. | 0.13 ± 0.005 n.s. | 38 ± 3.11 n.s. | 175 ± 7.57 n.s. | 38,981 ± 1091b |
| A. brasilense | 1.11 ± 0.10a | 8.42 ± 0.63a | 99 ± 4.08a | 97 ± 6.54b | 1084 ± 79.3a | 296 ± 4.14 | 0.13 ± 0.008 | 47 ± 8.18 | 192 ± 17.7 | 52,194 ± 7614b |
| B. subtilis | 0.78 ± 0.05b | 6.58 ± 0.31b | 81 ± 5.43c | 104 ± 4.18b | 819 ± 43.4b | 317 ± 8.37 | 0.12 ± 0.004 | 40 ± 3.84 | 174 ± 11.7 | 75,650 ± 8898a |
| P. fluorescens | 0.81 ± 0.09b | 6.16 ± 0.54b | 90 ± 2.37abc | 127 ± 8.42a | 850 ± 63.6b | 299 ± 9.16 | 0.12 ± 0.012 | 44 ± 6.31 | 186 ± 3.64 | 41,344 ± 6967b |
| R. tropici | 0.95 ± 0.06ab | 6.60 ± 0.06b | 96 ± 1.86ab | 102 ± 3.4b | 877 ± 35.7b | 302 ± 3.31 | 0.13 ± 0.004 | 48 ± 7.45 | 184 ± 11.3 | 47,528 ± 1528b |
| p value | 0.012 | 0.003 | 0.006 | 0.003 | 0.006 | 0.228 | 0.459 | 0.72 | 0.77 | 0.003 |
| Massai | ||||||||||
| Non-inoculated | 0.86 ± 0.06 n.s. | 6.56 ± 0.06bc | 92 ± 4.35b | 115 ± 8.84ab | 889 ± 61.1bc | 291 ± 14.3bc | 0.12 ± 0.003 n.s. | 26 ± 1.31c | 204 ± 13.9 n.s. | 65,717 ± 7486 n.s. |
| A. brasilense | 0.80 ± 0.08 | 5.91 ± 0.48c | 80 ± 2.13c | 89 ± 3.56c | 734 ± 25.5d | 330 ± 8.71a | 0.13 ± 0.003 | 54 ± 5.26a | 218 ± 16.6 | 55,375 ± 4692 |
| B. subtilis | 0.71 ± 0.06 | 5.54 ± 0.38c | 90 ± 9.15b | 137 ± 12.7a | 815 ± 58.6cd | 273 ± 13.7c | 0.13 ± 0.002 | 28 ± 3.97c | 207 ± 13.5 | 60,767 ± 4086 |
| P. fluorescens | 0.99 ± 0.08 | 8.25 ± 0.36a | 126 ± 7.36a | 122 ± 6.05ab | 1112 ± 56.4a | 297 ± 4.70abc | 0.12 ± 0.006 | 42 ± 0.76ab | 203 ± 20.6 | 65,631 ± 1807 |
| R. tropici | 0.93 ± 0.06 | 7.80 ± 0.56ab | 97 ± 2.19b | 106 ± 5.62bc | 972 ± 41.9ab | 319 ± 10.5ab | 0.14 ± 0.002 | 31 ± 4.20bc | 204 ± 6.86 | 64,013 ± 4834 |
| p value | 0.069 | 0.004 | <0.001 | 0.006 | <0.001 | 0.010 | 0.11 | <0.001 | 0.956 | 0.534 |
| Zuri | ||||||||||
| Non-inoculated | 0.98 ± 0.24 n.s. | 6.16 ± 0.84b | 76 ± 6.75b | 83 ± 14.1 n.s. | 786 ± 41.1b | 305 ± 5.99 n.s. | 0.112 ± 0b | 55 ± 3.29b | 201 ± 3.29 n.s. | 47,365 ± 6011d |
| A. brasilense | 1.29 ± 0.11 | 10.94 ± 1.29a | 130 ± 10a | 102 ± 7.98 | 1330 ± 116.9a | 326 ± 16.6 | 0.120 ± 0ab | 73 ± 1.59a | 226 ± 6.62 | 68,403 ± 7693bc |
| B. subtilis | 1.40 ± 0.17 | 9.06 ± 0.63a | 123 ± 10.5a | 92 ± 10.4 | 1181 ± 80.4a | 307 ± 10.3 | 0.128 ± 0a | 47 ± 5.76b | 195 ± 4.43 | 10,3847 ± 8402a |
| P. fluorescens | 1.20 ± 0.07 | 10.14 ± 0.2a | 120 ± 9a | 100 ± 7.74 | 1235 ± 56.7a | 330 ± 11.4 | 0.125 ± 0a | 55 ± 3.32b | 195 ± 12.3 | 62,285 ± 1919cd |
| R. tropici | 1.13 ± 0.10 | 8.88 ± 0.63a | 122 ± 14.8a | 102 ± 2.51 | 1232 ± 77a | 300 ± 5.6 | 0.131 ± 0a | 53 ± 9.05b | 197 ± 11.2 | 88,855 ± 10,595ab |
| p value | 0.267 | 0.002 | 0.005 | 0.416 | <0.001 | 0.235 | 0.006 | 0.041 | 0.069 | <0.001 |
| CV (%) | 16.58 | 14.75 | 14.06 | 12.32 | 11.63 | 4.34 | 4.29 | 9.05 | 5.65 | 17.97 |
| Treatments | N | P | K | Ca | Mg | S | B | Cu | Fe | Mn | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg) | (mg) | (mg) | (mg) | (mg) | (mg) | (µg) | (µg) | (µg) | (µg) | (µg) | |
| Tamani | |||||||||||
| Non-inoculated | 24.68 ± 3.46n.s.,1 | 11.59 ± 1.4 n.s. | 60.26 ± 9.72 n.s. | 7.01 ± 0.90 n.s. | 6.46 ± 0.81 n.s. | 3.25 ± 0.35 n.s. | 51.5 ± 4.52 n.s. | 7.65 ± 0.70b | 190 ± 26.0 n.s. | 1142 ± 202 n.s. | 154.00 ± 4.0 n.s. |
| A. brasilense | 24.75 ± 2.59 | 10.52 ± 1.71 | 46.37 ± 7.28 | 5.92 ± 0.49 | 4.68 ± 0.39 | 3.12 ± 0.26 | 50.6 ± 2.58 | 18.48 ± 2.55a | 173 ± 18.7 | 1030 ± 31.2 | 146.93 ± 19.5 |
| B. subtilis | 21.94 ± 1.77 | 12.03 ± 1.48 | 66.30 ± 7.07 | 6.99 ± 0.82 | 5.42 ± 0.66 | 3.37 ± 0.31 | 65.2 ± 11.8 | 20.07 ± 2.12a | 173 ± 18.0 | 904 ± 83.2 | 113.85 ± 21.8 |
| P. fluorescens | 25.77 ± 6.19 | 11.36 ± 2.15 | 51.02 ± 8.32 | 6.41 ± 0.74 | 4.99 ± 0.63 | 3.65 ± 1.02 | 64.1 ± 11.9 | 17.79 ± 2.95a | 209 ± 64.6 | 686 ± 134.9 | 117.80 ± 9.13 |
| R. tropici | 18.87 ± 2 | 8.26 ± 0.84 | 45.80 ± 3.88 | 5.53 ± 0.35 | 4.50 ± 0.35 | 2.65 ± 0.20 | 52.3 ± 4.49 | 15.02 ± 0.47a | 150 ± 46 | 711 ± 88.6 | 23.31 ± 13.7 |
| p value | 0.67 | 0.22 | 0.29 | 0.56 | 0.24 | 0.79 | 0.91 | <0.001 | 0.87 | 0.06 | 0.84 |
| Mombaça | |||||||||||
| Non-inoculated | 18.88 ± 1.06c | 7.82 ± 0.39b | 44.54 ± 4.64 n.s. | 3.93 ± 0.18c | 3.45 ± 0.17b | 2.82 ± 0.17c | 41.2 ± 1.76c | 9.81 ± 0.98 n.s. | 98 ± 3.66b | 620 ± 28.5b | 10.34 ± 0.60c |
| A. brasilense | 24.28 ± 1.75ab | 11.11 ± 0.44a | 48.48 ± 2.92 | 4.70 ± 0.32bc | 5.37 ± 0.51a | 3.70 ± 0.06ab | 94.0 ± 7.52a | 9.24 ± 0.65 | 174 ± 12.1a | 941 ± 70.5a | 84.78 ± 5.13a |
| B. subtilis | 26.97 ± 1.52a | 11.71 ± 0.53a | 56.15 ± 1.91 | 6.68 ± 0.44a | 5.92 ± 0.23a | 3.52 ± 0.16b | 68.4 ± 2.66b | 10.77 ± 1.45 | 220 ± 19.3a | 987 ± 36.6a | 67.72 ± 2.79b |
| P. fluorescens | 21.46 ± 0.88bc | 12.07 ± 0.37a | 55.85 ± 6.77 | 4.99 ± 0.377b | 5.65 ± 0.57a | 4.02 ± 0.02a | 89.4 ± 5.45a | 11.20 ± 1.39 | 189 ± 20.9a | 981 ± 50a | 77.27 ± 5.48ab |
| R. tropici | 23.72 ± 1.52ab | 11.01 ± 0.58a | 52.59 ± 3.17 | 5.17 ± 0.06b | 5.36 ± 0.07a | 3.44 ± 0.2b | 74.1 ± 7.02b | 11.64 ± 1.50 | 180 ± 11.3a | 1037 ± 29.1a | 89.34 ± 3.68a |
| p value | 0.044 | <0.001 | 0.25 | <0.001 | <0.001 | <0.001 | <0.001 | 0.70 | <0.001 | <0.001 | <0.001 |
| Tanzânia-1 | |||||||||||
| Non-inoculated | 24.82 ± 3.62ab | 9.48 ± 0.83b | 49.80 ± 3.11b | 3.99 ± 0.36b | 7.79 ± 1.23 n.s. | 3.27 ± 0.38ab | 100.3 ± 5.52a | 6.26 ± 0.59b | 117 ± 5.53b | 1036 ± 156 n.s. | 60 ± 15.4 n.s. |
| A. brasilense | 23.85 ± 2.75ab | 12.90 ± 1.07a | 76.12 ± 6.65a | 8.67 ± 0.63a | 7.72 ± 1.03 | 3.51 ± 0.23ab | 52.4 ± 4.86b | 7.28 ± 0.48b | 177 ± 10.8a | 982 ± 115 | 120 ± 6.39 |
| B. subtilis | 16.31 ± 1.66c | 11.65 ± 0.25ab | 42. ± 3.97b | 5.05 ± 0.79b | 6.33 ± 0.69 | 2.84 ± 0.24b | 38.5 ± 1.50c | 6.43 ± 0.90b | 143 ± 13.4ab | 1035 ± 81.3 | 107 ± 10.9 |
| P. fluorescens | 28.70 ± 1.21a | 13.90 ± 1.25a | 65.93 ± 2.80a | 9.32 ± 0.96a | 8.87 ± 1.35 | 3.90 ± 0.13a | 41.3 ± 1.89bc | 10.37 ± 0.93a | 189 ± 26.3a | 1211 ± 256 | 83 ± 26.6 |
| R. tropici | 19.58 ± 0.77bc | 11.43 ± ab | 47.61 ± 1.30b | 7.03 ± 0.69a | 7.40 ± 0.91 | 3.06 ± 0.08b | 28.9 ± 4.83 d | 7.20 ± 0.52 b | 158 ± 19.6ab | 1126 ± 138 | 96 ± 12.9 |
| p value | 0.004 | 0.045 | <0.001 | <0.001 | 0.61 | 0.039 | <0.001 | 0.004 | 0.048 | 0.94 | 0.08 |
| Quênia | |||||||||||
| Non-inoculated | 17.88 ± 2.14b | 8.39 ± 0.57b | 47.49 ± 7.19 n.s. | 2.45 ± 0.11c | 3.74 ± 0.36c | 2.93 ± 0.31b | 78.6 ± 10.2c | 12.03 ± 1.13 n.s. | 149 ± 15.1 n.s. | 811 ± 33.6c | 17.03 ± 1.06c |
| A. brasilense | 19.70 ± 0.55ab | 9.62 ± 0.25ab | 53.99 ± 5.79 | 3.23 ± 0.19b | 5.51 ± 0.32b | 3.18 ± 0.08ab | 167.3 ± 18.6a | 13.28 ± 2.68 | 230 ± 31.5 | 1037 ± 20.3a | 54.38 ± 2.7a |
| B. subtilis | 24.16 ± 2.22a | 11.36 ± 0.66a | 47.70 ± 3.81 | 3.13 ± 0.33bc | 5.45 ± 0.30b | 3.56 ± 0.27ab | 104.3 ± 8.72bc | 9.50 ± 1.34 | 198 ± 19.1 | 1092 ± 60.5a | 36.15 ± 3.96b |
| P. fluorescens | 20.30 ± 1.89ab | 9.68 ± 0.89ab | 49.93 ± 4.27 | 2.98 ± 0.13bc | 5.06 ± 0.38b | 2.87 ± 0.24ab | 128.7 ± 11.5ab | 8.87 ± 0.41 | 211 ± 30.4 | 986 ± 43.4ab | 41.90 ± 2.95b |
| R. tropici | 25.10 ± 0.54a | 11.02 ± 0.68a | 57.37 ± 2.17 | 3.97 ± 0.28a | 6.29 ± 0.09a | 3.81 ± 0.21a | 154.5 ± 14.7a | 13.53 ± 2.06 | 205 ± 17.4 | 877 ± 88.7bc | 40.54 ± 3.16b |
| p value | 0.02 | 0.023 | 0.53 | 0.002 | <0.001 | 0.044 | <0.001 | 0.25 | 0.19 | 0.006 | <0.001 |
| Massai | |||||||||||
| Non-inoculated | 20.56 ± 0.73a | 8.56 ± 0.92 n.s. | 37.65 ± 1.84 n.s. | 6.04 ± 0.49 n.s. | 3.28 ± 0.26b | 2.91 ± 0.06a | 39.57 ± 2.76a | 15.32 ± 1.54 | 164 ± 31.4 n.s. | 429 ± 23.5b | 39 ± 2.85b |
| A. brasilense | 13.79 ± 0.30c | 9.31 ± 0.27 | 34.71 ± 0.83 | 5.48 ± 0.36 | 4.35 ± 0.06ab | 2.01 ± 0.07b | 26.14 ± 1.87b | 15.76 ± 0.91 | 203 ± 38.2 | 483 ± 17.5ab | 113 ± 5.31a |
| B. subtilis | 17.91 ± 1.69ab | 10.35 ± 0.65 | 33.68 ± 4.66 | 5.28 ± 0.30 | 3.30 ± 0.33b | 2.78 ± 0.24a | 29.43 ± 2.45b | 12.66 ± 0.88 | 174 ± 46 | 664 ± 70.9a | 165 ± 18.1a |
| P. fluorescens | 15.53 ± 1.80bc | 9.92 ± 1.41 | 36.36 ± 4.36 | 6.44 ± 0.71 | 4.77 ± 0.45a | 2.53 ± 0.28ab | 31.60 ± 2.57b | 13.48 ± 2.12 | 164 ± 9.28 | 563 ± 57.9ab | 177 ± 32.6a |
| R. tropici | 18.22 ± 0.36ab | 10.69 ± 0.62 | 35.21 ± 1.02 | 6.09 ± 0.58 | 5.05 ± 0.60a | 2.76 ± 0.09a | 30.09 ± 1.57b | 5.00 ± 0.78 | 226 ± 16.4 | 667 ± 91.6a | 45 ± 35.8a |
| p value | 0.006 | 0.41 | 0.88 | 0.54 | 0.005 | 0.011 | 0.006 | 0.41 | 0.53 | 0.02 | < 0.001 |
| Zuri | |||||||||||
| Non-inoculated | 15.9 ± 0.18bc | 7.4 ± 0.32c | 36.1 ± 3.49b | 3.9 ± 1.0 n.s. | 5.1 ± 0.85 n.s. | 2.4 ± 0.04c | 89.9 ± 2.31c | 7.0 ± 1.01 n.s. | 131.1 ± 5.48a | 1054 ± 123n.s. | 51.7 ± 8.18b |
| A. brasilense | 19.6 ± 2.05ab | 9.2 ± 0.80ab | 64.0 ± 6.04a | 5.5 ± 0.59 | 7.4 ± 0.67 | 3.1 ± 0.13ab | 141.0 ± 12.3a | 10.2 ± 1.64 | 132.8 ± 1.26a | 1002 ± 140 | 76.3 ± 3.99a |
| B. subtilis | 20.0 ± 0.22a | 10.4 ± 0.54a | 59.6 ± 3.00a | 4.9 ± 0.24 | 7.2 ± 0.50 | 3.3 ± 0.07a | 130.5 ± 3.61ab | 7.0 ± 0.14 | 110.2 ± 2.73ab | 853 ± 92.2 | 70.8 ± 6.27ab |
| P. fluorescens | 17.0 ± 0.78abc | 9.0 ± 0.28ab | 52.5 ± 4.35a | 4.5 ± 0.32 | 5.5 ± 0.19 | 2.9 ± 0.09ab | 119.1 ± 4.90ab | 7.7 ± 0.95 | 90.9 ± 5.46b | 999 ± 34.6 | 79.9 ± 3.32a |
| R. tropici | 15.6 ± 1.11c | 8.1 ± 0.44bc | 50.1 ± 5.34a | 3.8 ± 0.87 | 6.2 ± 0.91 | 2.9 ± 0.26b | 113.4 ± 12.56bc | 6.9 ± 0.61 | 95.3 ± 8.50b | 1031 ± 100 | 83.2 ± 5.62a |
| p value | 0.009 | 0.004 | 0.001 | 0.28 | 0.089 | 0.002 | 0.002 | 0.38 | 0.005 | 0.68 | 0.015 |
| CV (%) | 0.84 | 8.85 | 13.88 | 21.26 | 15.09 | 7.59 | 10.88 | 5.17 | 11.95 | 12.88 | 19.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães, G.S.; Rondina, A.B.L.; de Oliveira Junior, A.G.; Jank, L.; Nogueira, M.A.; Hungria, M. Inoculation with Plant Growth-Promoting Bacteria Improves the Sustainability of Tropical Pastures with Megathyrsus maximus. Agronomy 2023, 13, 734. https://doi.org/10.3390/agronomy13030734
Guimarães GS, Rondina ABL, de Oliveira Junior AG, Jank L, Nogueira MA, Hungria M. Inoculation with Plant Growth-Promoting Bacteria Improves the Sustainability of Tropical Pastures with Megathyrsus maximus. Agronomy. 2023; 13(3):734. https://doi.org/10.3390/agronomy13030734
Chicago/Turabian StyleGuimarães, Gabriel Silva, Artur Berbel Lirio Rondina, Admilton Gonçalves de Oliveira Junior, Liana Jank, Marco Antonio Nogueira, and Mariangela Hungria. 2023. "Inoculation with Plant Growth-Promoting Bacteria Improves the Sustainability of Tropical Pastures with Megathyrsus maximus" Agronomy 13, no. 3: 734. https://doi.org/10.3390/agronomy13030734