
Opportunities and Challenges of Castor Bean (Ricinus communis L.) Genetic Improvement
by
 , and
, and
Michela Landoni
1,†,
Greta Bertagnon
2,†,
Martina Ghidoli
2,
Elena Cassani
2,
Fabrizio Adani
2 and
Roberto Pilu
2,* 1
Department of Earth and Environmental Sciences, University of Pavia, Via S. Epifanio 14, 27100 Pavia, Italy
2
Department of Agricultural and Environmental Sciences—Production, Landscape and Agroenergy, University of Milan, Via Celoria 2, 20133 Milan, Italy
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2023, 13(8), 2076; https://doi.org/10.3390/agronomy13082076
Submission received: 14 June 2023
/
Revised: 26 July 2023
/
Accepted: 4 August 2023
/
Published: 7 August 2023
(This article belongs to the Topic Advances in Industrial Crops Physioecology and Sustainable Cultivation)
Abstract
:Castor bean (Ricinus communis L.) originated in East Africa and then diffused to warm-temperate, subtropical, and tropical regions of the world. The high lipid content in the castor beans is extracted for use in pharmaceutical and industrial applications. The castor oil lipid profile is naturally composed of 90% ricinoleic acid and the remaining 10% is mainly composed of linoleic, oleic, stearic, and linolenic fatty acids. The highly toxic compound ricin within the seeds is insoluble in oil, making castor oil free from this toxin and safe to use for industrial and cosmetic applications. Among the main uses of castor oil are reported industrial uses such as component for lubricants, paints, coatings, polymers, emulsifiers, cosmetics, and medicinal uses as a laxative. There is also significant commercial potential for utilization of the whole castor bean plant such as animal feed, fertilizer, biofuel, and also for phytoremediation. Several breeding programs have been planned to improve the castor’s characteristics needed for its current or potential uses. In this review, after summarizing data on castor bean agronomy and uses, we focus on the main advances in Castor bean classical and biotechnological breeding programs, underlining the high potential of this oil crop. In particular, the main challenges of castor breeding programs are to increase yield, mainly through the selection of growth habits allowing mechanized harvest, and beneficial compound content, mainly the oil, and to decrease the toxic compounds content, mainly ricin.
1. Introduction
The castor bean plant (Ricinus communis L) belongs to the family Euphorbiaceae, genus Ricinus, Ricininae subtribe. Castor is found in various regions across the globe, including tropical, subtropical, and warm-temperate areas [1]. It includes annual and perennial species that can reach the size of 5–12 m [2]. Wild progenitors of castor are still growing in East Africa, in particular in Kenya and Ethiopia. A recent study based on a chromosome-level genome assembly has demonstrated that these countries can be considered castor’s center of origin. Wild progenitors have a woody-tree phenotype and are characterized by dehiscent capsules, small seeds, and one single elongated trunk. The domestication process probably occurred about 3200 years before present (YBP) in an area somewhere between east Africa and west Asia. The domestication process, through the selection of important plant traits such as plant height, diameter of the main culm and seed size, resulted in significant changes in plant morphology and architecture, determining the shift from a woody tree to a semi-woody annual plant. From the original area of domestication, castor plants were distributed all over the world: Europe (2500 YBP), India (2000 YBP), China (1400 YBP), and finally America (500 YBP) [3].
Castor leaves, deciduous in perennial cultivars, are 15–45 cm long, palmate, and alternate with 5–12 deep lobes with serrate margins. Different leaf colors have been observed: young leaves are commonly reddish to bronze and change to dark green as they mature. The flowers are borne in a rounded panicle inflorescence 40 cm long, located at the terminal part of the main and secondary branches. Flowers are unisexual, 1–1.5 cm in diameter with a short pedicel. No corolla is present. Male flowers are located at the base of the inflorescence while female flowers, relatively few in number, are borne toward the apex of the inflorescence and are characterized by a three-celled superior ovary and three red or green styles. Fruits are usually three-lobed, with smooth or spiny capsules, 1.5–2.5 cm long, brown, dehiscing in three cocci, each one-seeded. Seeds are ellipsoid, 9–17 mm long, compressed, with a mottled and shiny seed coat with a distinct caruncle at the base, white and copious endosperm, and narrow cotyledons. The germination of the seedlings is epigeal [2].
Flowering usually occurs within six months after seed germination. The reproduction of castor plants follows a mixed pollination system: selfing and outcrossing through anemophily and entomophily. In wild conditions, more than 80% of the seeds are generated by cross-pollination. Castor plants are monoecious, with the ratio of pistillate and staminate flowers in racemes varying according to different genotypes and environmental conditions [4]. The development of female flowers appears to be encouraged by moderate temperatures, while high temperatures promote male flowers. Moreover, the level of nutrients supplied plays a role in determining the proportion of male/female flowers: in young plants supplied with high levels of nutrients, the female flowers predominate, while the male flowers are more abundant in older plants with low levels of nutrients [5] (Figure 1).
2. Castor Bean Chemical Profile
Castor bean is an important crop used in several industrial applications, mainly due to its high levels of ricinoleic acid, which is able to confer unique properties to the oil extracted from its seeds. In fact, castor is cultivated in different tropical and sub-tropical areas all over the world and the demand for castor oil has been rising in recent years. Castor oil is distinguished by its primary triglyceride component, triricinolein. Moreover, it comprises a diverse array of minor biological compounds, including carotenoids, tocopherols, tocotrienols, phytosterols, phospholipids, phytochemicals, and phenolic compounds. These constituents collectively bestow the oil with remarkable qualities, providing oxidation stability, anti-inflammatory effects, and antioxidant properties [6]. In addition to its unique composition, the high-quality nature of castor oil is evident in its acid, anisidine, iodine, viscosity, and saponification values, which outperform those of other vegetable oils [6].
2.1. Oil Lipid Profile
Ricinus communis is cultivated for the extraction of oil from its seeds. Castor oil is a non-edible oil mainly used for pharmaceutical and industrial applications. It is a colorless to very pale-yellow liquid with a distinct taste and odor. Its boiling point is 313 °C (595 °F) and its density is 0.961 g/cm3 [7]. Castor oil stands out among vegetable oils due to its exclusive supply of ricinoleic acid, a hydroxylated fatty acid. This distinctive compound constitutes approximately 90% of the castor oil content. No other commercially available vegetable oil exhibits such a significant predominance of a single fatty acid. Notably, the concentration of ricinoleic acid appears to be minimally influenced by environmental conditions [8,9,10]. The primary component of its fatty acid profile is ricinoleic acid, accompanied by minor acids like linoleic, oleic, palmitic and stearic acid (Table 1).
2.2. Noxious Compounds: Ricine and Its Homologue
Unfortunately, castor also contains ricin, which represents the most toxic vegetable compound known [11]. The lethal dose for adult humans is 0.35–0.7 mg kg−1 of body weight by inhalation, whereas by ingestion has been estimated to be between 1 and 20 mg kg−1 of body weight. The lower level of ricin toxicity by ingestion is due to immune barriers present in the intestinal tract and the stomach acid environment [12].
Ricin is a dimeric glycoprotein constituted of A- and B-polypeptide chains covalently linked by a disulfide bond [13]. The B-chain binds to glycoproteins and glycolipids on the cell surface and helps the entrance and movement of the A-chain into animal cells. The enzymatic activity of the A-chain inactivates ribosomes by depurinating the first adenosine in the GAGA nucleotide sequence in the conserved loop of the 28 S rRNA subunit [14,15]. One thousand ribosomes per minute can be irreversibly inactivated by a single ricin A-chain, interrupting protein synthesis and resulting in cell death [16]. Several other plant species produce toxins similar to the ricin A-chain, highly concentrated in the seeds, but not as toxic as ricin because of the absence of an efficient B-chain [17]. According to a recent study, it seems that a 43 amino acids motif, located in the A subunit, is largely divergent between Ricinus communis and other plants producing ricin (Aleurites fordii and Jatropha curcas in particular) [18]. Among the amino acids encoded by the variable motif, Tyr 129 has been identified as pivotal for the efficiency of the A chain: if mutated, the activity of the A chain was reduced seven-fold [19].
Castor oil is free from ricin, as this toxin is insoluble in oil. Castor cake, the remaining industrial sub-product generated after extraction of the oil from castor seeds, is rich in valuable proteins and fiber and could potentially be used as animal feed. Unfortunately, due to the presence of ricin, it is generally used only as an organic fertilizer [17]. Castor bean residues have been successfully detoxified with a fermentation process carried out by some microorganisms such as Paecilomyces variotii and Aspergillus niger or by chemical treatment with calcium oxide and calcium hydroxide [20,21]. From the economic point of view, these processes are still not efficient enough to be used on a large scale [17,21]. For these reasons, the development of transgenic castor plants presenting low levels of ricin may be a possible solution for using castor meal as feed. Castor bean seeds also contain the ricin homologue R. communis agglutinin (RCA120), which is a tetrameric protein composed of two A-chains and two B-chains (both chains are very similar to the ricin ones, 90% and 84%, respectively). RCA120 presents a reduced toxicity but it causes blood coagulation when injected into mammalians’ bloodstream [22].
3. Uses
The high ricinoleic acid concentration allows the production of high purity derivates. The chemical industry uses castor oil as an essential component in several groups of products like lubricants and greases, inks, emulsifiers, bio-based polyurethanes, and cosmetics [9,10,31,32]. This wide range of applications is possible because of the different characteristics of castor oil derivatives, like high viscosity, high melting point, and insolubility in aliphatic petrochemical fuels and solvents, making it highly suitable as a lubricant under extreme conditions [10,33]. Castor oil is a lubricant that is environmentally friendly, as it exhibits good biodegradability and renewable properties. However, when used as a green lubricant, castor oil does have a few drawbacks. These include a low viscosity index and low oxidative stability, which can be attributed to the presence of unsaturated bonds [34,35,36]. Castor oil has long been used as a purgative and continues to be recognized as a safe and effective laxative. Many phytochemicals found in the plant tissues and seeds of castor have potential medicinal uses [37]. In several countries, castor oil is also considered an option for the production of biodiesel, because of its high content of ricinoleic acid, its solubility in alcohol, and its conversion performance of 90–95% [38]. Currently this use is not common, as the economic value of castor oil as an industrial oil is very high. In fact, castor oil is in high demand by the chemical industry to manufacture high quality products [17]. Another potential use of castor in the future is as an animal feed and organic fertilizer. Castor meal is not currently used as an animal feed because of the high levels of ricin. In the absence or significant reduction of ricin toxicity, castor meal would serve as an excellent protein source in animal feed formulations. Unfortunately, no process able to detoxicate the castor meal from ricin on an industrial scale has yet been developed, but an interesting option seems to be represented by lime (CaCO3) treatment, which is also able to increase the meal’s pH and Ca levels [39,40,41]. Due to the high N content, the fast rate of mineralization, and the anti-nematode effect, castor meal represents a good resource for crop fertilization all over the world. Castor meal contains 75 g/kg of N. Since the mineralization of N is very fast, castor meal should not be added to the soil at rates higher than 45 g/kg of soil [42,43]. Husks could also be used as fertilizer, but their high K content (45 g/kg) associated with a low N content (18.6 g/kg), make it necessary to blend castor husks with other N-rich organic materials in order to provide plants with a nutritionally balanced fertilizer [44,45]. R. communis is also grown in many landscapes as an ornamental plant, due to its big leaves, diversity of colors in stem and fruits, rapid growth, and drought tolerance. Castor fields can also be used for honey production: if hives are placed in a castor field, castor nectaries are regularly visited by honeybees (Apis mellifera), resulting in honey production [46]. This species may also be considered a good option for phytoremediation of soils contaminated with heavy metals. That is because castor is not a food crop and it is an hyperaccumulator of Pb [47] and Zn [48], and it is quite tolerant to Cd [49]. In particular, recent research demonstrated that a combined application of glutathione and citric acid increases castor plants’ Pb tolerance level [50].
Furthermore, castor bean can be used in preventing and alleviating soil compaction by generating numerous biopores within the soil profile, in particular in no-till soils. These biopores create pathways for the roots of subsequent crops to grow, resulting in enhanced root colonization at deeper levels. It helps reduce nutrient losses through leaching, facilitating nutrient cycling and contributing to the overall health and productivity of the soil [51,52].
Recently, castor bean has also been considered as a potentially important source of lupeol. Lupeol is a triterpene which has been studied for its anti-diabetes, anti-inflammation, and anti-cancer properties. Lupeol is also a precursor of betulinic acid (BA) and its derivatives, which have shown anti-HIV and anticancer properties [53]. Even though lupeol can be found in various plant species, its concentration is generally limited, but in castor, a high amount of this compound accumulates on the outer tissues, as an essential constituent of waxes [54]. Due to all these potential uses, the demand for castor oil in the global market has been rising in the recent years, with a supply not being at the moment able to satisfy the demand. In 2020, the three top world producers were India, Mozambique, and Brazil, with a seed production of 1.8 million, and 72 and 35 thousands of tons, respectively [55].
4. Agronomy
Castor bean cultivation is generally done by small farmers all over the world, mainly in India, Mozambique, China, and Brazil. The production in these countries is concentrated in arid and semiarid regions, where the rainfall is generally low, although the limited availability of water is the major factor affecting yield [56]. The culture system generally involves intercropping with food crops (such as maize and beans) with low adoption of technologies. The whole system of production, from planting to processing, is manual with little or no soil tillage and fertilization. Mostly local varieties are cultivated, and they are characterized by a long vegetative cycle and uneven seed maturation. In order to increase castor yields, the mechanization of the production process is pivotal, and breeding programs to develop a suitable plant architecture are currently the only way to make the mechanization of the crop cycle possible [17,57,58,59]. The annual varieties of castor require about four to nine months to reach maturity, while the perennials may continue yielding for 10–15 years. Capsules of non-shattering varieties (improved varieties) can be harvested when they are fully dry, while shattering types are harvested when they turn in color from green to yellow [2].
The central spike on the main rachis matures before the side-branches’ spikes, making necessary two to three pickings to complete the harvest. It is worth noting that harvesting of immature capsules should be avoided because of the negative impacts on oil content. Furthermore, it has been reported that before threshing, the capsule should be sun dried for four to five days [2]. Castor yield may vary from 1 to 3 tons of seed/ha depending on climatic conditions, crop management, and variety. The percentage of oil obtained can reach up to 50% by weight [56]. Castor seeds are characterized by slow, irregular, and cold-sensitive germination. Furthermore, 14–15 °C is the minimum tolerated temperature for germination, the optimum is 31 °C, and the maximum 36 °C. Germination and seedling emergence are often delayed by low soil temperature, resulting in irregular crop development. By implementing appropriate agronomic practices, such as selecting suitable cultivars, utilizing high-quality seeds, strategic planting dates, irrigation, soil fertilization, effective weed and pest management, disease control, mechanical harvesting, and postharvest management, seed yield in most regions of castor production can be significantly increased. A castor field yielding 2000 kg/ha of seed will remove 80 kg/ha of N, 18 kg/ha of P2O5, 32 kg/ha of K2O, 13 kg/ha of CaO, and 10 kg/ha of MgO from the soil [60]. In the early phases of development, the growth of the castor’s leaf area is slow. For this reason, weeds can quickly cover all the available soil surface. Trifluralin, Pendimethalin, and Clomazone are some of the pre-emergence herbicides that can be used in castor cultivation. These herbicides are useful to control monocotyledonous weeds, but less effective against broad-leaved plants [61]. For combatting the growth of broad-leaved weed plants, the only post-emergence herbicide that has been proven to be effective in castor cultivation is Chlorimuron-ethyl. An effective weed management program that does not lead to phytotoxicity on the castor crop is a treatment with a pre-emergence followed by a post-emergence herbicide applied at 20 DAE (days after emergence) [62]. Castor is affected by several diseases, but only a few of them are responsible for economic losses. The three major pathogenic fungi affecting castor are gray mold (Botryotinia ricini), vascular wilt (Fusarium oxysporum), and charcoal rot (Macrophomina phaseolina). Gray mold is considered to be the most serious castor disease all over the world. Unfortunately, only a few advances have been obtained in the management of this pathogen, mainly because B. ricini has a wide range of hosts within the Euphorbiaceae family, including weeds and ornamentals [63].
Although breeding programs have failed to isolate resistant genotypes, genotypes with moderate levels of tolerance have been identified [17]. The genetic characterization of disease resistance pathways will thus be a fundamental step in order to develop tolerant castor varieties. Castor oil is a non-edible product. For this reason, marginal areas should be chosen for castor production, in order not to subtract lands from food crop cultivations. In this scenario, tolerance to abiotic stresses is of particular importance. Castor plants are more sensitive to water stress in the early stages of growth. At the cellular level, water stress causes a reduction in callus initiation, nitrate reductase activity, and chlorophyll content [64]. Moreover, drought stress increases cuticular wax load [65] and abscisic acid concentration in the phloem sap, thus suggesting a role for ABA as a water stress signal also for sinks, not only for stomatal movement [66]. An important mechanism for drought tolerance in castor is osmotic adjustment. A study analyzing the association of osmotic adjustment (OA) with drought tolerance and yield in castor reported that the OA increased with increasing stress periods, with genotypes characterized by high osmotic adjustment (HOA) showing greater levels of proline, total soluble sugars (TSS), total free amino acids (FAA), and potassium in comparison with genotypes with low osmotic adjustment (LOA) in response to water stress. Furthermore, it was reported that HOA genotypes of castor produced significantly higher seed yield than LOA genotypes [67]. The mechanisms involved in drought resistance were analyzed in a study in which water stress was applied to castor plants grown in greenhouse conditions. Water stress was significantly associated with a reduction of plant growth, with a constant decrease of carbon assimilation and transpiration and the increased stomatal movement limitation observed suggesting that stomatal closure was the main factor responsible for photosynthesis reduction. The conclusions suggested by the authors were that drought resistance in castor seems to be associated to an early growth response with efficient stomatal control and high levels of net CO2 fixation [68]. Furthermore, it has been reported that when the water stress is removed, the castor plants completely recovered their normal photosynthetic activity in 24 h [69]. Another study conducted to analyze the association of OA with drought tolerance and yield in castor compared hybrids and their respective parents in response to moisture stress at primary spike development stage [67]. The authors described the existence of variability for OA among the different genotypes analyzed. Under water deficit, HOA genotypes were characterized by higher leaf RWC (relative water content), higher ELWRC (excised leaf water retention capacity) and were able to maintain higher leaf water potentials (Ψl) in comparison with LOA genotypes. Furthermore, HOA genotypes resulted in higher seed yield [67].
Growth and production of castor are also inhibited by high salinity in both the irrigation water and in the soil. The highest sensitivity is in the early stages of development with increasing salinity level resulting in delayed and reduced emergence of castor seeds [70,71]. In particular, increasing levels of Na salinity have been reported to damage the photosynthetic apparatus and induce proline accumulation [72]. Castor plants are extremely sensitive to hypoxia. Literature data reported that after 2–6 h of soil flooding, plants showed a reduction in CO2 uptake, stomatal conductance, leaf elongation, transpiration, root conductance, and abscisic acid production from roots [73]. Castor plants undergoing a continuous flooding are permanently damaged after three days and die after four days [17]. Successful cultivation of castor on marginal land relies on implementing appropriate agricultural techniques and selecting genotypes that are better adapted to challenging conditions. For example, the presence of high levels of exchangeable aluminum in acidic soil poses a significant challenge for castor cultivation. Aluminum toxicity occurs when the soil pH drops below 5.5, leading to low levels of exchangeable bases and organic matter. In such circumstances, toxic forms of aluminum become soluble in the soil solution, negatively impacting root growth and functionality [74,75].
Despite being a crop that requires minimal inputs and is suitable for cultivation in European climates, the limited mechanization available for harvesting castor beans restricts its production to underdeveloped or developing agronomy countries, where manual harvesting is feasible due to low labor costs [58]. The challenges associated with mechanically harvesting castor beans stem from the crop’s inherent traits of indeterminate growth and evergreen habit, leading to high production of fresh aerial biomass and uneven ripening of capsules [58,59]. These characteristics make direct combine harvesting practically unachievable: the large volume of fresh biomass easily obstructs the combine harvester’s cleaning mechanism, and determining the optimal harvest period is difficult, resulting in significant seed loss [59]. However, recent advancements have nearly resolved the issue of indeterminate growth in castor plants. Dwarf genotypes are now commercially available and have demonstrated promising performance even in marginal conditions [57]. Generally, dwarf hybrids exhibit a higher harvest index, lower seed dehiscence, and earlier flowering, which shortens the productive cycle [57]. In particular, the paper of Severino and coauthors showed that seed yield in temperate locations was not affected by plant density, whereas in tropical conditions, higher plant density resulted in increased productivity. During the reproductive phase, the seed yield showed the strongest correlation with plant leaf area and height, particularly at the beginning of this phase [57].
5. Genetic Resources
The genome of R. communis consists of 20 chromosomes (2n). A first draft with a 4.6× coverage was published in 2010 by Chan et al. [76], which studied the cultivated variety “Hale”. The size of the castor genome was estimated to be around 350 Mb with 31,000 genes [76]. A second genome sequence of an Ethiopian wild castor was released in 2022 by Lu et al. [18]. The result was more accurate, with a 170× coverage, the genome size 316 Mb for a total of 31,000 genes. The similarity between the two genomes is about 99.16%, suggesting that the domesticated varieties do not diverge very much from the wild ancestors [18]. R. communis germplasm is conserved in 10 different countries and includes a total number of 11,300 accessions. The National Bureau of Plant Genetic Resources, in India, is the major germplasm bank, which accounts a total of 4303 accessions of castor from 39 different countries. In Brazil, the germplasm bank of Embrapa includes 620 accessions. The 1043 accessions conserved in the USDA (United States Department of Agriculture) germplasm bank have been used for the quantification of oil and ricin. Great variability of ricin and oil content in the seeds has been reported [17,77].
The main world centers hosting castor germplasm banks are summarized in Table 2.
6. Genetic Improvement
One of the main challenges of castor breeding programs is to make mechanical harvest possible. The perennial nature together with the indeterminate growth habit of castor plants are the main obstacles to mechanical harvesting. Plants adapted to mechanized production should be short (1.0–1.5 m), with uniformly maturing racemes, and a low number of lateral branches [78,79]. Early maturation is a significant trait sought after for castor cultivation, particularly in regions with limited growing seasons or tropical areas where multiple crop cycles are practiced annually. Unfortunately, there is a negative correlation between early maturity and high seed yield. A search for traits of interest such as dwarfism and early flowering and maturation, focused on Indian [80,81] and Brazilian germplasm [57,82,83,84] was reported. In the study conducted by de Oliveira Neto and coauthors [57], a careful selection of castor bean accessions with considerable commercial value, especially for mechanized harvesting, has been identified. These selected accessions possess desirable traits such as ideal plant height, diameter, seed weight, and oil content. As a result, these specific genotypes hold great promise as potential contributors of favorable alleles in castor bean genetic breeding programs [57].
Decreasing the ricinoleic acid content and increasing the percentage of oleic acid would enhance performance of castor oil as a raw material for biodiesel [17]. These characteristics are present in a mutant conserved in the Brazilian germplasm bank of Embrapa [85] and are determined by a mutation in FAH12 oleate hydroxylase [86]. Another objective of castor genetic improvement is the development of castor plants in which the content of ricin in the seeds is strongly reduced or absent. Ricin is a dangerous toxin for human and animal health, since even though it is not contained in the extracted oil, its concentrations in the oil by-products, such as castor meal and husks, is high and prevents their use as feed or nutritional supplement. Some of the accessions present in the USDA germplasm bank have been used to create by traditional breeding the cultivar “Bringham”, characterized by a reduced ricin content [17].
6.1. Classical Breeding
The genetic diversity present in a crop species is the raw material, and the starting point for breeding programs, for example the isolation of a natural mutant named OLE-1 with High Oleic acid content.
OLE-1 represents a breakthrough among stable castor genotypes, displaying the highest proportion of oleic acid and the lowest proportion of ricinoleic acid reported to date. This exceptional profile makes it a highly promising source of elevated oleic acid levels, with valuable implications for applications in both the industrial and food sectors, where exceptional oxidative stability is crucial. Moreover, OLE-1 offers a unique opportunity for exploring the biosynthesis and genetics of ricinoleic acid content in castor [85]. Chemical and physical mutagenesis can also be used to increase the variability present in plants, as reported, for example, in cotton, where mutants with different fatty acid profile, fiber properties, and other important qualitative characteristics have been isolated [87,88,89,90,91].
Three types of radiation (γ rays, fast neutrons, and thermal neutrons) were applied to castor plants to generate mutations and consequently to increase genetic variability. The thermal neutron treatment has been used, for example, to reduce apical dominance, days to maturity and plant height. Gamma rays were used to develop pistillate lines resistant to wilt [92] and to isolate useful mutants with improved seed yield and other important characteristics [93].
Some papers have reported on the analysis of castor bean genetic diversity through the use of molecular markers such as AFLP (Amplified Fragment Length Polymorphism) and SSR (Simple Sequence Repeats) [26,38,65,66,94]. The data concerning the level of genetic variability present in castor beans collections are quite different in the different publications. While some papers reported low genetic diversity among the accessions analyzed, probably due, among other factors, to selective breeding, domestication, and samples not being representative of the population genetic variability, Pecina-Quintero et al. [38] reported a high level of genetic diversity when analyzing a collection of castor bean plants from Chiapas, Mexico by AFLP and SSR markers [38]. Once polymorphic molecular markers are found, they can be used not only to assess the genetic diversity in a collection but also for unequivocal varietal identification, phylogenetic analysis, and for marker assisted selected breeding programs. Various schemes of breeding programs have been used to improve castor bean. Mass selection consists in the selection of superior plants and the elimination of undesirable traits within a plant population. Mass selection is typically used for the selection of traits characterized by a high heritability. Concerning castor bean genetic improvement programs, mass selection was the most effective method to increase the frequency of pistillate plants [2]. The backcross method of selection is the most effective method for improving some simply inherited, qualitative characteristics in commercial cultivars or promising elite lines. This method has been used in castor to transfer monogenic traits such as dwarf habit, non-spiny capsules, stem color, plant height, and wilt resistance [95]. The pedigree method, based on five to six generations of selection of inbred lines, with subsequent progeny testing, has been used to select castor lines with high oil content and resistance to fusarium [96]. This same method was adopted in breeding programs aimed at increasing seed yield, with a selection scheme focused on long semi-compact spikes (>40 cm), number of capsules per raceme (>60), number of effective spikes per plant (>5), and single seed weight (>300 mg) [17]. Furthermore, the pedigree method, for example, has been reported to be used to develop several tall-type cultivars with late maturation in India. The single seed selection method was recently used to select fast germinating castor seeds at the suboptimal temperature of 22 °C. Moreover, the results showed that at 22 °C, the freshly harvested seeds are characterized by a low germination ability, which, however, can increase at higher temperatures (30 °C) or improve gradually during the storage [97]. Dwarf castor varieties are associated with higher yields and allow a more rational management in the field [98]. Plant height is a trait controlled by auxins, which are the plant regulators implied in regulation of cell division, elongation, and differentiation in the meristems (apical meristem and vascular cambia) [99]. Apical growth is often repressed when auxins’ transport or biosynthesis are impaired, leading to a major plant branching, dwarf and usually high yielding phenotype [100,101]. Among the different auxins, IAA (Indole-3-acetic acid) is one of the most physiologically active. In a recent study, based on BSA (Bulk Segregant Analysis), the Rc5NG4-1 gene, encoding for an IAA transport protein, was identified as candidate gene controlling castor height. An SNP (single nucleotide polymorphism, A/G), determining an amino acid substitution in the 5NG4 IAA transporter (from Y to C in the 7th helix loop), has been reported to be responsible for the dwarf phenotype of castor plants. In particular, the authors reported that in dwarf plants, the mutation in the Rc5NG4-1 gene is associated with a reduced capacity to transport free IAA from the apical bud to the node in comparison with the wild type [102]. The choice of the most suitable breeding techniques for obtaining high yielding varieties is strictly connected with the type of genetic interaction characterizing the different genes responsible for the inheritance of the quantitative trait under selection. A recent significant contribution was given by Mori et al. [103], who investigated the genetic architecture of oil content, seed yield and its related biometric traits through “generation mean analysis” in four different crosses for twelve generations [103]. Generation mean analysis is an efficient tool to understand the gene effects involved in the expression of a trait and the results of this study showed that total seed yield and oil constitution, which are quantitative traits, are governed by partial dominance to overdominance. Furthermore, the presence of non-allelic interactions for economically important traits in castor were previously demonstrated by several different authors. In particular, Solanki et al. in 2003 observed additive variance and epistasis in the inheritance of plant height, number of nodes up to the main raceme, length of the main raceme, and the number of capsules of the main raceme [104]. Similarly, Golakia et al. [105], in 2015, demonstrated the presence of additive, epistatic, and dominance variance in the number of nodes up to the main raceme, the length of the main raceme, and the total seed yield per plant [105]. Virani et al., in 2013, also confirmed the presence of all these gene effects in yield components in castor [106]. The presence of epistasis suggests that direct selection methods may not be suitable for improvement of castor seed yield. In fact, it would be difficult to fix useful genotypes because the positive effect of one gene might be covered by the negative effect of another gene. In conclusion, literature data showed that oil content, seed yield per plant, and its component traits are governed by varied levels of additive, dominance, and epistatic gene effects. When additive as well as non-additive effects are involved, breeders should adopt a breeding scheme able to exploit both types of gene effects, as for example the reciprocal recurrent selection scheme as reported by Mori and coauthors [103]. India is the pioneer in the production of hybrid castor in the world. The first commercial hybrid, GCH-3, was developed by combining different pistillate line genotypes from Texas, USA and India. After the development of an indigenous pistillate line, VP-1, several high yielding hybrids were released for cultivation in India and their success was immediate. The increase of seed yield in castor hybrids depends on both the genetic diversity and the individual combining ability of the parents [105,107,108]. Many of them are characterized by resistance to fusarium wilt and high seed yield potential [109].
6.2. New Breeding Techniques for Oleaginous Crops: GMO Technology and Genome Editing
Genes from different species can be introduced into the castor genome in order to confer agronomically important traits through genetic transformation techniques. Unfortunately, genetic transformation of castor remains challenging because of the lack of an appropriate protocol for castor plant regeneration [110]. In fact, this species is recalcitrant to in vitro regeneration, and the development of transgenic plants has been limited by the difficulties in this fundamental step necessary to regenerate plants from transformed calli. The rapid regeneration of castor plants holds great value for hybrid production, as utilizing dihaploids would significantly reduce the time and cost involved in generating inbred populations [17]. Successes in castor bean regeneration have been reported by Bertozzo and Machada who described a culture medium with reduced NO3/NH4, and lacking FeSO4 [111]. Agrobacterium tumefaciens is commonly used in genetic transformation to obtain transgenic plants expressing exogenous genes. Several methods have been developed to transfer genes into castor explants using A. tumefaciens or a particle gun [112], however, the efficiency of transformation in both cases is relatively low (0.8% and 1.4%, respectively). Recently, mainly due to the difficulties of the in vitro regeneration of explants, some researchers tested on R. communis a transformation technique via Agrobacterium originally developed for rice [113]. The method is based on the direct injection of Agrobacterium into the developing endosperm of castor oil fruits. The rate of transformation reached was 80% and a good proportion of viable seeds was also reported [114]. Several attempts to eliminate toxic and allergenic substances have been made by traditional breeding techniques. Castor lines with 70–75% decreases in ricin and RCA content were developed [115,116]. Unfortunately, the partial reduction obtained so far cannot be considered enough to ensure safe castor cultivation.
An example of successful application of GMO technology for ricin improvement was published by Sousa et al. [117] who managed to silence the ricin gene in the endosperm of castor bean seeds to produce non-toxic castor bean genotype, exploiting the RNA interference (RNAi) technique. The authors reported that by the ELISA test, ricin was not detected in transgenic seeds obtained through the biolistic transformation method [90]. Different studies reported the characterization of pathways involved in traits that are possible targets of breeding programs. Lupeol has been recognized as a fundamental wax component found in the epidermis of castor plants [118]. Its content varies significantly among the different castor varieties. Eleven enzymes, encoded by 21 different genes, are involved in lupeol synthesis in castor [119]. A study has recently been carried out in order to understand the mechanism regulating lupeol synthesis. A transcriptome analysis showed that several auxin-related genes have different patterns of expression in the hypocotyl, characterized by a low level of lupeol, and stem internodes, with a high level of lupeol, suggesting that auxin might be implied in the regulation of lupeol biosynthesis. Furthermore, several auxin-responsive cis-elements are found in promoter regions of two key enzymes involved in lupeol production (RcHMGR and RcLUS), of which the expression seemed to be induced by auxin treatments, and lupeol content was reported to be increased by auxin treatment and decreased by auxin transport inhibition [120]. Wang et al. [121] have identified a glutathione peroxidase gene family in the genome of R. communis. Glutathione peroxidases (GPXs) are key ROS scavengers that use reduced glutathione (GSH) or thioredoxin (Trx) to convert hydrogen peroxide (H2O2) into simple water [122,123,124]. A considerable number of studies have demonstrated that plant GPXs act as crucial regulators in response to diverse abiotic stresses such as salt, drought, and cold [125,126,127,128,129,130]. Searching in the R. communis genome database using known GPXs from other plant species, five GPXs were identified and named from RcGPX1 to RcGPX5. By the analysis of sequences upstream to the translation start site, several cis-acting elements responsive to stresses and plant hormones were identified. The expression patterns of RcGPXs under cold, salt, and drought stress in roots, stems, and leaves were investigated by qRT-PCR analysis and were found to be largely down or upregulated in the different tissues. One of the most serious environmental problems of agriculture is soil salinization, which affects crop growth, development, and productivity. About 20–50% of the world’s cultivated area suffers from salinity problems, and this area is growing. Hence, improving salt tolerance of crops has become an urgent task to enhance crop production. Rational breeding strategies are based on the knowledge of physiological and molecular mechanisms involved in plant responses to stresses [131,132,133]. Castor has good salt tolerance, but “early seedling” is the most vulnerable developmental stage for plants under stresses. Wang et al. [134] have recently reported that castor cotyledons are able to accumulate high amounts of Na+ and to provide more energy to help true leaves to cope with salt stress. In particular, proteomic analyses have shown that in high salt conditions the true leaves save carbon structures to synthesize more osmotic adjustment substances, while chlorophyll synthesis and electron transfer are enhanced to maintain a high photosynthesis rate [134]. The root is directly exposed to salinity in soil with the highest susceptibility to this stress during the seedling developmental stage. Under optimal growing conditions, the primary role of cotyledons is providing resources to support root development. Under salt stress the root/cotyledon ratio decreased from 0.52 to 0.27, suggesting that the cotyledon got more carbon than the root to maintain the development of the unique photosynthetic organ during the early seedling stage. Roots also presented an increased level of unsaturated fatty acids in order to enhance the cell wall fluidity and to protect cells from ion toxicity; the root also accumulates Na+ to maintain a high K+/Na+ ratio in the cotyledon and preserve metabolic activity [135]. In a recent study, Lei et al. [136] have identified the differentially expressed genes (DEGs) for differential salt tolerance in wild and cultivated castor plants. Multiple phytohormones and transcription factors are correlated with salt-tolerance. In salt stress conditions, 2C protein phosphatases (PP2C) homologs are upregulated. Six and four IAA homologs were significantly upregulated in cultivated and wild castors, respectively. Moreover, major jasmonate response genes are significantly upregulated under salt stress. Seedlings subjected to NaCl exhibited lower enrichment of DELLAs, which are growth inhibitors acting through the binding to gibberellins’ receptors [137]. Among all the genes differentially expressed in normal and salt stress conditions, comparing wild and cultivated castor plants, researchers also have identified multiple transcription factor families (ERF/AP2, WRKY and bHLH) [136].
Ricinus communis originates in tropical regions but is currently cultivated in many subtropical regions worldwide. In temperate regions, the early planting strategy is commonly used to avoid drought periods, thus improving the yield. Unfortunately, castor bean is sensitive to cold stress [138]. Therefore, it is imperative to understand the molecular mechanism of cold response by the identification of responsive genes/proteins to this stress to improve the cold tolerance of castor. Recently, an investigation on differential abundance protein species (DAPS) associated with cellular responses to cold stress was performed using isobaric tags for relative and absolute quantification (iTRAQ) technologies. This analysis revealed complex changes in DAPS between cold-stressed imbibed seeds and unstressed control seeds. The 127 DAPS identified were involved in translation and post-translational modification, stress responses, signal transduction, carbohydrate and energy metabolism, lipid transport, and metabolism [139].
Oleaginous crops such as soybean, rapeseed, peanut, and sunflower are essential for the production of edible oils and constitute the main source for the production of biodiesel. As they play a pivotal role in the global economy, genetic improvement programs are aimed at obtaining more productive lines, different composition in fatty acids and oil, and at the completion of the domestication process of species that have been only recently identified as oleaginous crops, for example Camelina sativa, reviewed by Ghidoli et al., 2023 [140]. These goals can be achieved not only through traditional breeding, but also by new breeding technologies (CRISPR, TALEN, ZFN), which allow the modification of specific genes encoding for traits of interest. In Table 3, the genes of oleaginous crops that have already been modified by exploiting these technologies are listed.
As shown in Figure 2, there are numerous methodologies available to improve R. comunis. In particular, through intraspecific and inter-specific crosses, new varieties of castor can be obtained, while, the use of recurrent backcrosses, mutational breeding, transgenesis, and genome editing techniques allows for the development of essentially derived varieties, which are subject to regulation under EU legislation [157].
7. Conclusions
Castor bean is a promising crop that originated in east Africa and west Asia and from its center of origin diffused all over the world in tropical, sub-tropical, and warm temperate regions. The oil extracted from its seed is suitable for many industrial purposes. It is currently used for pharmaceutical and industrial purposes, but other potential uses are under development, for example: the oil is a promising source of biofuel, and castor meal could be used as animal feed and as organic fertilizer, which also exhibits insecticide and nematicide function. The possibility to cultivate castor in marginal lands can overcome one of the main issues linked to the sustainable cultivation of non-edible crops, i.e., the soil competition with food crops. Other issues limiting castor uses are the presence of the highly toxic compound ricin in castor seeds, a plant architecture and development making mechanized harvest difficult, and the susceptibility to both biotic and abiotic stresses. To overcome these limitations, different breeding programs have been developed with the purpose of selecting the desirable traits such as the dwarf plant type, early flowering and maturity, high seed weight and oil content, reduced ricin content, and resistance to biotic and abiotic stresses. In addition to the classic breeding programs, the use of new breeding technologies is also achieving promising results with the aim of improving the traits of interest through the editing of the castor genome. A still open challenge in the field is the development of reliable protocols for the regeneration of castor bean plants.
The achievement of these goals will allow the development of castor plants with increased yield, quality, and safety, allowing the crop to satisfy the demand of the global market for the multiple purposes for which castor bean is and will be used.
Author Contributions
Conceptualization, R.P. and M.L.; writing—original draft preparation, M.L. and G.B.; writing—review and editing, M.G., E.C. and R.P.; visualization, M.L.; supervision, R.P., F.A. and M.L.; project administration, R.P.; funding acquisition, R.P. All authors have read and agreed to the published version of the manuscript.
Funding
Agritech National Research Centre and received funding from the European Union NextGenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR)–MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4–D.D. 1032 17/06/2022, CN00000022) to R.P. This manuscript reflects only the authors’ views and opinions; neither the European Union nor the European Commission can be considered responsible for them.
Data Availability Statement
Not applicable.
Acknowledgments
We wish to thank Lesley Currah for her editing and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weiss, E.A. Oilseed Crops; Blackwell Science: Oxford, UK, 2000; ISBN 0-632-05259-7. [Google Scholar]
- Chakrabarty, S.; Kalam Mohammad Aminul Islam, A.; Yaakob, Z.; Kalam Mohammad Mominul Islam, A. Castor (Ricinus communis): An Underutilized Oil Crop in the South East Asia. In Agroecosystems—Very Complex Environmental Systems; IntechOpen: London, UK, 2021. [Google Scholar]
- Xu, W.; Wu, D.; Yang, T.; Sun, C.; Wang, Z.; Han, B.; Wu, S.; Yu, A.; Chapman, M.A.; Muraguri, S.; et al. Genomic Insights into the Origin, Domestication and Genetic Basis of Agronomic Traits of Castor Bean. Genome Biol. 2021, 22, 113. [Google Scholar] [CrossRef]
- Zimmerman, L.H.; Smith, J.D. Production of F1 Seed in Castorbeans by Use of Sex Genes Sensitive to Environment. Crop Sci. 1966, 6, 406–409. [Google Scholar] [CrossRef]
- Shifriss, O. Growth and Sexuality of Ricinus communis L. in a Constant Environment. Am. Nat. 1964, 98, 187–189. [Google Scholar] [CrossRef]
- Yeboah, A.; Ying, S.; Lu, J.; Xie, Y.; Amoanimaa-Dede, H.; Boateng, K.G.A.; Chen, M.; Yin, X. Castor Oil (Ricinus communis): A Review on the Chemical Composition and Physicochemical Properties. Food Sci. Technol. 2021, 41, 399–413. [Google Scholar] [CrossRef]
- Castor|Kaiima. Available online: https://kaiima.com/castor/ (accessed on 10 May 2023).
- da Silva Ramos, L.C.; Tango, J.S.; Savi, A.; Leal, N.R. Variability for Oil and Fatty Acid Composition in Castorbean Varieties. J. Am. Oil Chem. Soc. 1984, 61, 1841–1843. [Google Scholar] [CrossRef]
- Ogunniyi, D.S. Castor Oil: A Vital Industrial Raw Material. Bioresour. Technol. 2006, 97, 1086–1091. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, H.; Meier, M.A.R. Castor Oil as a Renewable Resource for the Chemical Industry. Eur. J. Lipid Sci. Technol. 2010, 112, 10–30. [Google Scholar] [CrossRef]
- Tumer, N. Ricin Toxins. Toxins 2020, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Nagler-Anderson, C. Man the Barrier! STRATEGIC DEFENCES in the Intestinal Mucosa. Nat. Rev. Immunol. 2001, 1, 59–67. [Google Scholar] [CrossRef]
- Lappi, D.A.; Kapmeyer, W.; Beglau, J.M.; Kaplan, N.O. The Disulfide Bond Connecting the Chains of Ricin. Proc. Natl. Acad. Sci. USA 1978, 75, 1096. [Google Scholar] [CrossRef]
- Endo, Y.; Tsurugi, K. RNA N-Glycosidase Activity of Ricin A-Chain. Mechanism of Action of the Toxic Lectin Ricin on Eukaryotic Ribosomes. J. Biol. Chem. 1987, 262, 8128–8130. [Google Scholar] [CrossRef] [PubMed]
- Parikh, B.A.; Tortora, A.; Li, X.P.; Tumer, N.E. Ricin Inhibits Activation of the Unfolded Protein Response by Preventing Splicing of the HAC1 mRNA. J. Biol. Chem. 2008, 283, 6145–6153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsnes, S.; Fernandez-Puentes, C.; Carrasco, L.; Vazquez, D. Ribosome Inactivation by the Toxic Lectins Abrin and Ricin. Kinetics of the Enzymic Activity of the Toxin A-Chains. Eur. J. Biochem. 1975, 60, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Severino, L.S.; Auld, D.L.; Baldanzi, M.; Cândido, M.J.D.; Chen, G.; Crosby, W.; Tan, D.; He, X.; Lakshmamma, P.; Lavanya, C.; et al. A Review on the Challenges for Increased Production of Castor. Agron. J. 2012, 104, 853–880. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Pan, C.; Fan, W.; Liu, W.; Zhao, H.; Li, D.; Wang, S.; Hu, L.; He, B.; Qian, K.; et al. A Chromosome-Level Genome Assembly of Wild Castor Provides New Insights into Its Adaptive Evolution in Tropical Desert. Genom. Proteom. Bioinform. 2022, 20, 42–59. [Google Scholar] [CrossRef]
- Ready, M.P.; Kim, Y.; Robertus, J.D. Site-Directed Mutagenesis of Ricin A-Chain and Implications for the Mechanism of Action. Proteins Struct. Funct. Bioinform. 1991, 10, 270–278. [Google Scholar] [CrossRef]
- Madeira, J.V.; Macedo, J.A.; Macedo, G.A. Detoxification of Castor Bean Residues and the Simultaneous Production of Tannase and Phytase by Solid-State Fermentation Using Paecilomyces variotii. Bioresour. Technol. 2011, 102, 7343–7348. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, K.V.; Deus-de-Oliveira, N.; Godoy, M.G.; Guimarães, Z.A.S.; Nascimento, V.V.; de Melo, E.J.T.; Freire, D.M.G.; Dansa-Petretski, M.; Machado, O.L.T. Simultaneous Allergen Inactivation and Detoxification of Castor Bean Cake by Treatment with Calcium Compounds. Braz. J. Med. Biol. Res. 2012, 45, 1002. [Google Scholar] [CrossRef] [Green Version]
- Harley, S.M.; Beevers, H. Lectins in Castor Bean Seedlings. Plant Physiol. 1986, 80, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Sotelo-Leyva, C.; Salinas-Sánchez, D.O.; Peña-Chora, G.; Trejo-Loyo, A.G.; González-Cortázar, M.; Zamilpa, A. Insecticidal Compounds in Ricinus communis L. (Euphorbiaceae) to Control Melanaphis sacchari Zehntner (Hemiptera: Aphididae). Fla. Entomol. 2020, 103, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Salem, N.; Bachrouch, O.; Sriti, J.; Msaada, K.; Khammassi, S.; Hammami, M.; Selmi, S.; Boushih, E.; Koorani, S.; Abderraba, M.; et al. Fumigant and Repellent Potentials of Ricinus communis and Mentha pulegium Essential Oils against Tribolium castaneum and Lasioderma serricorne. Int. J. Food Prop. 2018, 20, S2899–S2913. [Google Scholar] [CrossRef] [Green Version]
- Salinas-Sánchez, D.O.; Flores-Franco, G.; Aviles-Montes, D.; Valladares-Cisneros, M.G.; Ataide, D.M.A.; Mendoza-Catalan, M.A.; Sotelo-Leyva, C. Bioactivity of a Linoleic Acid-Rich Fraction of Ricinus communis L. (Euphorbiaceae) Leaves against the Yellow Sugarcane Aphid, Sipha flava (Hemiptera: Aphididae). J. Food Prot. 2021, 84, 1524–1527. [Google Scholar] [CrossRef]
- Elhaj, W.E.; Osman, A.A.; Elawad, L.M.E. Efficacy of Ricinus communis L., Cassia occidentalis L. and Bacillus thuringiensis against Helicoverpa armigera Hübner (Lepidoptera: Noctuidae). J. Agron. Res. 2021, 3, 46–53. [Google Scholar] [CrossRef]
- Kouakou, Y.Y.F.R.; Kra, K.D.; Diallo, H.A. Effects of Two-Dried Castor Leaf Formulations on the Population Dynamics and Pathological Activities of Root-Lesion and Root-Knot Nematodes on Water Yam. Int. J. Plant Soil Sci. 2023, 35, 33–41. [Google Scholar] [CrossRef]
- Yao, Y.; Regis, F.; Kra, K.D.; Assiri, K.P. Nematicidal Effectiveness of Products Stemming from Dried Leaves of Castor-Oil Plant (Ricinus communis L.) on Meloidogyne and Pratylenchus, Yam Pathogenic Nematodes in Côte d’ Ivoire. Int. J. Agron. Agric. Res. 2017, 11, 57–68. [Google Scholar]
- Izidoro, A., Jr.; Silva, E.J.; Tarini, G.; Bordin, J.C.; Silva, B.A.; Ambrosano, L.; Dias-Arieira, C.R. Aqueous Extract of Castor Bean Seed Cake for the Control of Pratylenchus Brachyurus in Soybean. Nematropica 2021, 51, 1–8. [Google Scholar]
- Adomako, J.; Kwoseh, C. Effect of Castor Bean (Ricinus communis L.) Aqueous Extracts on the Performance of Root-Knot Nematodes (Meloideogyne spp.) on Tomato (Solanum lycopersicum L.). J. Sci. Technol. 2013, 33, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Budavari, S.; O’Neil, M.J.; Smith, A.; Heckelman, P.A.; Kinneary, J.F. The Merck Index: An Encyclopedia of Chemicals, Drugs, and Biologicals. J. Hazard. Mater. 1996, 30, 373. [Google Scholar]
- Oprea, S. Synthesis and Properties of Polyurethane Elastomers with Castor Oil as Crosslinker. JAOCS J. Am. Oil Chem. Soc. 2010, 87, 313–320. [Google Scholar] [CrossRef]
- Yao, L.; Hammond, E.G.; Wang, T.; Bhuyan, S.; Sundararajan, S. Synthesis and Physical Properties of Potential Biolubricants Based on Ricinoleic Acid. JAOCS J. Am. Oil Chem. Soc. 2010, 87, 937–945. [Google Scholar] [CrossRef]
- Zeng, Q. The Lubrication Performance and Viscosity Behavior of Castor Oil under High Temperature. Green Mater. 2021, 10, 51–58. [Google Scholar] [CrossRef]
- Sen, B.; Gupta, M.K.; Mia, M.; Pimenov, D.Y.; Mikolajczyk, T. Performance Assessment of Minimum Quantity Castor-Palm Oil Mixtures in Hard-Milling Operation. Materials 2021, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- González, M.; Gallego, R.; Romero, M.A.; González-Delgado, J.A.; Arteaga, J.F.; Valencia, C.; Franco, J.M. Impact of Natural Sources-Derived Antioxidants on the Oxidative Stability and Rheological Properties of Castor Oil Based-Lubricating Greases. Ind. Crop. Prod. 2016, 87, 297–303. [Google Scholar] [CrossRef]
- Morris, B. Phytochemical Traits in the Genetic Resources of Castor Bean. Curr. Top. Plant Biol. 2004, 5, 63–67. [Google Scholar]
- Pecina-Quintero, V.; Anaya-López, J.L.; Núñez-Colín, C.A.; Zamarripa-Colmenero, A.; Montes-García, N.; Solís-Bonilla, J.L.; Aguilar-Rangel, M.R. Assessing the Genetic Diversity of Castor Bean from Chiapas, México Using SSR and AFLP Markers. Ind. Crop. Prod. 2013, 41, 134–143. [Google Scholar] [CrossRef]
- Gowda, N.K.S.; Pal, D.T.; Bellur, S.R.; Bharadwaj, U.; Sridhar, M.; Satyanarayana, M.L.; Prasad, C.S.; Ramachandra, K.S.; Sampath, K.T. Evaluation of Castor (Ricinus communis) Seed Cake in the Total Mixed Ration for Sheep. J. Sci. Food Agric. 2009, 89, 216–220. [Google Scholar] [CrossRef]
- Diniz, L.L.; Valadares Filho, S.C.; Campos, J.M.S.; Valadares, R.F.D.; Da Silva, L.D.; Monnerat, J.P.I.S.; Benedeti, P.B.; De Oliveira, A.S.; Pina, D.S. Effects of Castor Meal on the Growth Performance and Carcass Characteristics of Beef Cattle. Asian-Australas. J. Anim. Sci. 2010, 23, 1308–1318. [Google Scholar] [CrossRef]
- Diniz, L.L.; Filho, S.C.V.; de Oliveira, A.S.; Pina, D.S.; da Silva, L.D.; Benedeti, P.B.; Baião, G.F.; Campos, J.M.S.; Valadares, R.F.D. Castor Bean Meal for Cattle Finishing: 1—Nutritional Parameters. Livest. Sci. 2011, 135, 153–167. [Google Scholar] [CrossRef]
- Gupta, A.P.; Antil, R.S.; Narwal, R.P. Utilization of Deoiled Castor Cake for Crop Production. Arch. Agron. Soil Sci. 2006, 50, 389–395. [Google Scholar] [CrossRef]
- Lima, R.L.S.; Severino, L.S.; Sampaio, L.R.; Sofiatti, V.; Gomes, J.A.; Beltrão, N.E.M. Blends of Castor Meal and Castor Husks for Optimized Use as Organic Fertilizer. Ind. Crop. Prod. 2011, 33, 364–368. [Google Scholar] [CrossRef] [Green Version]
- Lima, R.D.L.S.D.; Severino, L.S.; Silva, M.I.D.L.; Vale, L.S.D.; Beltrão, N.E.D.M. Recipients Volume and Substrate Composition for Castor Seedlings Production. Web Sci. 2006, 30, 480–486. [Google Scholar] [CrossRef]
- Lima, R.D.L.S.D.; Severino, L.S.; Silva, M.I.D.L.; Jerônimo, J.F.; Vale, L.S.D.; Beltrão, N.E.D.M. Substrates for Castor Seedlings Production Composed by Blends of Five Organic Materials. Web Sci. 2006, 30, 474–479. [Google Scholar] [CrossRef]
- Milfont, M.; Freitas, B.; Rizzardo, R.; Guimarães, M. Honey Production by Africanized Honey Bees in Castor Bean Cropping. Cencia Rural 2009, 39, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- Romeiro, S.; Lagôa, A.M.M.A.; Furlani, P.R.; De Abreu, C.A.; De Abreu, M.F.; Erismann, N.M. Lead Uptake and Tolerance of Ricinus communis L. Braz. J. Plant Physiol. 2006, 18, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Cai, Q. Zinc Tolerance and Accumulation in Eight Oil Crops. J. Plant Nutr. 2010, 33, 982–997. [Google Scholar] [CrossRef]
- Niu, Z.X.; Sun, L.N.; Sun, T.H.; Li, Y.S.; Hong, W. Evaluation of Phytoextracting Cadmium and Lead by Sunflower, Ricinus, Alfalfa and Mustard in Hydroponic Culture. J. Environ. Sci. 2007, 19, 961–967. [Google Scholar] [CrossRef]
- Zeng, F.; Mallhi, Z.I.; Khan, N.; Rizwan, M.; Ali, S.; Ahmad, A.; Hussain, A.; Alsahli, A.A.; Alyemeni, M.N. Combined Citric Acid and Glutathione Augments Lead (Pb) Stress Tolerance and Phytoremediation of Castorbean through Antioxidant Machinery and pb Uptake. Sustainability 2021, 13, 4073. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Pivetta, L.A. Mechanical and Biological Approaches to Alleviate Soil Compaction in Tropical Soils: Assessed by Root Growth and Activity (rb Uptake) of Soybean and Maize Grown in Rotation with Cover Crops. Soil Use Manag. 2017, 33, 141–152. [Google Scholar] [CrossRef]
- Orji, O.A.; Eke, I.P. Effect of Mulch Materials on Soil Physico-Chemical Properties and the Performance of Castor Bean Plant (Ricinus communis) in Rivers State, Nigeria. Indian J. Anim. Res. 2018, 52, 649–654. [Google Scholar] [CrossRef]
- Zhang, D.M.; Xu, H.G.; Wang, L.; Li, Y.J.; Sun, P.H.; Wu, X.M.; Wang, G.J.; Chen, W.M.; Ye, W.C. Betulinic Acid and Its Derivatives as Potential Antitumor Agents. Med. Res. Rev. 2015, 35, 1127–1155. [Google Scholar] [CrossRef]
- Saleem, M. Lupeol, a Novel Anti-Inflammatory and Anti-Cancer Dietary Triterpene. Cancer Lett. 2009, 285, 109–115. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 10 May 2023).
- Mubofu, E.B. Castor Oil as a Potential Renewable Resource for the Production of Functional Materials. Sustain. Chem. Process. 2016, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Neto, S.S.; De Paula Manjavachi, M.K.; Zeffa, D.M.; Sartori, M.M.P.; Zanotto, M.D. Morphological Characterization and Selection of Castor Bean Accessions for Mechanized Production. Pesqui. Agropecu. Trop. 2019, 49, 1–9. [Google Scholar] [CrossRef]
- Latterini, F.; Stefanoni, W.; Cavalaris, C.; Karamoutis, C.; Pari, L.; Alexopoulou, E. Effectiveness of Three Terminating Products on Reducing the Residual Moisture in Dwarf Castor Plants: A Preliminary Study of Direct Mechanical Harvesting in Central Greece. Agronomy 2022, 12, 146. [Google Scholar] [CrossRef]
- Stefanoni, W.; Latterini, F.; Malkogiannidis, V.; Salpiggidis, V.; Alexopoulou, E.; Pari, L. Mechanical Harvesting of Castor Bean (Ricinus communis L.) with a Combine Harvester Equipped with Two Different Headers: A Comparison of Working Performance. Energies 2022, 15, 2999. [Google Scholar] [CrossRef]
- Canecchio Filho, V.; Freire, E.S. Adubação da Mamoneira: I—Experiências Preliminares. Bragantia 1958, 17, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Manickam, S.; Kalaiselvan, P.; Subramaniyan, K.; Venkatachalam, S.R. Role of Herbicide in Castor Based Intercropping System. J. Phytol. Res. 2009, 22, 291–294. [Google Scholar]
- Sofiatti, V.; Severino, L.S.; Silva, F.M.O.; Silva, V.N.B.; Brito, G.G. Pre and Postemergence Herbicides for weed Control in Castor Crop. Ind. Crop. Prod. 2012, 37, 235–237. [Google Scholar] [CrossRef]
- Soares, J.D. Gray Mold of Castor: A Review. In Plant Pathology; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Manjula, K.; Sarma, P.S.; Nageshwar Rao, T.; Thatikunta, R. In Vitro Screening of Castor Genotypes for Stress Tolerance Seed Physiology View Project Horticulture View Project. J. Oilseeds Res. 2001, 18, 292–293. [Google Scholar]
- Lakshmamma, P.; Lakshmi, P.; Lavanya, C.; Anjani, K. Growth and Yield of Different Castor Genotypes Varying in Drought Tolerance. Ann. Arid Zone 2009, 48, 35–39. [Google Scholar]
- Zhong, W.; Hartung, W.; Komor, E.; Schobert, C. Phloem Transport of Abscisic Acid in Ricinus communis L. Seedlings. Plant Cell Environ. 1996, 19, 471–477. [Google Scholar] [CrossRef]
- Babita, M.; Maheswari, M.; Rao, L.M.; Shanker, A.K.; Rao, D.G. Osmotic Adjustment, Drought Tolerance and Yield in Castor (Ricinus communis L.) Hybrids. Environ. Exp. Bot. 2010, 69, 243–249. [Google Scholar] [CrossRef]
- Sausen, T.L.; Rosa, L.M.G. Growth and Limitations to Carbon Assimilation in Ricinus communis (Euphorbiaceae) under Soil Water Stress Conditions. Acta Bot. Bras. 2010, 24, 648–654. [Google Scholar] [CrossRef]
- Funk, J.L.; Zachary, V.A. Physiological Responses to Short-Term Water and Light Stress in Native and Invasive Plant Species in Southern California. Biol. Invasions 2010, 12, 1685–1694. [Google Scholar] [CrossRef]
- Pinheiro, H.A.; Silva, J.V.; Endres, L.; Ferreira, V.M.; de Albuquerque Câmara, C.; Cabral, F.F.; Oliveira, J.F.; de Carvalho, L.W.T.; dos Santos, J.M.; dos Santos Filho, B.G. Leaf Gas Exchange, Chloroplastic Pigments and Dry Matter Accumulation in Castor Bean (Ricinus communis L.) Seedlings Subjected to Salt Stress Conditions. Ind. Crop. Prod. 2008, 27, 385–392. [Google Scholar] [CrossRef]
- Silva, S.M.S.; Alves, A.N.; Gheyi, H.R.; Beltrão, N.E.D.M.; Severino, L.S.; Soares, F.A.L. Growth and Production of Two Cultivars Of Castor Bean under Saline Stress|Desenvolvimento e Producao de Duas Cultivares de Mamoneira Sob Estresse Salino. Rev. Bras. Eng. Agric. Ambient. 2008, 12, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wan, S.; Zhou, J.; Yang, Z.; Qin, P. Leaf Chlorophyll Fluorescence, Hyperspectral Reflectance, Pigments Content, Malondialdehyde and Proline Accumulation Responses of Castor Bean (Ricinus communis L.) Seedlings to Salt Stress Levels. Ind. Crop. Prod. 2010, 31, 13–19. [Google Scholar] [CrossRef]
- Else, M.A.; Coupland, D.; Dutton, L.; Jackson, M.B. Decreased Root Hydraulic Conductivity Reduces Leaf Water Potential, Initiates Stomatal Closure and Slows Leaf Expansion in Flooded Plants of Castor Oil (Ricinus communis) Despite Diminished Delivery of ABA from the Roots to Shoots in Xylem Sap. Physiol. Plant. 2001, 111, 46–54. [Google Scholar] [CrossRef]
- Lourdes, R.D.E.; Lima, S.D.E.; Severino, L.I.V.S.; Ferreira, G.B. Soil Exchangeable Aluminum Influencing the Growth and Leaf Tissue Macronutrients Content of Castor Plants. Rev. Caatinga 2014, 27, 10–15. [Google Scholar]
- de Freitas, L.B.; Fernandes, D.M.; Maia, S.C.M.; Pivetta, L.G.; Zanotto, M.D. Aluminum Tolerance in Castor Bean Lines. Pesqui. Agropecu. Trop. 2018, 48, 299–305. [Google Scholar] [CrossRef]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft Genome Sequence of the Oilseed Species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinkerton, S.D.; Rolfe, R.; Auld, D.L.; Ghetie, V.; Lauterbach, B.F. Selection of Castor for Divergent Concentrations of Ricin and Ricinus communis Agglutinin. Crop Sci. 1999, 39, 353–357. [Google Scholar] [CrossRef]
- Baldanzi, M.; Pugliesi, C. Selection for Non-Branching in Castor, Ricinus communis L. Plant Breed. 1998, 117, 392–394. [Google Scholar] [CrossRef]
- Baldanzi, M.; Fambrini, M.; Pugliesi, C. Redesign of the Castorbean Plant Body Plan for Optimal Combine Harvesting. Ann. Appl. Biol. 2003, 142, 299–306. [Google Scholar] [CrossRef]
- Anjani, K. Castor Genetic Resources: A Primary Gene Pool for Exploitation. Ind. Crop. Prod. 2012, 35, 1–14. [Google Scholar] [CrossRef]
- Anjani, K.; Raoof, M.A.; Prasad, M.S.L.; Duraimurugan, P.; Lucose, C.; Yadav, P.; Prasad, R.D.; Lal, J.J.; Sarada, C. Trait-Specific Accessions in Global Castor (Ricinus communis L.) Germplasm Core Set for Utilization in Castor Improvement. Ind. Crop. Prod. 2018, 112, 766–774. [Google Scholar] [CrossRef]
- dos S. de Oliveira, F.; Bassegio, D.; Ramos, A.R.; da Silva, G.H.; da Silva, J.; Zanotto, M.D.; Fernandes, D.M. Phosphate Fertilization of Low Size Castor Bean in Conventional and Narrow Cultivation in Second Cropping Season. J. Exp. Agric. Int. 2019, 29, 1–11. [Google Scholar] [CrossRef]
- da Silva, J.; Ramos, A.R.; Amorim, D.J.; Zanotto, M.D.; Sartori, M.M.P. Productive Potential and Castor Bean Selection of the FCA-PB Cultivar Progenies. Rev. Ceres 2020, 67, 42–51. [Google Scholar] [CrossRef]
- Severino, L.S.; Auld, D.L.; Vale, L.S.; Marques, L.F. Plant Density Does Not Influence Every Castor Plant Equally. Ind. Crop. Prod. 2017, 107, 588–594. [Google Scholar] [CrossRef]
- Rojas-Barros, P.; De Haro, A.; Muñoz, J.; Fernández-Martínez, J.M. Isolation of a Natural Mutant in Castor with High Oleic/Low Ricinoleic Acid Content in the Oil. Crop Sci. 2004, 44, 76–80. [Google Scholar] [CrossRef]
- Venegas-Calerón, M.; Sánchez, R.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Molecular and Biochemical Characterization of the OLE-1 High-Oleic Castor Seed (Ricinus communis L.) Mutant. Planta 2016, 244, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, C.N.; Hendon, B.R.; Mishra, D.; Rieff, J.M.; Lowery, C.C.; Lambert, K.C.; Witt, T.W.; Oswalt, S.J.; Bechere, E.; Smith, C.W.; et al. Cotton (Gossypium hirsutum L.) Mutants with Reduced Levels of Palmitic Acid (C16:0) in Seed Lipids. Euphytica 2019, 215, 112. [Google Scholar] [CrossRef]
- Hendon, B.R.; Bechere, E.; Witt, T.W.; Kelly, B.R.; Mishra, D.; Auld, D.L. Genetic Improvement of Naked-Tufted Seed Mutants in Upland Cotton (Gossypium hirsutum L.). Euphytica 2019, 215, 81. [Google Scholar] [CrossRef]
- Bechere, E.; Zeng, L.; Auld, D. Registration of Five Upland Cotton Mutant Germplasm Lines with Superior Fiber Length, Strength, and Uniformity. J. Plant Regist. 2018, 12, 107–111. [Google Scholar] [CrossRef]
- Percy, R.; Hendon, B.; Bechere, E.; Auld, D. Qualitative Genetics and Utilization of Mutants. Cotton 2015, 57, 155–185. [Google Scholar] [CrossRef]
- Brown, N.; Wayne Smith, C.; Hague, S.; Auld, D.; Hequet, E.; Joy, K.; Jones, D. Within-Boll Yield Characteristics and Their Correlation with Fiber Quality Parameters Following Mutagenesis of Upland Cotton, TAM 94L-25. Crop Sci. 2015, 55, 1513–1523. [Google Scholar] [CrossRef]
- Lavanya, C.; Gopinath, V. Inheritance Studies for Morphological Characters and Sex Expression in Pistillate Lines of Castor (Ricinus communis L.). Indian J. Genet. Plant Breed. 2008, 68, 275–282. [Google Scholar]
- Ahmed, H.M.; Sarwar, G.; ul Haq, M.A. Genetic Variability and Interdependence of Morphological Traits in Castorbean (Ricinus communis L) Mutants. Songklanakarin J. Sci. Technol. 2012, 34, 279–286. [Google Scholar]
- Allan, G.; Williams, A.; Rabinowicz, P.D.; Chan, A.P.; Ravel, J.; Keim, P. Worldwide Genotyping of Castor Bean Germplasm (Ricinus communis L.) Using AFLPs and SSRs. Genet. Resour. Crop Evol. 2008, 55, 365–378. [Google Scholar] [CrossRef]
- Milani, M.; de Medeiros Nbreg, M.B. Castor Breeding. In Plant Breeding from Laboratories to Fields; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Moshkin, V.A.; Dvoryadkina, A.G. Cytology and Genetics of Qualitative Characteristics; Castor: New Delhi, India, 1986. [Google Scholar]
- Severino, L.S. Single-Seed Selection of Fast-Germinating Genotypes of Castor (Ricinus communis). Ind. Crop. Prod. 2023, 194, 116307. [Google Scholar] [CrossRef]
- Hu, W.; Chen, L.; Qiu, X.; Lu, H.; Wei, J.; Bai, Y.; He, N.; Hu, R.; Sun, L.; Zhang, H.; et al. Morphological, Physiological and Proteomic Analyses Provide Insights into the Improvement of Castor Bean Productivity of a Dwarf Variety in Comparing with a High-Stalk Variety. Front. Plant Sci. 2016, 7, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, E.; Kurczyńska, E.U.; Friml, J. Cellular Events during Interfascicular Cambium Ontogenesis in Inflorescence Stems of Arabidopsis. Protoplasma 2014, 251, 1125–1139. [Google Scholar] [CrossRef] [PubMed]
- Kebrom, T.H. A Growing Stem Inhibits Bud Outgrowth—The Overlooked Theory of Apical Dominance. Front. Plant Sci. 2017, 8, 1874. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A Role for Flavin Monooxygenase-like Enzymes in Auxin Biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, A.; Li, F.; Xu, W.; Han, B.; Cheng, X.; Liu, A. Bulked Segregant Analysis Reveals Candidate Genes Responsible for Dwarf Formation in Woody Oilseed Crop Castor Bean. Sci. Rep. 2021, 11, 6277. [Google Scholar] [CrossRef]
- Mori, K.K.; Patel, J.; Rani, K.; Mori, V.K.; Kumar, M.; Ajay, B.C. Deciphering Higher Order Non-Allelic Interactions for Quantitative Characters through Twelve Generation Mean Analysis in Castor (Ricinus communis L.). Genet. Resour. Crop Evol. 2022, 69, 1759–1785. [Google Scholar] [CrossRef]
- Solanki, S.S.; Deora, V.S.; Singh, D.P. Combining Ability of New Castor (Ricinus communis L.) Pistillate Lines: MCP-1-1. J. Oilseeds Res. 2004, 21, 274–276. [Google Scholar]
- Golakia, P.R.; Poshiya, V.K.; Monpara, B.A. Identification of Superior Donor Parents for Earliness through Combining Ability in Castor. Int. J. Res. Plant Sci. 2015, 5, 26–31. [Google Scholar]
- Virani, H.P.; Dhedhi, K.K.; Dhaduk, H.L. Gene Effects of Seed Yield and Component Traits in Castor Crosses. PKV Res. J. 2013, 37, 8–13. [Google Scholar]
- Lavanya, C.; Chandramohan, Y. Combining Ability and Heterosis for Seed Yield and Yield Components in Castor. J. Oilseeds Res. 2003, 20, 220–224. [Google Scholar]
- Ramana, P.V.; Lavanya, C.; Ratnasree, P. Combining Ability and Heterosis Studies under Rainfed Conditions in Castor (Ricinus communis L.). Indian J. Genet. Plant Breed. 2005, 65, 325–326. [Google Scholar]
- Lavanya, C.; Solanki, S.S. Crop Improvement of Castor. The Challenges Ahead. Research and Development in Castor. Present Status and Future Strategies. Indian Soc. Oilseeds Res. Hyderabad 2010, 36–55. Available online: https://scholar.google.co.in/citations?view_op=view_citation&hl=en&user=8KzYu1cAAAAJ&citation_for_view=8KzYu1cAAAAJ:hC7cP41nSMkC (accessed on 14 May 2023).
- Sailaja, M.; Tarakeswari, M.; Sujatha, M. Stable Genetic Transformation of Castor (Ricinus communis L.) via Particle Gun-Mediated Gene Transfer Using Embryo Axes from Mature Seeds. Plant Cell Rep. 2008, 27, 1509–1519. [Google Scholar] [CrossRef]
- Bertozzo, F.; Machado, I.S. Meios de Cultura No Desenvolvimento de Ápices Caulinares de Mamoneira (Ricinus communis L.) In Vitro. Ciência Agrotecnol. 2010, 34, 1477–1482. [Google Scholar] [CrossRef] [Green Version]
- Sujatha, M.; Sailaja, M. Stable Genetic Transformation of Castor (Ricinus communis L.) via Agrobacterium Tumefaciens-Mediated Gene Transfer Using Embryo Axes from Mature Seeds. Plant Cell Rep. 2005, 23, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhou, B.; Yang, Y.; Mei, J.; Zhao, X.; Guo, X.; Huang, X.; Tang, D.; Liu, X. Piercing and Vacuum Infiltration of the Mature Embryo: A Simplified Method for Agrobacterium-Mediated Transformation of Indica Rice. Plant Cell Rep. 2009, 28, 1065–1074. [Google Scholar] [CrossRef]
- Sánchez-Álvarez, A.; Ruíz-López, N.; Moreno-Pérez, A.J.; Martínez-Force, E.; Garcés, R.; Salas, J.J. Agrobacterium-Mediated Transient Gene Expression in Developing Ricinus communis Seeds: A First Step in Making the Castor Oil Plant a Chemical Biofactory. Front. Plant Sci. 2019, 10, 1410. [Google Scholar]
- Auld, D.L.; Pinkerton, S.D.; Boroda, E.; Lombard, K.A.; Murphy, C.K.; Kenworthy, K.E.; Becker, W.D.; Rolfe, R.D.; Ghetie, V. Registration of TTU-LRC Castor Germplasm with Reduced Levels of Ricin and RCA120. Crop Sci. 2003, 43, 746–747. [Google Scholar] [CrossRef]
- Auld, D.L.; Rolfe, R.D.; McKeon, T.A. Development of Castor with Reduced Toxicity. J. New Seeds 2008, 3, 61–69. [Google Scholar] [CrossRef]
- Sousa, N.L.; Cabral, G.B.; Vieira, P.M.; Baldoni, A.B.; Aragão, F.J.L. Bio-Detoxification of Ricin in Castor Bean (Ricinus communis L.) Seeds. Sci. Rep. 2017, 7, 15385. [Google Scholar] [CrossRef] [Green Version]
- Guhling, O.; Hobl, B.; Yeats, T.; Jetter, R. Cloning and Characterization of a Lupeol Synthase Involved in the Synthesis of Epicuticular Wax Crystals on Stem and Hypocotyl Surfaces of Ricinus communis. Arch. Biochem. Biophys. 2006, 448, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, R.; Lu, W.; Zhou, Z.; Jiang, X.; Zhao, H.; Yang, B.; Lü, S. Transcriptome Analysis Identifies Key Genes Involved in the Regulation of Epidermal Lupeol Biosynthesis in Ricinus communis. Ind. Crop. Prod. 2021, 160, 113100. [Google Scholar] [CrossRef]
- Li, D.; Pan, C.; Lu, J.; Zaman, W.; Zhao, H.; Zhang, J.; Lü, S. Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor. Molecules 2021, 26, 2978. [Google Scholar] [CrossRef] [PubMed]
- Maiorino, F.M.; Brigelius-Flohé, R.; Aumann, K.D.; Roveri, A.; Schomburg, D.; Flohé, L. Diversity of glutathione peroxidases. Methods Enzymol. 1995, 252, 38–48. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant Glutathione Peroxidases: Emerging Role of the Antioxidant Enzymes in Plant Development and Stress Responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Passaia, G.; Margis-Pinheiro, M. Glutathione Peroxidases as Redox Sensor Proteins in Plant Cells. Plant Sci. 2015, 234, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Roxas, V.P.; Lodhi, S.A.; Garrett, D.K.; Mahan, J.R.; Allen, R.D. Stress Tolerance in Transgenic Tobacco Seedlings That Overexpress Glutathione S-Transferase/Glutathione Peroxidase. Plant Cell Physiol. 2000, 41, 1229–1234. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Miyao, K.; Gaber, A.; Takeda, T.; Kanaboshi, H.; Miyasaka, H.; Shigeoka, S. Enhancement of Stress Tolerance in Transgenic Tobacco Plants Overexpressing Chlamydomonas Glutathione Peroxidase in Chloroplasts or Cytosol. Plant J. 2004, 37, 21–33. [Google Scholar] [CrossRef]
- Navrot, N.; Collin, V.; Gualberto, J.; Gelhaye, E.; Hirasawa, M.; Rey, P.; Knaff, D.B.; Issakidis, E.; Jacquot, J.P.; Rouhier, N. Plant Glutathione Peroxidases Are Functional Peroxiredoxins Distributed in Several Subcellular Compartments and Regulated during Biotic and Abiotic Stresses. Plant Physiol. 2006, 142, 1364–1379. [Google Scholar] [CrossRef] [Green Version]
- Passaia, G.; Spagnolo Fonini, L.; Caverzan, A.; Jardim-Messeder, D.; Christoff, A.P.; Gaeta, M.L.; de Araujo Mariath, J.E.; Margis, R.; Margis-Pinheiro, M. The Mitochondrial Glutathione Peroxidase GPX3 Is Essential for H2O2 Homeostasis and Root and Shoot Development in Rice. Plant Sci. 2013, 208, 93–101. [Google Scholar] [CrossRef]
- Kim, Y.J.; Jang, M.G.; Noh, H.Y.; Lee, H.J.; Sukweenadhi, J.; Kim, J.H.; Kim, S.Y.; Kwon, W.S.; Yang, D.C. Molecular Characterization of Two Glutathione Peroxidase Genes of Panax Ginseng and Their Expression Analysis against Environmental Stresses. Gene 2014, 535, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, T.; Manna, M.; Kaul, T.; Pandey, S.; Reddy, C.S.; Reddy, M.K. Genome-Wide Dissection of Arabidopsis and Rice for the Identification and Expression Analysis of Glutathione Peroxidases Reveals Their Stress-Specific and Overlapping Response Patterns. Plant Mol. Biol. Report. 2015, 33, 1413–1427. [Google Scholar] [CrossRef]
- Wang, X.; Liu, X.; An, Y.Q.C.; Zhang, H.; Meng, D.; Jin, Y.; Huo, H.; Yu, L.; Zhang, J. Identification of Glutathione Peroxidase Gene Family in Ricinus communis and Functional Characterization of RcGPX4 in Cold Tolerance. Front. Plant Sci. 2021, 12, 707127. [Google Scholar] [CrossRef]
- Li, B.; He, L.; Guo, S.; Li, J.; Yang, Y.; Yan, B.; Sun, J.; Li, J. Proteomics Reveal Cucumber Spd-Responses under Normal Condition and Salt Stress. Plant Physiol. Biochem. PPB 2013, 67, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Xu, J.; Chen, T.; Tao, A.; Qi, J. Proteomic Changes in Kenaf (Hibiscus cannabinus L.) Leaves under Salt Stress. Ind. Crop. Prod. 2016, 91, 255–263. [Google Scholar] [CrossRef]
- Veeranagamallaiah, G.; Jyothsnakumari, G.; Thippeswamy, M.; Chandra Obul Reddy, P.; Surabhi, G.K.; Sriranganayakulu, G.; Mahesh, Y.; Rajasekhar, B.; Madhurarekha, C.; Sudhakar, C. Proteomic Analysis of Salt Stress Responses in Foxtail Millet (Setaria italica L. cv. Prasad) Seedlings. Plant Sci. 2008, 175, 631–641. [Google Scholar] [CrossRef]
- Wang, Y.; Peng, X.; Salvato, F.; Wang, Y.; Yan, X.; Zhou, Z.; Lin, J. Salt-Adaptive Strategies in Oil Seed Crop Ricinus communis Early Seedlings (Cotyledon vs. True Leaf) Revealed from Proteomics Analysis. Ecotoxicol. Environ. Saf. 2019, 171, 12–25. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Yang, F.; Zhou, W.; Mao, S.; Lin, J.; Yan, X. Untargeted LC-MS-Based Metabolomics Revealed Specific Metabolic Changes in Cotyledons and Roots of Ricinus communis during Early Seedling Establishment under Salt Stress. Plant Physiol. Biochem. 2021, 163, 108–118. [Google Scholar] [CrossRef]
- Djakovic-Petrovic, T.; De Wit, M.; Voesenek, L.A.C.J.; Pierik, R. DELLA Protein Function in Growth Responses to Canopy Signals. Plant J. 2007, 51, 117–126. [Google Scholar] [CrossRef]
- Lei, P.; Liu, Z.; Hu, Y.; Kim, H.C.; Liu, S.; Liu, J.; Xu, L.; Li, J.; Zhao, Y.; Yu, Z.; et al. Transcriptome Analysis of Salt Stress Responsiveness in the Seedlings of Wild and Cultivated Ricinus communis L. J. Biotechnol. 2021, 327, 106–116. [Google Scholar] [CrossRef]
- Jiang, X.J.; Wen, X. The Effect of Temperature on the Germination Rates on Castor Bean. Seeds 2008, 27, 67–69. [Google Scholar]
- Wang, X.; Li, M.; Liu, X.; Zhang, L.; Duan, Q.; Zhang, J. Quantitative Proteomic Analysis of Castor (Ricinus communis L.) Seeds during Early Imbibition Provided Novel Insights into Cold Stress Response. Int. J. Mol. Sci. 2019, 20, 355. [Google Scholar] [CrossRef] [Green Version]
- Ghidoli, M.; Frazzini, S.; De Benedetti, S.; Sangiorgio, S.; Landoni, M.; Scarafoni, A.; Rossi, L.; Pilu, R. Genetic Improvement of Camelina sativa (L.) Crantz: Opportunities and Challenges. Agronomy 2023, 12, 1562. [Google Scholar] [CrossRef]
- Al Amin, N.; Ahmad, N.; Wu, N.; Pu, X.; Ma, T.; Du, Y.; Bo, X.; Wang, N.; Sharif, R.; Wang, P. CRISPR-Cas9 Mediated Targeted Disruption of FAD2-2 Microsomal Omega-6 Desaturase in Soybean (Glycine max.L). BMC Biotechnol. 2019, 19, 9. [Google Scholar] [CrossRef]
- Wang, J.; Kuang, H.; Zhang, Z.; Yang, Y.; Yan, L.; Zhang, M.; Song, S.; Guan, Y. Generation of Seed Lipoxygenase-Free Soybean Using CRISPR-Cas9. Crop J. 2020, 8, 432–439. [Google Scholar] [CrossRef]
- Ma, J.; Sun, S.; Whelan, J.; Shou, H. CRISPR/Cas9-Mediated Knockout of GmFATB1 Significantly Reduced the Amount of Saturated Fatty Acids in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 3877. [Google Scholar] [CrossRef] [PubMed]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-Mediated Genome Editing of the Fatty Acid Desaturase 2 Gene in Brassica napus. Plant Physiol. Biochem. PPB 2018, 131, 63–69. [Google Scholar] [CrossRef]
- Zhang, K.; Nie, L.; Cheng, Q.; Yin, Y.; Chen, K.; Qi, F.; Zou, D.; Liu, H.; Zhao, W.; Wang, B.; et al. Effective Editing for Lysophosphatidic Acid Acyltransferase 2/5 in Allotetraploid Rapeseed (Brassica napus L.) Using CRISPR-Cas9 System. Biotechnol. Biofuels 2019, 12, 225. [Google Scholar] [CrossRef]
- Xie, T.; Chen, X.; Guo, T.; Rong, H.; Chen, Z.; Sun, Q.; Batley, J.; Jiang, J.; Wang, Y. Targeted Knockout of BnTT2 Homologues for Yellow-Seeded Brassica napus with Reduced Flavonoids and Improved Fatty Acid Composition. J. Agric. Food Chem. 2020, 68, 5676–5690. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Yu, P.; Tian, X.; Guo, L.; Tu, J.; Shen, J.; Yi, B.; Fu, T.; Wen, J.; Liu, K.; et al. DELLA Proteins BnaA6.RGA and BnaC7.RGA Negatively Regulate Fatty Acid Biosynthesis by Interacting with BnaLEC1s in Brassica napus. Plant Biotechnol. J. 2021, 19, 2011–2026. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fu, M.; Li, H.; Wang, L.; Liu, R.; Liu, Z.; Zhang, X.; Jin, S. High-Oleic Acid Content, Nontransgenic Allotetraploid Cotton (Gossypium hirsutum L.) Generated by Knockout of GhFAD2 Genes with CRISPR/Cas9 System. Plant Biotechnol. J. 2021, 19, 424. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Zhu, J.; Gong, L.; He, L.; Lee, C.; Han, S.; Chen, C.; He, G. Mutagenesis of FAD2 Genes in Peanut with CRISPR/Cas9 Based Gene Editing. BMC Biotechnol. 2019, 19, 24. [Google Scholar] [CrossRef] [PubMed]
- Morineau, C.; Bellec, Y.; Tellier, F.; Gissot, L.; Kelemen, Z.; Nogué, F.; Faure, J.D. Selective Gene Dosage by CRISPR-Cas9 Genome Editing in Hexaploid Camelina sativa. Plant Biotechnol. J. 2017, 15, 729–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant Enhancement of Fatty Acid Composition in Seeds of the Allohexaploid, Camelina sativa, Using CRISPR/Cas9 Gene Editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Lyzenga, W.J.; Harrington, M.; Bekkaoui, D.; Wigness, M.; Hegedus, D.D.; Rozwadowski, K.L. CRISPR/Cas9 Editing of Three CRUCIFERIN C Homoeologues Alters the Seed Protein Profile in Camelina sativa. BMC Plant Biol. 2019, 19, 292. [Google Scholar] [CrossRef] [Green Version]
- Ozseyhan, M.E.; Kang, J.; Mu, X.; Lu, C. Mutagenesis of the FAE1 Genes Significantly Changes Fatty Acid Composition in Seeds of Camelina sativa. Plant Physiol. Biochem. PPB 2018, 123, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aznar-Moreno, J.A.; Durrett, T.P. Simultaneous Targeting of Multiple Gene Homeologs to Alter Seed Oil Production in Camelina sativa. Plant Cell Physiol. 2017, 58, 1260–1267. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Zeng, W.; Chen, W.; Wu, Y.; Wen, G.; Chen, X.; Wang, Q.; Zhou, J.; Li, Y.; Yang, Z.; et al. HaCYC2c Regulating the Heteromorphous Development and Functional Differentiation of Florets by Recognizing HaNDUA2 in Sunflower. Plant Cell Rep. 2022, 41, 1025–1041. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.A.; Venegas-Calerón, M.; Du, Z.Y.; Garcés, R.; Tanner, J.A.; Chye, M.L.; Martínez-Force, E.; Salas, J.J. Characterization and Function of a Sunflower (Helianthus annuus L.) Class II Acyl-CoA-Binding Protein. Plant Sci. 2020, 300, 110630. [Google Scholar] [CrossRef]
- CPVO. Essentially Derived Varieties. Available online: https://cpvo.europa.eu/sites/default/files/documents/articles/EDV_presentation_PlantumNL_March_2006_BK.pdf (accessed on 18 June 2023).
Figure 1.
Castor bean plants at different developing stages. (A) Mature plant at flowering; (B) Inflorescent with female flower (red pistil on the top of the inflorescence) and male flower (yellow stamen at the base of the inflorescence); (C) capsules at physiological maturity; (D) dry capsules; (E) trilobate dry capsule and seeds.
Figure 1.
Castor bean plants at different developing stages. (A) Mature plant at flowering; (B) Inflorescent with female flower (red pistil on the top of the inflorescence) and male flower (yellow stamen at the base of the inflorescence); (C) capsules at physiological maturity; (D) dry capsules; (E) trilobate dry capsule and seeds.

Figure 2.
Synoptic scheme of R. communis genetic improvement methodologies. 1 Backcross; 2 Genes of interest; 3 Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Associated Protein 9; 4 Techniques regulated by EU legislation [157].
Figure 2.
Synoptic scheme of R. communis genetic improvement methodologies. 1 Backcross; 2 Genes of interest; 3 Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Associated Protein 9; 4 Techniques regulated by EU legislation [157].
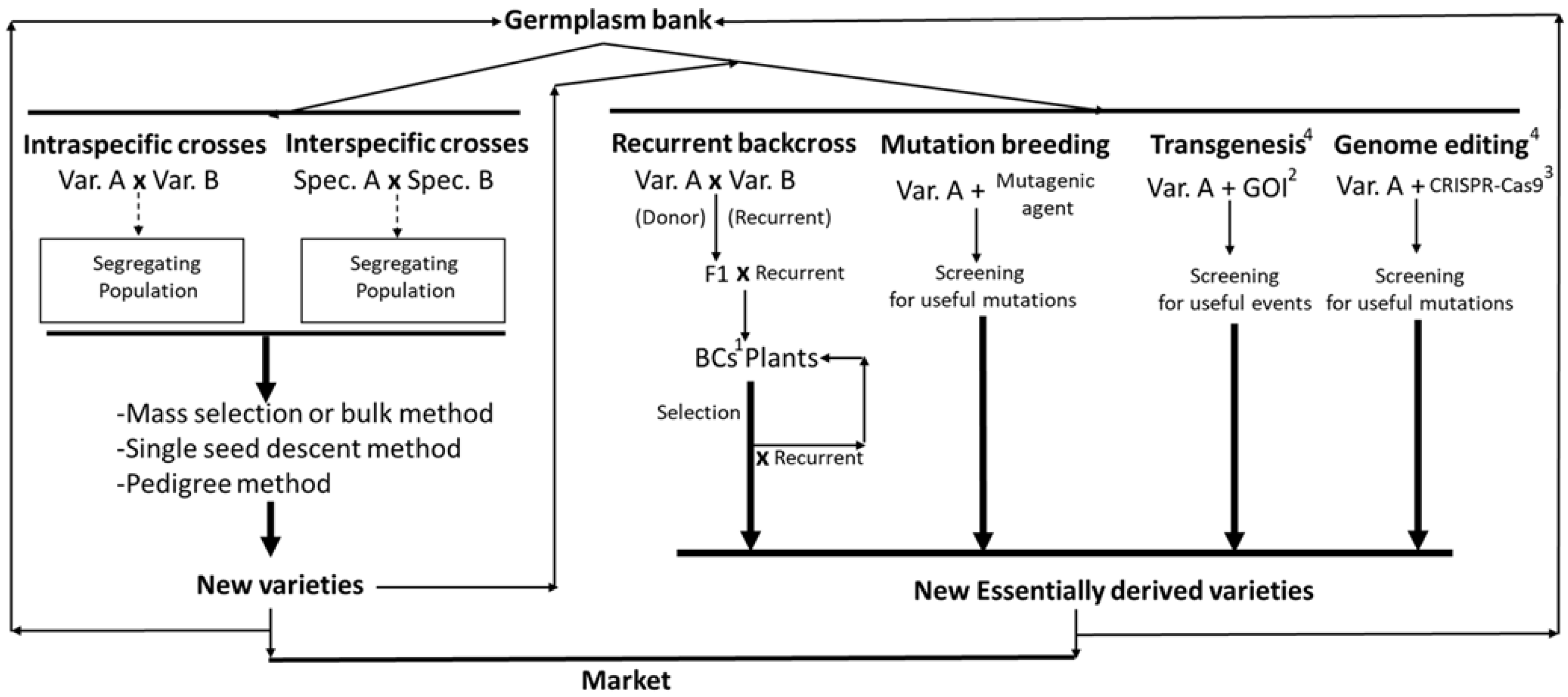

{kind=link}
{kind=link}
Table 1.
Chemical composition of main fatty acid in castor oil.
| Fatty Acid | Amount (%) | Chemical Structure |
|---|---|---|
| Ricinoleic (C18:1-OH) | 88–92 | 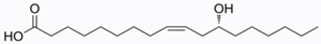 |
| Linoleic (C18:2) | 4.2–4.8 |  |
| Oleic (C18:1) | 2.3–3.4 |  |
| Palmitic (C16:0) | 0.8–1.1 |  |
| Stearic (C18:0) | 0.6–1.1 |  |
Table 2.
Main world germplasm banks harboring genetic resources of castor bean [17].
Table 2.
Main world germplasm banks harboring genetic resources of castor bean [17].
| Institution | Country | Nr. of Accessions |
|---|---|---|
| National Bureau of Plant Genetic Resources | India | 4307 |
| Institute of Crop Science (CAAS) and Institute of Oil Crops Research (CAAS) | China | 3341 |
| Embrapa, Empresa Baiana de Desenvolvimento Agricola S.A. and Instituto Agroanomico de Campinas (I.A.C.) | Brazil | 1348 |
| USDA, ARS, PGRCU and USDA, ARS, NCGRP | United States | 1043 |
| C.I. La Selva-CORPOICA | Colombia | 424 |
| N.I. Vavilov All-Russian Scientific Research Institute of Plant Industry | Russia | 423 |
| Institute of Oil Crops | Ukraine | 255 |
| Biodiversity Conservation and Research Institute | Ethiopia | 232 |
| National Dryland Farming Research Station and National Genebank of Kenya, Crop Plant Genetic Resource Centre, KARI | Kenya | 173 |
| Maize Research Institute and Institute of Field and Vegetable Crops | Serbia | 112 |
| Agricoltural Research Station Teleorman | Romania | 66 |
| Total: | 11,300 |
Table 3.
Oleaginous plant species and genes that have been modified in the recent years by new genome editing technologies.
Table 3.
Oleaginous plant species and genes that have been modified in the recent years by new genome editing technologies.
| Plant | Genes | Mutant Phenotypes | Citations |
|---|---|---|---|
| soybean | FAD2-2 | Increase of oleic acid content and decrease of linoleic acid | [141] |
| GmLox1/2/3 | Free of lipoxygenase | [142] | |
| GmFATB1 | Significant reduction of saturated fatty acids content in seeds | [143] | |
| rapeseed | BnFAD2 | Significant increase in the content of oleic acid | [144] |
| BnLPAT2/5 | Seeds of the mutants were wizened and showed enlarged oil bodies, disrupted distribution of protein bodies, and increased accumulation of starch in mature seeds | [145] | |
| BnTT2 | Yellow-seeded rapeseed with reduced flavonoids and improved fatty acid composition | [146] | |
| BnaLEC1 | Reduced seed oil content and C18:1, increase of C18:2 | [147] | |
| cotton | GhFAD2 | High-oleic acid content, no transgenic allotetraploid cotton (Gossypium hirsutum L.) generated by knockout of GhFAD2 genes with CRISPR/Cas9 system | [148] |
| peanut | AhFAD2 | Mutagenesis of FAD2 genes in peanut with CRISPR/Cas9 based gene editing (NO plant regeneration) | [149] |
| false flax | CsFAD2 | Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa | [150] |
| CsFAD2 | Decreased polyunsaturated fatty acids and concomitant increase of oleic acid in the oil | [151] | |
| CsCRUC | CRISPR/Cas9 editing of three CRUCIFERIN C homeologs alters the seed protein profile in Camelina sativa | [152] | |
| CsFAE1 | Mutagenesis of the FAE1 genes significantly changes fatty acid composition in seeds of Camelina sativa | [153] | |
| CsDGAT1 | Simultaneous targeting of multiple gene homeologs to alter seed oil production in Camelina sativa | [154] | |
| CsPDAT1 | |||
| sunflower | HaCYC2c | Overexpression of HaCYC2c leads to the transformation of floral symmetry and influences the fertility of disc florets for the optimization of reproductive efficiency | [155] |
| HaACBP1 | HaACBP1 plays a role in the transport and trafficking of acyl-CoAs during seed development | [156] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Landoni, M.; Bertagnon, G.; Ghidoli, M.; Cassani, E.; Adani, F.; Pilu, R. Opportunities and Challenges of Castor Bean (Ricinus communis L.) Genetic Improvement. Agronomy 2023, 13, 2076. https://doi.org/10.3390/agronomy13082076
AMA Style
Landoni M, Bertagnon G, Ghidoli M, Cassani E, Adani F, Pilu R. Opportunities and Challenges of Castor Bean (Ricinus communis L.) Genetic Improvement. Agronomy. 2023; 13(8):2076. https://doi.org/10.3390/agronomy13082076
Chicago/Turabian StyleLandoni, Michela, Greta Bertagnon, Martina Ghidoli, Elena Cassani, Fabrizio Adani, and Roberto Pilu. 2023. "Opportunities and Challenges of Castor Bean (Ricinus communis L.) Genetic Improvement" Agronomy 13, no. 8: 2076. https://doi.org/10.3390/agronomy13082076
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.