
Historical Trends Analysis of Main Agronomic Traits in South China Inbred Indica Rice Varieties since Dwarf Breeding

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Materials
2.2. Field Experiments
2.3. Phenotypic Investigation
2.4. Statistical Analysis
3. Results
3.1. Performance of Agronomic Traits of Inbred Indica Rice under Different Environments in South China
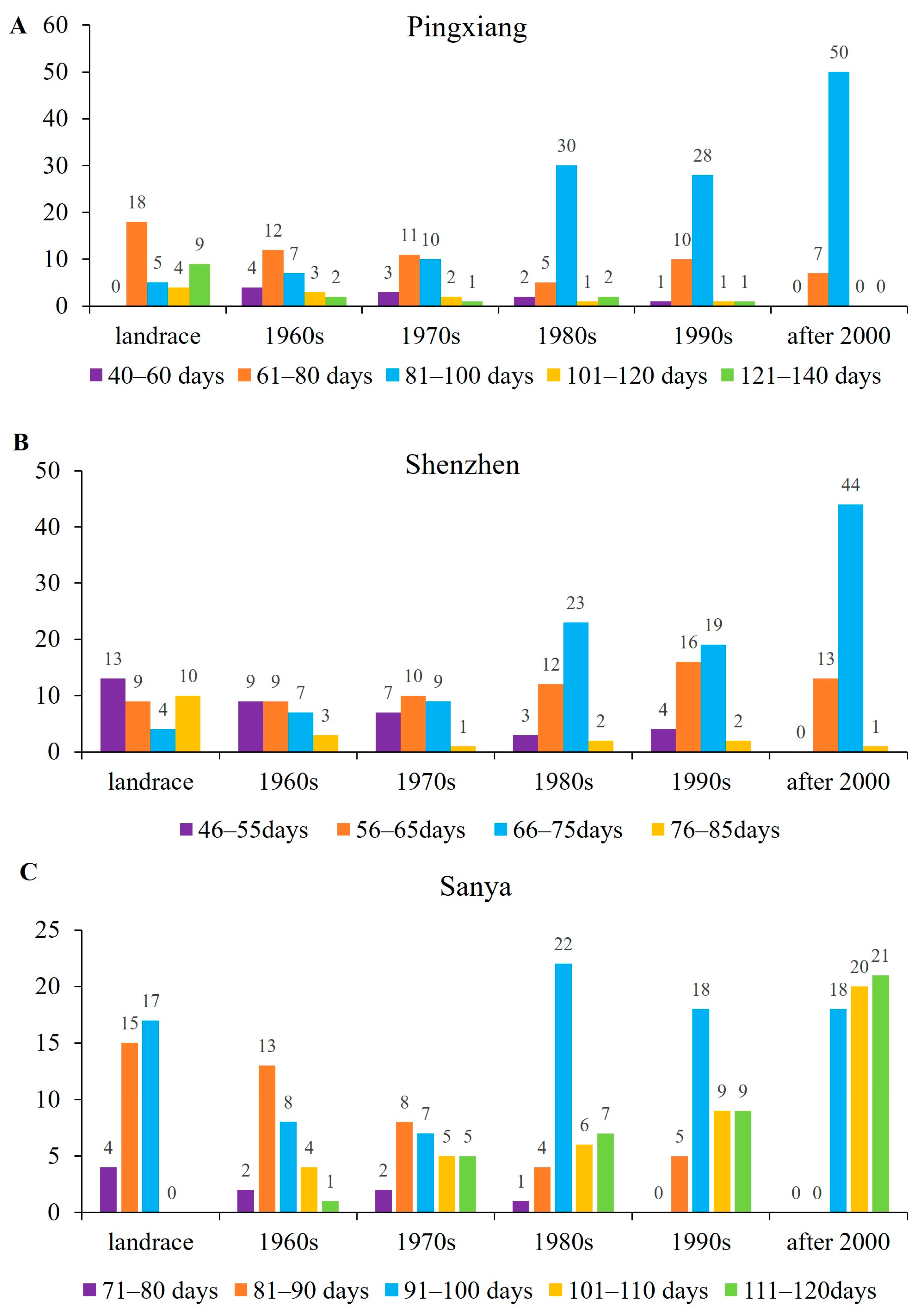
3.1.1. Heading Date
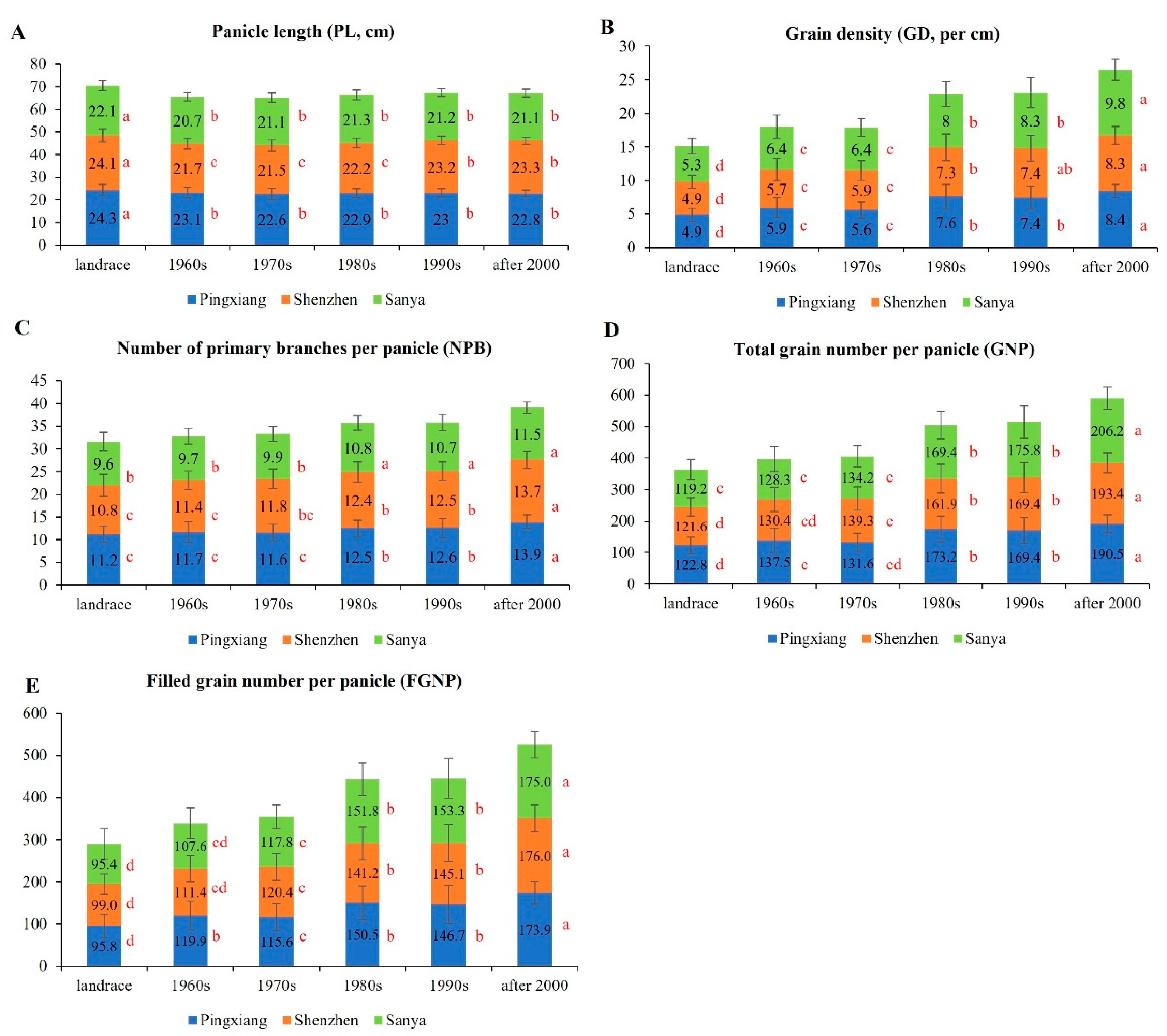
3.1.2. Plant Type-Related Traits
3.1.3. Panicle Type-Related Traits
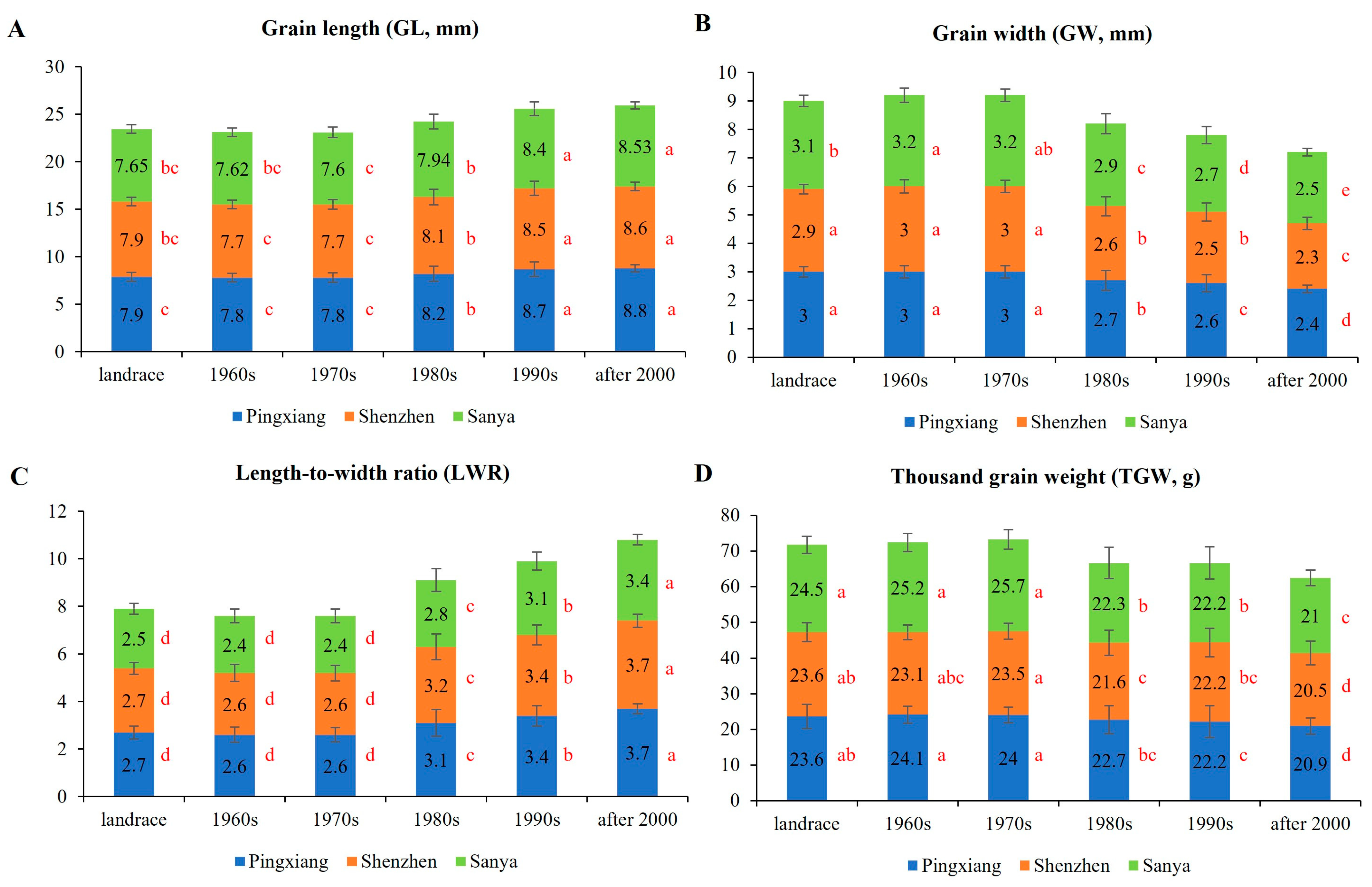
3.1.4. Grain Type-Related Traits
3.1.5. Yield-Related Traits
3.2. Correlations among Important Agronomic Traits and Grain Yield per Plant
3.3. Path Analysis of Agronomic Traits and Grain Yield per Plant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeng, D.; Tian, Z.; Rao, Y.; Dong, G.; Yang, Y.; Huang, L.; Leng, Y.; Xu, J.; Sun, C.; Zhang, G.; et al. Rational design of high-yield and superior-quality rice. Nat. Plants 2017, 20, 17031. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semidwarfing gene, sd-1: Rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.B.; Tang, Q.Y.; Zou, Y.B. Current status and challenges of rice production in China. Plant Prod. Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef]
- Ikeda, M.; Miura, K.; Aya, K.; Kitano, H.; Matsuoka, M. Genes offering the potential for designing yield-related traits in rice. Curr. Opin. Plant Biol. 2013, 16, 213–220. [Google Scholar] [CrossRef]
- Bai, X.; Wu, B.; Xing, Y. Yield-related QTLs and their applications in rice genetic improvement. J. Integr. Plant Biol. 2012, 54, 300–311. [Google Scholar] [CrossRef]
- Yang, S.R. New trend in rice breeding for super high yield combination of ideal plant morphology with favorable vigor. J. Shengyang Agric. Univ. 1987, 18, 4–5. (In Chinese) [Google Scholar]
- Chen, W.F.; Xu, Z.J.; Zhang, L.B.; Zhang, W.Z.; Ma, D.R. Theories and practices of breeding japonica rice for super high yield. Sci. Agric. Sin. 2007, 40, 869–874. (In Chinese) [Google Scholar]
- Yuan, L.P. Hybrid rice breeding for super high yield. Hybrid Rice 1997, 12, 1–6. (In Chinese) [Google Scholar]
- Zhou, K.D.; Wang, X.D.; Li, S.G.; Li, P.; Li, H.Y.; Huang, G.S.; Liu, T.Q.; Shen, M.S. The study on heavy panicle type of intersubspecific hybrid rice (Oryza sativa L.). Sci. Agric. Sin. 1997, 30, 91–93. (In Chinese) [Google Scholar]
- Cheng, S.H.; Cao, L.Y.; Chen, S.G.; Zhu, D.F.; Wang, X.; Min, S.K.; Zhai, H.Q. Conception of late-stage vigor super hybrid rice and its biological significance. Chin. J. Rice Sci. 2005, 19, 280–284. (In Chinese) [Google Scholar]
- Huang, Y.X. Foundation and new development on the scientific system of rice ecological breeding. Chin. Rice Res. Newsl. 2000, 8, 13–14. [Google Scholar]
- Huang, Y.X. Ecological breeding engineering for maximum yield, super high quality of Chinese super rice with semidwarf and “early growth and deep root”. Guangdong Agric. Sci. 2001, 3, 2–6. (In Chinese) [Google Scholar]
- Peng, S.B.; Cassman, K.G.; Virmani, S.S.; Sheehy, J.; Khush, G.S. Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential. Crop Sci. 1999, 39, 1552–1559. [Google Scholar] [CrossRef]
- Peng, S.B.; Laza, R.C.; Visperas, R.M.; Sanico, A.L.; Cassman, K.G.; Khush, G.S. Grain yield of rice cultivars and lines developed in the Philippines since 1966. Crop Sci. 2000, 40, 307–314. [Google Scholar] [CrossRef]
- Yang, J.C.; Wang, P.; Liu, L.J.; Wang, Z.Q.; Zhu, Q.S. Evolution characteristics of grain yield and plant type for mid-season indica rice cultivars. Acta Agron. Sin. 2006, 32, 945–955. (In Chinese) [Google Scholar]
- Tabien, R.E.; Samonte, S.O.; McClung, A.M. Forty-eight years of rice improvement in texas since the release of cultivar bluebonnet in 1944. Crop Sci. 2008, 48, 2097–2106. [Google Scholar] [CrossRef]
- Chen, Z.; Bu, Q.; Liu, G.; Wang, M.; Wang, H.; Liu, H.; Li, X.; Li, H.; Fang, J.; Liang, Y.; et al. Genomic decoding of breeding history to guide breeding-by-design in rice. Natl. Sci. Rev. 2023, 10, nwad029. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Huang, C.W. Studies on the evolutionary changes in yield and related characters of rice cultivars grown in the south China region. J. South China Agric. Univ. 1988, 9, 17–29. (In Chinese) [Google Scholar]
- Liu, C.G.; Zhang, G.Q.; Zhou, H.Q.; Feng, D.J.; Zheng, H.B. Genetic improvement of yield and plant-type traits of inbred indica rice cultivars in south China. Sci. Agric. Sin. 2010, 43, 3901–3911. (In Chinese) [Google Scholar]
- Mann, C.C. Crop scientists seek a new revolution. Science 1999, 283, 310–314. [Google Scholar] [CrossRef]
- Mohapatra, P.K.; Sahu, K. Heterogeneity of primary branch development and spikelet survival in rice panicle in relation to assimilates of primary branches. J. Exp. Bot. 1991, 42, 871–879. [Google Scholar] [CrossRef]
- Liao, Y.P.; Chen, Z.M.; He, X.Y.; Chen, S.J.; Chen, Y.H. Sink, source and flow characteristics of rice variety Yuexiangzhan with high harvest index. Chin. J. Rice Sci. 2001, 15, 73–76. [Google Scholar]
- Horton, P. Prospects for crop improvement through the genetic manipulation of photosynthesis: Morphological and biochemical aspects of light capture. J. Exp. Bot. 2000, 51, 475–485. [Google Scholar] [CrossRef]
- Liu, C.G.; Zhou, H.Q.; Feng, D.J.; Zhou, X.Q.; Chen, D.G.; Li, L.J.; Li, J.C.; Zhang, G.Q.; Chen, Y.D. Analysis of main agronomic traits affecting yield level in inbred indica rice cultivars in south China. Chin. J. Rice Sci. 2012, 26, 182–188. [Google Scholar]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef]
- Song, X.; Meng, X.; Guo, H.; Cheng, Q.; Jing, Y.; Chen, M.; Liu, G.; Wang, B.; Wang, Y.; Li, J.; et al. Targeting a gene regulatory element enhances rice grain yield by decoupling panicle number and size. Nat. Biotechnol. 2022, 40, 1403–1411. [Google Scholar] [CrossRef]
- Feng, F.; Li, Y.; Qin, X.; Liao, Y.; Siddique, K.H.M. Changes in Rice Grain Quality of Indica and Japonica Type Varieties Released in China from 2000 to 2014. Front. Plant Sci. 2017, 8, 1863. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Chen, S.; Gan, Z.; Lu, Q.; Yan, Z.; Chen, G.; Lin, S.; Yang, W.; Zhao, J.; Ba, Y.; et al. Grain shape is a factor affecting the stigma exsertion rate in rice. Front. Plant Sci. 2023, 14, 1087285. [Google Scholar] [CrossRef]
- Ramli, A.B.; Rafii, M.Y.; Latif, M.A.; Saleh, G.B.; Omar, O.B.; Puteh, A.B. Generation mean analysis of grain quality traits in selected rice populations derived from different amylose characteristics. J. Sci. Food Agric. 2016, 96, 1593–1600. [Google Scholar] [CrossRef]
- Chang, T.T.; Li, C.C.; Vergara, B.S. Component analysis of duration from seeding to heading in rice by the basic vegetative phase and the photoperiod-sensitive phase. Euphytica 1969, 18, 79–91. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, F.; Zhou, X.; Poh, T.X.; Xie, L.; Shen, J.; Yang, L.; Song, S.; Yu, H.; Chen, Y. The tetratricopeptide repeat protein OsTPR075 promotes heading by regulating florigen transport in rice. Plant Cell 2022, 34, 3632–3646. [Google Scholar] [CrossRef] [PubMed]
- Osnato, M. Evolution of flowering time genes in rice: From the paleolithic to the anthropocene. Plant Cell Environ. 2023, 46, 1046–1059. [Google Scholar] [CrossRef]
- Xu, L.; Yuan, S.; Man, J. Changes in rice yield and yield stability in China during the past six decades. J. Sci Food Agric. 2020, 100, 3560–3569. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ma, Q.; Yu, H.; Li, Y.; Zhou, L.; He, Q.; Xu, Z.; Zhou, G. Responses of plant biomass and yield component in rice, wheat, and maize to climatic warming: A meta-analysis. Planta 2020, 252, 90. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xue, Y.; Li, J. Towards molecular breeding and improvement of rice in China. Trends Plant Sci. 2005, 10, 610–614. [Google Scholar] [CrossRef]
- Wu, B.; Hu, W.; Xing, Y.Z. The history and prospect of rice genetic breeding in China. Yi Chuan 2018, 40, 841–857. (In Chinese) [Google Scholar]
- Bai, S.; Yu, H.; Wang, B.; Li, J. Retrospective and perspective of rice breeding in China. J. Genet Genom. 2018, 45, 603–612. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Zhao, Y.; Nie, W.; Zhang, Q.; Liu, Z.; Jiang, Y.; Chen, K.; Yu, N.; Luan, X.; Li, W.; et al. Historical Trends Analysis of Main Agronomic Traits in South China Inbred Indica Rice Varieties since Dwarf Breeding. Agronomy 2023, 13, 2159. https://doi.org/10.3390/agronomy13082159
Feng X, Zhao Y, Nie W, Zhang Q, Liu Z, Jiang Y, Chen K, Yu N, Luan X, Li W, et al. Historical Trends Analysis of Main Agronomic Traits in South China Inbred Indica Rice Varieties since Dwarf Breeding. Agronomy. 2023; 13(8):2159. https://doi.org/10.3390/agronomy13082159
Chicago/Turabian StyleFeng, Xiaomin, Ying Zhao, Wenlong Nie, Qiang Zhang, Zhixia Liu, Yijun Jiang, Kai Chen, Ning Yu, Xin Luan, Wenlong Li, and et al. 2023. "Historical Trends Analysis of Main Agronomic Traits in South China Inbred Indica Rice Varieties since Dwarf Breeding" Agronomy 13, no. 8: 2159. https://doi.org/10.3390/agronomy13082159