
Overexpression of the Peanut AhDGAT3 Gene Increases the Oil Content in Soybean
by
Yang Xu
1,2,†,
Fan Yan
1,†,
Zhengwei Liang
2,
Ying Wang
1, 
Jingwen Li
1,
Lei Zhao
1,
Xuguang Yang
1,
Qingyu Wang
1,* and
Jingya Liu
1,* 1
College of Plant Science, Jilin University, Changchun 130062, China
2
Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China
*
Authors to whom correspondence should be addressed.
†
Yang Xu and Fan Yan are co-first authors.
Agronomy 2023, 13(9), 2333; https://doi.org/10.3390/agronomy13092333
Submission received: 14 August 2023
/
Revised: 30 August 2023
/
Accepted: 31 August 2023
/
Published: 7 September 2023
(This article belongs to the Special Issue Advances in Crop Molecular Breeding and Genetics)
Abstract
:Soybean (Glycine max) is the main oilseed crop that provides vegetable oil for human nutrition. The main objective of its breeding research is to increase the total oil content. In the Kennedy pathway, Diacylglycerol acyltransferase (DGAT) is a rate-limiting enzyme that converts diacylglycerol (DAG) to triacylglycerol (TAG). Here, the AhDGAT3 gene was cloned from peanut and overexpressed in the wild-type (WT) Arabidopsis. The total fatty acid content in T3 AhDGAT3 transgenic Arabidopsis seeds was 1.1 times higher on average than that of the WT. Therefore, AhDGAT3 was transferred into the WT (JACK), and four T3 transgenic soybean lines were obtained, which proved to be positive using molecular biological detection. Specific T-DNA insertion region location information was also obtained via genome re-sequencing. The results of high-performance gas chromatography showed that the contents of oleic acid (18:1) composition and total fatty acids in transgenic soybean plants were significantly higher than that of the WT. However, linoleic acid (18:2) was much lower compared to the WT. The agronomic trait survey showed that the quantitative and yield traits of AhDGAT3 transgenic soybean were better than those of the WT. These results suggest that fatty acids in transgenic soybeans, especially oleic acid and total fatty acid, are enhanced by the over-expression of AhDGAT3.
1. Introduction
Soybean (Glycine max) and peanut (Arachis hypogaea L.) are the main sources of vegetable protein and oil, which are the most important bioenergy resources for humans to achieve sustainable development in the world. Peanut is recognized as one of the world’s major oil crops and is widely planted and cultivated in China. The oil content of peanut seeds accounts for more than 50% of the seeds’ dry weight. Among several common edible oil crops, the total oil content of peanut is second only to sesame (Sesamum indicum L.) but is higher than many other common oil crops, such as rapeseed (Brassica campestris L.), soybean, and cottonseed (Gossypium spp.) [1]. More than 80% of unsaturated fatty acids with high quality are stored in peanut seeds, which is approximately 2.5 times more than the two major oil crops, soybean and corn [2]. Therefore, it will be an innovative discovery if the functional genes that regulate the oil content in peanuts can be transformed into other common oil crops.
Soybean is a diploid plant that developed from an ancient tetraploid and contains a set of pathways to synthesize complex lipids. The oil synthesis pathways of soybean involve the process from synthesis to desaturation and the final formation of TAG. There are two methods for the synthesis of soybean fatty acids: one method involves their function as glycerin and phospholipid to form cell membranes, and the other is their storage in seeds, mainly to form TAG [3]. TAG is the main form of oil and fat stored in all kinds of living creatures and is very important for the formation of oil in plant seeds [4]. In addition, many enzymes are involved in the lipid synthesis pathway, including the expression and regulation of different genes. Therefore, it is much more complex to reveal the molecular regulation mechanism of changes in the content of total fatty acids and various fatty acid compositions [5].
DGAT is a rate-limiting enzyme in the Kennedy pathway and plays a key role in the synthesis and accumulation of TAG, which is mainly responsible for the conversion of DAG to TAG. There are four types of DGATs in the DGAT family, including DGAT1, DGAT2, DGAT3, and WS/DGAT [6]. DGAT1 is a member of the Acyl-CoA cholesterol acyltransferase family, which was first cloned in mice and then in Arabidopsis [7,8]. The DGAT1 gene was subsequently cloned in many other plants, including nasturtium, castor, maize, tobacco, and rape [9,10,11,12,13]. The DGAT2 gene was cloned in Mortierella ramanniana and Arabidopsis [14,15], castor [16], and tung tree [17]. The DGAT3 gene has only been identified in peanut [18] and Arabidopsis [19]. However, the DGAT3 protein sequence showed low homology with those of the DGAT1 and DGAT2 subfamilies. WS/DGAT was cloned in fungal microorganisms and has rarely been studied so far.
There are also four types of DGAT genes in peanuts. AhDGAT1 is involved in the lipid synthesis of yeast [20]. AhDGAT2 was found to significantly increase the content of fatty acids in Escherichia coli [21]. The AhDGAT3 gene is a soluble enzyme located in the cytoplasm with an unclear function [18]. However, the in-depth functional research on AhWSD/DGAT is still very limited. Peanut oil has a high nutritional value, and it is easily digested and absorbed by the human body from the composition and proportion of fatty acids in peanut seeds. Peanut oil is mainly composed of 12 fatty acid compositions, in which the sum of palmitic acid (16:0), oleic acid (18:1), and linoleic acid (18:2) accounts for about 90% of the total fatty acids. Kamisaka et al. found that DGAT was purified from the liposome in a lipid-producing fungus [21]. Lardizabal observed that overexpression of UrDGAT2A significantly increased the oil content in transgenic soybean seeds [22]. Chen et al. found that GmDGAT2D was overexpressed in hairy roots, increasing the 18:1 and 18:2 TAG content, whereas overexpression of GmDGAT1A increased the 18:3 TAG content. The overexpression of GmDGAT2D increased 18:1 TAG production and decreased 18:3 TAG in mutant seeds [23]. However, no study has reported that the AhDGAT3 gene is heterologously overexpressed in soybean or verified its further functions and regulation mechanisms so far.
In our study, the AhDGAT3 gene was cloned from peanut seeds, and the role of AhDGAT3 in regulating the total fatty acid content, fatty acid compositions, growth, and development was characterized in transgenic soybean by heterologously overexpressing AhDGAT3 to provide a theoretical basis for further verifying its function in soybean.
2. Materials and Methods
2.1. Plant Materials
Total RNAs were extracted from 30-day-old immature peanut seeds after flowering and used for cloning AhDGAT3 by RT-PCR. Mature seeds of WT soybean (JACK) were used as a negative control for subsequent experiments. JACK was a common soybean variety and receptor for soybean cotyledon node genetic transformation because of its high genetic transformation efficiency. T3 AhDGAT3 transgenic soybean lines were selected and randomly sampled for specificity analyses of gene expression levels and fatty acid compositions. All soybean and peanut seeds were planted in the crop genetics and breeding station in May. Tissues including roots, stems, leaves, flowers, different period of pods after flowering, and mature seeds were sampled randomly for our experiments at particular time. Arabidopsis (Col-0) was used to transform the AhDGAT3 gene and measure the content of fatty acid compositions. The specific disinfection, vernalization, and culture methods of Arabidopsis referred to the relevant literature [24]. The mature seeds of the WT and 5 transgenic Arabidopsis lines after harvesting were selected to extract their DNA, RNA, and protein and measure the total fatty acid content and 5 key fatty acid compositions. All samples were selected and collected randomly from 3 different lines and stored in −80 °C freezer after freezing in liquid nitrogen.
2.2. Isolation and Sequence Analysis of AhDGAT Gene in Peanut
The complete coding sequence of the AhDGAT3 gene was inquired, obtained, and accessed on 4 May 2020 using the NCBI Nucleotide Blast tool (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and peanut genome database (http://www.peanutbase.org). The conserved sequences and regions of DGAT3 in different plants were aligned, analyzed and accessed on 5 May 2020 using the online software MEME 5.5.4 (https://meme-suite.org/meme/tools/meme). Total RNAs were sampled and extracted from 30-day-old immature peanut seeds after flowering. Then, the extracted RNAs were reverse-transcribed into cDNAs, which were used as a template for cloning the AhDGAT3 using RT-PCR method. The specific PCR primers were designed according to the cDNA sequence using Primer Premier 5.0 software. All the RT-PCR primer pairs are provided in Table 1. The RT-PCR reaction contained the forward and reverse primers, cDNA, dNTP mixture, PCR reaction buffer, and double distilled water. After being sequenced in the Biosciences Company (Comate, Changchun, China) for accuracy, the PCR product containing target gene would be constructed to the cloning vector pMD18-T for further studies.
2.3. Arabidopsis and Soybean Transformation of AhDGAT3
The open reading frame of the AhDGAT3 gene was constructed into an overexpression vector pTF101-35s using heat-shock method, and the DH5α competent cells were treated by CaCl2, which needed to be prepared in advance. Then, the resultant plasmid pTF101-AhDGAT3 was transformed into Agrobacterium tumefaciens EHA105 using heat-shock method for the genetic transformation of Arabidopsis and soybean. Then, the resultant plasmid was transformed into Arabidopsis Col-0 using Agrobacterium transformation and floral-dip method [25] and transformed into soybean JACK (WT) using A. tumefaciens to infect the soybean cotyledon nodes. The transformed Arabidopsis and soybean were screened on the MS medium, which contained 5 mg L−1 glufosinate-ammonium. Then, T0 transgenic Arabidopsis and soybeans were detected using PCR and bar strip, which is a simple and accurate method to identify the positive plants. All the positive AhDGAT3 transgenic Arabidopsis and soybean lines were bred to T3 generation for subsequent experiments.
2.4. Molecular Detection of AhDGAT3 Transgenic Soybean
The regenerated plants were transplanted and cultivated in the plant tissue incubator. Leaves of T0 transgenic plants were sampled and identified using PCR and bar strip for positive detection. Transgenic plants were tested via RT-PCR, from which the DNA of AhDGAT3 transgenic soybean leaves was extracted and used as the PCR template. All the specific primer pairs were provided in Table 1 and used to amplify the AhDGAT3 gene (1438 bp) and bar gene (220 bp). Southern blot was performed to verify that the AhDGAT3 gene has been transformed into soybean successfully at the DNA level. The bar and AhDGAT3 genes were chosen as DNA probes, which referred to the DIG High Prime DNA Labeling and Detection Starter Kit II. Western blot was performed to verify the expression level of AhDGAT3 protein in transgenic soybeans on a qualitative level. Protein of JACK leaves was extracted as the negative control. The primary monoclonal antibody, which was required for the experiment, was Mouse Monoclonal with 1:5000 titers. The HRP Goat Anti-Mouse lgG was chosen to be the secondary antibody with 1:6000 titers. ELISA was performed to determine whether the PAT protein was expressed in transgenic soybeans on a quantitative level. The specific experiment methods all referred to the relevant literature [24].
2.5. Genome Re-Sequencing
Four T3 AhDGAT3 transgenic soybean strains were sent to Biotechnology company (Biomarker, Beijing, China) for genome re-sequencing analysis. After the Genomic DNA samples were qualified, the DNA was fragmentated using ultrasonic mechanical interrupt method, and then the end of the DNA fragment was purified and repaired. Then, 3’ and A ends of the DNA were sequenced, connected with joints, and then the DNA fragment was selected and sequenced. Finally, the genomic library was constructed, qualified, and sequenced with Illumina instrument. After the quality of the sequenced original reads (double-ended sequences) was evaluated, the clean reads were selected to be compared with the reference genome sequences, including the mutation detection and annotation of SNP. InDel and mutations were carried out on the basis of sequence alignment. The functional genes were finally discovered and annotated at DNA level.
The AhDGAT3 transgenic soybeans (R01–R04), respectively, were aligned to the reference genome and exogenous T-DNA sequence data, according to the comparison results. The short end of two kinds of matching sequences was found, and the sample data were aligned to the reference genome and the exogenous insertion sequence, respectively. According to the result of alignment, two types of paired ends were found. The insertion sequence was aligned to the reference genome by blast for homology assessment. The assembled contig sequences were aligned to the reference genome by blast, and then the contig sequences were selected to be compared to the regions of chromosomes according to the alignment results. These regions were verified by IGV screenshots, and then the region location information where AhDGAT3 was inserted was obtained.
2.6. Quantitative RT-PCR Analysis
Total RNAs were extracted from the WT soybean, peanut, and AhDGAT3 transgenic soybean tissues, including vegetative growth (root, stem, and leaf) and reproductive growth (different period of pods after flowering and mature seeds). Then, RNAs were reverse-transcribed into cDNAs using RNAiso Plus and M-MLV Kit. All the qRT-PCR primer pairs of the AhDGAT3 and internal reference genes including β-Tubulin and Actin genes, which were used to standardize the data [26,27,28], are provided in Table 1. The qRT-PCR conditions referred to the instructions of TAKARA Biotechnology Company. All the qRT-PCR samples followed the principles of three biological and technical replicates.
2.7. Measurement of Fatty Acid Content in Arabidopsis and Soybean
The total fatty acids of AhDGAT3 transgenic Arabidopsis were extracted using the methylester method. The fatty acid samples were tested and analyzed using a flame ionization detector (Agilent 7890A GC system, Santa Clara, CA, USA) and referring to the methods described in the literature [29]. Five key fatty acid compositions (16:0, 18:0, 18:1, 18:2, and 18:3) were standardized as the control. The total fatty acids of mature seeds in AhDGAT3 transgenic soybeans were extracted using the hydrolysis method (GB5009.168-2016). Two standard sample solutions of the two methods were formulated and used to measure different peak values at different time. And then, we plotted the standard curve in different concentration gradients. All the values were plugged into the standard curve for calculation. The specific experiment methods both referred to the relevant literature [24].
2.8. The Agronomic Trait Analysis of WT and Transgenic Soybeans
After JACK and T3 AhDGAT3 transgenic soybean plants matured, the following agronomic traits were analyzed for each plant, including plant height, main stem node number, effective branch number, 100-seed weight, pod number per plant, seed number per plant, seed weight per plant, and podding height. We randomly selected 30 WT (JACK) and 40 T3 transgenic soybeans (10 of each line). Each result was the average of one soybean line. The 10-day to 50-day pods after flowering of JACK and AhDGAT3 transgenic soybeans were randomly selected and assessed for pod length and width. The pods were measured 3 times. And then, the average length and width of a single soybean were calculated. The results of the agronomic traits were determined by tape measure, vernier calipers, grain counting machines, rulers, and scales.
2.9. Statistical Analysis
All statistical data were calculated and analyzed using SPSS 20.0 (IBM Corp, Armonk, NY, USA) software. Significant differences among the means of samples were compared at p < 0.05 (significant difference) or p < 0.01 (extremely significant difference), based on an independent-sample t-test.
3. Results
3.1. Isolation and Characterization of AhDGAT3 Gene
In order to verify the functions of AhDGAT3, all the AhDGAT family genes were searched, obtained, and accessed on 4 May 2020 from the plant comparative genome database, Phytozome (http://phytozome-next.jgi.doe.gov/), and the Peanut genome database, (http://www.peanutbase.org/), for homologous search. Four candidate genes were identified, named AhDGAT1 (XM_016346849), AhDGAT2 (XM_016095296), AhDGAT3 (XM_016339125), and AhWSD/DGAT (XM_016109460). The AhDGAT1, AhDGAT2, and AhDGAT3 genes were closely related but were far related to the AhWSD/DGAT. In the DGAT3 conserved domain, Motif1 contains the characteristic fatty-acid-binding protein sequence KSGSIALLQEFERVVGAEG, and Motif2 contains the CKCMGKCKSAPNVRIQNSTAD conserved sequences. Motif3 contains the speculated structural motif NPLCIGVGLEDVDAIVA, and Motif4 contains the DDLQGNLTWDAAEVLMKQLEQVRAEEKELKKKQKQEKKAKL conserved sequences. The subfamily also contains the KKRVLFDDL active sites of the acyltransferase family [15]. In addition, this subfamily also contains the thioesterase characteristic sequence TNPDCESSSSSSSSESESES (Figure 1).
3.2. Oil Content in Seeds of Different Genotype Arabidopsis Plants
To reveal and verify the functions of AhDGAT3 in the accumulation of oil in seeds, the AhDGAT3 gene was transformed into the WT Arabidopsis (Col-0) plants. Mature seeds from T3 AhDGAT3 transgenic lines were sampled to measure the oil content. The results showed that the content of 18:1 composition in AhDGAT3 transgenic Arabidopsis seeds was significantly higher than that of the WT, reaching 1.3 times on average, respectively (Figure 2A). However, the contents of 16:0, 18:2, and 18:3 compositions in the transgenic lines were slightly lower than those in the WT. The total fatty acid content in four AhDGAT3 transgenic soybean lines was also enhanced a lot (Figure 2B).
3.3. Overexpression AhDGAT3 Gene in Soybean
To verify the further functions of AhDGAT3 in soybean, four T3 AhDGAT3 transgenic soybean lines were obtained and detected via a series of molecular biology methods, including the bar strip, Southern blot, and Western blot. The results showed that AhDGAT3 was transformed into a JACK cultivar background (Figure 3). ELISA analysis revealed that the expression level in transgenic soybeans was higher than that of the WT (Table 2).
The 1038 bp AhDGAT3 gene was inserted into the pTF101-35s vector with the Xba I/Sac I restriction site (Figure 3C). The distances between the bar gene probe and the left/right boundaries of T-DNA were approximately 9 kb and 2 kb, respectively. The distances between the target gene and the left/right boundaries of T-DNA were approximately 9.6 kb and 1 kb, respectively (Figure 3C). The results of the Southern blot showed that only one hybridization band appeared in four AhDGAT3 transgenic lines (OE-1 to OE-4), and no hybridization band was detected in the WT. This suggested that exogenous T-DNA was integrated in the form of single-copy DNA in four AhDGAT3 transgenic lines (Figure 3B,D). Specific insertion sites were analyzed and studied via genome re-sequencing in transgenic soybean. The AhDGAT3 gene was translated into the protein and synthesized monoclonal antibodies to identify the transgenic soybeans. The Western blot results showed that there was an apparent hybridization signal at 38 KD, indicating that the AhDGAT3 gene was over-expressed at the protein level in transgenic plants (Figure 3E).
The ELISA results showed that the AhDGAT3 protein was not expressed in the WT (negative control), and no value was calculated using a standard curve. The AhDGAT3 protein of four transgenic lines was detected and calculated using a standard curve. AhDGAT3 expression levels in the vegetative growth stage were lower than those of the reproductive growth stage. AhDGAT3 level accumulation was highest in mature seeds as the most important organ for oil storage, which was approximately two to five times higher than that of the nutritious organs. This result can further indicate that AhDGAT3 proteins are expressed in four transgenic soybean lines (Table 2).
3.4. Genome Re-Sequencing of AhDGAT3 Transgenic Soybean
In this analysis, the genomic DNA from four T3 transgenic soybean leaves was re-sequenced. Clean data with a total data volume of 92.59 Gbp were obtained, and Q30 reached 90.51%. The average contrast ratio of the sample to the W82 reference genome was 98.88%. The depth of the average coverage was 21× and the depth of the genome coverage was 98.28%, which made sure at least one base was covered.
The re-sequencing data of four transgenic soybeans (R01–R04) were compared with reference genome and exogenous T-DNA sequences, and all short sequences that could be compared with exogenous T-DNA sequences were selected for the analysis. The assembled contig sequences were blasted to the reference genome. And then, the contig sequences were selected to be compared to the regions of chromosomes according to the alignment results. These regions were verified by IGV screen shots to obtain the location information of exogenous insertion fragments. All the results of gene insertion information are shown in Table 3.
3.5. Expression Analysis of AhDGAT3 in WT and Transgenic Soybean
Tissues in vegetative and reproductive growth stages from JACK and transgenic soybeans were collected, and the total RNAs were extracted to verify gene over-expression in the transgenic soybean using qRT-PCR. The results showed that AhDGAT3 expression in transgenic soybeans was significantly (p < 0.01) higher than that of the WT in the leaves and 10- and 40-day-old pods after flowering. The AhDGAT3 expression level in the flower and 30-day-old pods of transgenic soybeans was significantly higher (p < 0.05) than that in the WT (Figure 4).
3.6. AhDGAT3 Over-Expression Enhances Fatty Acid Content in Transgenic Soybean
Mature soybean seeds (0.01 g) were collected and sampled with three biological replicates, including the WT (JACK) and AhDGAT3 transgenic lines, which were used for oil determination and analysis. The results of high-performance gas chromatography (HPGC) showed no significant difference in the content of 16:0, 18:0, and 18:3 fatty acid compositions between AhDGAT3 transgenic soybeans and JACK. However, the difference of the 18:1 content was significant, with the transgenic line being 27.3% higher than JACK, respectively. The content of the 18:2 fatty acid compositions was significantly lower than JACK. The total fatty acid content of the T3 AhDGAT3 transgenic soybean was up to 4% higher than that of JACK (Figure 5).
3.7. AhDGAT3 Overexpression Improved the Agronomic Traits of Transgenic Soybean
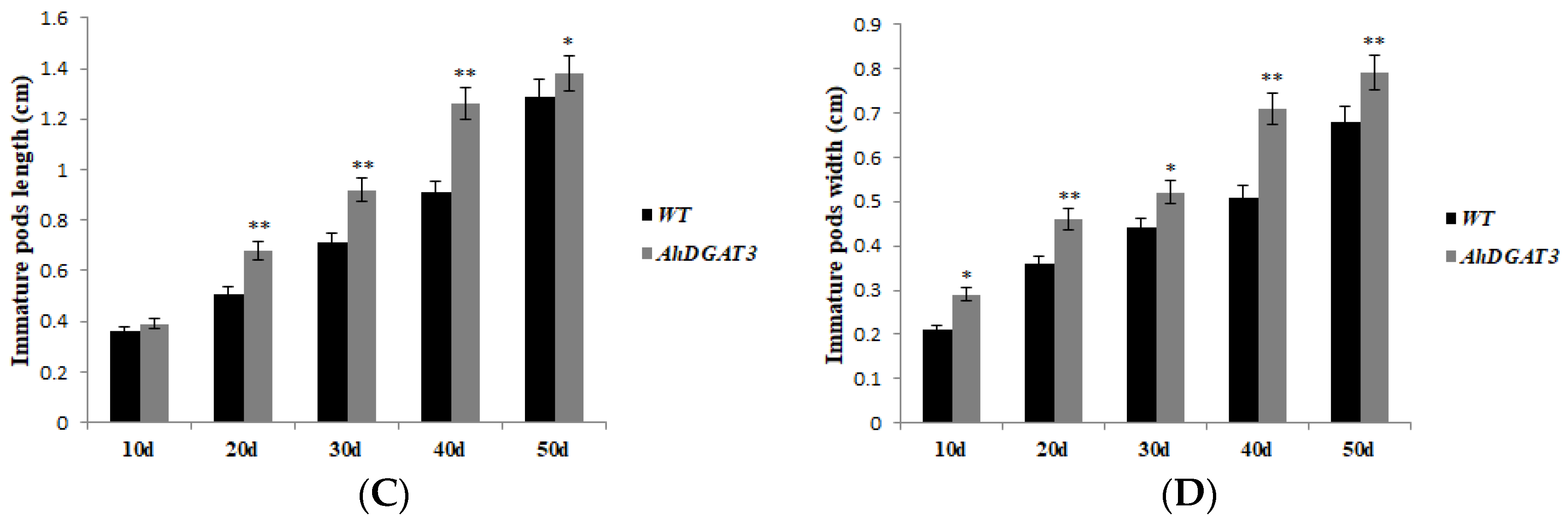
The analysis of the agronomic traits in the WT and AhDGAT3 transgenic soybean seeds showed that plant height, effective branch number, pod number per plant, and seed weight per plant in the AhDGAT3 transgenic soybean were significantly superior to the WT (Figure 6A and Table 4). The size of 10-day to 50-day pods after flowering of the AhDGAT3 transgenic soybean was significantly bigger than the WT at different stages of reproductive growth (Figure 6B–D). The results showed that overexpression of the AhDGAT3 gene may also affect the growth and development of soybean.
4. Discussion
Soybean is an important food and oil crop in the world, accounting for a significant proportion [30]. How to effectively improve vegetable oil content and the composition ratio of unsaturated fatty acids is an important breeding goal for us. Heterologous gene expression is an important means to improve soybean oil content.
Recently, studies on how to improve the oil content of plant seeds have mainly focused on the lipid metabolism pathway. All kinds of functional enzyme genes and transcription factors, including PDAT, FAD, SAD, LEC1, LEC2, Dof, and ABI3, were cloned and studied, which proved to play an important role [31]. It is a major technical method to verify the coordinated expression of multiple genes in metabolic pathways via genetic transformation technology [32]. However, as an enzyme gene that plays a key catalytic role in the conversion process of DAG to TAG. DGAT plays an important role in regulating the accumulation of fatty acids and lipid synthesis in oil crops [33]. The oil content of seeds depends on quantitative heredity and is regulated by all kinds of factors, including the synthesis and accumulation of fatty acids and the development of seeds [34,35].
Compared to animal fatty acids, various unsaturated fatty acids in plants are very beneficial to our health. The demand for high-oleic-acid vegetable oil is also increasing with the improvement of our living standards. The results showed that AhDGAT3 was over-expressed in Arabidopsis and soybean. The contents of 18:1 unsaturated fatty acid compositions in transgenic plant seeds were significantly increased. This result is consistent with recent research reports [23]. AtDGAT3 was involved in a soluble cytosolic process in the circulation of linoleic and linolenic acid to TAG during the degradation and breakdown of seed oil [19]. However, no in-depth studies showed the functions of the AhDGAT3 gene using heterologous transformation and genetic engineering technology between peanut and soybean. Most recent studies on DGAT genes mainly focus on functional analysis in Arabidopsis. The UrDGAT2 gene of Umbelopsis ramanniana was successfully transferred into soybean and over-expressed in seeds. The UrDGAT2 transgenic soybean showed a higher oil content [22]. TAG, as the main storage in oil crop seeds, provided enough C sources for seed germination and development. The up-regulation of the DGAT1 gene was involved in the fatty acid mobilization and the catalyzation in the reverse reaction of TAG synthesis during seed germination and development [36,37]. The results of the agronomic traits showed that the phenotype of the AhDGAT3 transgenic soybean was better than that of the WT soybean in terms of the size of different period pods, the number of effective branches, the number of seeds per plant, pod number per plant, main stem node number, and the weight of seeds per plant at harvest time. We speculated that AhDGAT3 was also closely related to the growth and development of soybeans. Moreover, the next step is to study the internal regulation mechanism of AhDGAT3. We aimed to develop high-quality transgenic soybean lines with high oil content and yield, creating excellent soybean germplasm resources.
5. Conclusions
We characterized the major functions of AhDGAT3 in Arabidopsis and soybean. The content of total fatty acids and 18:1 compositions in the AhDGAT3-overexpressed transgenic soybean was significantly higher than those of the WT (JACK). The re-sequencing of AhDGAT3 transgenic soybean revealed that T-DNA was integrated into the soybean genome as a single copy. Overexpression of the AhDGAT3 gene can affect the size of soybean immature pods and a series of agronomic traits. Therefore, we also speculate that this gene may also play a key role in soybean growth and development, which needs further study. Our results indicated that molecular-assisted breeding is a very efficient and fast way of increasing the oil content of soybean seeds, which can also be used in the modulation of other agronomic traits of soybean in the future.
Author Contributions
Conceptualization, Y.X., F.Y. and Q.W.; Data curation, F.Y. and Y.W.; Formal analysis, Y.X., Z.L., J.L. (Jingwen Li) and Y.W.; Funding acquisition, Q.W.; Investigation, Y.X. and X.Y.; Methodology, F.Y. and L.Z.; Project administration, Q.W.; Resources, J.L. (Jingwen Li); Software, J.L. (Jingya Liu); Supervision, Q.W.; Validation, Q.W.; Visualization, F.Y.; Writing—original draft, Y.X.; Writing—review and editing, Y.X. and F.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science and Technology Development Plan Project of Jilin Province (No. 20220202009NC), the National Natural Science Foundation of China (No. 32101689), and the Science and Technology research project of Education Department of Jilin Province (No. JJKH20221034KJ).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liao, B.S. Analysis on the competitiveness of peanut oil industry in China. J. Peanut 2003, 32, 11–15. [Google Scholar]
- Li, X.D.; Cao, Y.L.; Hu, Y.P.; Xiao, L.; Wu, Y.; Wu, G.; Lu, C. Study on fatty acid accumulation patterns in peanut seed development. Chin. J. Oil Crops 2009, 31, 157–162. [Google Scholar]
- Mekhedov, S.; De Ilárduya, O.M.; Ohlrogge, J. Toward a functional catalog of the plant genome. A survey of genes for lipid biosynthesis. Plant Physiol. 2000, 122, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Barthole, G.; Lepiniec, L.; Rogowsky, P.M.; Baud, S. Controlling lipid accumulation in cereal grains. Plant Sci. 2012, 185–186, 33–39. [Google Scholar] [CrossRef]
- Liu, Z.J. Cloning of Genes Involved in Seed Oil Content and Transgenic Research of Cotton. Ph.D. Thesis, China Agricultural University, Beijing, China, 2013. [Google Scholar]
- Yen, C.E.; Stone, S.J.; Koliwad, S.; Harris, C.; Farese, R.V., Jr. DGAT enzymes and triacylglyrol biosynthesis. J. Lipid Res. 2008, 49, 2283–2301. [Google Scholar] [CrossRef] [PubMed]
- Cases, S.; Smith, S.J.; Zheng, Y.W. Identification of a gene encoding an acyl-Co A: Diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13018–13023. [Google Scholar] [CrossRef]
- Hobbs, H.D.; Chaofu, L.; Hills, M. Cloning of a c DNA encoding acyltransferase from Arabidopsis thaliana and its functional expression. FEBS Lett. 1999, 452, 145–149. [Google Scholar] [CrossRef]
- Xu, J.; Francis, T.; Mietkiewska, E.; Giblin, E.M.; Barton, D.L.; Zhang, Y.; Zhang, M.; Taylor, D.C. Cloning and characterization of an acyl Co A-dependent diacylglycerol acyltransferase 1 (DGAT1) gene from Tropaeolummajus, and a study of the functional motifs of the DGAT protein using site-directed mutagenesis to modify enzyme activity and oil content. Plant Biotechnol. J. 2008, 6, 799–818. [Google Scholar] [CrossRef]
- He, X.H.; Grace, Q.C.; Lin, J.T.; McKeon, T.A. Regulation of diacylglycerol acyltransferase in developing seeds of castor. Lipids 2004, 39, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Allen, W.B.; Roesler, K.; Williams, M.E.; Zhang, S.; Li, J.; Glassman, K.; Ranch, J.; Nubel, D.; Solawetz, W.; et al. A phenylalanine in DGAT is a key determinant of oil content and composition in maize. Nat. Genet. 2008, 40, 367–372. [Google Scholar] [CrossRef]
- Bouvier-Navé, P.; Benveniste, P.; Oelkers, P.; Sturley, S.L.; Schaller, H. Expression in yeast and tobacco of plant c DNAs encoding acyl-CoA: Diacylglycerol acyltransferase. Eur. J. Biochem. 2000, 267, 85–96. [Google Scholar] [CrossRef]
- Nykiforuk, C.L.; Laroche, A.; Weselake, R.J. Isolation and characterization of a c DNA encoding a second putative diacylglycerol acyltransferase from a microspore-derived cell suspension culture of Brassica napus L. cv Jet Neuf. Plant Physiol. 1999, 121, 1957–1959. [Google Scholar]
- Salanoubat, M.; Lemcke, K.; Rieger, M.; Ansorge, W.; Unseld, M.; Fartmann, B.; Valle, G.; Blöcker, H.; Perez-Alonso, M.; Obermaier, B.; et al. Sequence and analysis of chromosome 3 of the plant Arabidopsis thaliana. Nature 2000, 408, 820–822. [Google Scholar]
- Lardizabal, K.D.; Mai, J.T.; Wagner, N.W.; Wyrick, A.; Voelker, T.; Hawkins, D.J. DGAT2 is a new diacylglycerol acyltransferase gene family: Purification, cloning, and expression in insect cells of two polypeptides from Mortierella ramanniana with diacylglycerol acyltransferase activity. J. Biol. Chem. 2001, 276, 38862–38869. [Google Scholar] [CrossRef] [PubMed]
- Kroon, J.T.; Wei, W.; Simon, W.J.; Slabas, A.R. Identification and functional expression of a type 2 acyl-Co A: Diacylglycerol acyltransferase (DGAT2) in developing castor bean seeds which has high homology to the major triglyceride biosynthetic enzyme of fungi and animals. Phytochemistry 2006, 67, 2541–2549. [Google Scholar] [CrossRef]
- Shockey, J.M.; Gidda, S.K.; Chapital, D.C.; Kuan, J.-C.; Dhanoa, P.K.; Bland, J.M.; Rothstein, S.J.; Mullen, R.T.; Dyer, J.M. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell 2006, 18, 2294–2313. [Google Scholar] [CrossRef]
- Saha, S.; Enugutti, B.; Rajakumari, S.; Rajasekharan, R. Cytosolic triacylglycerol biosynthetic pathway in oilseeds. Molecular cloning and expression of peanut cytosolic diacylglycerol acyltransferase. Plant Physiol. 2006, 141, 1533–1543. [Google Scholar] [CrossRef]
- Hernandez, M.L.; Whitehead, L.; He, Z.; Gazda, V.; Gilday, A.; Kozhevnikova, E. A cytosolic acyltransferase contributes to triacylglycerol synthesis in sucrose-rescued Arabidopsis seed oil catabolism mutants. Plant Physiol. 2012, 160, 215–225. [Google Scholar] [CrossRef]
- Zheng, L.; Shockey, J.; Guo, F.; Shi, L.; Li, X.; Shan, L.; Wan, S.; Peng, Z. Discovery of a new mechanism for regulation of plant triacylglycerol metabolism: The peanut diacylglycerol acyltransferase-1 gene family transcriptome is highly enriched in alternative splicing variants. J. Plant Physiol. 2017, 219, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Kamisaka, Y.; Mishra, S.; Nakahara, T. Purification and characterization of diacylglycerol acyltransferase from the lipid body fraction of an oleagious fungus. J. Biol. Chem. 1997, 121, 1107–1114. [Google Scholar]
- Lardizabal, K.; Effertz, R.; Levering, C.; Mai, J.; Pedroso, M.C.; Jury, T.; Aasen, E.; Gruys, K.; Bennett, K. Expression of Umbelopsis ramanniana DGAT2A in seed increases oil in soybean. Plant Physiol. 2008, 148, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.B.; Wang, J.J.; Zhang, G.Y. Two types of soybean diacylglycerol acyltransferases are differentially involved in triacylglycerol biosynthesis and response to environmental stresses and hormones. Sci. Rep. 2016, 6, 28541. [Google Scholar] [CrossRef]
- Xu, Y.; Yan, F.; Liu, Y.; Wang, Y.; Gao, H.; Zhao, S.; Wang, Q.; Li, J. Quantitative proteomic and lipidomics analyses of high oil content GmDGAT1-2 transgenic soybean, illustrates the regulatory mechanism of lipoxygenase and oleosin. Plant Cell Rep. 2021, 40, 2303–2323. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Bo, J.; Liu, B.; Bi, Y.; Hou, W.; Wu, C. Validation of Internal Control for Gene Expression Study in Soybean by Quantitative Real-Time PCR. BMC Mol. Biol. 2008, 9, 59. [Google Scholar]
- Hu, R.; Fan, C.; Li, H.; Zhang, Q. Evaluation of Putative Reference Genes for Gene Expression Normalization in Soybean by Quantitative Real-Time RT-PCR. BMC Mol. Biol. 2009, 10, 93. [Google Scholar] [CrossRef]
- Le, D.T.; Aldrich, D.L.; Valliyodan, B.; Watanabe, Y.; Ha, C.V.; Nishiyama, R.; Guttikonda, S.K.; Quach, T.N.; Gutierrez-Gonzalez, J.J.; Tran, L.S.P.; et al. Evaluation of Candidate Reference Genes for Normalization of Quantitative RT-PCR in Soybean Tissues under Various Abiotic Stress Conditions. Edite par Christian Schönbach. PLoS ONE 2012, 7, e46487. [Google Scholar] [CrossRef]
- Browse, J.; McCourt, P.J.; Somerville, C.R. Fatty acid composition of leaf lipids determined after combined digestion and fatty acid methyl ester formation from fresh tissue. Anal. Biochem. 1986, 152, 141–145. [Google Scholar] [CrossRef]
- Broun, P.; Gettner, S.; Somerville, C. Genetic engineering of plant lipids. Annu. Rev. Nutr. 1999, 19, 197–216. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-lipidmetabolism. Arab. Book 2010, 8, e0133. [Google Scholar] [CrossRef]
- Sun, Q.X. Cloning and Characterization of Genes Involved in the Biosynthesis of Very Long Chain Polyunsaturated Fatty Acid and the Reconstitution of This Pathway in Crop Plants. Ph.D. Thesis, Shandong Agricultural University, Taian, China, 2011. [Google Scholar]
- Jako, C.; Kumar, A.; Wei, Y.D.; Zou, J.; Barton, D.L.; Giblin, E.M.; Covello, P.S.; Taylor, D.C. Seed-specific over-expression of an Arabidopsis cDNA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight. Plant Physiol. 2001, 126, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Ohlrogge, J. Supply of triacylglycerol in developing embryos. Plant Physiol. 1999, 120, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.L.; Isleib, T.G. Genetic analysis of fatty acids in American peanuts. Chin. J. Oil Crops 2000, 22, 35–37. [Google Scholar]
- Zou, J.; Wei, Y.D.; Jako, C.; Kumar, A.; Selvaraj, G.; Taylor, D.C. The Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol acyltransferase gene. Plant J. 1999, 19, 645–653. [Google Scholar] [CrossRef]
- Feussner, I.; Kuhn, H.; Wasternack, C. Lipoxygenase-dependent degradation of storage lipids. Trends Plant Sci. 2001, 6, 268–273. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Partial alignment of DGAT3 deduced amino acids in different plants. Sequences were aligned using MEGA 7.0 software. Different shapes of lines represent four different special functional domains and two special sites. Accession numbers of 9 genes in GenBank are as follows: Arachis hypogaea, AAX62735.1; Glycine max, XP_003542403.1; Glycine soja, XP_028209585.1; Brassica rapa, RID49571.1; Lotus japonicas, AFK37850.1; Lupinus angustifolius, XP_019419028.1; Medicago truncatula, XP_003609890.1; Prosopis alba, XP_028760027.1.
Figure 1.
Partial alignment of DGAT3 deduced amino acids in different plants. Sequences were aligned using MEGA 7.0 software. Different shapes of lines represent four different special functional domains and two special sites. Accession numbers of 9 genes in GenBank are as follows: Arachis hypogaea, AAX62735.1; Glycine max, XP_003542403.1; Glycine soja, XP_028209585.1; Brassica rapa, RID49571.1; Lotus japonicas, AFK37850.1; Lupinus angustifolius, XP_019419028.1; Medicago truncatula, XP_003609890.1; Prosopis alba, XP_028760027.1.

Figure 2.
(A) Content of 5 fatty acid compositions in the WT and AhDGAT3 transgenic Arabidopsis lines. (B) Total fatty acid content in the WT and AhDGAT3 transgenic Arabidopsis seeds. The data represent the average of 3 independent repetitions of experiments. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference. The peaks and peak time of each group were palmetto acid (16:0), 11.672 min; stearic acid (18:0), 16.011 min; oleic acid (18:1), 16.471 min; linoleic acid (18:2), 17.779 min; and linolenic acid (18:3), 20.345 min.
Figure 2.
(A) Content of 5 fatty acid compositions in the WT and AhDGAT3 transgenic Arabidopsis lines. (B) Total fatty acid content in the WT and AhDGAT3 transgenic Arabidopsis seeds. The data represent the average of 3 independent repetitions of experiments. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference. The peaks and peak time of each group were palmetto acid (16:0), 11.672 min; stearic acid (18:0), 16.011 min; oleic acid (18:1), 16.471 min; linoleic acid (18:2), 17.779 min; and linolenic acid (18:3), 20.345 min.
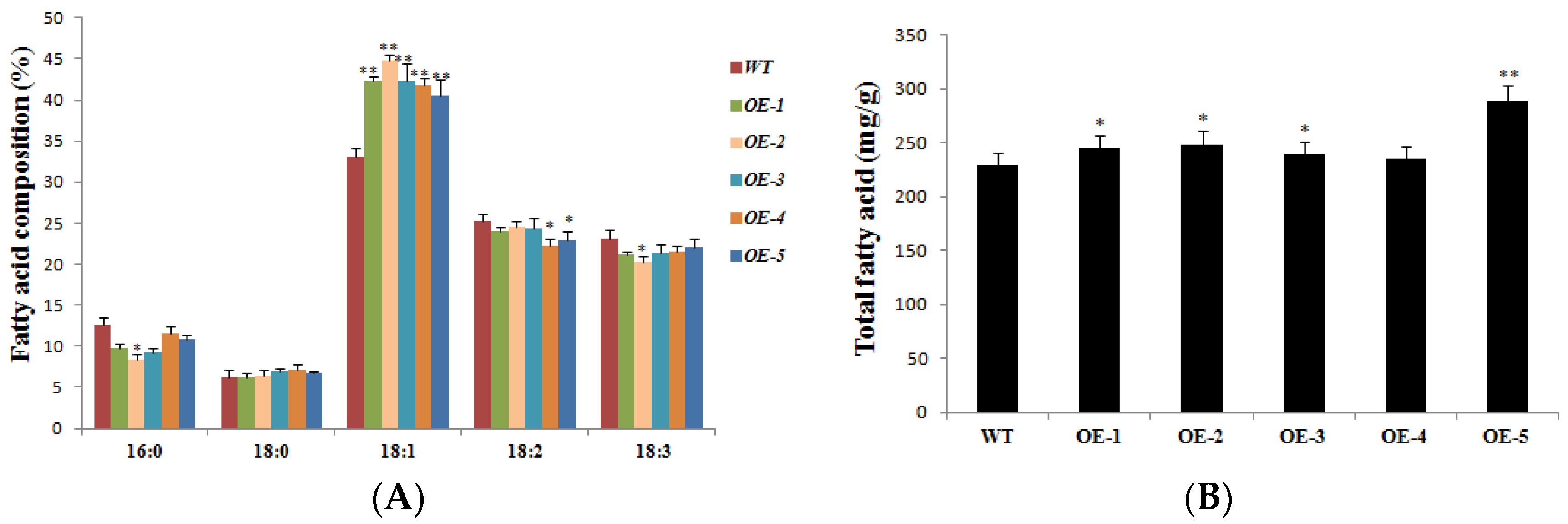
Figure 3.
Positive detection of AhDGAT3 transgenic soybean lines. (A) Bar strip analysis of 4 AhDGAT3 transgenic soybeans. (B) Southern blot of T3 transgenic soybeans using the bar gene fragment as a probe. M represents DNA marker; P represents recombinant plasmid (positive control); WT represents JACK (negative control). (C) Map of pTF101-AhDGAT3 recombinant plasmid in AhDGAT3 transgenic soybeans. (D) Southern blot of T3 transgenic soybeans using the target gene fragment as a probe. M represents DNA marker; P represents recombinant plasmid (positive control); WT represents JACK (negative control). (E) Western blot of T3 transgenic soybeans. M represents pre-stained protein marker; P represents AhDGAT3 protein (positive control); WT represents JACK (negative control).
Figure 3.
Positive detection of AhDGAT3 transgenic soybean lines. (A) Bar strip analysis of 4 AhDGAT3 transgenic soybeans. (B) Southern blot of T3 transgenic soybeans using the bar gene fragment as a probe. M represents DNA marker; P represents recombinant plasmid (positive control); WT represents JACK (negative control). (C) Map of pTF101-AhDGAT3 recombinant plasmid in AhDGAT3 transgenic soybeans. (D) Southern blot of T3 transgenic soybeans using the target gene fragment as a probe. M represents DNA marker; P represents recombinant plasmid (positive control); WT represents JACK (negative control). (E) Western blot of T3 transgenic soybeans. M represents pre-stained protein marker; P represents AhDGAT3 protein (positive control); WT represents JACK (negative control).
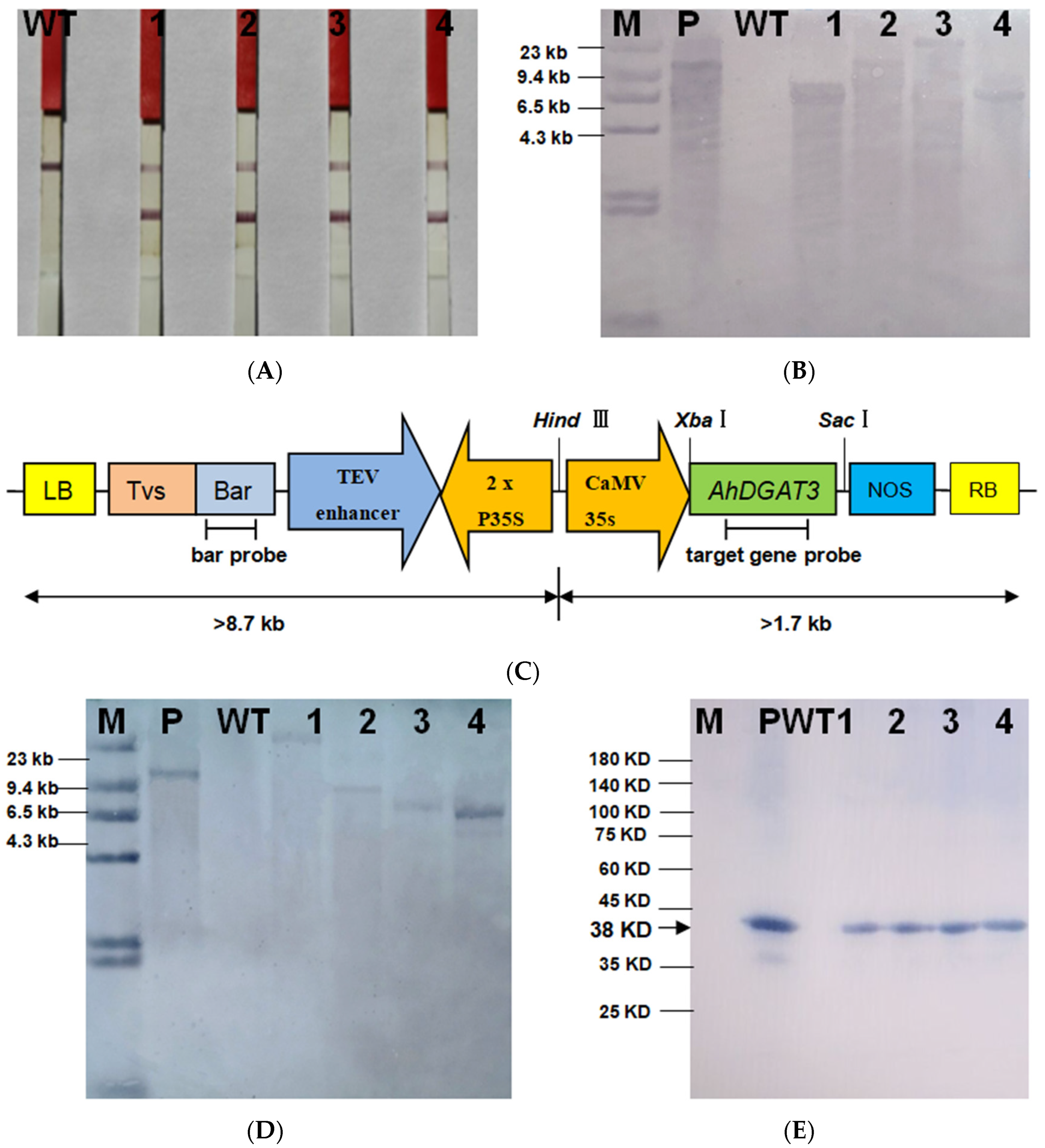
Figure 4.
Expression levels of AhDGAT3 in different tissues of soybean. The data represent the average of 3 independent experiments ± SD. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference.
Figure 4.
Expression levels of AhDGAT3 in different tissues of soybean. The data represent the average of 3 independent experiments ± SD. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference.
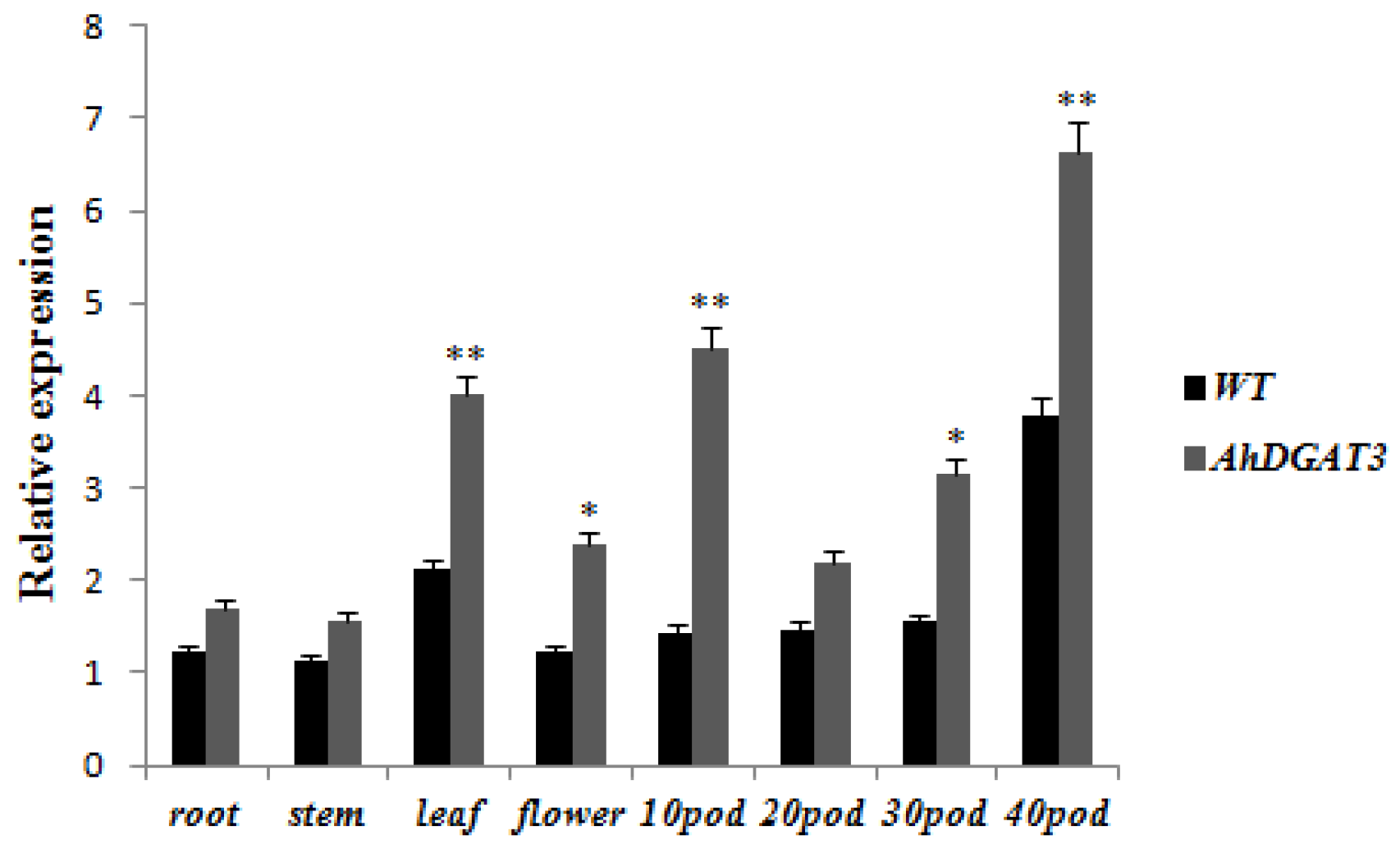
Figure 5.
Content of different fatty acid compositions and total fatty acids in soybean seeds of different genotypes. (A) The content of different fatty acid compositions in JACK and 4 different AhDGAT3 transgenic lines (OE-1 to 4) of soybean, measured by HPGC. (B) Total fatty acid of 4 AhDGAT3 overexpressing soybean seeds. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference.
Figure 5.
Content of different fatty acid compositions and total fatty acids in soybean seeds of different genotypes. (A) The content of different fatty acid compositions in JACK and 4 different AhDGAT3 transgenic lines (OE-1 to 4) of soybean, measured by HPGC. (B) Total fatty acid of 4 AhDGAT3 overexpressing soybean seeds. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference.
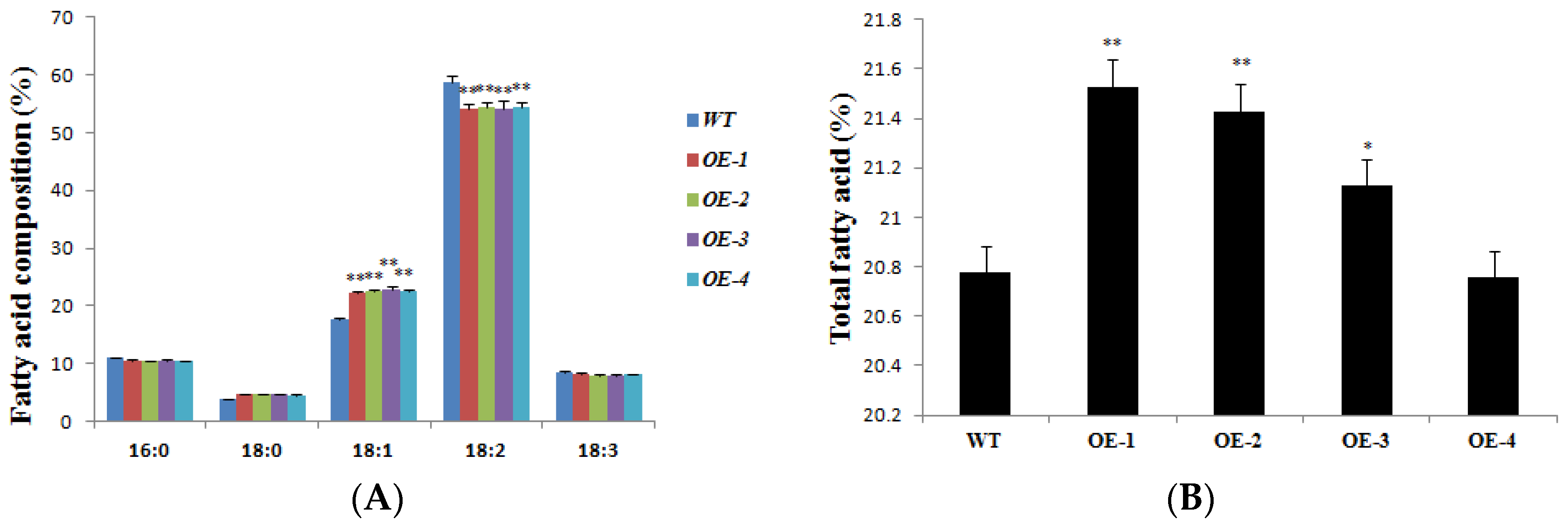
Figure 6.
Phenotypic observation of AhDGAT3 transgenic soybean and the WT. (A) Three AhDGAT3 transgenic lines (OE-1 to OE-3) and the WT in mature period. (B) The size of 10-day to 50-day immature pods after flowering in the WT and AhDGAT3 transgenic soybean. (C) The difference of 10-day to 50-day immature pods’ length between the WT and AhDGAT3 transgenic lines. (D) The difference of 10-day to 50-day immature pods’ width between the WT and AhDGAT3 transgenic lines. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference.
Figure 6.
Phenotypic observation of AhDGAT3 transgenic soybean and the WT. (A) Three AhDGAT3 transgenic lines (OE-1 to OE-3) and the WT in mature period. (B) The size of 10-day to 50-day immature pods after flowering in the WT and AhDGAT3 transgenic soybean. (C) The difference of 10-day to 50-day immature pods’ length between the WT and AhDGAT3 transgenic lines. (D) The difference of 10-day to 50-day immature pods’ width between the WT and AhDGAT3 transgenic lines. Error bars indicate the standard error. * p < 0.05 and ** p < 0.01 represent the significant difference.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Gene specific primer pairs and information used for PCR and molecular detection.
| Purpose | Gene Name (Amplification Length) | Accession Number | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|---|
| RT-PCR | AhDGAT3 (1438 bp) | XM_016339125 | AATAGAAATAGAAATGTGATAATGG | ACAAATCAGGCTCTGGAAGTT |
| qRT-PCR | AhDGAT3 (140 bp) | XM_016339125 | AGAATGGAACCGCTATGT | CTCTGCCCTTACTTGCTC |
| β-Tubulin (185 bp) | GMU12286 | GGAAGGCTTTCTTGCATTGGTA | AGTGGCATCCTGGTACTGC | |
| Actin (155 bp) | J01298 | GTCCTTTCAGGAGGTACAACC | CCTTGAAGTATCCTATTGAGC | |
| Detection | Bar (220 bp) | GTCTGCACCATCGTCAACCACTACA | AGACGTACACGGTCGACTCGGCCGT | |
| AhDGAT3 (304 bp) | XM_016339125 | AGAATGGAACCGCTATGT | CTCTGCCCTTACTTGCTC |
Table 2.
AhDGAT3 protein levels of different tissues in JACK and 4 transgenic lines (OE-1 to OE-4). Data represent the average values of three biological replicates. NA means that the value is not detected and calculated through the standard curve, and there were no detection results.
Table 2.
AhDGAT3 protein levels of different tissues in JACK and 4 transgenic lines (OE-1 to OE-4). Data represent the average values of three biological replicates. NA means that the value is not detected and calculated through the standard curve, and there were no detection results.
| Material | JACK | OE-1 (ng/g) | OE-2 (ng/g) | OE-3 (ng/g) | OE-4 (ng/g) |
|---|---|---|---|---|---|
| Nodules | NA | 196.2 ± 10.2 | 220.6 ± 14.3 | 200.5 ± 13.8 | 233.9 ± 14.9 |
| Root | NA | 236.1 ± 19.2 | 389.5 ± 8.7 | 248.2 ± 1.5 | 415.5 ± 20.4 |
| Stem | NA | 203.7 ± 14.9 | 208.7 ± 6.9 | 200.6 ± 7.9 | 317.1 ± 11.7 |
| Leave | NA | 483.7 ± 16.7 | 523.7 ± 14.4 | 511.3 ± 10.2 | 655.1 ± 11.4 |
| Flower | NA | 288.3 ± 27.2 | 385.1 ± 17.7 | 348.6 ± 11.5 | 417.6 ± 10.5 |
| 10-day Pod | NA | 803.8 ± 14.8 | 841.7 ± 9.2 | 816.9 ± 3.8 | 869.5 ± 6.9 |
| 20-day Pod | NA | 676.1 ± 22.9 | 706.6 ± 10.5 | 796.1 ± 4.6 | 787.1 ± 27.1 |
| 30-day Pod | NA | 700.6 ± 15.9 | 804.1 ± 5.4 | 753.5 ± 32.3 | 883.7 ± 8.8 |
| 40-day Pod | NA | 913.2 ± 5 | 985.4 ± 3.8 | 951.4 ± 20.3 | 1014.4 ± 11.2 |
| Seed | NA | 949.3 ± 17.9 | 1096.9 ± 11.1 | 1004.6 ± 16.9 | 1166.1 ± 22.1 |
Table 3.
The insertion region location information of four AhDGAT3 transgenic strains (R01–R04).
| Sample | Chromosome | Integration Sites | Integrated Way |
|---|---|---|---|
| R01 | 3 | 36,280,392 | Single copy |
| R02 | 10 | 19,795,197 | Single copy |
| R03 | 11 | 32,500,049 | Single copy |
| R04 | 14 | 8,451,776 | Single copy |
Table 4.
Agronomic traits in the WT and AhDGAT3 transgenic soybean (OE-1 to 3). The data for sample mean ± standard error (p < 0.01). Significantly different results are indicated by different letters (a, b): a represents no significant difference; b represents significant difference.
Table 4.
Agronomic traits in the WT and AhDGAT3 transgenic soybean (OE-1 to 3). The data for sample mean ± standard error (p < 0.01). Significantly different results are indicated by different letters (a, b): a represents no significant difference; b represents significant difference.
| Material | WT | OE-1 | OE-2 | OE-3 |
|---|---|---|---|---|
| Plant height (cm) | 101.4 ± 0.14 a | 114.1 ± 0.17 b | 109.2 ± 0.21 b | 105.1 ± 0.27 a |
| Effective branch number | 4 ± 0.21 a | 8 ± 0.16 b | 7 ± 0.28 b | 6 ± 0.12 b |
| Pod number per plant | 78 ± 0.14 a | 188 ± 0.11 b | 168 ± 0.18 b | 161 ± 0.27 b |
| Seed number per plant | 155 ± 0.16 a | 316 ± 0.29 b | 293 ± 0.19 b | 289 ± 0.31 b |
| 100-seed weight (g) | 13.5 ± 0.21 a | 16.3 ± 0.22 b | 15.6 ± 0.24 a | 16.9 ± 0.17 b |
| Seed weight per plant (g) | 25.7 ± 0.17 a | 57.2 ± 0.24 b | 49.7 ± 0.26 b | 45.7 ± 0.26 b |
| Main stem node number | 18 ± 0.25 a | 22 ± 0.26 b | 21 ± 0.21 b | 21 ± 0.24 b |
| Podding height (g) | 8.4 ± 0.29 a | 10.1 ± 0.31 a | 9.9 ± 0.31 a | 10.1 ± 0.27 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, Y.; Yan, F.; Liang, Z.; Wang, Y.; Li, J.; Zhao, L.; Yang, X.; Wang, Q.; Liu, J. Overexpression of the Peanut AhDGAT3 Gene Increases the Oil Content in Soybean. Agronomy 2023, 13, 2333. https://doi.org/10.3390/agronomy13092333
AMA Style
Xu Y, Yan F, Liang Z, Wang Y, Li J, Zhao L, Yang X, Wang Q, Liu J. Overexpression of the Peanut AhDGAT3 Gene Increases the Oil Content in Soybean. Agronomy. 2023; 13(9):2333. https://doi.org/10.3390/agronomy13092333
Chicago/Turabian StyleXu, Yang, Fan Yan, Zhengwei Liang, Ying Wang, Jingwen Li, Lei Zhao, Xuguang Yang, Qingyu Wang, and Jingya Liu. 2023. "Overexpression of the Peanut AhDGAT3 Gene Increases the Oil Content in Soybean" Agronomy 13, no. 9: 2333. https://doi.org/10.3390/agronomy13092333
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.