
The Development of Multi-Resistant Rice Restorer Lines and Hybrid Varieties by Pyramiding Resistance Genes against Blast and Brown Planthopper
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
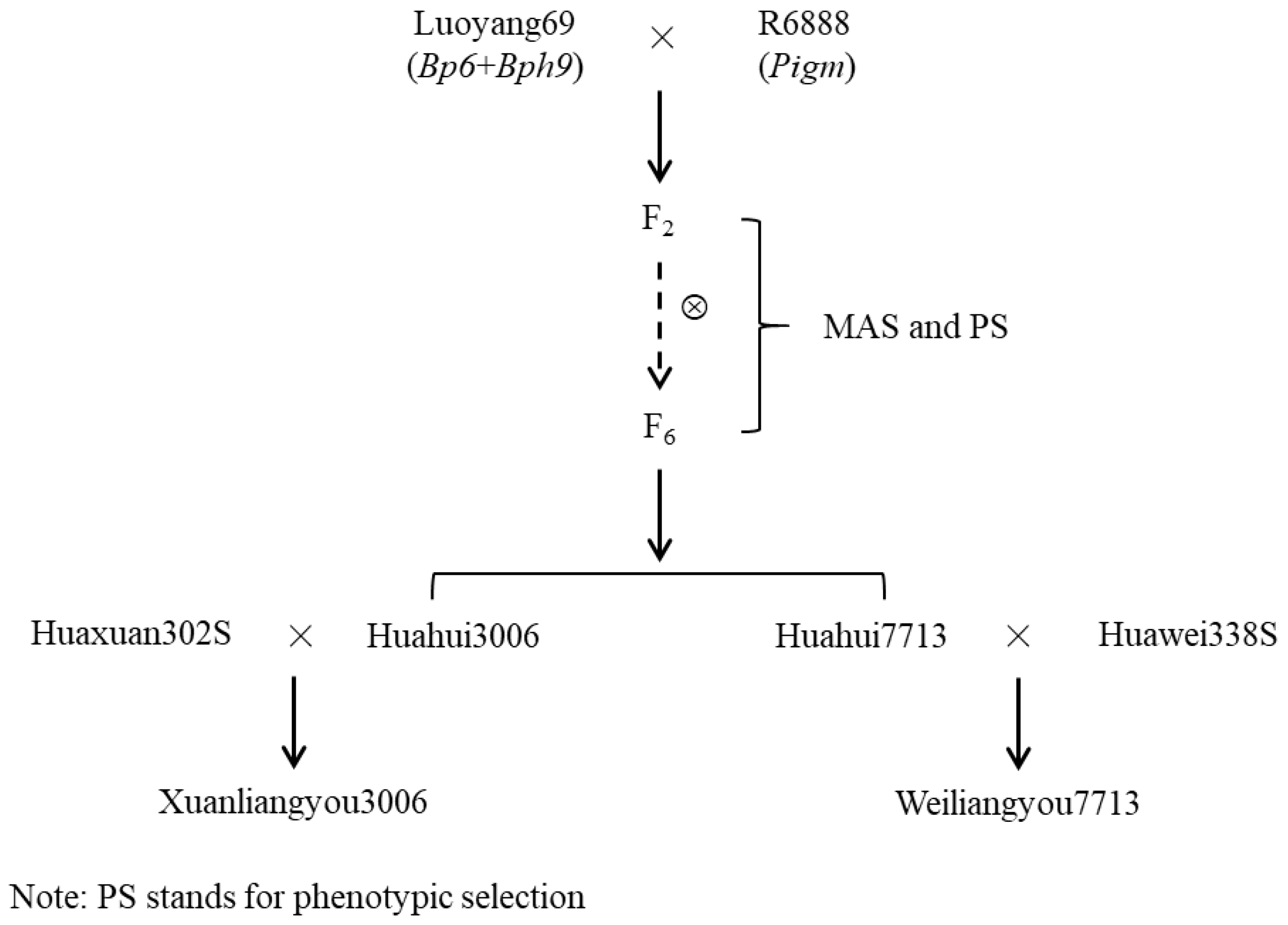
2.1. Plant Materials
2.2. Primer Design of the ARMS Markers of Bph6 and Bph9
2.3. Evaluation of BPH and Blast Resistance
2.4. MAS of New Hybrid Rice Restorer Lines with High-Level BPH and Blast Resistance
2.5. Evaluation of Main Agronomic and Grain-Quality Traits
3. Results
3.1. Development and Validation of Molecular Markers for Bph6 and Bph9
3.2. Development of New Restorer Lines with BPH and Blast Resistance by MAS
3.3. New Hybrid-Rice Variety Development with Blast and BPH Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, P.; Jiang, H.; Luo, L.; Ye, C.; Xiao, Y. Pyramiding of Multiple Genes to Improve Rice Blast Resistance of Photo-Thermo Sensitive Male Sterile Line, without Yield Penalty in Hybrid Rice Production. Plants 2023, 12, 1389. [Google Scholar] [CrossRef] [PubMed]
- Skamnioti, P.; Gurr, S.J. Against the grain: Safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef]
- Yan, L.; Luo, T.; Huang, D.; Wei, M.; Ma, Z.; Liu, C.; Qin, Y.; Zhou, X.; Lu, Y.; Li, R.; et al. Recent Advances in Molecular Mechanism and Breeding Utilization of Brown Planthopper Resistance Genes in Rice: An Integrated Review. Int. J. Mol. Sci. 2023, 24, 12061. [Google Scholar] [CrossRef]
- Cheng, X.; Zhu, L.; He, G. Towards understanding of molecular interactions between rice and the brown planthopper. Mol. Plant 2013, 6, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.; Zhao, Y.; Du, B.; Chen, R.; Zhu, L.; He, G. Genomics of interaction between the brown planthopper and rice. Curr. Opin. Insect Sci. 2017, 19, 82–87. [Google Scholar] [CrossRef]
- Shi, S.; Wang, H.; Nie, L.; Tan, D.; Zhou, C.; Zhang, Q.; Li, Y.; Du, B.; Guo, J.; Huang, J.; et al. Bph30 confers resistance to brown planthopper by fortifying sclerenchyma in rice leaf sheaths. Mol. Plant 2021, 14, 1714–1732. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Kaur, P.; Kishore, A.; Vikal, Y.; Singh, K.; Neelam, K. Recent advances in genomics-assisted breeding of brown planthopper (Nilaparvata lugens) resistance in rice (Oryza sativa). Plant Breed. 2020, 139, 1052–1066. [Google Scholar] [CrossRef]
- Jairin, J.; Phengrat, K.; Teangdeerith, S.; Vanavichit, A.; Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breed. 2007, 19, 35–44. [Google Scholar] [CrossRef]
- Cohen, M.B.; Alam, S.N.; Medina, E.B.; Bernal, C.C. Brown planthopper, Nilaparvata lugens, resistance in rice cultivar IR64: Mechanism and role in successful N. lugens management in Central Luzon. Philippines. Entomol. Exp. Appl. 1997, 85, 221–229. [Google Scholar] [CrossRef]
- Myint, K.K.M.; Fujita, D.; Matsumura, M.; Sonoda, T.; Yoshimura, A.; Yasui, H. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor. Appl. Genet. 2012, 124, 495–504. [Google Scholar] [CrossRef]
- Hu, J.; Cheng, M.; Gao, G.; Zhang, Q.; Xiao, J.; He, Y. Pyramiding and evaluation of three dominant brown planthopper resistance genes in the elite indica rice 9311 and its hybrids. Pest Manag. Sci. 2013, 69, 802–808. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, P.K.; Arya, M.; Singh, N.K.; Singh, U.S. Molecular screening of blast resistance genes in rice using SSR markers. Plant Pathol. J. 2015, 31, 12. [Google Scholar] [CrossRef] [PubMed]
- Devanna, B.N.; Jain, P.; Solanke, A.U.; Das, A.; Thakur, S.; Singh, P.K.; Kumari, M.; Dubey, H.; Jaswal, R.; Pawar, D.; et al. Understanding the Dynamics of Blast Resistance in Rice-Magnaporthe oryzae Interactions. J. Fungi 2022, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, Y.; Abe, Y.; Takagi, H.; Abe, A.; Shimizu, M.; Ito, K.; Kanzaki, E.; Oikawa, K.; Kourelis, J.; Langner, T.; et al. Disentangling the complex gene interaction networks between rice and the blast fungus identifies a new pathogen effector. PLoS Biol. 2023, 21, e3001945. [Google Scholar] [CrossRef]
- Nogoy, F.M.; Song, J.Y.; Ouk, S.; Rahimi, S.; Kwon, S.W.; Kang, K.K.; Cho, Y.G. Current applicable DNA markers for marker assisted breeding in abiotic and biotic stress tolerance in rice (Oryza sativa L.). Plant Breed. Biotechnol. 2016, 4, 271–284. [Google Scholar] [CrossRef]
- Zhao, H.; Yao, W.; Ouyang, Y.; Yang, W.; Wang, G.; Lian, X.; Xing, Y.; Chen, L.; Xie, W. RiceVarMap: A comprehensive database of rice genomic variations. Nucleic Acids Res. 2015, 43, D1018–D1022. [Google Scholar] [CrossRef]
- Ye, S.; Dhillon, S.; Ke, X.; Collins, A.R.; Day, I.N. An efficient procedure for genotyping single nucleotide polymorphisms. Nucleic Acids Res. 2001, 29, e88. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, G.; Sivaranjani, A.K.P.; Pandey, M.K.; Sakthivel, K.; Rani, N.S.; Sudarshan, I.; Prasad, G.S.V.; Neeraja, C.N.; Sundaram, R.M.; Viraktamath, B.C.; et al. Development of a PCR-based SNP marker system for effective selection of kernel length and kernel elongation in rice. Mol. Breed. 2010, 26, 735–740. [Google Scholar] [CrossRef]
- Ramkumar, G.; Prahalada, G.D.; Hechanova, S.L.; Vinarao, R.; Jena, K.K. Development and validation of SNP-based functional co-dominant markers for two major disease resistance genes in rice (O. sativa L.). Mol. Breed. 2015, 35, 1–11. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, A.; Wang, F.; Wang, J.; Bi, J.; Kong, D.; Zhang, F.; Luo, L.; Liu, G.; Yu, X. Development and validation of a PCR-based functional marker system for identifying the low amylose content-associated gene Wxhp in rice. Breed. Sci. 2019, 69, 702–706. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, J.; Wang, Z.; Jing, S.; Wang, Y.; Ouyang, Y.; Cai, B.; Xin, X.; Liu, X.; Zhang, C.; et al. Allelic diversity in an NLR gene BPH9 enables rice to combat planthopper variation. Proc. Natl. Acad. Sci. USA 2016, 113, 12850–12855. [Google Scholar] [CrossRef]
- Guo, J.; Xu, C.; Wu, D.; Zhao, Y.; Qiu, Y.; Wang, X.; Ouyang, Y.; Cai, B.; Liu, X.; Jing, S.; et al. Bph6 encodes an exocyst-localized protein and confers broad resistance to planthoppers in rice. Net. Genet. 2018, 50, 297–306. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Liu, H.; Zeng, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Marker assisted pyramiding of Bph6 and Bph9 into elite restorer line 93-11 and development of functional marker for Bph9. Rice 2017, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Du, Y.; Yu, J.; Zhang, R.; Yu, M.; Cao, H.; Song, T.; Pan, X.; Liang, D.; Liu, Y. Molecular Detection and Analysis of Blast Resistance Genes in Rice Main Varieties in Jiangsu Province, China. Agronomy 2023, 13, 157. [Google Scholar] [CrossRef]
- Qin, P.; Jiang, N.; Hu, X.; Zhu, Y.; Deng, Z.; Bai, Z.; Fu, J.; Zhang, X.; Wang, K.; Chen, S.; et al. Breeding a series of new rice restorer lines with blast resistance and low-to-intermediate apparent amylose contents by integrating marker-assisted and phenotypic selection. J. Phytopathol. 2021, 169, 667–677. [Google Scholar] [CrossRef]
- Guo, J.; Wang, H.; Guan, W.; Guo, Q.; Wang, J.; Yang, J.; Peng, Y.; Shan, J.; Gao, M.; Shi, S.; et al. A tripartite rheostat controls self-regulated host plant resistance to insects. Nature 2023, 618, 799–807. [Google Scholar] [CrossRef]
- Jena, K.K.; Kim, S.M. Current status of brown planthopper (BPH) resistance and genetics. Rice 2010, 3, 161–171. [Google Scholar] [CrossRef]
- Kim, J.; An, X.; Yang, K.; Miao, S.; Qin, Y.; Hu, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Molecular Mapping of a New Brown Planthopper Resistance Gene Bph43 in Rice (Oryza sativa L.). Agronomy 2022, 12, 808. [Google Scholar] [CrossRef]
- Hu, J.; Li, X.; Wu, C.; Yang, C.; Hua, H.; Gao, G.; Xiao, J.; He, Y. Pyramiding and evaluation of the brown planthopper resistance genes Bph14 and Bph15 in hybrid rice. Mol. Breed. 2012, 29, 61–69. [Google Scholar] [CrossRef]
- Jiang, H.; Feng, Y.; Bao, L.; Li, X.; Gao, G.; Zhang, Q.; He, Y. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding. Mol. Breed. 2012, 30, 1679–1688. [Google Scholar] [CrossRef]
- Wang, H.; Ye, S.; Mou, T. Molecular breeding of rice restorer lines and hybrids for brown planthopper (BPH) resistance using the Bph14 and Bph15 genes. Rice 2016, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ma, T.; Zhang, A.; Ong, K.; Luo, Z.; Li, Z.; Yang, J.; Yin, Z. Marker-assisted breeding of Chinese elite rice cultivar 9311 for disease resistance to rice blast and bacterial blight and tolerance to submergence. Mol. Breed. 2017, 37, 106. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, Z.; Lang, X.; Du, B.; Liu, K.; Yang, G.; Hu, G.; Li, S.; He, G.; You, A. Development and validation of a PCR-based functional marker system for the brown planthopper resistance gene Bph14 in rice. Breed. Sci. 2013, 63, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Chen, Z.; Chen, Z.; Zhou, Y.; Wang, Z.; Wang, F.; Chen, S. Allele-specific marker-based assessment revealed that the rice blast resistance genes Pi2 and Pi9 have not been widely deployed in Chinese indica rice cultivars. Rice 2016, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Pranathi, K.; Viraktamath, B.C.; Neeraja, C.N.; Balachandran, S.M.; Hariprasad, A.S.; Koteswara Rao, P.; Revathi, P.; Senguttuvel, P.; Hajira, S.K.; Balachiranjeevi, C.H.; et al. Development and validation of candidate gene-specific markers for the major fertility restorer genes, Rf4 and Rf3 in rice. Mol. Breed. 2016, 36, 145. [Google Scholar] [CrossRef]
- Yap, R.; Hsu, Y.C.; Wu, Y.; Lin, Y.; Kuo, C. Multiplex PCR genotyping for five bacterial blight resistance genes applied to marker-assisted selection in rice (Oryza sativa). Plant Breed. 2016, 135, 309–317. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.; Li, M.; Zhao, W.; Sun, H.; Zhao, Q. A multiplex PCR system for detection of wide compatibility allele S5-n and erect panicle allele dep1 in rice. Crop Breed. Appl. Biot. 2017, 17, 250–258. [Google Scholar] [CrossRef]
- Hayashi, K.; Yoshida, H.; Ashikawa, I. Development of PCR-based allele-specific and InDel marker sets for nine rice blast resistance genes. Theor. Appl. Genet. 2006, 113, 251–260. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Marker Name | Primer Name | Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|---|
| Bph6 | B6 | SF | AGGGCCTCTGGCGCTCTAC | 361(−)/283(+) |
| SR | AATGTGAAAGTGCAATTAGAAGGT | |||
| RF | ATAGTGAAGTTGAATCCGAAGG | |||
| RR | AGTGACTCAGCCTTGTGTTTCG | |||
| Bph9 | B9 | IF | ACCATTGTTAGGCAGTTGTTCA | 316(−)/230(+) |
| OR | ATTCGACTCCCTTTCTTGTTATCT | |||
| OF | AATGTCGCACCCAGCAGC | |||
| IR | CAGCCTCCTGAAGAGATCTTTCA | |||
| BPH1/9 | B1/9 | SF | GGCGAGTTGATGAAATAAAGG | 473(−)/545(+) |
| SR | GTTCTTTGCATCGCTGTCTC | |||
| RR | TTGCTCCCACCGAAGTCA | |||
| RF | AGGGAATGGTGTAGTAGCA | |||
| Pigm | GM | OF | TGCTGAACAAGGTGTAGAGGTA | 448(−)/621(+) |
| OR | GCCAGGTCCTACTTCCACAAAA | |||
| IF | GTGACATCCAGTCCTACACAATCTAA | |||
| IR | CACGGAACTCGTTTTCGTC |
| Lines | Genes of Pyramiding | Number of Lines | Seedling BPH Resistance Score | Seedling Blast Resistance Score |
|---|---|---|---|---|
| Gm69 lines | Bph6 + Bph9 + Pigm | 6 | 2.75 ± 0.37 c | 3.76 ± 0.41 a |
| Gm6 lines | Bph6 + Pigm | 8 | 3.56 ± 0.54 b | 3.62 ± 0.42 a |
| Gm9 lines | Bph9 + Pigm | 10 | 3.26 ± 0.19 b | 3.43 ± 0.52 a |
| R6888 | Pigm | 8.20 ± 0.26 a | 3.63 ± 0.41 a | |
| Luoyang69 | Bph6 + Bph9 | 2.73 ± 0.15 c | 8.23 ± 0.15 b | |
| 9311 | 7.90 ± 0.16 a | 8.33 ± 0.33 b |
| Line | GY (t/ha) | DTH (d) | PH (cm) | PL (cm) | NGP | SF (%) | TGW (g) | AAC (%) | GC (mm) | GT | HRR (%) | LWR | CD (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Huahui3006 | 8.43 ± 0.2 b | 125 ± 0.6 c | 114.7 ± 2.6 b | 23.8 ± 0.7 a | 220.6 ± 27.4 b | 88.9 ± 1.9 a | 27.5 ± 1.9 a | 16.1 ± 0.1 c | 62 ± 2.0 b | 6.0 ± 0.2 a | 64.2 ± 1.2 b | 3.4 ± 0.08 a | 3.6 ± 0.2 c |
| Huahui7713 | 8.67 ± 0.2 ab | 126 ± 1.4 b | 114.3 ± 1.2 b | 23.2 ± 0.9 b | 230.6 ± 9.7 b | 88.2 ± 1.8 a | 27.8 ± 1.8 a | 16.4 ± 0.1 b | 63 ± 1.7 ab | 6.0 ± 0.2 a | 63.1 ± 2.5 b | 3.3 ± 0.03 b | 4.6 ± 0.3 b |
| R6888 | 8.31 ± 0.2 b | 129 ± 1.2 a | 117.6 ± 1.6 a | 22.3 ± 1.0 b | 258.2 ± 20.2 a | 85.8 ± 1.6 b | 27.3 ± 1.1 a | 15.5 ± 0.2 d | 65 ± 1.5 a | 3.0 ± 0.1 b | 64.1 ± 1.8 b | 3.3 ± 0.07 b | 4.41 ± 0.3 b |
| Luoyang69 | 8.52 ± 0.3 a | 120 ± 0.8 d | 112.3 ± 3.0 b | 23.8 ± 0.9 a | 171.4 ± 14.2 c | 88.01 ± 1.1 ab | 29.4 ± 1.4 a | 17.3 ± 0.2 a | 65 ± 1.4 a | 6.2 ± 0.3 a | 69.5 ± 1.7 a | 2.9 ± 0.12 c | 7.05 ± 0.2 a |
| Line | Genotype | Blast Resistance | BPH Resistance | |||||
|---|---|---|---|---|---|---|---|---|
| Bph6 | Bph9 | Pigm | Seedling | Panicle Blast Severity Index | Incidence of Infected Panicles | Integrated Resistance Index | ||
| R6888 | − | − | + | 3 | 3 | 3 | 3 | HS |
| Luoyang69 | + | + | − | 9 | 9 | 9 | 9 | R |
| Huahui3006 | + | + | + | 3 | 3 | 3 | 3 | R |
| Huahui7713 | + | + | + | 3 | 3 | 3 | 3 | R |
| XuanLiangYou3006 | +/− | +/− | +/− | 3 | 3 | 3 | 3 | R |
| WeiLiangYou7713 | +/− | +/− | +/− | 3 | 3 | 3 | 3 | R |
| Variety | Year | Agronomic Traits | Grain Quality Traits | Blast Resistance | BPH Resistance | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| GY (t/ha) | DTM (d) | AAC (%) | GC (mm) | CD (%) | HRR (%) | IDI | HSPBS | |||
| Weiliangyou7713 | 2019 | 10.22 (CK + 6.27%) | 138.0 (CK + 4.6) | 15 | 82 | 0.8 | 68.9 | 3.5 | 5 | R |
| 2020 | 9.61 (CK + 8.76%) | 135.9 (CK + 2.8) | 14.3 | 82 | 0.9 | 65.9 | 3 | 5 | R | |
| Xuanliangyou3006 | 2020 | 9.13 (CK + 3.24%) | 137.5 (CK + 5.6) | 16.8 | 60 | 1.4 | 66.3 | 3.4 | 3 | MR |
| 2021 | 9.39 (CK + 1.89%) | 139.0 (CK + 8.1) | 15.4 | 61 | 7 | 64.9 | 2.6 | 3 | MR | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.; Qin, P.; Liu, K.; Jiang, N.; Yan, T.; Zhang, X.; Fu, C.; He, G.; Wang, K.; Yang, Y. The Development of Multi-Resistant Rice Restorer Lines and Hybrid Varieties by Pyramiding Resistance Genes against Blast and Brown Planthopper. Agronomy 2024, 14, 878. https://doi.org/10.3390/agronomy14050878
Deng Z, Qin P, Liu K, Jiang N, Yan T, Zhang X, Fu C, He G, Wang K, Yang Y. The Development of Multi-Resistant Rice Restorer Lines and Hybrid Varieties by Pyramiding Resistance Genes against Blast and Brown Planthopper. Agronomy. 2024; 14(5):878. https://doi.org/10.3390/agronomy14050878
Chicago/Turabian StyleDeng, Zhao, Peng Qin, Kaiyu Liu, Nan Jiang, Tianze Yan, Xuanwen Zhang, Chenjian Fu, Guangcun He, Kai Wang, and Yuanzhu Yang. 2024. "The Development of Multi-Resistant Rice Restorer Lines and Hybrid Varieties by Pyramiding Resistance Genes against Blast and Brown Planthopper" Agronomy 14, no. 5: 878. https://doi.org/10.3390/agronomy14050878