
Identification of Barley (Hordeum vulgare L.) Autophagy Genes and Their Expression Levels during Leaf Senescence, Chronic Nitrogen Limitation and in Response to Dark Exposure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. DNA and Protein Sequence Analysis
2.3. RNA Purification and RT-qPCR Analysis
2.4. HvATG5 Cloning and Agrobacterium tumefaciens-Mediated Transformation
2.5. Chlorophyll Content Determination
2.5.1. Concanamycin A Treatment
2.5.2. Statistics
3. Results
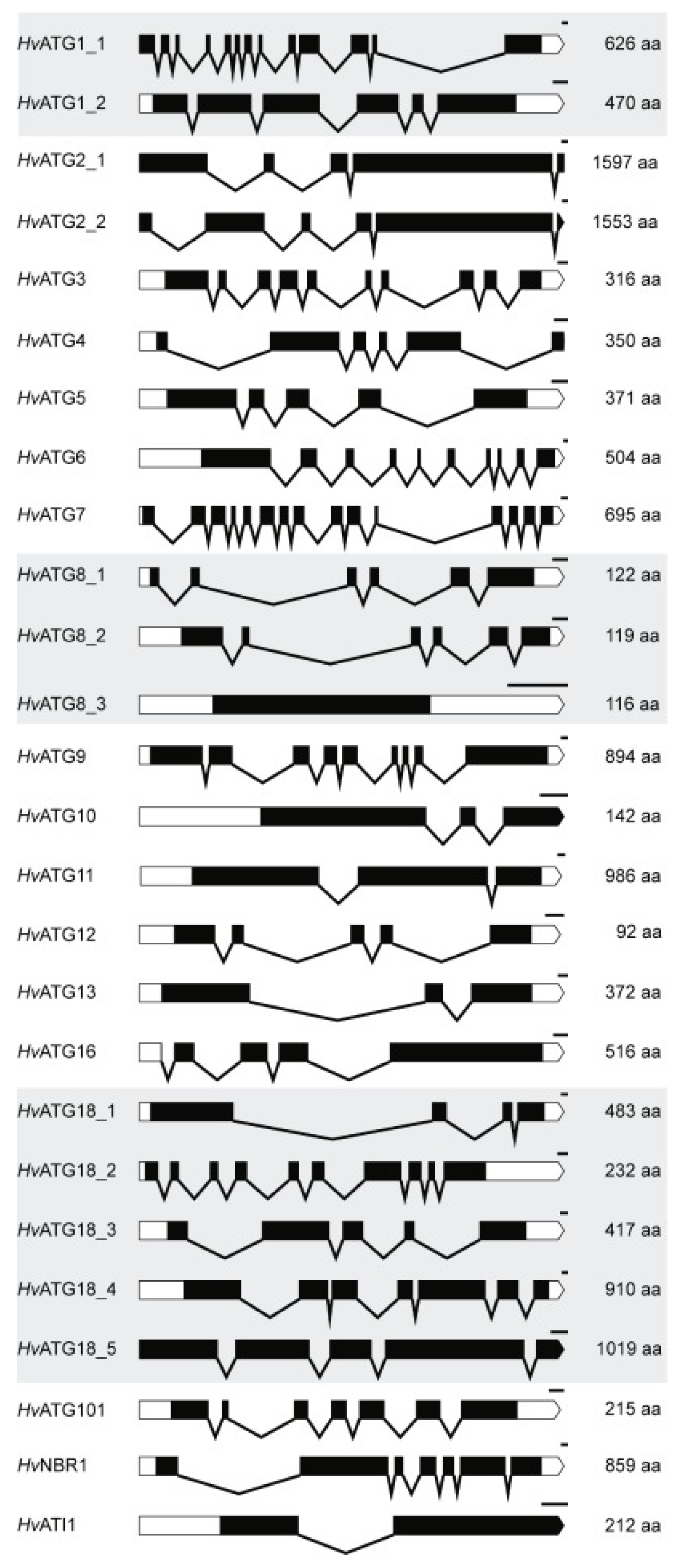
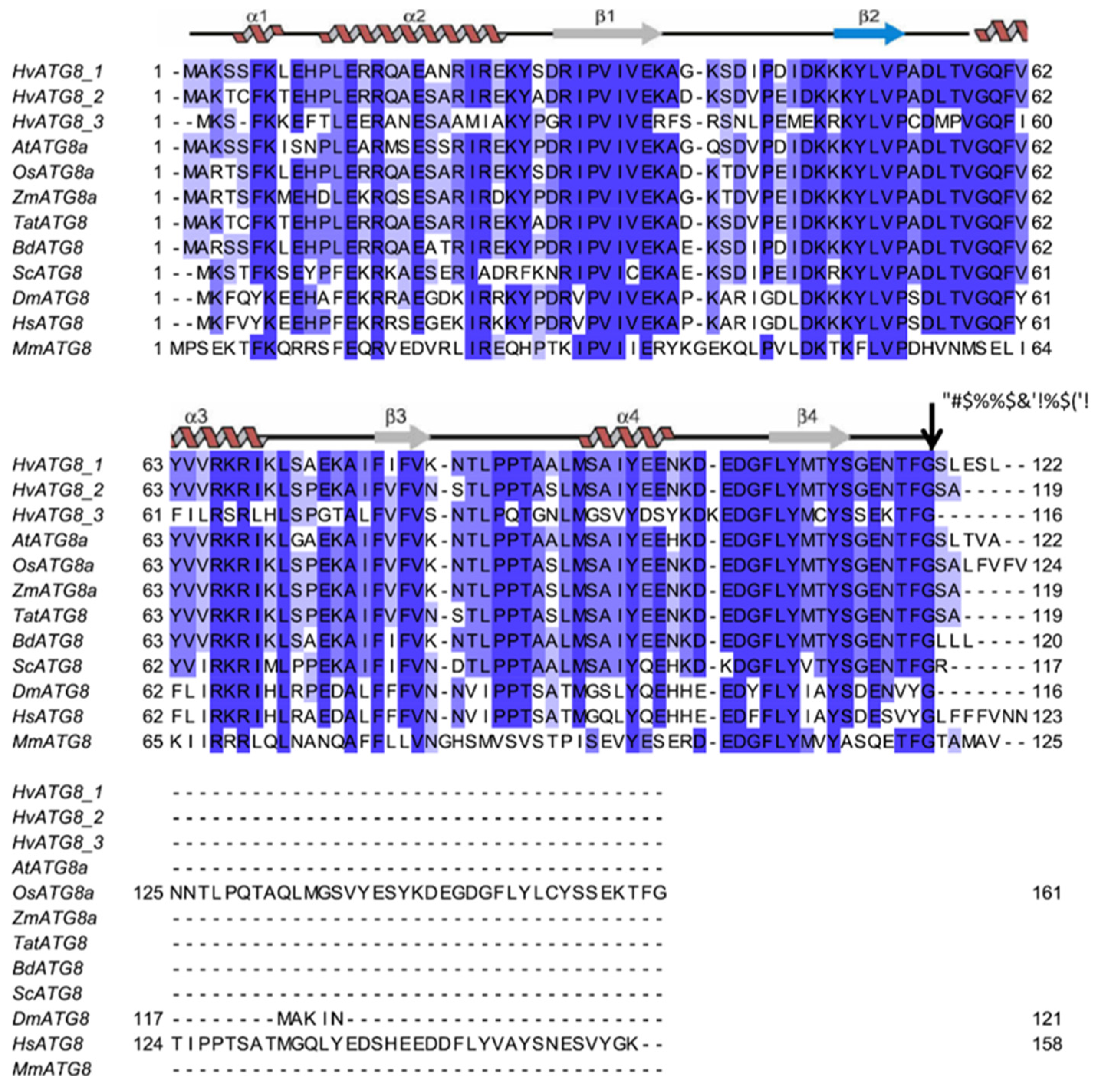
3.1. Identification of ATG, NBR1 and ATI Genes in Barley
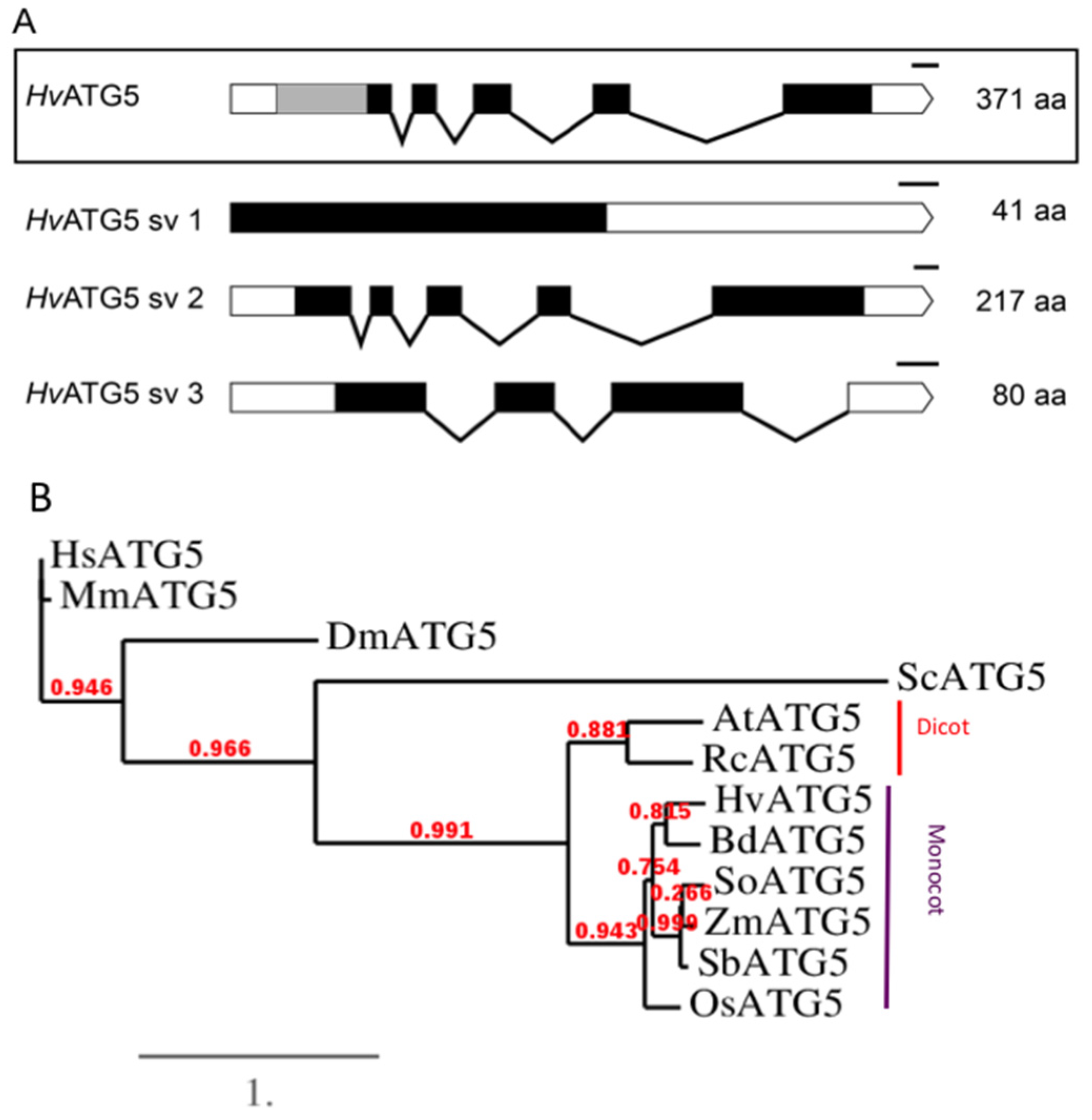
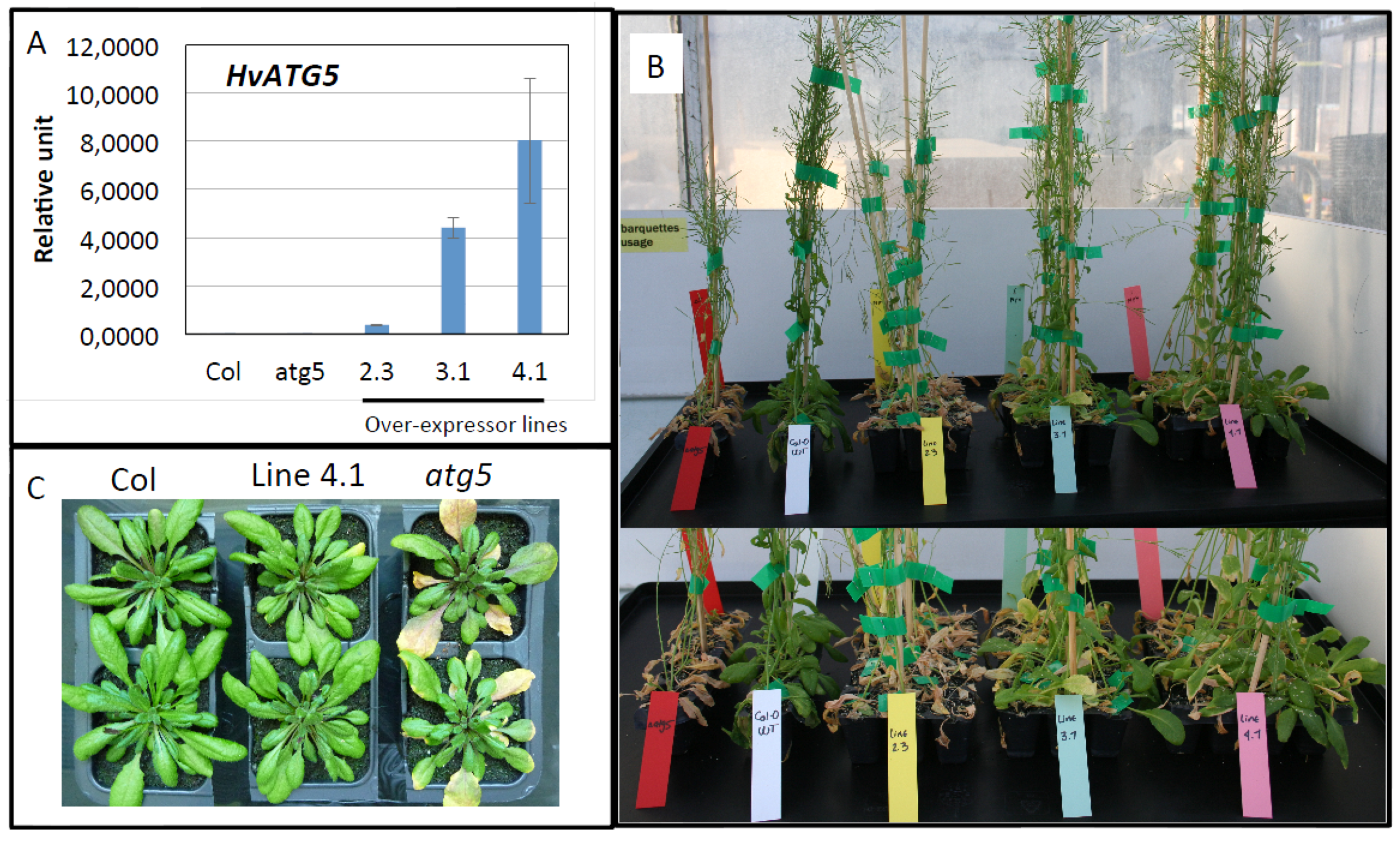
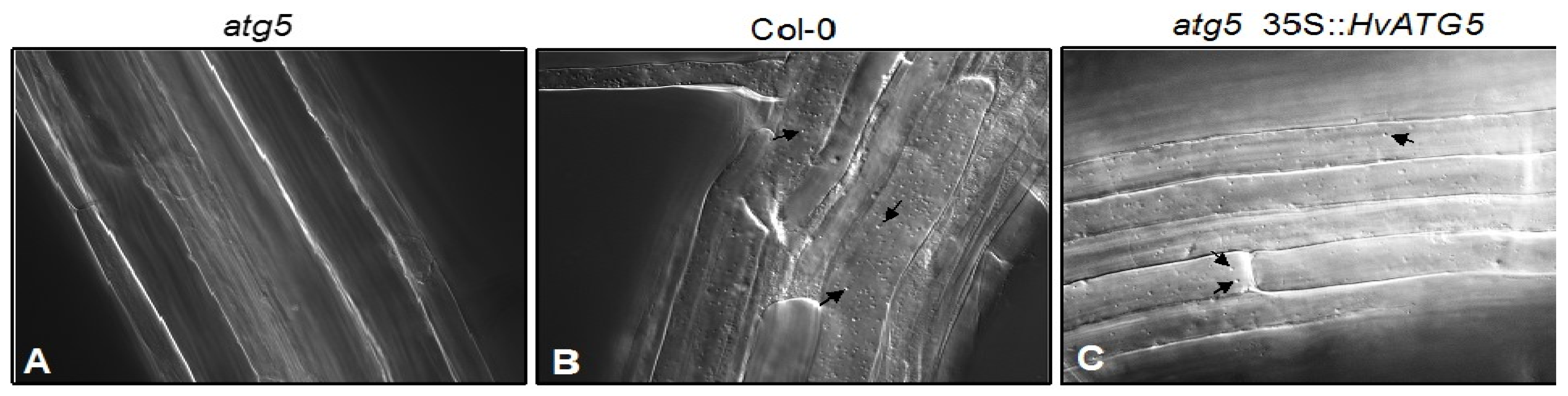
3.2. HvATG5 Description and Functional Complementation
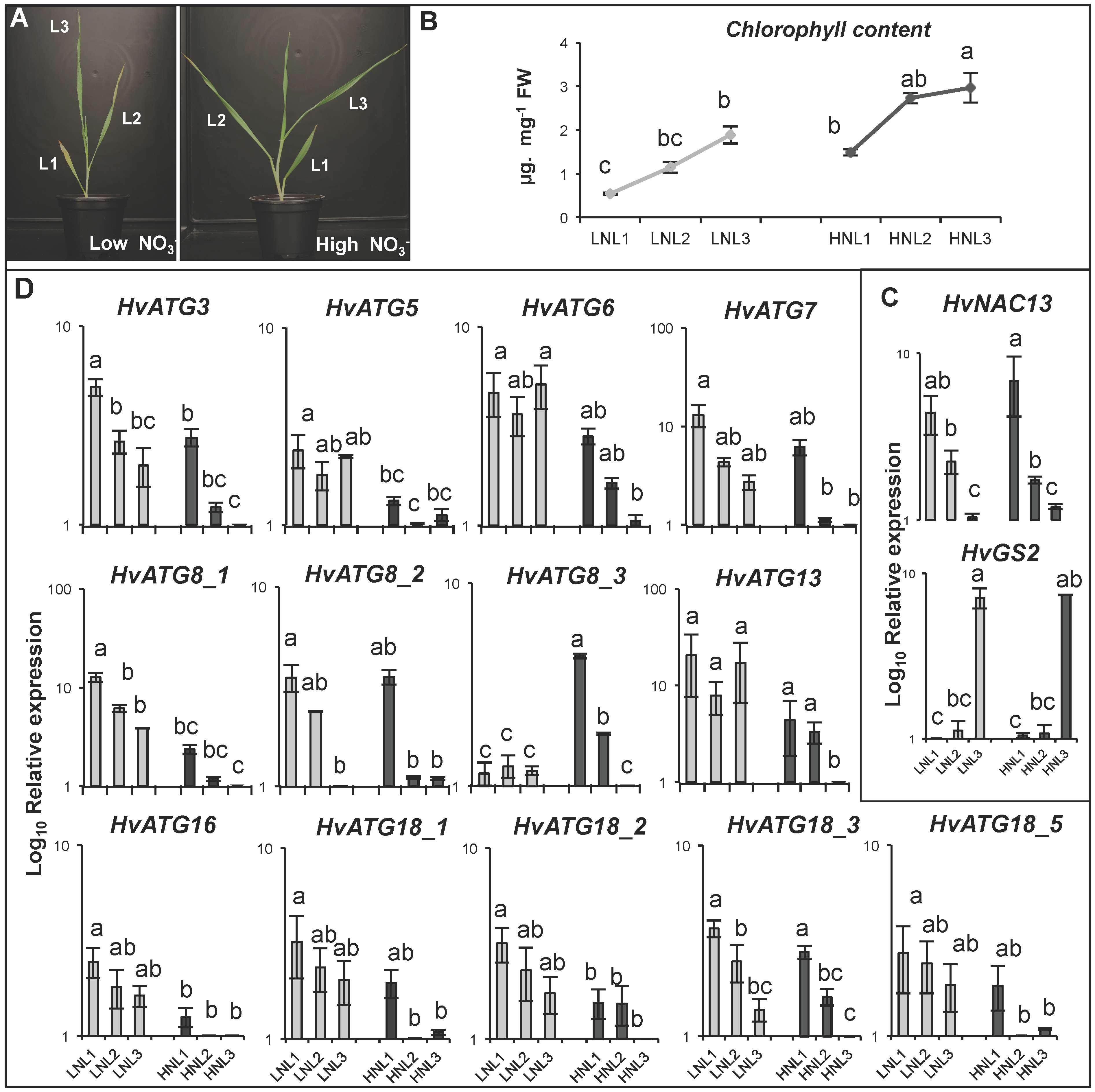
3.3. Expression Patterns of HvATG Genes during Leaf Senescence and Nutritional Stress
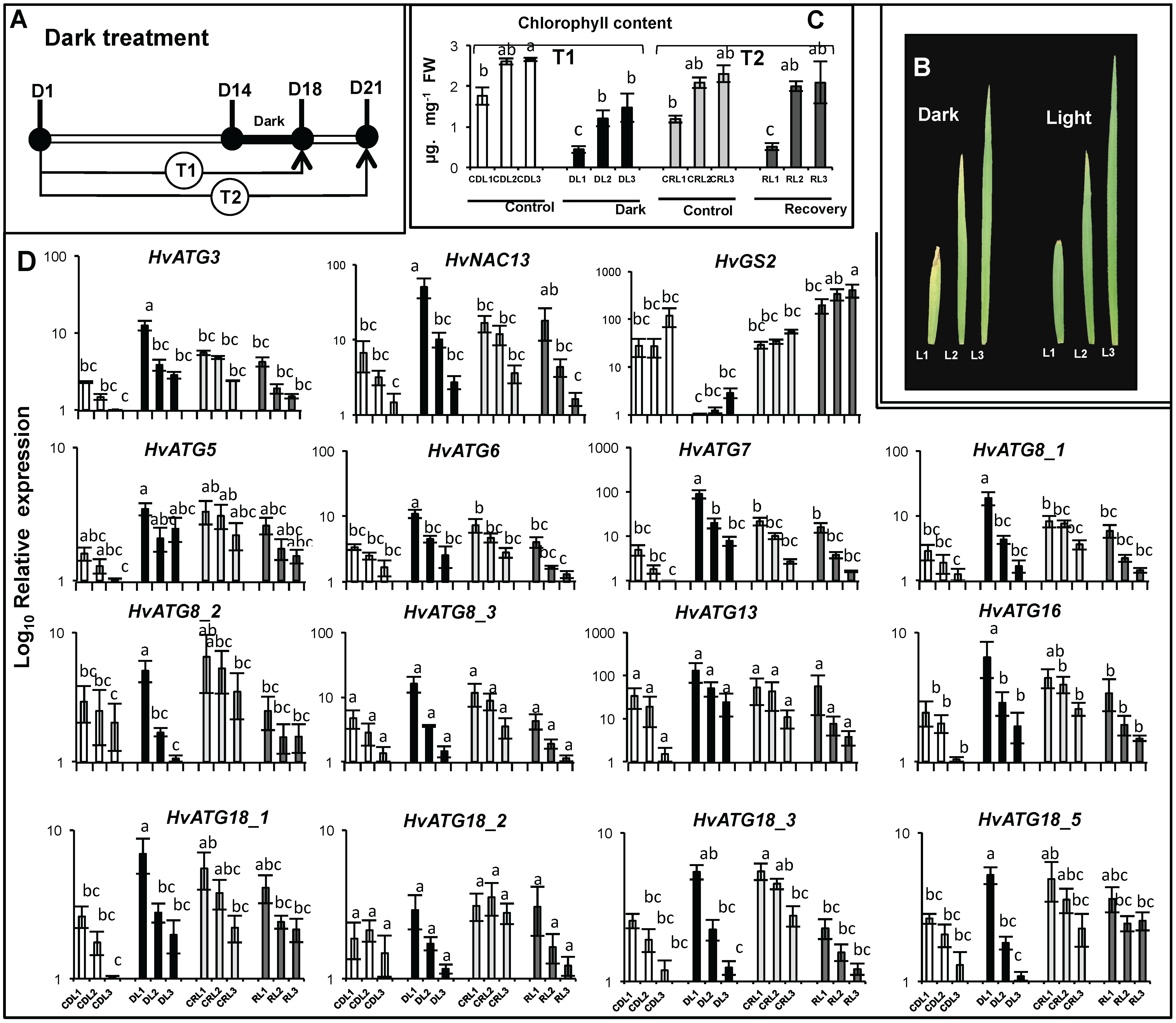
3.4. HvATG Transcript Levels in Seedling Leaves During Senescence and in Response to Nitrate Shortage or Dark Treatment
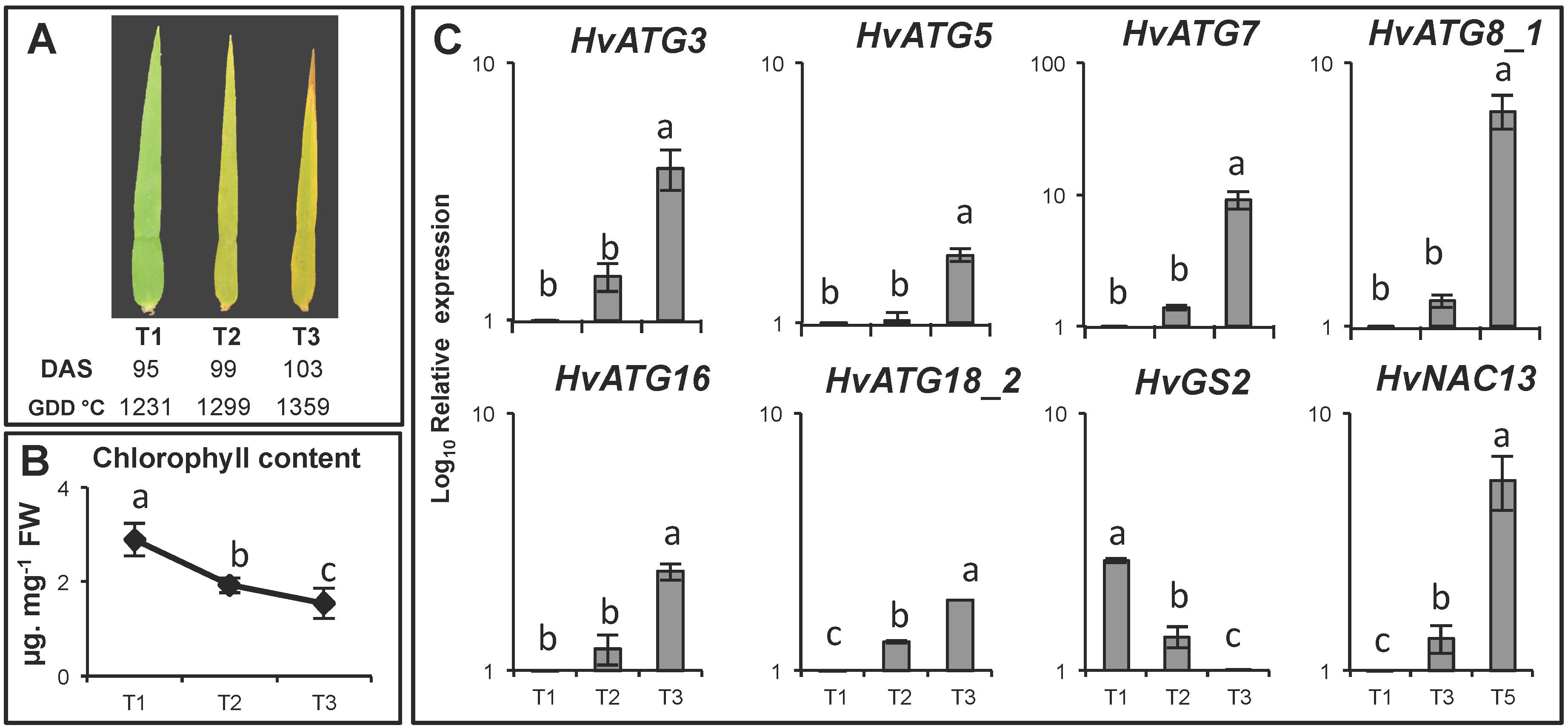
3.5. HvATG Transcript Levels in the Flag Leaf During Senescence
4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tsukada, M.; Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of saccharomyces-cerevisiae. Febs Lett. 1993, 333, 169–174. [Google Scholar] [CrossRef]
- Baba, M.; Takeshige, K.; Baba, N.; Ohsumi, Y. Ultrastructural analysis of the autophagic process in yeast—Detection of autophagosomes and their characterization. J. Cell Biol. 1994, 124, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Takeshige, K.; Baba, M.; Tsuboi, S.; Noda, T.; Ohsumi, Y. Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J. Cell Biol. 1992, 119, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Doelling, J.H.; Walker, J.M.; Friedman, E.M.; Thompson, A.R.; Vierstra, R.D. The apg8/12-activating enzyme apg7 is required for proper nutrient recycling and senescence in arabidopsis thaliana. J. Biol. Chem. 2002, 277, 33105–33114. [Google Scholar] [CrossRef] [PubMed]
- Hanaoka, H.; Noda, T.; Shirano, Y.; Kato, T.; Hayashi, H.; Shibata, D.; Tabata, S.; Ohsumi, Y. Leaf senescence and starvation-induced chlorosis are accelerated by the disruption of an arabidopsis autophagy gene. Plant Physiol. 2002, 129, 1181–1193. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Contento, A.L.; Bassham, D.C. Atatg18a is required for the formation of autophagosomes during nutrient stress and senescence in arabidopsis thaliana. Plant J. 2005, 42, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Chung, T.J.; Suttangkakul, A.; Vierstra, R.D. The ATG autophagic conjugation system in maize: Atg transcripts and abundance of the atg8-lipid adduct are regulated by development and nutrient availability. Plant Physiol. 2009, 149, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Liu, T.; Ouyang, J.; Wang, R.; Fan, T.; Zhang, M. Genome-wide identification, classification, and expression analysis of autophagy-associated gene homologues in rice (oryza sativa l.). DNA Res. 2011, 18, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.R.; Doelling, J.H.; Suttangkakul, A.; Vierstra, R.D. Autophagic nutrient recycling in arabidopsis directed by the ATG8 and ATG12 conjugation pathways. Plant Physiol. 2005, 138, 2097–2110. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Vierstra, R.D. Autophagy: A multifaceted intracellular system for bulk and selective recycling. Trends Plant Sci. 2012, 17, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bassham, D.C. Autophagy: Pathways for self-eating in plant cells. Ann. Rev. Plant Biol. 2012, 63, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Svenning, S.; Lamark, T.; Krause, K.; Johansen, T. Plant NBR1 is a selective autophagy substrate and a functional hybrid of the mammalian autophagic adapters NBR1 and p62/sqstm1. Autophagy 2011, 7, 993–1010. [Google Scholar] [CrossRef] [PubMed]
- Zientara-Rytter, K.; Lukomska, J.; Moniuszko, G.; Gwozdecki, R.; Surowiecki, P.; Lewandowska, M.; Liszewska, F.; Wawrzynska, A.; Sirko, A. Identification and functional analysis of JOKA2, a tobacco member of the family of selective autophagy cargo receptors. Autophagy 2011, 7, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Honig, A.; Avin-Wittenberg, T.; Galili, G. Selective autophagy in the aid of plant germination and response to nutrient starvation. Autophagy 2012, 8, 838–839. [Google Scholar] [CrossRef] [PubMed]
- Avin-Wittenberg, T.; Honig, A.; Galili, G. Variations on a theme: Plant autophagy in comparison to yeast and mammals. Protoplasma 2012. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, S.; Avin-Wittenberg, T.; Galili, G. Involvement of autophagy in the direct er to vacuole protein trafficking route in plants. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Guiboileau, A.; Avila-Ospina, L.; Yoshimoto, K.; Soulay, F.; Azzopardi, M.; Marmagne, A.; Lothier, J.; Masclaux-Daubresse, C. Physiological and metabolic consequences of autophagy defisciency for the management of nitrogen and protein resources in arabidopsis leaves depending on nitrate availability. New Phytol. 2013, 199, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Guiboileau, A.; Yoshimoto, K.; Soulay, F.; Bataillé, M.; Avice, J.; Masclaux-Daubresse, C. Autophagy machinery controls nitrogen remobilization at the whole-plant level under both limiting and ample nitrate conditions in arabidopsis. New Phytol. 2012, 194, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chung, T.; Pennington, J.G.; Federico, M.L.; Kaeppler, H.F.; Kaeppler, S.M.; Otegui, M.S.; Vierstra, R.D. Autophagic recycling plays a central role in maize nitrogen remobilization. Plant cell 2015, 27, 1389–1408. [Google Scholar] [CrossRef] [PubMed]
- Wada, S.; Hayashida, Y.; Izumi, M.; Kurusu, T.; Hanamata, S.; Kanno, K.; Kojima, S.; Yamaya, T.; Kuchitsu, K.; Makino, A.; et al. Autophagy supports biomass production and nitrogen use efficiency at the vegetative stage in rice. Plant Physiol. 2015, 168, U60–U721. [Google Scholar] [CrossRef] [PubMed]
- Kurusu, T.; Koyano, T.; Hanamata, S.; Kubo, T.; Noguchi, Y.; Yagi, C.; Nagata, N.; Yamamoto, T.; Ohnishi, T.; Okazaki, Y.; et al. Osatg7 is required for autophagy-dependent lipid metabolism in rice postmeiotic anther development. Autophagy 2014, 10, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Merkulova, E.A.; Guiboileau, A.; Naya, L.; Masclaux-Daubresse, C.; Yoshimoto, K. Assessment and optimization of autophagy monitoring methods in arabidopsis roots indicate direct fusion of autophagosomes with vacuoles. Plant Cell Physiol. 2014, 55, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Clément, G.; Anne, P.; Routaboul, J.; Guiboileau, A.; Soulay, F.; Shirasu, K.; Yoshimoto, K. Stitching together the multiple dimensions of autophagy using metabolomic and transcriptomic analyses reveals new impacts of autophagy defects on metabolism, development and plant response to environment. Plant Cell 2014, 26, 1857–1877. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.; Waugh, R.; Langridge, P.; Close, T.; Wise, R.; Graner, A.; Matsumoto, T.; Sato, K.; Schulman, A.; Muehlbaue, G.; et al. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–717. [Google Scholar]
- Altschul, S.F.; Lipman, D.J. Protein database searches for multiple alignments. Proc. Natl. Acad. Sci. USA 1990, 87, 5509–5513. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Higgins, D.; Gibson, T. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenol oxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Noda, N.N.; Ohsumi, Y.; Inagaki, F. Atg8-family interacting motif crucial for selective autophagy. Febs Lett. 2010, 584, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Noda, N.N.; Satoo, K.; Fujioka, Y.; Kumeta, H.; Ogura, K.; Nakatogawa, H.; Ohsumi, Y.; Inagaki, F. Structural basis of ATG8 activation by a homodimeric E1, ATG7. Mol. Cell 2011, 44, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Noda, N.N.; Kobayashi, T.; Adachi, W.; Fujioka, Y.; Ohsumi, Y.; Inagaki, F. Structure of the novel c-terminal domain of vacuolar protein sorting 30/autophagy-related protein 6 and its specific role in autophagy. J. Biol. Chem. 2012, 287, 16256–16266. [Google Scholar] [CrossRef] [PubMed]
- Papinski, D.; Schuschnig, M.; Reiter, W.; Wilhelm, L.; Barnes, C.A.; Maiolica, A.; Hansmann, I.; Pfaffenwimmer, T.; Kijanska, M.; Stoffel, I.; et al. Early steps in autophagy depend on direct phosphorylation of Atg9 by the ATG1 kinase. Mol. Cell 2014, 53, 515–515. [Google Scholar] [CrossRef]
- Jao, C.C.; Ragusa, M.J.; Stanley, R.E.; Hurley, J.H. What the n-terminal domain of Atg13 looks like and what it does a horma fold required for ptdins 3-kinase recruitment. Autophagy 2013, 9, 1112–1114. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, Y.; Noda, N.N.; Fujii, K.; Yoshimoto, K.; Ohsumi, Y.; Inagaki, F. In vitro reconstitution of plant Atg8 and Atg12 conjugation systems essential for autophagy. J. Biol. Chem. 2008, 283, 1921–1928. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Hanaoka, H.; Sato, S.; Kato, T.; Tabata, S.; Noda, T.; Ohsumi, Y. Processing of Atg8s, ubiquitin-like proteins, and their deconjugation by Atg4s are essential for plant autophagy. Plant Cell 2004, 16, 2967–2983. [Google Scholar] [CrossRef] [PubMed]
- Avila-Ospina, L.; Marmagne, A.; Talbotec, J.; Krupinska, K.; Masclaux-Daubresse, C. The identification of new cytosolic glutamine synthetase and asparagine synthetase genes in barley (hordeum vulgare l.), and their expression during leaf senescence. J. Exp. Bot. 2015, 66, 2013–2026. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.R.; Suttangkakul, A.; Vierstra, R.D. The Atg12-conjugating enzyme Atg10 is essential for autophagic vesicle formation in arabidopsis thaliana. Genetics 2008, 178, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Slavikova, S.; Ufaz, S.; Avin-Wittenberg, T.; Levanony, H.; Galili, G. An autophagy-associated Atg8 protein is involved in the responses of arabidopsis seedlings to hormonal controls and abiotic stresses. J. Exp. Bot. 2008, 59, 4029–4043. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Contento, A.L.; Nguyen, P.Q.; Bassham, D.C. Degradation of oxidized proteins by autophagy during oxidative stress in arabidopsis. Plant Physiol. 2007, 143, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Contento, A.L.; Bassham, D.C. Disruption of autophagy results in constitutive oxidative stress in arabidopsis. Autophagy 2007, 3, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xiong, Y.; Bassham, D.C. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy 2009, 5, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Jikumaru, Y.; Kamiya, Y.; Kusano, M.; Consonni, C.; Panstruga, R.; Ohsumi, Y.; Shirasu, K. Autophagy negatively regulates cell death by controlling NPR1-dependent salicylic acid signaling during senescence and the innate immune response in arabidopsis. Plant Cell 2009, 21, 2914–2927. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Wang, F.; Zheng, Z.; Fan, B.; Chen, Z. A critical role of autophagy in plant resistance to necrotrophic fungal pathogens. Plant J. 2011, 66, 953–968. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Suzuki, T.; Hattori, M.; Yoshimoto, K.; Ohsumi, Y.; Moriyasu, Y. Atatg genes, homologs of yeast autophagy genes, are involved in constitutive autophagy in arabidopsis root tip cells. Plant Cell Physiol. 2006, 47, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Shpilka, T.; Weidberg, H.; Pietrokovski, S.; Elazar, Z. Atg8: An autophagy-related ubiquitin-like protein family. Genome Biol. 2011, 12, 226. [Google Scholar] [CrossRef] [PubMed]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.-S.; Penfold, C.A.; Jenkins, D.; et al. High-resolution temporal profiling of transcripts during arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [PubMed]
- Avila-Ospina, L.; Moison, M.; Yoshimoto, K.; Masclaux-Daubresse, C. Autophagy, plant senescence, and nutrient recycling. J. Exp. Bot. 2014, 65, 3799–3811. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, J.; Gregersen, P.L.; Krupinska, K. Identification of predominant genes involved in regulation and execution of senescence-associated nitrogen remobilization in flag leaves of field grown barley. J. Exp. Bot. 2014, 65, 2963–3973. [Google Scholar] [CrossRef] [PubMed]
- Rose, T.L.; Bonneau, L.; Der, C.; Marty-Mazars, D.; Marty, F. Starvation-induced expression of autophagy-related genes in arabidopsis. Biol. Cell 2006, 98, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Xiao, D.; Liu, D.; Chai, W.; Gong, Q.; Wang, N.N. Heterologous expression of Atg8c from soybean confers tolerance to nitrogen deficiency and increases yield in arabidopsis. PLoS ONE 2012, 7, e37217. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | BAC Clones | Gene ID EMBL-EBI | Accession No. NCBI or UniProt *** | H.vulgare Genomic seq. | No. of AA Residues | Identity to | ||
|---|---|---|---|---|---|---|---|---|
| S. cerevisiae | A. thaliana * | O. sativa ** | ||||||
| HvATG1_1 | NIASHv2141K06 | MLOC_55869 | AK371799 | x_contig_40347 | 626 | 28% | 45% | 69% |
| HvATG1_2 | - | MLOC_1553 | M0UXZ8 *** | x_contig_113713 | 470 | 18% | 28% | 29% |
| HvATG2_1 | - | MLOC_14121 | M0UTQ2 *** | x_contig_1567298 | 1597 | 17% | 34% | 69% |
| HvATG2_2 | - | MLOC_4729 | M0WB65 *** | x_contig_135441 | 1553 | 17% | 32% | 66% |
| HvATG3 | FLbaf179k14 | MLOC_66486 | AK252967 | x_contig_51626 | 316 | 26% | 71% | 89% |
| HvATG4 | - | MLOC_62924 | M0XTP6 *** | x_contig_47099 | 350 | 30% | 54% | 77% |
| HvATG5 | NIASHv2006J04 | MLOC_70253 | AK362511 | x_contig_57680 | 371 | 13% | 48% | 72% |
| HvATG6 | NIASHv2011D22 | MLOC_71271 | AK362923, AM075824 | x_contig_59463 | 504 | 19% | 37% | 85% |
| HvATG7 | NIASHv2065A17 | MLOC_20377 | AK367931 | x_contig_158944 | 695 | 28% | 34% | 49% |
| HvATG8_1 | FLbaf129h07 | MLOC_74964 | AK251678 | x_contig_66871 | 122 | 71% | 86% | 87% |
| HvATG8_2 | FLbaf5e12 | MLOC_18032 | AK248733 | x_contig_1578994 | 119 | 73% | 81% | 93% |
| HvATG8_3 | FLbaf77m18 | MLOC_62061 | AK250515 | x_contig_46162 | 116 | 48% | 47% | 50% |
| HvATG9 | NIASHv2013F07 | MLOC_54359 | AM085509, AK363183 | x_contig_39071 | 890 | 13% | 48% | 79% |
| HvATG10 | - | MLOC_73839 | M0YWL5 *** | X_contig_64537 | 142 | 10% | 24% | 50% |
| HvATG11 | - | MLOC_53194 | AK365876 | x_contig_38134 | 986 | 18% | 44% | 71% |
| HvATG12 | - | MLOC_55583 | - | x_contig_40085 | 92 | 35% | 74% | 73% |
| HvATG13 | NIASHv2035H08 | MLOC_12860 | AK365609 | x_contig_1564279 | 540 | 10% | 24% | 76% |
| HvATG16 | NIASHv1141N15 | MLOC_66915 | AK361491 | x_contig_52278 | 516 | 6% | 50% | 77% |
| HvATG18_1 | NIASHv2028H07 | MLOC_56544 | AK364793 | x_contig_40934 | 483 | 16% | 58% | 77% |
| HvATG18_2 | NIASHv2141H12 | MLOC_74982 | AK371787 | x_contig_6690 | 232 | 16% | 17% | 17% |
| HvATG18_3 | NIASHv2025P14 | MLOC_56913 | AK364502 | x_contig_41239 | 385 | 19% | 42% | 38% |
| HvATG18_4 | NIASHv2073H03 | MLOC_4865 | AK368421 | x_contig_135595 | 912 | 7% | 6% | 6% |
| HvATG18_5 | NIASHv2002C04, NIASHv2139D21 | MLOC_24797 | AK362065, AK371649 | x_contig_1655679 | 1019 | 6% | 6% | 5% |
| M. musculus | A. thaliana * | O. sativa ** | ||||||
| HvATG101 | - | MLOC_40301 | M0W150 *** | x_contig_2558024 | 215 | 24% | 58% | 81% |
| HvNBR1 | - | MLOC_74717 | M0YZ84 *** | X_contig_66412 | 859 | 23% | 40% | 71% |
| HvATI1 | - | MLOC_80474 | M0ZAS9 *** | X_contig_9070 | 212 | - | 32% | 58% |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avila-Ospina, L.; Marmagne, A.; Soulay, F.; Masclaux-Daubresse, C. Identification of Barley (Hordeum vulgare L.) Autophagy Genes and Their Expression Levels during Leaf Senescence, Chronic Nitrogen Limitation and in Response to Dark Exposure. Agronomy 2016, 6, 15. https://doi.org/10.3390/agronomy6010015
Avila-Ospina L, Marmagne A, Soulay F, Masclaux-Daubresse C. Identification of Barley (Hordeum vulgare L.) Autophagy Genes and Their Expression Levels during Leaf Senescence, Chronic Nitrogen Limitation and in Response to Dark Exposure. Agronomy. 2016; 6(1):15. https://doi.org/10.3390/agronomy6010015
Chicago/Turabian StyleAvila-Ospina, Liliana, Anne Marmagne, Fabienne Soulay, and Céline Masclaux-Daubresse. 2016. "Identification of Barley (Hordeum vulgare L.) Autophagy Genes and Their Expression Levels during Leaf Senescence, Chronic Nitrogen Limitation and in Response to Dark Exposure" Agronomy 6, no. 1: 15. https://doi.org/10.3390/agronomy6010015
APA StyleAvila-Ospina, L., Marmagne, A., Soulay, F., & Masclaux-Daubresse, C. (2016). Identification of Barley (Hordeum vulgare L.) Autophagy Genes and Their Expression Levels during Leaf Senescence, Chronic Nitrogen Limitation and in Response to Dark Exposure. Agronomy, 6(1), 15. https://doi.org/10.3390/agronomy6010015