Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB)


1
Karaisali Vocational School, Cukurova University, Adana 01770, Turkey
2
Department of Horticulture, Inonu University, Malatya 44280, Turkey
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(7), 127; https://doi.org/10.3390/agronomy8070127
Submission received: 8 May 2018
/
Revised: 6 July 2018
/
Accepted: 17 July 2018
/
Published: 23 July 2018
(This article belongs to the Special Issue Berry Crop Production and Protection)
Abstract
:All living organisms perceive mechanical signals, regardless of their taxonomic classifications or life habits. Because of their immobility, plants are influenced by a variety of environmental stresses, such as mechanical stress, during their growth and development. Plants develop physiological behaviors to adapt to their environment for long-term development and evolution. Sound-induced stress—an abiotic stress factor—is an example of mechanical stress and is caused by sound waves generated by different sources. This stress has a negative effect on the development and growth of plants. The strawberry plants evaluated in this study were exposed to three different sound intensity levels (95, 100, 105 dB) at a constant frequency of 1000 Hz. In strawberry plants, stress induced by sound waves is thought to trigger increased production of secondary metabolites as a defense mechanism. To determine the effect of sound applications, the fresh and dry weights of the roots and shoots were measured in strawberry plants, and the pH, total soluble solids (Brix), titratable acidity, vitamin C, total sugar, total acid, and total phenols were analyzed in the fruits. Results show that the sound stress, which was produced at a constant frequency (1000 Hz) and different sound levels (95, 100, 105 dB), affects the growth parameters of the plant and several quality parameters of the fruit.
Keywords:
abiotic stress; fruit quality parameters; ascorbic acid; biomass; sound waves; frequency; dB1. Introduction
Worldwide, strawberry (Fragaria × ananassa) is commonly consumed either in its fresh form or after it is processed. Strawberry reaches its full size and ripens within 30 days; it is a non-climacteric fruit. This growth period is dependent on light, temperature, soil composition, and some cultivation conditions [1]. In addition to being a fruit that is consumed for its taste, strawberry contains carbohydrates, vitamin C, and some antioxidant compounds (e.g., phenolics and flavonoids) [2]. Secondary plant metabolites are compounds with no fundamental roles in the life processes of plants, but they are important for the plant’s ability to interact with its environment for adaptation and defense [3].
The ability to sense and respond to physical stimuli is of key importance to all living things. Light, temperature, and chemical signals are among the environmental stimuli detected by living organisms. Some of these stimuli are related to physical–mechanical stimuli (i.e., differences in mechanical forces or pressures detected by a living cell). Due to the force of gravity straining self-loading and inner growth, and mechanical loads of snow, ice, fruit, wind, rainfall, touch, sound, and hydration (turgor pressure) may be perceived by a cell. All living organisms perceive mechanical signals, regardless of their taxonomic classifications or life habits (sessile vs. motile). Because of their immobility, plants are influenced by environmental stress, such as mechanical stress, during their growth and development [4,5,6,7]. Plants develop physiological behaviors to adapt to the environment for long-term development and evolution. In previous studies, it was reported that plants are capable of responding to wind, touch, electric fields, magnetic fields, and ultraviolet rays [8,9]. Sato et al. [10] reported that, under mechanical stress, chloroplasts in plant cells were re-localized by an active motor system. Erner and Jaffe [11] reported that the contents of ethylene (C2H4) and abscisic acid (ABA) were increased in plants. They also reported that the gibberellin (GA) and indoleacetic acid (IAA) contents were decreased under mechanical stress. Secondary plant metabolites are compounds with no fundamental roles in the life processes of plants; however, they are vital elements in plants’ interaction with the environment for adaptation and defense mechanisms.
Sound waves and sonication act as forms of abiotic stress on plants [12]. It is well known that plants absorb and resonate some sound frequencies from the external realm [13,14,15]. Sound waves had significant dual effects on the root development of Actinidia chinensis plantlets (p < 0.05). The root activity, total length, and the number of roots were increased by the stimulation from sound waves; however, the cell membrane permeability decreased Increasing ATP content in cells means that anabolism is strengthened. At 1 kHz and 100 dB, the soluble protein content and SOD activity were reported to increase. On the other hand, when sound wave stimulation exceeded 1 kHz and 100 dB, these indices were reduced [16,17,18].
2. Sound Waves and Sound Magnitude
Sound is a mechanical vibration wave that travels in a medium that consists of certain materials. According to physicists, ‘sound’ is the molecular diffusion of an energy source in air medium. Sound consists of vibrations in the air that are sensed by our brains after traveling as waves in a medium and stimulating our ears. Sound waves take the form of sinus waves. The distance between two peaks is the wavelength, and the number of wave peaks measured within 1 s is the frequency (Figure 1a,b). In other words, the frequency of a wave depends on the frequency of the vibrations of the particles in the medium (e.g., air) through which the wave travels. Frequency is computed by measuring the vibrations in time. The number of vibrations in 1 s is expressed in units of Hertz (1 Hertz = 1 cycle/s).
The term ‘sound magnitude level’ refers to the logarithm of an energy-physical magnitude. A decibel measures the perceived sound level; it is the noise level unit. The basic sound magnitude parameter for the sound magnitude level is I0, which is the hearing limit at 1000 Hz:
The magnitude of sound is in proportion to the square of the sound pressure (I ~ ), and the level of the sound pressure is found with the equation:
Here, the basic sound pressure p0 at 1000 Hz is accepted as 2 × 10−5 Pa at the hearing limit. Sound measurement devices directly show the sound pressure level in decibel units using these equations. The sound power level, on the other hand, is the measurement of the sound power diffusing in any direction from the source and is expressed in a logarithmic manner, as in the case of sound magnitude level.
Using a reference power of 1 picowatt/m2, i.e., 10−12 W/m2, the SPL (sound power level) is calculated as:
WSource = The total power diffused by the source
WReference = 10−12 W/m2 [19].
Strawberry is a fruit that is consumed worldwide and is cultivated in open and greenhouse systems. It was determined in previous studies conducted on a variety of fruits and vegetables that stress conditions cause increased production of secondary metabolites as a defense mechanism. However, since a plant’s development is negatively affected when it is exposed to abiotic and biotic stresses, these stresses cause losses in yield. To date, no studies have investigated the effect of sound waves on the quality parameters of the strawberry fruit. Under stress conditions, plants increase production of compounds such as phenolic compounds and ascorbic acid to protect themselves. Therefore, the concentrations of nutritional compounds (which accumulate because of sound stress) are expected to increase under stress. In this study, we aimed to increase the quality parameters in the strawberry via sound waves at 1000 Hz and three different frequencies (95, 100, and 105 dB) without harming the plant. The effects of sound stress on the strawberry fruit’s total soluble solids, titratable acidity, total sugars, total acids, pH total phenolic, and ascorbic acid were measured.
3. Materials and Methods
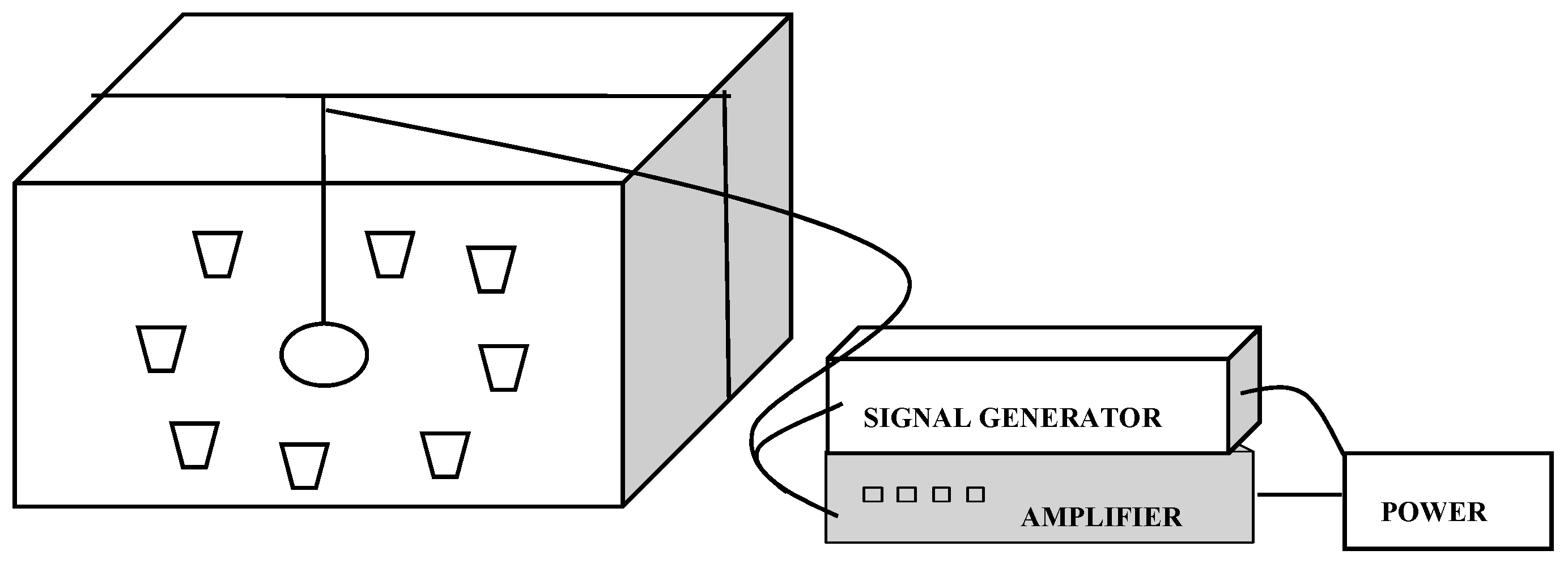
In the present study, a sound amplifier that was capable of transmitting sounds at different decibel values, along with a decibel indicator, was used. A signal generator that was capable of being adjusted was used as a frequency oscillator for creating the 1000 Hz frequency. Three 2 × 2 m chambers that were prepared specifically for the experiment and whose four sides could be opened were used. Furthermore, a sound level meter (noise measurement device) was used as a sound measurement device. Also, speakers that could produce 360° sound were used. The glass was 4 mm thick in the chambers. Between the glass was a 10.5 mm space.
Strawberry plants (Fragaria × ananassa Duch. cv Festival) in 4 L pots of turf + perlite (1:1) were used as the plant material. Hoagland nutrient solution ((M): Ca(NO3)2·4H2O, 3.0 × 10−3; K2SO4, 0.90 × 10−3; MgSO4·7H2O, 1.0 × 10−3; KH2PO4, 0.2 × 10−3; H3BO3, 1.0 × 10−5; 10−4 M FeEDTA, MnSO4·H2O, 1.0 × 10−6; CuSO4·5H2O, 1.0 × 10−7; (NH)6Mo7O24·4H2O, 1.0 × 10−8; ZnSO4·7H2O, 1 × 10−6) was used to water the plants [20]. The plants were placed in the sound chambers for the purpose of measuring the effect of sound stress.
A total of four special sound chambers were used to conduct the study. The chambers were placed in a plastic greenhouse at Research Fields of Cukurova University, Karaisalı Vocational High School (36°59′ N, 35°18′ E, 20 m above sea level), Adana, Turkey. A randomized complete block experimental design was applied in the study (3 replicates, 10 plants in each replicate). Then, different sound waves were directed at the pots. The speaker was 65 cm from each pot (Figure 2). Sound magnitudes of 95, 100, and 105 dB were directed at the pots in the sound chambers. For 30 days, the sound waves were delivered once per day for 1 h in the morning, between 10:00 and 11:00 a.m. The plants were in closed chambers when the sound waves were emitted.
It has been stated by specialists that being exposed to sound exceeding 85 dB might be dangerous. For this reason, dB values above 85 dB were selected. To compare results, control plants were placed in a chamber in which no sound applications were made.
The experiment was designed as a randomized complete block experimental design with 3 replicates, 10 plants in each replicate. A total of four chambers were used. The chambers could only detect the sound within, as the chambers were built in such a way that no other sound could enter. IBM SPSS Statistics 20 software was used for data analysis. The mean values of the fruit parameters for the three sound frequencies were compared using an ANOVA test. The effects of sound on the fruit parameters were considered significant at p ≤ 0.05.
3.1. Measurements and Analyses in Strawberry Plants and Fruits
The trial was started on 16 April 2014 and finalized on 16 May 2014. The fresh and dry weights of the roots in strawberry plants and the fresh and dry weights of the green parts were measured every 10 days for a total of three measurement points throughout the trial. The pH, total soluble solids (Brix), titratable acidity, vitamin C, total sugar, total acid, and total phenol contents of the fruits were analyzed. For the purpose of preparing ultrapure water (18.2 MΩ cm), the Millipore System (Millipore Corp., Bedford, MA, USA) was used. The chromatography reagent standards and solvents were obtained from Sigma Chemical, Co. (St. Louis, MO, USA).
3.2. Determination of Total Soluble Solids (TSS) and Titratable Acidity (TTA)
A hand-type refractometer (ATAGO ATC-1, Tokyo, Japan) was used to determine the total soluble solids in the juice of each sample. For the purpose of determining the total titratable acidity levels, the acid–base titration method was applied. The juice (1 mL) and distilled water (50 mL) were added to a conical bottle to titrate with aqueous NaOH (0.1 N) to obtain pH 8.1. Total acid content was determined in citric acid equivalents and is reported as the mean value of triplicate analyses.
3.3. Extraction of Sugars and Acids (TS and TA)
One gram of the sample was weighed and powdered with liquid nitrogen. The sample was added to 20 mL of aqueous ethanol (80%, v/v) and the solution placed in a screw-cap Eppendorf tube and then in an ultrasonic bath where it was sonicated for 15 min at 80 °C. It was then filtered through filter paper (the extraction was repeated three more times). The filtered extracts were mixed and evaporated in a boiling water bath until dry. Distilled water (2 mL) was used to dissolve the precipitation, and the resulting solution was filtered using Whatman nylon syringe filters (0.45 µm pore size, 13 mm diameter) before HPLC analysis. For organic acid extraction, liquid nitrogen was used to powder the homogenate (1 g of frozen sample), which was then weighed. Then, it was mixed with 20 mL aqueous metaphosphoric acid (3%) at room temperature for 30 min with a shaker. The mixture was then filtered and its volume was increased to 25 mL using the same solvent. It was then used for HPLC analysis [21].
3.4. HPLC of Organic Acid and Sugars
There is a built-in degasser, pump, and controller coupled to a photodiode array detector (Shimadzu SPD 10A vp) in the high-performance liquid chromatographic apparatus (Shimadzu LC 10A vp, Kyoto, Japan). The device also has an automatic injector with a 20 µL injection volume and is interfaced with a computer with Class VP Chromatography Manager Software (Shimadzu, Japan). The separation process was performed with a 250 × 4.6 mm i.d., 5 µm, reverse-phase Ultrasphere ODS analytical column (Beckman, Fullerton, CA, USA). The column was run at room temperature with a flow rate of 1 mL min−1. The process was performed with a 0.1 a.u.f.s. sensitivity (wavelengths between 200 and 360 nm). The elution was isocratic with 0.5% aqueous metaphosphoric acid. The retention times of the components were compared using an in-house PDA library to identify the components that had authentic standards under analytical conditions and UV spectra. Between injections, there was a 10 min equilibrium time. The Shimadzu LC-10 A vp device was used for separating the sugar on a 150 × 4.6 mm i.d., 5 µm, reverse-phase Nucleosil NH2 analytical column (Shimadzu, Tokyo, Japan) at room temperature with a 1 mL min−1 flow rate [21].
3.5. Determination of Total Phenolic Content (TPC) and Ascorbic Acid (AA) in Strawberry Fruits
In order to determine the total phenolic content (TPC), the Folin–Ciocalteu method was employed. After homogenization with a T18, IKA Homogenizer, Germany, 5 g of the frozen fruits with 25 mL ethanol was centrifuged at 3500× g for 3 min. Filter paper was used to filter the supernatant. Then, 2 mL of 10% Folin–Ciocalteu reagent was added to 0.4 mL of the extract. After this, it was left idle for 2–3 min. Finally, 1.6 mL (7.5%) of Na2CO3 solution was added to the mixture, which was incubated for 1 h in the dark, after which is was measured at 765 nm in a spectrophotometer (UV-1201, Shimadzu, Kyoto, Japan) against a blank solution (0.4 mL water + 2 mL Folin–Ciocalteu reagent + 1.6 mL Na2CO3). Using the gallic acid standard, the total phenolic content was computed as 1 mg gallic acid equivalent (GAE) 100 g−1. The results are reported as mg/gallic acid equivalents per gram/dry weight. The Merck RQflex reflectometer was employed to analyze the ascorbic acid content (AA) in the samples by adopting the protocol for the juice of red fruit. The results are given as mg ascorbic acid/100 g fresh sample [21].
4. Results and Discussion
4.1. Results of the Weight Measurements in Strawberry Plants in Fresh and Dry Roots and Shoots
Table 1 shows that the difference between the fresh and dry weight values of the roots was statistically significant for all three measurement dates. As the sound level increased, the root growth regressed. For the measurement that was made 10 days after the initial sound application, the lowest root weight was determined to belong to plants subjected to 105 dB sound, followed by those exposed to the 100 dB level. The highest fresh and dry root weight values were found to be in the control plants. The fresh and dry root weights of plants at the 95 dB level were equal to those of the control plants. For the root weight measurements that were made 20 days after the initial sound application, it was determined that the lowest fresh and dry values were in the plants that received sound at the 105 dB level, and the highest values were measured in the control plants. With respect to fresh weight measurements, the sound levels fall into different statistical groups; for dry weight measurements, only the application at 105 dB is significantly different from the others. Similar results were obtained for the measurements that were made 30 days after the initial sound application. For the fresh root weight, the control plants and the plants at the 95 dB level had similar values and fall into the same statistical group. The weight decreased at 100 dB, the control group plants are included in the group with the plants at 95 dB; the treatment at 105 dB, having the lowest root weight values, is in its own group as it was significantly different from all other applications (Table 1).
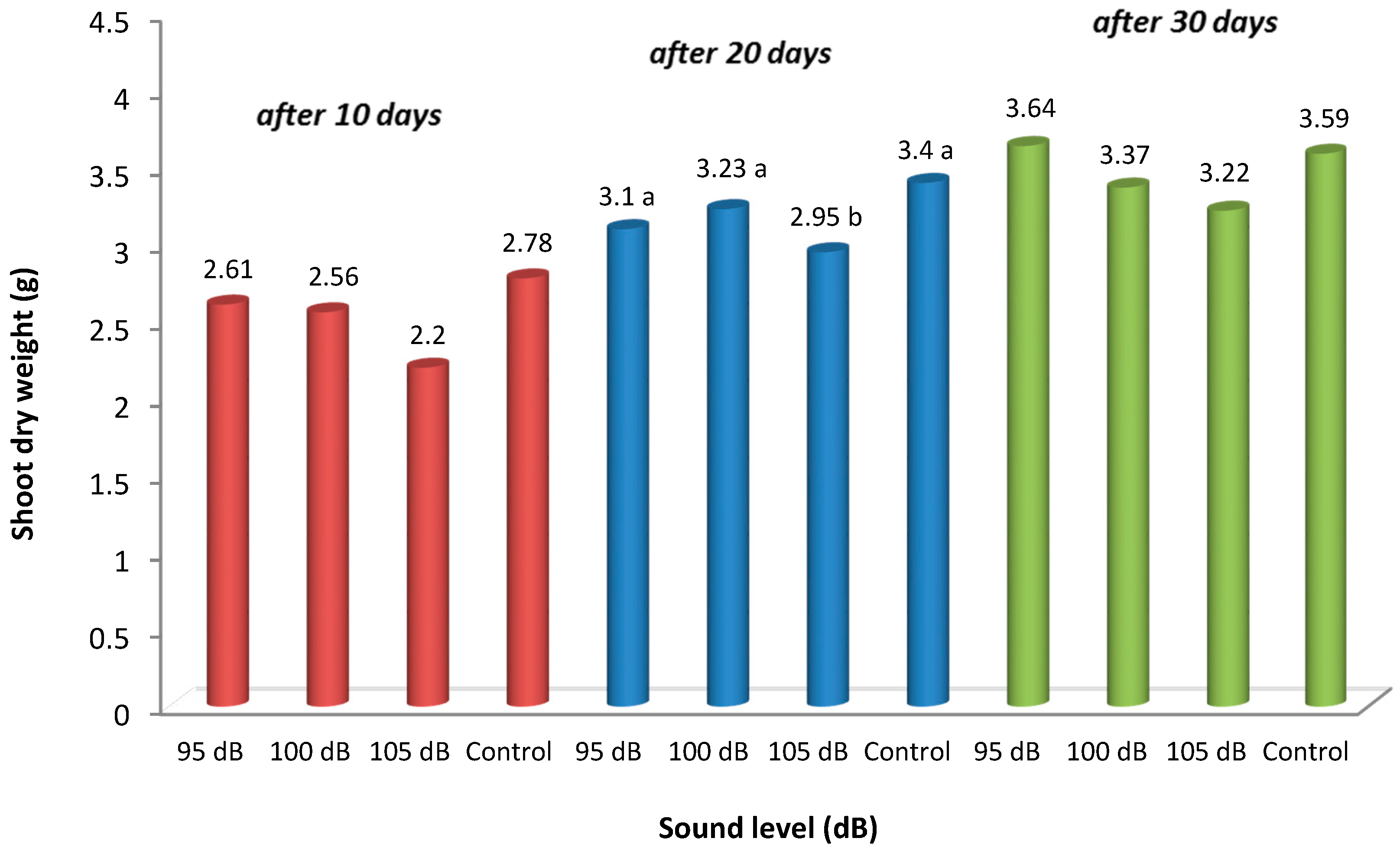
The measurement results in Table 1 for fresh and dry weights of shoots show there was no statistically significant difference in these values between the applications for the measurements made 10 days after the onset of the sound application. However, like the root weights, the sound magnitude negatively affected shoot weight, the control plants had the highest values, and weight decreased at 105 dB. In the second measurement, made 20 days after the initial sound application, the difference between the applications was found to be statistically significant in terms of shoot fresh and dry weight values. For the root fresh weight results, while 100 and 105 dB treatments are in the same statistical group with the lowest values, the control plants, with the highest values, falls into a separate group, showing a significant difference compared to the 100 and 105 dB level applications. The 95 dB sound application is in an intermediary group. For the measurements made 30 days after the initial sound application, the fresh shoot weight results were determined to be significant at a statistical level. The control plants had the highest fresh weight values, and the plants at 95 dB had the second highest. The lowest shoot fresh weights were found in plants at the 100 dB and 105 dB levels; they were significantly different from the other two applications (control and 95 dB). For the last measurement date, the differences between the applications were not found to be significant at a statistical level in terms of shoot dry weight values. However, it was determined that the shoot dry weight values were lower in the plants that were exposed to sounds at high levels (Table 1 and Figure 3).
At 105 dB, as the sound level increased, the rate of the decrease in root dry weights was 44–46% for all measurement dates compared with the control plants. The decrease in root dry weight values at the 100 dB sound level was determined to occur at a rate of 8–26% compared to the control plants. The shoot dry weight values at 105 dB decreased at a rate of 10.3–21% compared to the control plants. The shoot dry weight values at the 100 dB sound level decreased at a rate of 5–8% compared to the control plants (Figure 3). The 95 dB level is included in the same group as the controls for most measurements, although 95 dB did not cause much weight loss in the roots and shoots.
Energy metabolism (for example, sugar, lipid, and photosynthesis) is influenced as abiotic stress increases [22,23,24,25]. For this reason, it is possible that metabolic responses to abiotic stress are gradual and complex. Abiotic stress also influences various cellular processes like growth, photosynthesis, carbon partitioning, carbohydrate–lipid metabolism, osmotic homeostasis, protein synthesis, and gene expression [26,27,28].
4.2. Results of the Analysis of Strawberry Plants
When we consider the results of the analysis made 30 days after the initial sound applications at different levels (in Table 2), we see that pH values were similar for all applications, ranging between 3.36 and 3.39. Similarly, total soluble solids (TSS) and titratable acidity (TTA) values were not significantly affected by the sound applications at different levels. The TTS results were similar, ranging between 8.2 and 8.4. The TTA results were between 7.35 and 7.42. Although there were differences in total acid (TA) for different applications, this difference was not at a statistically significant level. As the sound level increased, the acid rates in the fruits increased. While the total acid was 21.36 in the fruits at the 105 dB sound level, this value was 19.55 in the control plants (Table 2).
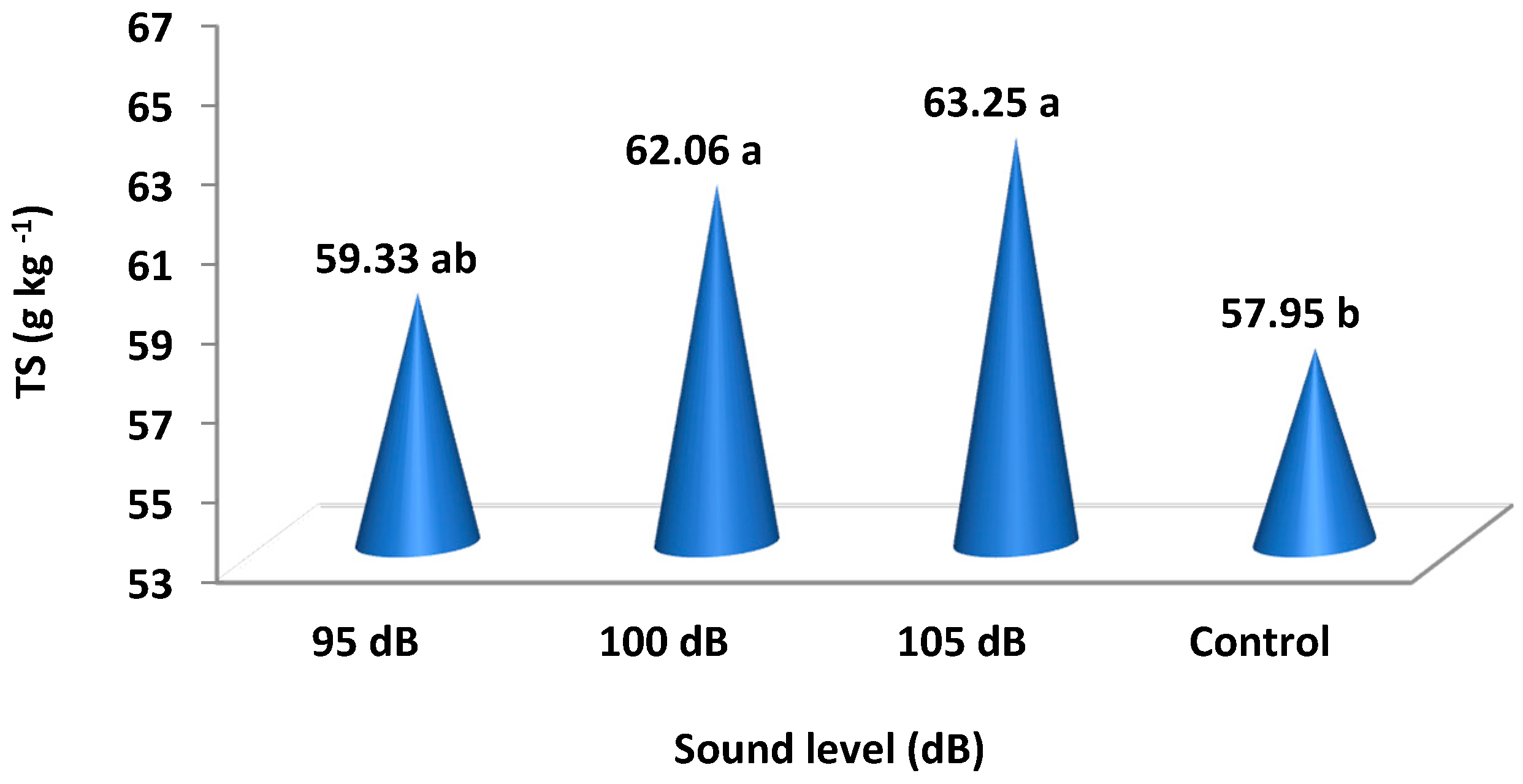
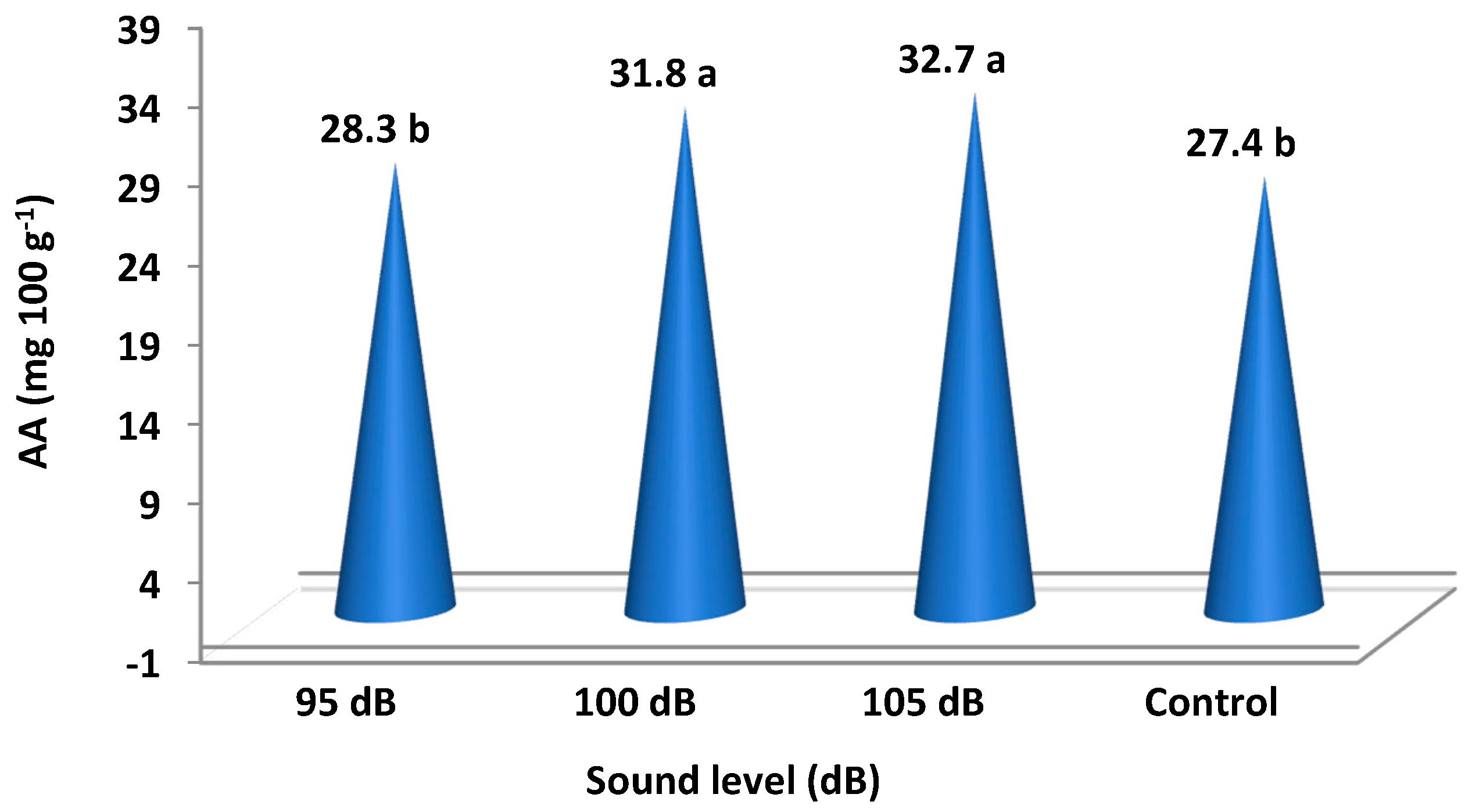
It was determined that different sound levels caused statistically significant differences in total sugar (TS), total phenol content (TPC), and ascorbic acid (AA). Total sugar increased for the applications of 105 and 100 dB sound levels, falling into the same statistical group with values of 62.06 and 63.25. The control group fruits, falling into a separate group, were determined to have 57.95 total sugar. Plants exposed to the 95 dB sound level had total sugar that falls into the intermediary group with a value of 59.33. Total phenol content also increased with the sound magnitude; the phenol contents of the fruits at 105 dB was determined to be 288.0 and is included in a separate group. Total phenol content was 275.5 in the control group plants, which was the lowest value, and it was 279.5 for plants at the 95 dB sound level; these values are in the same statistical group. Total phenol content was 282.0 for the application at 100 dB, which is in the intermediary group. Ascorbic acid (AA) results were similar to the patterns for TPC. While the ascorbic acid of the control plants was 27.4, which is the lowest value, it was 28.3 at 95 dB; the control and 95 dB treatment are in the same group. The ascorbic acid values for the 100 dB and 105 dB sound levels were 31.8 and 32.7, respectively. These are the highest values, and these two applications were found to be in the same group (Table 2).
Total sugar (TS), total phenol content (TPC), and ascorbic acid (AA), which were detected at statistically significant levels in the fruit analyses, increased with the increasing sound magnitudes. Total sugar increased at a rate of 9% in strawberry fruits at 105 dB compared to the control group, and it increased at a rate of 7% at 100 dB (Figure 4). Total phenol content increased at a rate of 4.5% at 105 dB and at a rate of 2.4% at 100 dB. Ascorbic acid increased at a rate of 19% at 105 dB compared to the control group and at a rate of 16% at 100 dB (Table 2 and Figure 5).
The diversity in the structure and function of secondary metabolites makes them necessary because they are of critical importance to the survival of plants under stress conditions [32]. Many environmental stresses (high/low temperature, drought, alkalinity, salinity, UV stress, and pathogen infection) have the potential for damaging plants [3]. In laboratory conditions, production of secondary metabolites was induced anew by using elicitation [3,33]. Several researchers applied various elicitors to improve secondary metabolite production in cultures of plant cells, tissues, and organs [34]. Nutrient stress has an important influence on phenolic levels in plant tissues [3,34]. Pathogen attack, UV irradiation, high-intensity light, wounds, nutrient deficiency, temperature, herbicide treatment, and other environmental stress factors increase the accumulation of phenylpropanoid [35,36,37]. The effects of some secondary plant products on growing conditions are high in terms of the metabolic pathways that are responsible for accumulation of the related natural products.
Sugar accumulation is a common result of abiotic stress (e.g., glucose, fructose, and sucrose accumulate, along with other osmolytes, during cold treatment [38,39,40]). It was traditionally believed that osmolyte accumulation protected plant cells (either by osmotic adjustment or by stabilizing membranes and proteins); however, in time, another role of osmolytes was proposed to be the regulation of redox or sugar signaling. These influences might, for instance, involve hexokinase-dependent signaling or interactions between trehalose synthesis and sugar and ABA signaling [41,42]. It was been reported that ABA synthesis and signaling are important components in sugar signaling. In plants, sugars play important roles as both nutrients and signal molecules. Both glucose and sucrose are recognized as pivotal integrating regulatory molecules that control gene expression related to plant metabolism, stress resistance, growth, and development [43,44,45]. It was recently proposed that soluble sugars, especially when they are present at higher concentrations, might act as reactive oxygen species (ROS) scavengers themselves [46]. All abiotic stresses generate ROS, potentially leading to oxidative damage affecting crop yield and quality. In addition to the well-known classical antioxidant mechanisms, sugars and sugar-metabolizing enzymes have entered the picture as important players in the defense against oxidative stress [47].
Abiotic stress may be used in preharvest activities to improve the quality and yield of products [48]. For instance, vitamin C may be improved in plants that are exposed to high-intensity light or in plants that have less frequent irrigation [49].
The environment affects the ascorbic acid concentration in the fruits and leaves [50,51]. The level of synthesis controls the regulation of ascorbate levels in cells [52,53]. Recycling and degradation (Pallanca and Smirnoff, Green and Fry [53,54]) and transport of this molecule in cells or between organs (Horemans, Foyer, and Asard [55]) are also controlled by the synthesis. During the stress response and the adaptation to stress, the recycling pathway is important. Reduced ascorbate, an antioxidant, is oxidized into an unstable radical (monodehydroascorbate) in oxidative stress conditions; the oxidized molecules then dissociate into ascorbate and dehydroascorbate.
Alessandra Ferrandino and Claudio Lovisolo [56] claimed that abiotic stress modified the growth and development in all plant organs of grapevine plants. At the berry level, the response to abiotic stress drives the accumulation of secondary metabolites in berry pulps, seeds, and skins as a defense against cell damage. Viticultural trials may be designed to control plant stress response to increase secondary metabolite concentrations.
Exposure of plants to unfavorable environmental conditions (e.g., heavy metals, drought, nutrient deficiency, salt stress) can increase the production of reactive oxygen species (ROS). To protect themselves against these toxic oxygen intermediates, plants employ antioxidant defense systems [57]. To control the levels of ROS and to protect cells under stress conditions, plant tissues contain several enzymes that scavenge ROS (SOD, CAT, peroxidases, and glutathione peroxidase), detoxify LP products (glutathione-S-transferases, phospholipid-hydroperoxide glutathione peroxidase, and ascorbate peroxidase), and a network of low molecular mass antioxidants (ascorbate, glutathione, phenolic compounds, and tocopherols) [57]. Secondary metabolites are involved in protective functions in response to both biotic and abiotic stress conditions. As determined in a recent study, environmental factors increase the concentrations of phytochemicals [58]. Stress predominates among all the factors that enhance the concentrations of phytochemicals in fruits and vegetables. This makes sense when it is considered that all stress types (biotic/abiotic) are conducive to oxidative stress in plants [59], and oxidative signaling controls synthesis and accumulation of secondary metabolites [60]. Plants produce phenolic compounds as a defensive mechanism to biotic/abiotic stresses [61].
In the strawberry plant in the fruit development stage, sound frequency stress promoted metabolite accumulation, which resulted in an improvement in fruit quality.
5. Conclusions
In the present study, different sound waves with different decibel values at a constant frequency were applied, and they caused decreases in the weight of roots and the green parts of the strawberry plants. However, these different sound waves also caused some increases in several quality parameters in the fruits at a statistically significant level. This increase, which also enhances the value of the fruit and is important for human health, is positive. Salinity, drought, high temperature, and irreversible abiotic stress factors reduce the growth and development of the plants, causing major losses in terms of yield; plants even die under continuous stress. In further studies, greenhouse trials may be performed at sound levels that do not affect plant development and yield at significant levels but increase fruit quality. In this way, sound applications that do not damage the soil and plant but increase the quality parameters in a positive manner may be recommended for greenhouse cultivation. In the present study, the weight losses were found to be greater in plants at the 105 dB sound level. At 95 dB, on the other hand, the results were close to those of the control group plants which were not exposed to sound. For this reason, 1000 Hz and 100 dB sound levels may be used for the strawberry plant to increase the quality of its fruits. However, in the future, it should be investigated whether it affects the yield in greenhouse designs.
Author Contributions
H.O. and O.A. conceived and designed the experiments; H.O. performed parts of physics and sound system of the experiments; O.A. performed the parts related to plants and O.A. analyzed the data; O.A. contributed reagents/materials/analysis tools; H.O. and O.A. wrote the paper.
Funding
We thank Cukurova University Scientific Research Projects Directorate for their financial support. And also, we thank to Ebru Kafkas and her laboratory team for strawberry fruits analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Klein, B.P.; Perry, A.K. Ascorbic acid and vitamin A activity in selected vegetables from different geographical areas of the United States. J. Food Sci. 1982, 47, 941–945. [Google Scholar] [CrossRef]
- Robards, K.; Prenzler, P.D.; Tucker, G.; Swatsitang, P.; Glover, W. Phenolic compounds and their role in oxidative processes in fruits. Food Chem. 1999, 66, 401–436. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozkurt, H.; Altuntas, O. The Effect of Sound Waves at Different Frequencies upon the Plant Element Nutritional Uptake of Snake Plant (Sansevieria trifasciata) Plants. Indian J. Sci. Technol. 2016, 9, 48–55. [Google Scholar] [CrossRef]
- Ozkurt, H.; Altuntas, O.; Bozdogan, E. The Effects of Sound Waves upon Plant Nutrient Elements Uptake of Sword Fern (Nephrolepis exaltata) Plants. J. Basic Appl. Sci. Res. 2016, 6, 9–15. [Google Scholar]
- Jaffe, M.J.; Leopold, A.C.; Staples, R.C. Thigmo responses inplants and fungi. Am. J. Bot. 2002, 89, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Baluška, F.; Šamaj, J.; Wojtazek, P.; Volkmann, D.; Menzel, D. Cytoskeleton–plasma membrane–cell wall continuum in plants. Emerging links revisited. Plant Physiol. 2003, 133, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Mary, M.; Braam, J. The Arabidopsis TCH4 xyloglucan endotransglyco-sylase. Plant Physiol. 1997, 115, 181–190. [Google Scholar]
- Sistrunk, M.L.; Antosiewicz, D.M.; Purugganan, M.M.; Braam, J. Arabidopsis TCH3 encodes a novel Ca2+ binding protein and shows environmentally induced and tissue-specific regulation. Plant Cell 1994, 6, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kadota, A.; Wada, M. Mechanically induced avoidance response of chloroplasts in fern protonemal cells. Plant Physiol. 1999, 121, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Erner, Y.; Jaffe, M.J. Thigmomorphogenesis: The involvement of auxin and abscisic acid in growth retardation due to mechanical perturbation. Plant Cell Physiol. 1982, 23, 935–941. [Google Scholar]
- Wang, L.; Weller, C.L. Recent advances in extraction of nutraceuticals from plants. Trends Food Sci. Technol. 2006, 17, 300–312. [Google Scholar] [CrossRef]
- Hou, T.Z.; Li, M.D. Experimental evidence of a plant meridian system: IV. The effects of acupuncture on growth and metabolism of Phaseolus vulgaris L. beans. Am. J. Chin. Med. 1997, 25, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Z.; Luan, J.Y.; Wang, J.Y.; Li, M.D. Experimental evidence of a plant meridian system: III. The sound characteristics of Phylodendron (Alocasia) and effects of acupuncture on those properties. Am. J. Chin. Med. 1994, 22, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Z.; Re, Z.W.; Li, M.D. Experimental evidence of a plant meridian system: II. The effects of needle acupuncture on the; temperature changes of soybean (Glycine max). Am. J. Chin. Med. 1994, 22, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.C.; Wang, B.C.; Duan, C.R.; Dai, C.Y.; Jia, Y.; Wang, X.J. Brief study on physiological effects of sound field on actinidia Chinese callus. J. Chongqing Univ. 2002, 25, 79–84. (In Chinese) [Google Scholar]
- Yang, X.C.; Wang, B.C.; Duan, C.R. Effects of sound stimulation on energy metabolism of Actinidia chinensis callus. Colloids Surf. B Biointerfaces 2003, 30, 67–72. [Google Scholar]
- Yang, X.C.; Wang, B.C.; Ye, M. Effects of different sound intensities on root development of Actinidia Chinese plantlet. Chin. J. Appl. Environ. Biol. 2004, 10, 274–276. (In Chinese) [Google Scholar]
- Taş, F. Internal Acoustic Research and Improvement Methods in a Commercial Vehicle. Ph.D. Thesis, Istanbul Technical University, Istanbul, Turkey, 2010. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. In Circular California Agricultural Experiment Station; University of California: Berkeley, CA, USA, 1938; pp. 347–461. [Google Scholar]
- Kafkas, E.; Koşar, M.; Paydaş, S.; Kafkas, S.; Başer, K.H.C. Quality characteristics of strawberry genotypes at different maturation stages. Food Chem. 2007, 100, 1229–1236. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.; Bohlman, M.C.; Quilici, D. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; Harter, K. The AtGenExpress global stress expression data set: Protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, S.B.; Caverzan, A.; Teixeira, F.K.; Lazzarotto, F.; Silveira, J.A.; Ferreira-Silva, S.L.; Abreu-Neto, J.; Margis, R.; Margis-Pinheiro, M. Cytosolic APx knockdown indicates an ambiguous redox responses in rice. Phytochemistry 2010, 71, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Britt, A.B. Repair of DNA damage induced by solar UV. Photosynth. Res. 2004, 81, 105–112. [Google Scholar] [CrossRef]
- Hilal, M.; Parrado, M.F.; Rosa, M.; Gallardo, M.; Orce, L.; Massa, E.M.; Prado, F.E. Epidermal lignin deposition in quinoa cotyledons in response to UV-B radiation. Photochem. Photobiol. 2004, 79, 205–210. [Google Scholar] [CrossRef]
- Ibañez, S.; Rosa, M.; Hilal, M.; González, J.A.; Prado, F.E. Leaves of Citrus aurantifolia exhibit a different sensibility to solar UV-B radiation according to development stage in relation to photosynthetic pigments and UV-B absorbing compounds production. J. Photochem. Photobiol. B Biol. 2008, 90, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Seigler, D.S. Plant Secondary Metabolism; Kluwer Academic Publishers: Boston, MA, USA, 1998. [Google Scholar]
- DiCosmo, F.; Misawa, M. Eliciting secondary metabolism in plant cell cultures. Trends Biotechnol. 1985, 3, 318. [Google Scholar] [CrossRef]
- Sudha, G.; Ravishankar, G.A. Influence of methyl jasmonate and salicylic acid in the enhancement of capsaicin production in cell suspension cultures of Capsicum frutescens Mill. Curr. Sci. 2003, 85, 1212–1217. [Google Scholar]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Fowler, S.; Fiehn, O.; Thomashow, M.F. A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 15243–15248. [Google Scholar] [CrossRef] [PubMed]
- Chalker-Scott, L.; Fnchigami, L.H. The role of phenolic compounds in plant stress responses. In Low Temperature Stress Physiology in Crops; Paul, H.L., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1989; p. 40. [Google Scholar]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingler, A.; Roitsch, T. Metabolic regulation of leaf senescence: Interactions of sugar signalling with biotic and abiotic stress responses. Plant Biol. 2008, 10, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef] [Green Version]
- Avonce, N.; Leyman, B.; Mascorro-Gallardo, J.O.; Van Dijck, P.; Thevelein, J.M.; Iturriaga, G. The Arabidopsis trehalose-6-P synthase AtTPS1 gene is a regulator of glucose, abscisic acid, and stress signalling. Plant Physiol. 2004, 136, 3649–3659. [Google Scholar] [CrossRef] [PubMed]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S.C.M. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramon, M.; Rolland, F.; Sheen, J. Sugar sensing and signaling. The Arabidopsis Book. Am. Soc. Plant Biol. 2008. [Google Scholar] [CrossRef]
- Van den Ende, W.; Valluru, R. Sucrose, sucrosyl oligosaccharides, and oxidative stress: Scavenging and salvaging? J. Exp. Bot. 2009, 60, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Bolouri-Moghaddam, M.R.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalt, W.; Ryan, D.A.J.; Duy, J.C.; Prior, R.L.; Ehlenfeldt, M.K.; Vander Kloet, S.P. Interspecific variation in anthocyanins, phenolics, and antioxidant capacity among genotypes of highbush and lowbush blueberries (Vaccinium section cyanococcus spp.). J. Agric. Food Chem. 2001, 49, 4761–4767. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Dumas, Y.; Dadomo, M.; Di Lucca, G.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidantcontent of tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Yu, J.; Gomez, F.; Fernández, L.; McIntosh, L.; Foyer, C.H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot. 2006, 57, 1621–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N.; Conklin, P.L.; Loewus, F.A. Biosynthesis of ascorbic acid in plants: A renaissance. Annu. Rev. Plant Biol. 2001, 52, 437–467. [Google Scholar] [CrossRef] [PubMed]
- Pallanca, J.E.; Smirnoff, N. The control of ascorbic acid synthesis and turnover in pea seedlings. J. Exp. Bot. 2000, 51, 669–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.A.; Fry, S.C. Vitamin C degradation in plant cells via enzymatic hydrolysis of 4-O-oxalyl-l-threonate. Nature 2005, 433, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Horemans, N.; Foyer, C.H.; Asard, H. Transport and action of ascorbate at the plant plasma membrane. Trends Plant Sci. 2000, 5, 263–267. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Poiroux-Gonord, F.; Bidel, L.P.; Fanciullino, A.L.; Gautier, H.; Lauri-Lopez, F.; Urban, L. Health benefits of vitamins and secondary metabolites of fruits and vegetables and prospects to increase their concentrations by agronomic approaches. J. Agric. Food Chem. 2010, 58, 12065–12082. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, J.; Hippeli, S.; Elstner, E.F. Plant’s defence and its benefits for animals and medicine: Role of phenolics and terpenoids in avoiding oxygen stress. Plant Physiol. Biochem. 2002, 40, 471–478. [Google Scholar] [CrossRef]
- Kunz, D.A.; Chen, J.L.; Pan, G. Accumulation of α-keto acids as essential components in cyanide assimilation by Pseudomonas fluorescens NCIMB 11764. Appl. Environ. Microbiol. 1998, 64, 4452–4459. [Google Scholar] [PubMed]
- English-Loeb, G.; Stout, M.J.; Duffey, S.S. Drought stress in tomatoes: Changes in plant chemistry and potential nonlinear consequences for insect herbivores. Oikos 1997, 79, 456–468. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Low-frequency sinus wave; (b) high-frequency sinus wave.

Figure 2.
Illustration of the trial design.

Figure 3.
Shoot dry weight changes. Measurements were made at 10-day intervals in plants that were exposed to different sound levels. The means of different letters are significantly different (p < 0.05).
Figure 3.
Shoot dry weight changes. Measurements were made at 10-day intervals in plants that were exposed to different sound levels. The means of different letters are significantly different (p < 0.05).

Figure 4.
Total sugars (TS) in strawberry fruits 30 days after exposure to different sound waves. The means of different letters are significantly different (p < 0.05).
Figure 4.
Total sugars (TS) in strawberry fruits 30 days after exposure to different sound waves. The means of different letters are significantly different (p < 0.05).

Figure 5.
Total ascorbic acid (AA) in strawberry fruits 30 days after exposure to different sound waves. The means of different letters are significantly different (p < 0.05).
Figure 5.
Total ascorbic acid (AA) in strawberry fruits 30 days after exposure to different sound waves. The means of different letters are significantly different (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fresh and dry weight changes in the roots and shoots. Measurements were made at 10-day intervals in plants that were exposed to different sound levels.
Table 1.
Fresh and dry weight changes in the roots and shoots. Measurements were made at 10-day intervals in plants that were exposed to different sound levels.
| 10 Days after the Sound Treatment | 20 Days after the Sound Treatment | 30 Days after the Sound Treatment | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sound Intensity Treatments | RFW (g) | RDW (g) | SFW (g) | SDW (g) | RFW (g) | RDW (g) | SFW (g) | SDW (g) | RFW (g) | RDW (g) | SFW (g) | SDW (g) |
| 95 dB | 7.94 a | 1.10 a | 10.92 | 2.61 | 9.50 ab | 1.38 a | 17.84 ab | 3.10 a | 12.50 a | 1.72 a | 21.25 a | 3.64 |
| 100 dB | 5.77 b | 0.85 b | 10.33 | 2.56 | 8.98 b | 1.32 a | 16.32 b | 3.23 a | 9.32 b | 1.40 ab | 19.18 b | 3.37 |
| 105 dB | 4.74 b | 0.64 b | 9.13 | 2.20 | 6.30 c | 0.78 b | 15.05 b | 2.95 b | 8.86 c | 0.96 b | 18.52 b | 3.22 |
| Control | 7.34 a | 1.15 a | 11.15 | 2.78 | 10.57 a | 1.43 a | 19.52 a | 3.40 a | 13.03 a | 1.77 a | 22.92 a | 3.59 |
The means in the columns followed by different letters are significantly different (p < 0.05); RFW: root fresh weight; RDW: root dry weight; SFW: shoot fresh weight; SDW: shoot dry weight.
Table 2.
Measurements of several quality parameters of the fruits taken from plants 30 days after exposure to different sound levels
Table 2.
Measurements of several quality parameters of the fruits taken from plants 30 days after exposure to different sound levels
| Sound Intensity Treatments | pH | TSS (%) | TTA (g kg−1) | TS (g kg−1) | TA (g kg−1) | TPC (mg 100 g−1 Gallic Acid) | AA (mg 100 g−1) |
|---|---|---|---|---|---|---|---|
| 95 dB | 3.36 | 8.4 | 7.37 | 59.33 ab | 19.82 | 279.5 b | 28.3 b |
| 100 dB | 3.38 | 8.3 | 7.41 | 62.06 a | 20.62 | 282.0 ab | 31.8 a |
| 105 dB | 3.39 | 8.4 | 7.42 | 63.25 a | 21.36 | 288.0 a | 32.7 a |
| Control | 3.36 | 8.2 | 7.35 | 57.95 b | 19.55 | 275.5 b | 27.4 b |
The means in the columns followed by different letters are significantly different (p < 0.05); TSS: total soluble solids; TS: total sugars; TTA: titratable acidity; TA: total acids; TPC: total phenolic content; AA: ascorbic acid.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ozkurt, H.; Altuntas, O. Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB). Agronomy 2018, 8, 127. https://doi.org/10.3390/agronomy8070127
AMA Style
Ozkurt H, Altuntas O. Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB). Agronomy. 2018; 8(7):127. https://doi.org/10.3390/agronomy8070127
Chicago/Turabian StyleOzkurt, Halil, and Ozlem Altuntas. 2018. "Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB)" Agronomy 8, no. 7: 127. https://doi.org/10.3390/agronomy8070127
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.