Compared to Australian Cultivars, European Summer Wheat (Triticum aestivum) Overreacts When Moderate Heat Stress Is Applied at the Pollen Development Stage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions and Heat Stress Treatment
2.2. Physiological Measurements
2.3. Histological Analysis
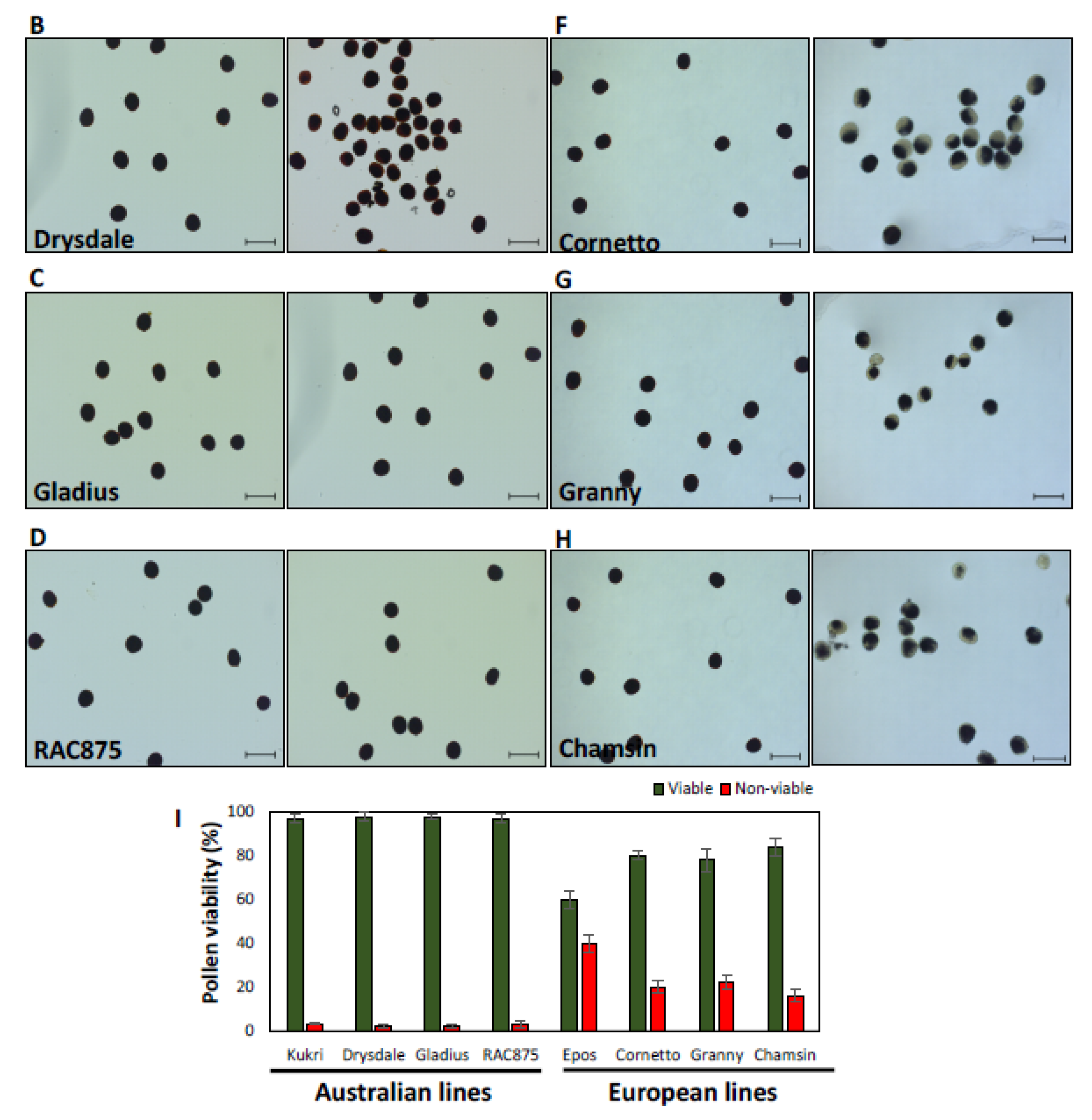
2.4. Microscopy and Pollen Viability Assay
2.5. RNA Isolation and RT-qPCR
2.6. Statistical Analysis
3. Results
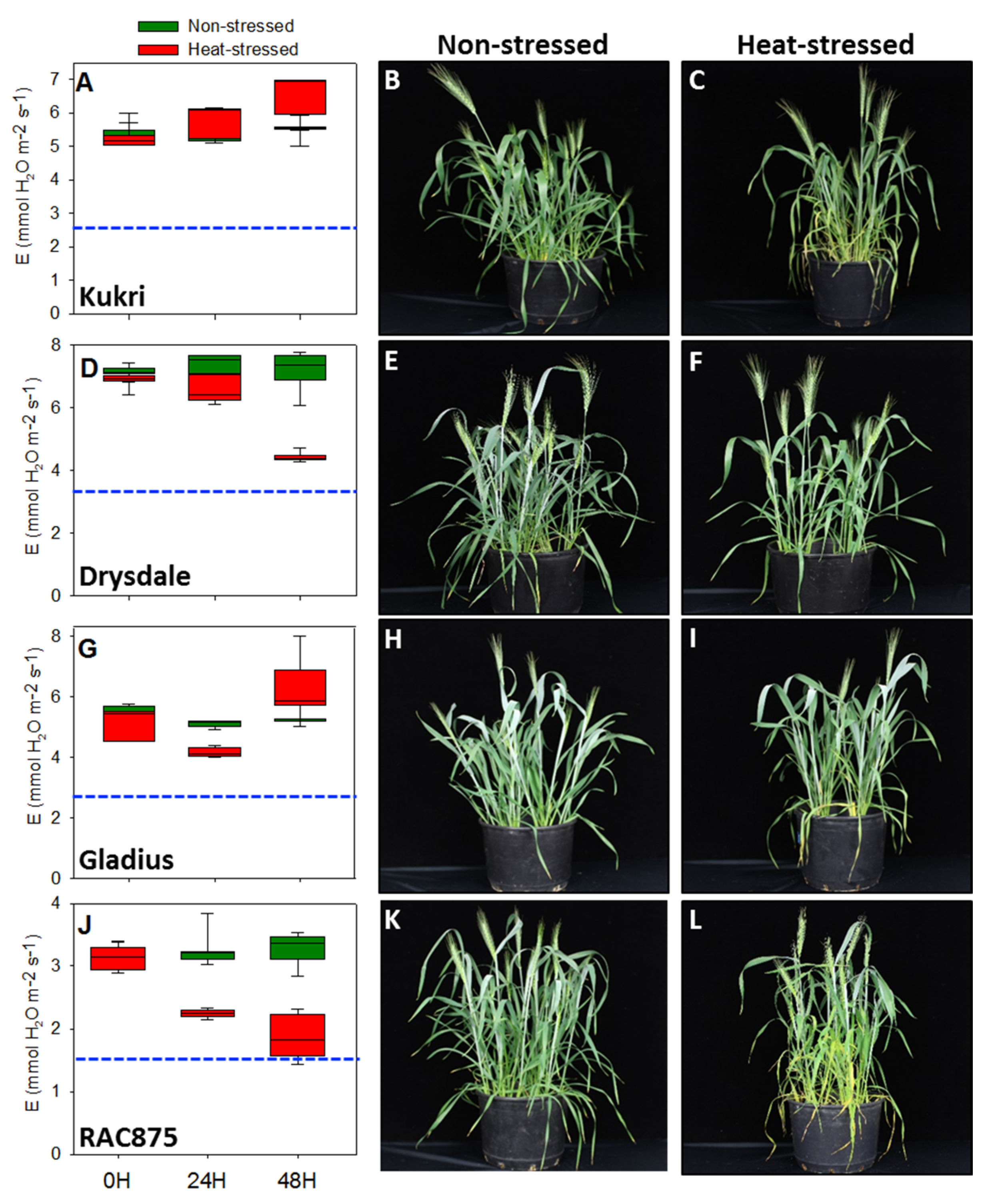
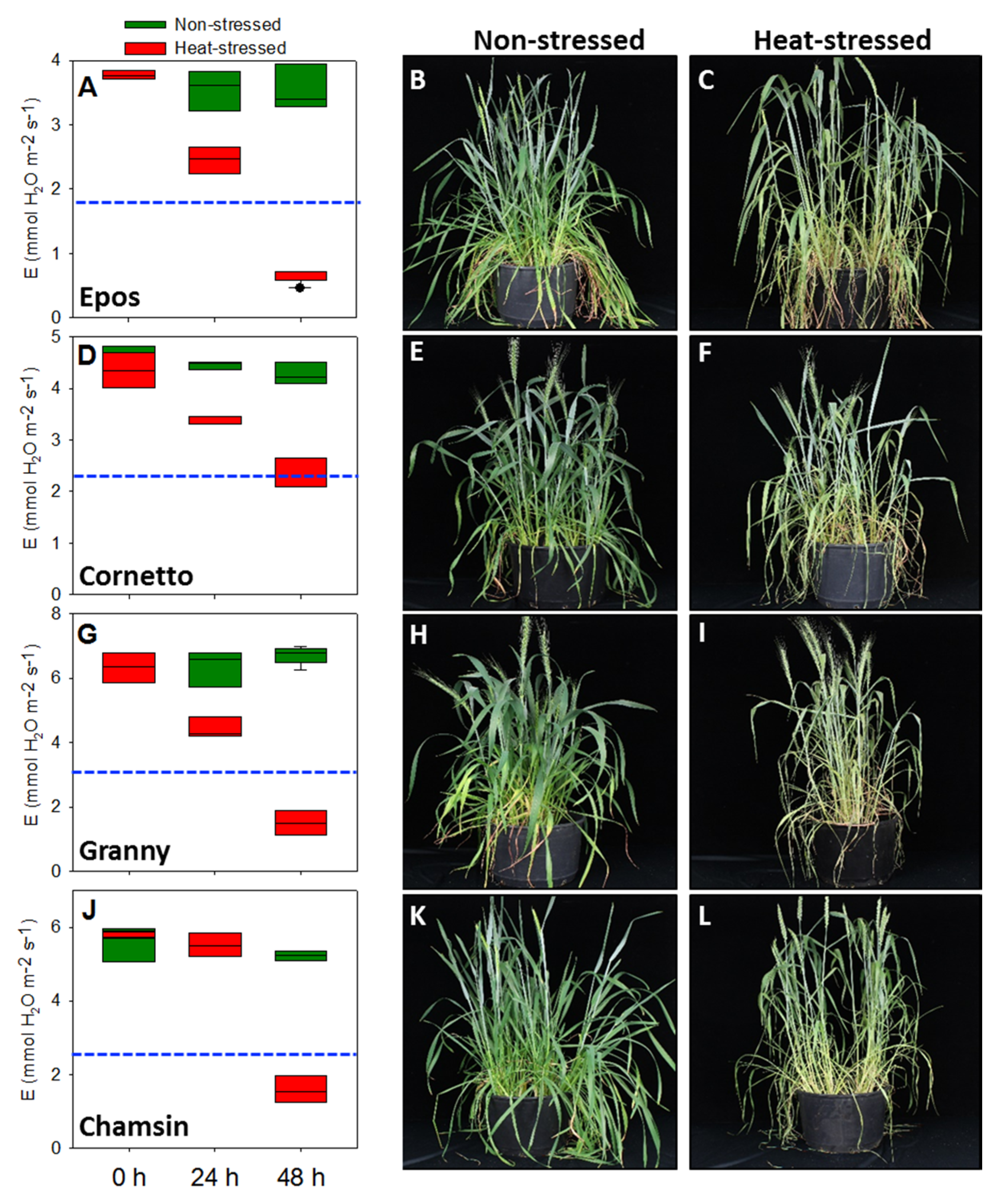
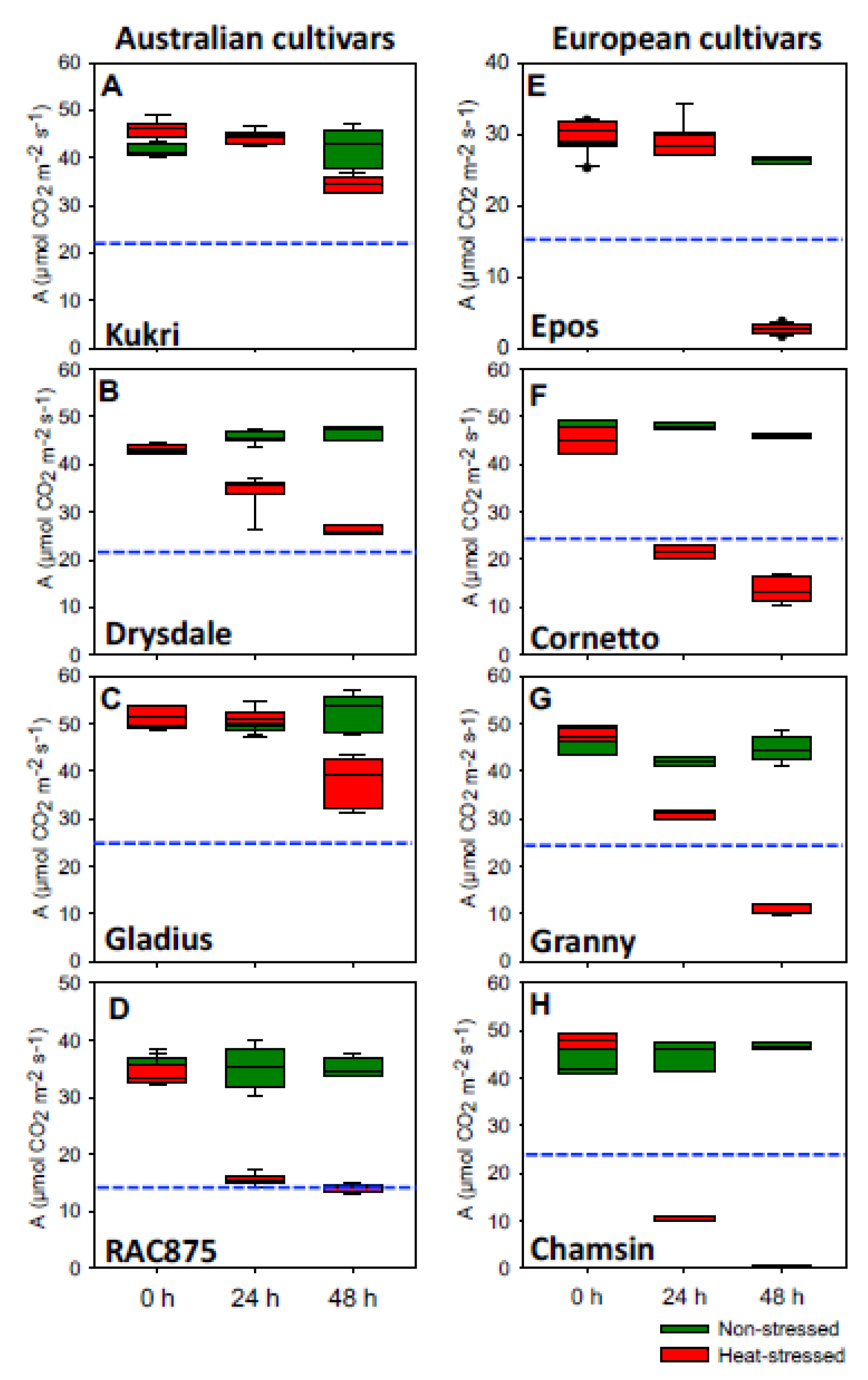
3.1. Physiological Responses of Australian and European Summer Wheat Cultivars under Heat Stress
3.1.1. Transpiration Rate Was Strongly Reduced in European Cultivars
3.1.2. Photosynthesis Rate Was Strongly Reduced in European Cultivars
3.2. Analysis of Male Reproductive Structures after a Short-Term Moderate Heat Stress
3.2.1. Anther Morphology Was Not Significantly Modified after Moderate Heat Stress
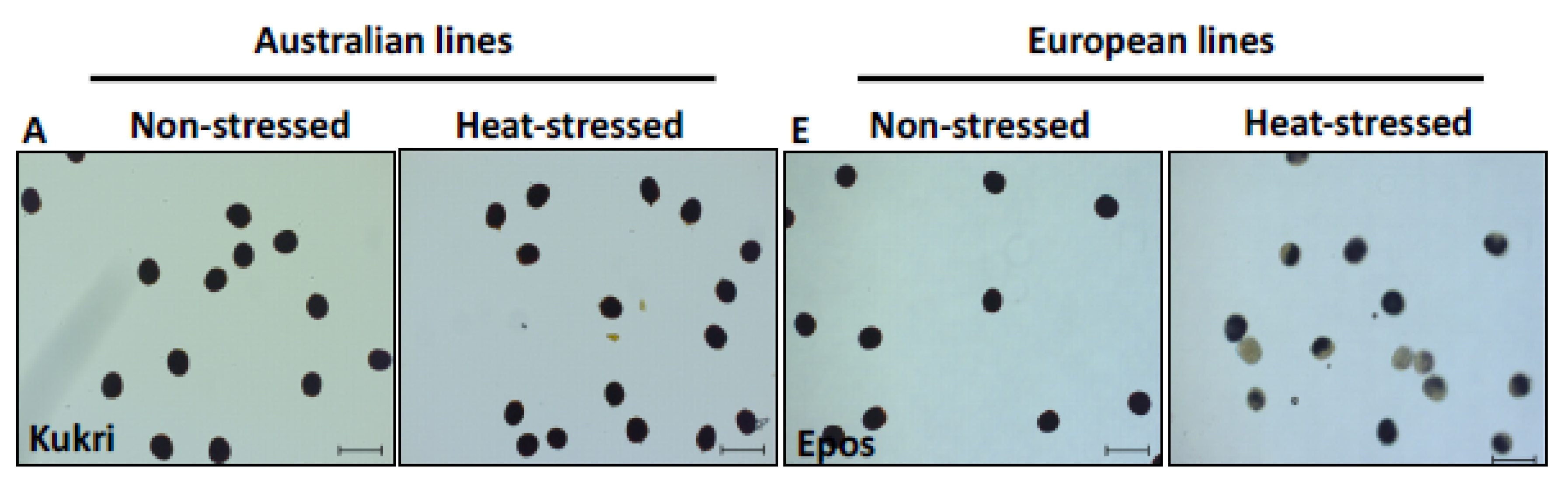
3.2.2. Pollen Viability Was Only Affected in European Cultivars after Moderate Heat Stress
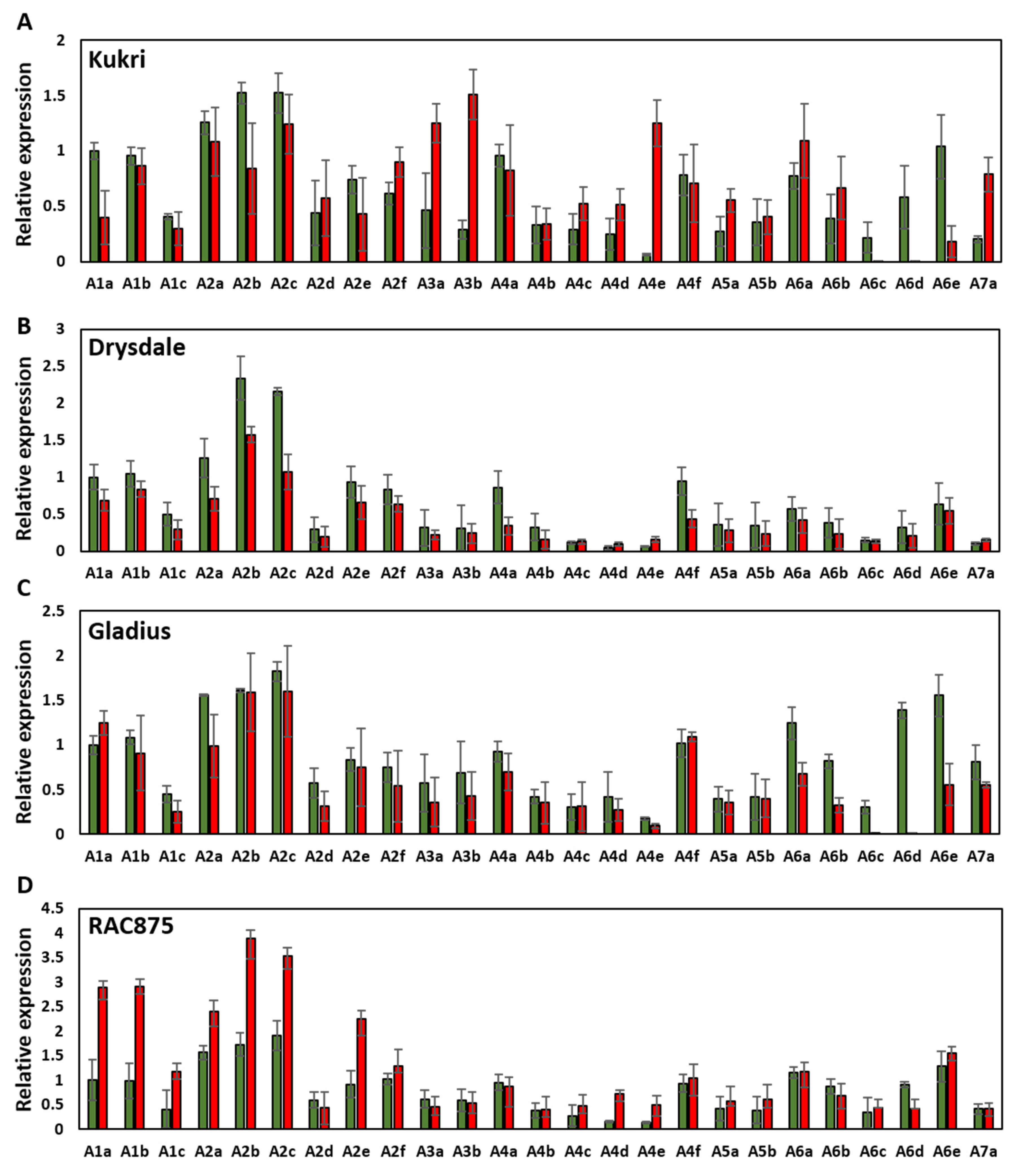
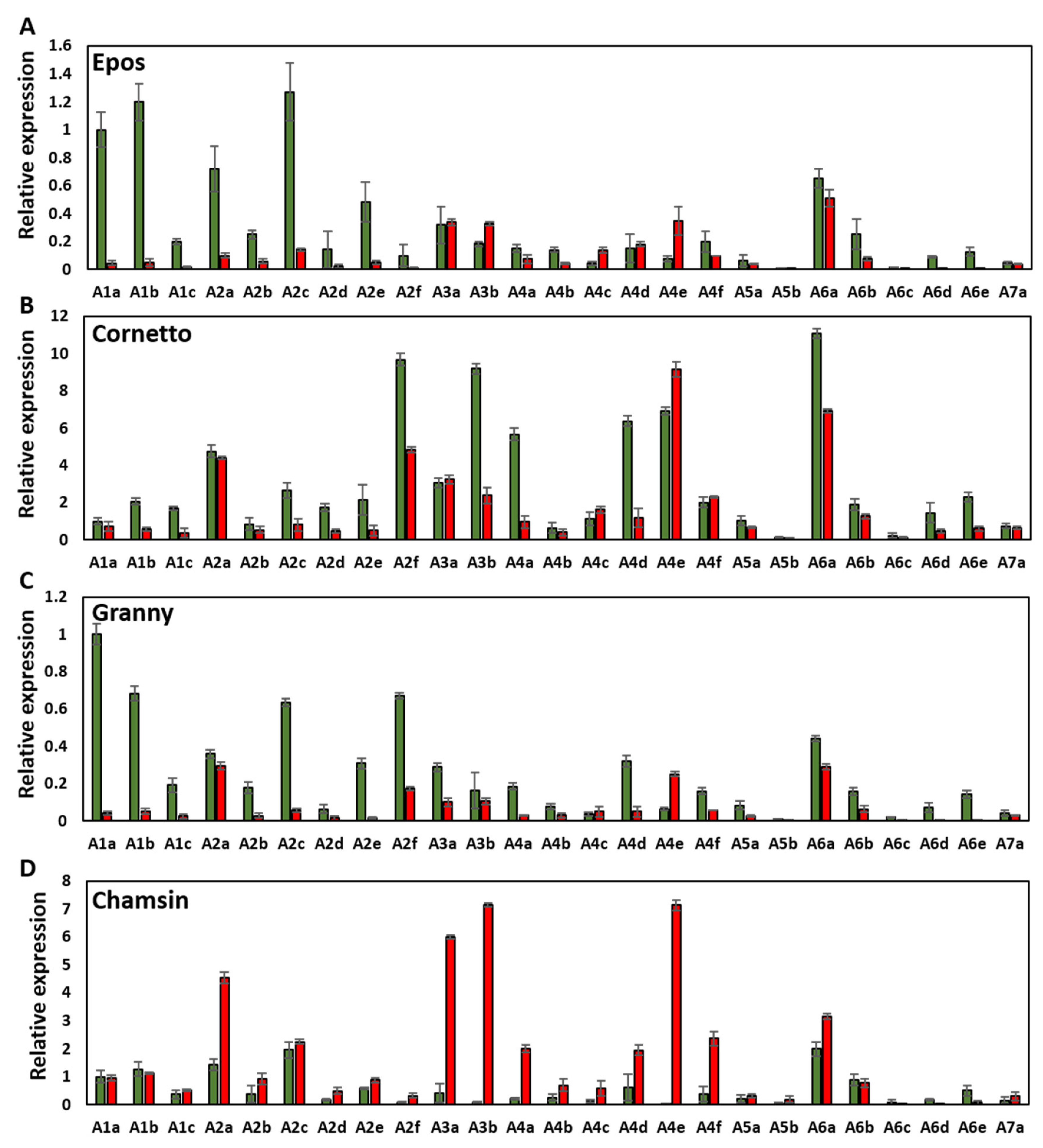
3.3. Heat Shock Factors Genes Were Strongly Regulated in European, but Not in Australian Cultivars
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergauwen, D.; De Smet, I. From early farmers to Norman Borlaug—The making of modern wheat. Curr. Biol. 2017, 27, R858–R862. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.Y.; Chenu, K.; Dreccer, M.F.; Chapman, S.C. Breeding for the future: What are the potential impacts of future frost and heat events on sowing and flowering time requirements for Australian bread wheat (Triticum aestivium) varieties? Glob. Chang. Biol. 2012, 18, 2899–2914. [Google Scholar] [CrossRef] [PubMed]
- USDA Foreign Agriculture Service. Wheat: World Markets and Trade; Office of Global Analysis-USDA: Washington, DC, USA, 2018.
- Agricultural Production—Crops; Eurostat: Luxembourg, 2017.
- Joukhadar, R.; Daetwyler, H.D.; Bansal, U.K.; Gendall, A.R.; Hayden, M.J. Genetic diversity, population structure and ancestral origin of Australian wheat. Front. Plant Sci. 2017, 8, 2115. [Google Scholar] [CrossRef] [PubMed]
- Wieland, T. Scientific theory and agricultural practice: Plant breeding in Germany from the late 19th to the early 20th century. J. Hist. Biol. 2006, 39, 309–343. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Kobza, J.; Edwards, G.E. Influences of leaf temperature on photosynthetic carbon metabolism in wheat. Plant Physiol. 1987, 83, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Weis, E. Reversible heat-inactivation of the Calvin Cycle—A possible mechanism of the temperature regulation of photosynthesis. Planta 1981, 51, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Camejo, D.; Rodriguez, P.; Morales, A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcon, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Yu, X.Q.; Kjaer, K.H.; Rosenqvist, E.; Ottosen, C.O.; Wu, Z. Screening and validation of tomato genotypes under heat stress using F-v/F-m to reveal the physiological mechanism of heat tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Sailaja, B.; Subrahmanyam, D.; Neelamraju, S.; Vishnukiran, T.; Rao, Y.V.; Vijayalakshmi, P.; Voleti, S.R.; Bhadana, V.P.; Mangrauthia, S.K. Integrated physiological, biochemical, and molecular analysis identifies important traits and mechanisms associated with differential response of rice genotypes to elevated temperature. Front. Plant Sci. 2015, 6, 1044. [Google Scholar] [CrossRef] [PubMed]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Tripp, J.; Winkelhaus, S.; Tschiersch, B.; Theres, K.; Nover, L.; Scharf, K.D. In the complex family of heat stress transcription factors, HSfA1 has a unique role as master regulator of thermotolerance in tomato. Genes Dev. 2002, 16, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.P.; Sadat, S.; Drenth, J.; McIntyre, C.L. The heat shock factor family from Triticum aestivum in response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes. J. Exp. Bot. 2014, 65, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Begcy, K.; Dresselhaus, T. Epigenetic responses to abiotic stresses during reproductive development in cereals. Plant Reprod. 2018, 31. [Google Scholar] [CrossRef]
- Chen, C.; Begcy, K.; Liu, K.; Folsom, J.J.; Wang, Z.; Zhang, C.; Walia, H. Heat stress yields a unique MADS box transcription factor in determining seed size and thermal sensitivity. Plant Physiol. 2016, 171, 606–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folsom, J.J.; Begcy, K.; Hao, X.; Wang, D.; Walia, H. Rice Fertilization-Independent Endosperm1 regulates seed size under heat stress by controlling early endosperm development. Plant Physiol. 2014, 165, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Monneveux, P.; Pastenes, C.; Reynolds, M.P. Limitations to photosynthesis under light and heat stress in three high-yielding wheat genotypes. J. Plant Physiol. 2003, 160, 657–666. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Geelen, D. The impact of environmental stress on male reproductive development in plants: Biological processes and molecular mechanisms. Plant Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, J.F.; Talle, B.; Wilson, Z.A. Anther and pollen development: A conserved developmental pathway. J. Integr. Plant Biol. 2015, 57, 876–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, Z.A.; Zhang, D.B. From Arabidopsis to rice: Pathways in pollen development. J. Exp. Bot. 2009, 60, 1479–1492. [Google Scholar] [CrossRef] [PubMed]
- Begcy, K.; Dresselhaus, T. Tracking maize pollen development by the Leaf Collar Method. Plant Reprod. 2017, 30, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, B.; Singh, S.D.; Nagarajan, S.; Aggarwal, P.K. Impact of temperature on phenology and pollen sterility of wheat varieties. Aust. J. Crop Sci. 2011, 5, 1039–1043. [Google Scholar]
- Hlavacova, M.; Klem, K.; Smutna, P.; Skarpa, P.; Hlavinka, P.; Novotna, K.; Rapantova, B.; Trnka, M. Effect of heat stress at anthesis on yield formation in winter wheat. Plant Soil Environ. 2017, 63, 139–144. [Google Scholar] [Green Version]
- Omidi, M.; Siahpoosh, M.R.; Mamghani, R.; Modarresi, M. The influence of terminal heat stress on meiosis abnormalities in pollen mother cells of wheat. Cytologia 2014, 79, 49–58. [Google Scholar] [CrossRef]
- Pradhan, G.P.; Prasad, P.V.V.; Fritz, A.K.; Kirkham, M.B.; Gill, B.S. Effects of drought and high temperature stress on synthetic hexaploid wheat. Funct. Plant Biol. 2012, 39, 190–198. [Google Scholar] [CrossRef]
- Liu, B.; Asseng, S.; Liu, L.L.; Tang, L.; Cao, W.X.; Zhu, Y. Testing the responses of four wheat crop models to heat stress at anthesis and grain filling. Glob. Chang. Biol. 2016, 22, 1890–1903. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Bramley, H.; Palta, J.A.; Siddique, K.H.M. Heat stress in wheat during reproductive and grain-filling phases. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.; Condon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Begcy, K.; Mariano, E.D.; Gentile, A.; Lembke, C.G.; Zingaretti, S.M.; Souza, G.M.; Menossi, M. A novel stress-induced sugarcane gene confers tolerance to drought, salt and oxidative stress in transgenic tobacco plants. PLoS ONE 2012, 7, e44697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattiello, L.; Begcy, K.; da Silva, F.R.; Jorge, R.A.; Menossi, M. Transcriptome analysis highlights changes in the leaves of maize plants cultivated in acidic soil containing toxic levels of Al(3+). Mol. Biol. Rep. 2014, 41, 8107–8116. [Google Scholar] [CrossRef] [PubMed]
- Begcy, K.; Walia, H. Drought stress delays endosperm development and misregulates genes associated with cytoskeleton organization and grain quality proteins in developing wheat seeds. Plant Sci. 2015, 240, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Fracheboud, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) grown at control or high temperature. Environ. Exp. Bot. 2004, 52, 123–129. [Google Scholar] [CrossRef]
- Cheng, C.H.; Mccomb, J.A. In vitro germination of wheat pollen on raffinose medium. New Phytol. 1992, 120, 459–462. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Liu, P.; Li, G.; Dong, S.T.; Wang, F.H.; Kong, L.A.; Zhang, J.W. Effect of heat stress on the photosynthetic characteristics in flag leaves at the grain-filling stage of different heat-resistant winter wheat varieties. J. Agron. Crop Sci. 2014, 200, 143–155. [Google Scholar] [CrossRef]
- Chytyk, C.J.; Hucl, P.J.; Gray, G.R. Leaf photosynthetic properties and biomass accumulation of selected western Canadian spring wheat cultivars. Can. J. Plant Sci. 2011, 91, 305–314. [Google Scholar] [CrossRef]
- Driever, S.M.; Lawson, T.; Andralojc, P.J.; Raines, C.A.; Parry, M.A.J. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes. J. Exp. Bot. 2014, 65, 4959–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, P.; Basnayake, J.; Inman-Bamber, G.; Lakshmanan, P.; Natarajan, S.; Stokes, C. Genetic variation in transpiration efficiency and relationships between whole plant and leaf gas exchange measurements in Saccharum spp. and related germplasm. J. Exp. Bot. 2016, 67, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Sadras, V.O.; Lawson, C.; Montoro, A. Photosynthetic traits in Australian wheat varieties released between 1958 and 2007. Field Crop Res. 2012, 134, 19–29. [Google Scholar] [CrossRef]
- Ford, K.L.; Cassin, A.; Bacic, A. Quantitative proteomic analysis of wheat cultivars with differing drought stress tolerance. Front. Plant Sci. 2011, 2, 44. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra, C.P.; Garcia-Pereira, M.J.; Gracia, M.P.; Igartua, E.; Casas, A.M.; Contreras-Moreira, B. Large differences in gene expression responses to drought and heat stress between elite barley cultivar Scarlett and a spanish landrace. Front. Plant Sci. 2017, 8, 647. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, B.; Jager, K.; Feher, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.W.; Wang, L.J.; Zhao, J.Y.; Fan, P.G.; Li, S.H. Effect and after-effect of water stress on the distribution of newly-fixed C-14-photoassimilate in micropropagated apple plants. Environ. Exp. Bot. 2007, 60, 484–494. [Google Scholar] [CrossRef]
- Sudhir, P.; Murthy, S.D.S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The plant heat stress transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Bharti, K.; von Koskull-Doring, P.; Bharti, S.; Kumar, P.; Tintschl-Korbitzer, A.; Treuter, E.; Nover, L. Tomato heat stress transcription factor HsfB1 represents a novel type of general transcription coactivator with a histone-like motif interacting with the plant CREB binding protein ortholog HAC1. Plant Cell 2004, 16, 1521–1535. [Google Scholar] [CrossRef] [PubMed]
- Giorno, F.; Wolters-Arts, M.; Grillo, S.; Scharf, K.D.; Vriezen, W.H.; Mariani, C. Developmental and heat stress-regulated expression of HsfA2 and small heat shock proteins in tomato anthers. J. Exp. Bot. 2010, 61, 453–462. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety Name | Country of Origin | Flowering Time |
|---|---|---|
| Kukri | Australia | 40 ± 1 day |
| Drysdale | Australia | 48 ± 1.5 days |
| Gladius | Australia | 49 ± 1.8 days |
| RAC875 | Australia | 45 ± 2.3 days |
| Epos | Czech Republic | 52 ± 2.4 days |
| Cornetto | Germany | 57 ± 1.1 days |
| Granny | Czech Republic | 50 ± 1.6 days |
| Chamsin | Germany | 53 ± 2 days |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Begcy, K.; Weigert, A.; Egesa, A.O.; Dresselhaus, T. Compared to Australian Cultivars, European Summer Wheat (Triticum aestivum) Overreacts When Moderate Heat Stress Is Applied at the Pollen Development Stage. Agronomy 2018, 8, 99. https://doi.org/10.3390/agronomy8070099
Begcy K, Weigert A, Egesa AO, Dresselhaus T. Compared to Australian Cultivars, European Summer Wheat (Triticum aestivum) Overreacts When Moderate Heat Stress Is Applied at the Pollen Development Stage. Agronomy. 2018; 8(7):99. https://doi.org/10.3390/agronomy8070099
Chicago/Turabian StyleBegcy, Kevin, Anna Weigert, Andrew Ogolla Egesa, and Thomas Dresselhaus. 2018. "Compared to Australian Cultivars, European Summer Wheat (Triticum aestivum) Overreacts When Moderate Heat Stress Is Applied at the Pollen Development Stage" Agronomy 8, no. 7: 99. https://doi.org/10.3390/agronomy8070099