Mineral Composition and Antioxidant Status of Tomato with Application of Selenium

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Irrigation Experiment
2.2. Foliar Application Experiment
2.3. Statistical Analyses
3. Results
3.1. Plant Biomass
3.1.1. Irrigation Experiment
3.1.2. Foliar Application Experiment
3.2. Macronutrient Content in the Different Organs of Tomato Plants
3.2.1. Irrigation Experiment
3.2.2. Foliar Application Experiment
3.3. Selenium Content in the Different Organs of Tomato Plants
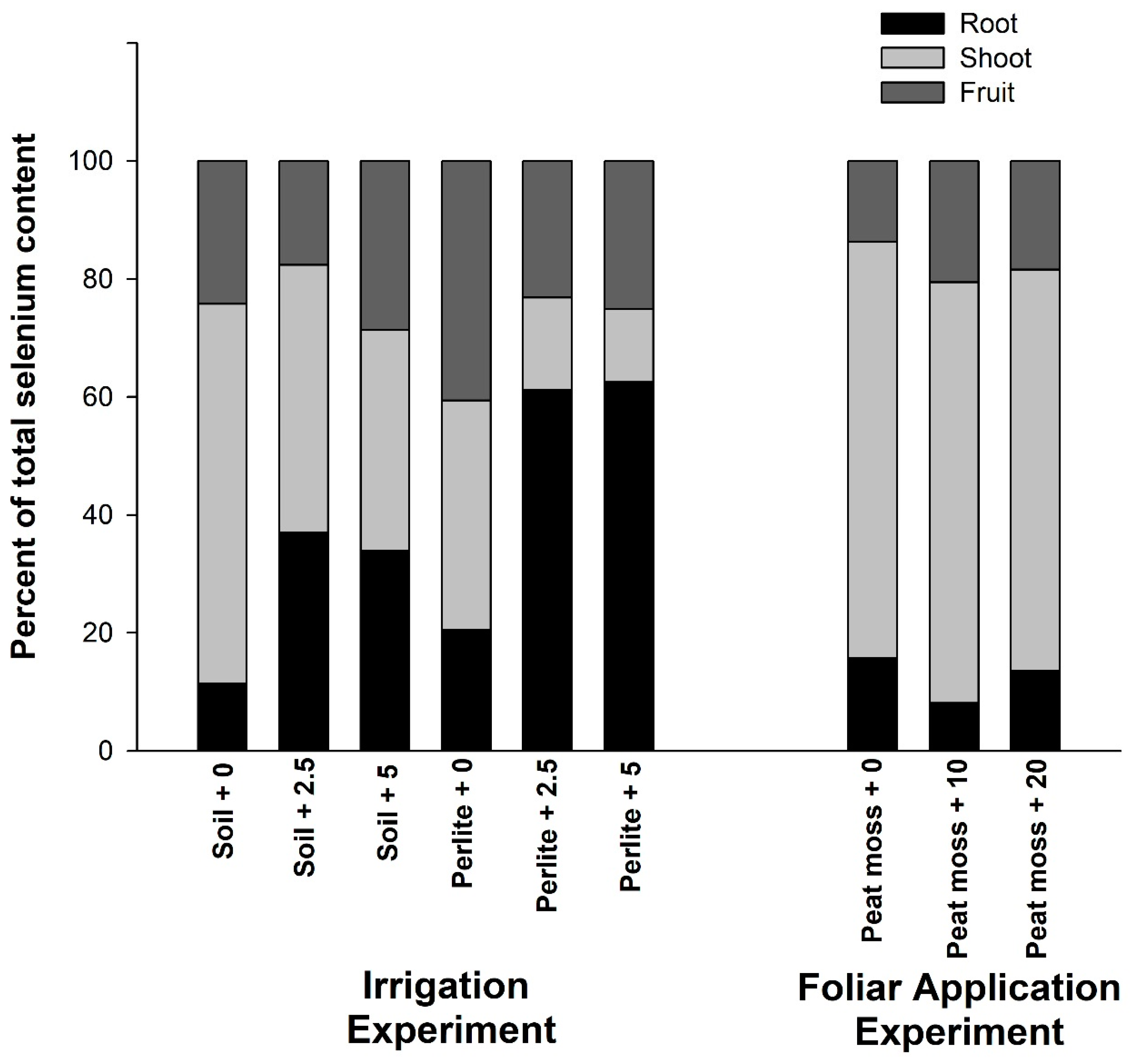
3.4. Relative Distribution of Selenium in Different Organs of the Plant
3.5. Total Antioxidant Capacity in Tomato Fruits
3.6. Correlation Matrix between the Concentrations of Selenium for the Different Organs of Tomato Plants against the Mineral Contents, Biomass, and Antioxidant Status of the Fruit
4. Discussion
4.1. Biomass
4.2. Mineral Concentrations
4.3. Selenium Concentration
4.4. Antioxidant Status in Tomato Fruits
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Great, B. Dietary Reference Values for Food Energy and Nutrients for the United Kingdom: Report of the Panel on Dietary Reference Values of the Committee on Medical Aspects of Food Policy, 8th ed.; Department of Health: London, UK, 1991.
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; McGrath, S.P.; Zhao, F.-J.; Breward, N. Biofortification of UK food crops with selenium. Proc. Nutr. Soc. 2006, 65, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F. Selenium in global food systems. Br. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H.E. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, F.M. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology; Selinus, O., Alloway, B., Centeno, J., Finkelman, R., Fuge, R., Lindh, U., Smedley, P., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 375–416. [Google Scholar]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Hartikainen, H.; Xue, T.; Piironen, V. Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil 2000, 225, 193–200. [Google Scholar] [CrossRef]
- Boldrin, P.; Faquin, V.; Ramos, S.; Boldrin, K.; Ávila, F.; Guilherme, L. Soil and foliar application of selenium in rice biofortification. J. Food Compos. Anal. 2013, 31, 238–244. [Google Scholar] [CrossRef]
- Schiavon, M.; dall’Acqua, S.; Mietto, A.; Pilon-Smits, E.A.H.; Sambo, P.; Masi, A.; Malagoli, M. Selenium fertilization alters the chemical composition and antioxidant constituents of tomato (Solanum lycopersicon L.). J. Agric. Food Chem. 2013, 61, 10542–10554. [Google Scholar] [CrossRef] [PubMed]
- Mimmo, T.; Tiziani, R.; Valentinuzzi, F.; Lucini, L.; Nicoletto, C.; Sambo, P.; Scampicchio, M.; Pii, Y.; Cesco, S. Selenium biofortification in fragaria × ananassa: Implications on strawberry fruits quality, content of bioactive health beneficial compounds and metabolomic profile. Front. Plant Sci. 2017, 8, 1887. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, E.; Cidade, M.; Heerdt, G.; Ribessi, R.; Morgon, N.; Cadore, S. Effect of selenite and selenate application on mineral composition of lettuce plants cultivated under hydroponic conditions: Nutritional balance overview using a multifaceted study. J. Braz. Chem. Soc. 2017, 29, 371–379. [Google Scholar] [CrossRef]
- Schiavon, M.; Berto, C.; Malagoli, M.; Trentin, A.; Sambo, P.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Selenium biofortification in radish enhances nutritional quality via accumulation of methyl-selenocysteine and promotion of transcripts and metabolites related to glucosinolates, phenolics, and amino acids. Front. Plant Sci. 2016, 7, 1371. [Google Scholar] [CrossRef] [PubMed]
- Germ, M.; Kreft, I.; Stibilj, V.; Urbanc-Berčič, O. Combined effects of selenium and drought on photosynthesis and mitochondrial respiration in potato. Plant Physiol. Biochem. 2007, 45, 162–167. [Google Scholar] [CrossRef] [PubMed]
- González-Morales, S.; Pérez-Labrada, F.; García-Enciso, E.L.; Leija-Martínez, P.; Medrano-Macías, J.; Dávila-Rangel, I.E.; Juárez-Maldonado, A.; Rivas-Martínez, E.N.; Benavides-Mendoza, A. Selenium and sulfur to produce allium functional crops. Molecules 2017, 22, 558. [Google Scholar] [CrossRef] [PubMed]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A.S. Selenium in Higher Plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Mitani, N.; Yamaji, N.; Shen, R.F.; Ma, J.F. Involvement of silicon influx transporter OsNIP2;1 in selenite uptake in rice. Plant Physiol. 2010, 153, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Galinha, C.; Sánchez-Martínez, M.; Pacheco, A.M.; do Carmo Freitas, M.; Coutinho, J.; Maçãs, B.; Almeida, A.S.; Pérez-Corona, M.T.; Madrid, Y.; Wolterbeek, H.T. Characterization of selenium-enriched wheat by agronomic biofortification. Journal of food science and technology. J. Food Sci. Technol. 2015, 52, 4236–4245. [Google Scholar] [CrossRef] [PubMed]
- Cartes, P.; Gianfreda, L.; Mora, M.L. Uptake of selenium and its antioxidant activity in ryegrass when applied as selenate and selenite forms. Plant Soil 2005, 276, 359–367. [Google Scholar] [CrossRef]
- Zhang, M.; Tang, S.; Huang, X.; Zhang, F.; Pang, Y.; Huang, Q. Selenium uptake, dynamic changes in selenium content and its influence on photosynthesis and chlorophyll fluorescence in rice (Oryza sativa L.). Environ. Exp. Bot. 2014, 107, 39–45. [Google Scholar] [CrossRef]
- Mozafariyan, M.; Mohammad, M.K.; Hawrylak-Nowak, B. Ameliorative effect of selenium on tomato plants grown under salinity stress. Arch. Agron. Soil Sci. 2016, 62, 1368–1380. [Google Scholar] [CrossRef]
- Jiang, Y.; Zeng, Z.H.; Bu, Y.; Ren, C.Z.; Li, J.Z.; Han, J.J.; Tao, C.; Zhang, K.; Wang, X.X.; Lu, G.X.; et al. Effects of selenium fertilizer on grain yield, Se uptake and distribution in common buckwheat (Fagopyrum esculentum Moench). Plant Soil Environ. 2015, 61, 371–377. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef] [Green Version]
- The United States Department of Agriculture. United States Standards for Grades of Fresh Tomatoes; Fresh Products Branch: Washington, DC, USA, 1997.
- Association of Official Agricultural Chemists. Official Methods of Analysis, 15th ed.; AOAC: Washington, DC, USA, 1990; ISBN 9780935584752. [Google Scholar]
- Fick, K.R.; Miller, S.M.; Funk, J.D.; McDowell, L.R.; Houser, R.H. Methods of Mineral Analysis for Plant and Animal Tissues; AID Publication: Gainesville, FL, USA, 1976. [Google Scholar]
- Association of Official Agricultural Chemists. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Miller, N.J.; Rice-Evans, C.; Davies, M.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [PubMed]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer Texts in Statistics 103; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7137-0. [Google Scholar]
- Hamilton, S.J. Review of selenium toxicity in the aquatic food chain. Sci. Total Environ. 2004, 326, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Lehotai, N.; Kolbert, Z.; Peto, A.; Feigl, G.; Ordog, A.; Kumar, D.; Tari, I.; Erdei, L. Selenite-induced hormonal and signalling mechanisms during root growth of Arabidopsis thaliana L. J. Exp. Bot. 2012, 63, 5677–5687. [Google Scholar] [CrossRef] [PubMed]
- Simojoki, A.; Xue, T.; Lukkari, K.; Pennanen, A.; Hartikainen, H. Allocation of added selenium in lettuce and its impact on roots. Agric. Food Sci. 2003, 12, 155–164. [Google Scholar] [CrossRef]
- Randle, W.M.; Block, E.; Littlejohn, M.H.; Putman, D.; Bussard, M.L. Onion (Allium cepa L.) Thiosulfinates respond to increasing sulfur fertility. J. Agric. Food Chem. 1994, 42, 2085–2088. [Google Scholar] [CrossRef]
- Rios, J.; Rosales, M.A.; Blasco, B.M.; Cervilla, L.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and growth-promoting effect of selenium on senescing lettuce. Plant Soil 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Kápolna, E.; Laursen, K.; Husted, S.; Larsen, E. Bio-fortification and isotopic labelling of Se metabolites in onions and carrots following foliar application of Se and 77Se. Food Chem. 2012, 133, 650–657. [Google Scholar] [CrossRef]
- Kápolna, E.; Hillestrøm, P.R.; Laursen, K.H.; Husted, S.; Larsen, E.H. Effect of foliar application of selenium on its uptake and speciation in carrot. Food Chem. 2009, 115, 1357–1363. [Google Scholar] [CrossRef]
- Hladun, K.; Parker, D.; Tran, K.; Trumble, J. Effects of selenium accumulation on phytotoxicity, herbivory, and pollination ecology in radish (Raphanus sativus L.). Environ. Pollut. 2013, 172, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Põldma, P.; Tõnutare, T.; Viitak, A.; Luik, A.; Moor, U. Effect of selenium treatment on mineral nutrition, bulb size, and antioxidant properties of garlic (Allium sativum L.). J. Agric. Food Chem. 2011, 59, 5498–5503. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Jianzhou, C.; Xueli, H.; Binbin, L.; Jingmin, L.; Zhaowei, Y. Effects of selenium on agronomical characters of winter wheat exposed to enhanced ultraviolet-B. Ecotoxicol. Environ. Saf. 2013, 92, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, T.; Bayoumi, Y.; Alshaal, T.; Elhawat, N.; Sztrik, A.; El-Ramady, H. Selenium fortification induces growth, antioxidant activity, yield and nutritional quality of lettuce in salt-affected soil using foliar and soil applications. Plant Soil 2017, 421, 245–258. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Lin, Z.-Q.; Broadley, M. Selenium Biofortification. In Selenium in Plants; Pilon-Smits, E.A.H., Winkel, H.E., Lin, Z.-Q., Eds.; Springer: Cham, Sweden, 2017; pp. 231–255. [Google Scholar]
- Castillo-Godina, R.G.; Foroughbakhch-Pournavab, R.; Benavides-Mendoza, A. Effect of selenium on elemental concentration and antioxidant enzymatic activity of tomato plants. J. Agric. Sci. Technol. 2016, 18, 233–244. [Google Scholar]
- Smoleń, S.; Kowalska, I.; Sady, W. Assessment of biofortification with iodine and selenium of lettuce cultivated in the NFT hydroponic system. Sci. Hortic. 2014, 166, 9–16. [Google Scholar] [CrossRef]
- Iqbal, M.; Iqbal, H.; Liaqata, H.; Arslan Ashraf, M.; Rasheed, R.; Rehman, A.U. Exogenously applied selenium reduces oxidative stress and induces heat tolerance in spring wheat. Plant Physiol. Biochem. 2015, 94, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Pazurkiewicz-Kocot, K.; Galas, W.; Kita, A. The effect of selenium on the accumulation of some metals in Zea mays L. plants treated with indole-3-acetic acid. Cell. Mol. Biol. Lett. 2003, 8, 97–103. [Google Scholar] [PubMed]
- Hawrylak-Nowak, B. Effect of selenium on selected macronutrients in maize plants. J. Elem. 2008, 13, 513–519. [Google Scholar]
- Feng, R.; Wei, C.; Tu, S.; Wu, F. Effects of Se on the uptake of essential elements in Pteris vittata L. Plant Soil 2009, 325, 123–132. [Google Scholar] [CrossRef]
- Nawaz, F.; Ahmad, R.; Ashraf, M.; Waraich, E.; Khan, S. Effect of selenium foliar spray on physiological and biochemical processes and chemical constituents of wheat under drought stress. Ecotoxicol. Environ. Saf. 2015, 113, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Petra, M., Ed.; Academic Press: Boston, MA, USA, 2012; p. 89. [Google Scholar]
- Campbell, C.R. Reference Sufficiency Ranges for Plant Analysis in the Southern Region of the United States; Southern Region Agricultural Experiment Station: Raleigh, NC, USA, 2000; p. 134. [Google Scholar]
- Marles, R. Mineral nutrient composition of vegetables, fruits and grains: The context of reports of apparent historical declines. J. Food Compos. Anal. 2017, 56, 93–103. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; Taylor & Francis Group: Oxford, UK, 2011; ISBN 9781420093681. [Google Scholar]
- Pyrzynska, K. Selenium speciation in enriched vegetables. Food Chem. 2009, 114, 1183–1191. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Q.; Hu, C.; Zhao, X.; Duan, B.; Zhao, Z. Regulatory effects of sulfur on oilseed rape (Brassica napus L.) response to selenite. Soil Sci. Plant Nutr. 2016, 62, 247–253. [Google Scholar] [CrossRef]
- Asher, C.J.; Butler, G.W.; Peterson, P.J. Selenium transport in root Systems of tomato. J. Exp. Bot. 1977, 28, 279–291. [Google Scholar] [CrossRef]
- Li, H.-F.; McGrath, S.P.; Zhao, F.-J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Wang, Q.; Luo, Z.; Yu, Y.; Jiang, R.; Li, H. Effects of root iron plaque on selenite and selenate dynamics in rhizosphere and uptake by rice (Oryza sativa). Plant Soil 2015, 388, 255–266. [Google Scholar] [CrossRef]
- Valeria, P.; Alberto, A.; Paola, F.; Pifferi, P.G. Foliar application of selenite and selenate to potato (Solanum tuberosum): Effect of a ligand agent on selenium content of tubers. J. Agric. Food Chem. 2000, 48, 4749–4751. [Google Scholar] [CrossRef]
- Wojcik, P. Uptake of mineral nutrients from foliar fertilization. J. Fruit Ornam. Plant Res. 2004, 12, 201–218. [Google Scholar]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Kong, L.; Wang, M.; Bi, D. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress. Plant Growth Regul. 2005, 45, 155–163. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, Y.; Shi, G.; Zhang, X. Selenium delays tomato fruit ripening by inhibiting ethylene biosynthesis and enhancing the antioxidant defense system. Food Chem. 2017, 219, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Businelli, D.; D’Amato, R.; Onofri, A.; Tedeschini, E.; Tei, F. Se-enrichment of cucumber (Cucumis sativus L.), lettuce (Lactuca sativa L.) and tomato (Solanum lycopersicum L. Karst) through fortification in pre-transplanting. Sci. Hortic. 2015, 197, 697–704. [Google Scholar] [CrossRef]
- Kiferle, C.; Gonzali, S.; Holwerda, H.T.; Ibaceta, R.R.; Perata, P. Tomato fruits: A good target for iodine biofortification. Front. Plant Sci. 2013, 4, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Treatments Substrate + Se (mg L−1) | Fresh Weight of Roots | Root Dry Weight | Fresh Weight of Shoots | Shoot Dry Weight | Fruit Fresh Weight | Fruit Dry Weight |
|---|---|---|---|---|---|---|
| g plant−1 | g plant−1 | g plant−1 | ||||
| Irrigation Experiment 0, 2.5, and 5 mg L−1 Selenium in Fertilizer Solution | ||||||
| Soil + 0 | 416 b † | 54.0 c | 1964 a | 279.9 a | 2852 ab | 175.0 a |
| Soil + 2.5 | 417 b | 55.3 bc | 2049 a | 291.9 a | 2744 ab | 166.8 ab |
| Soil + 5 | 255 c | 33.3 d | 1405 b | 187.5 b | 2425 b | 149.6 b |
| Perlite + 0 | 488 a | 60.5 ab | 1509 b | 204.4 b | 2930 a | 185.7 a |
| Perlite + 2.5 | 489 a | 61.4 a | 1013 c | 128.5 c | 2872 a | 167.2 ab |
| Perlite + 5 | 154 d | 18.4 e | 430 d | 58.1 d | 939 c | 54.4 c |
| ANOVA significance | ||||||
| Substrate (S) | ns | ns | ** | ** | ** | ** |
| Concentration (C) | ** | ** | ** | ** | ** | ** |
| S × C | ** | ** | ** | ** | ** | ** |
| Foliar Application Experiment 0, 10, and 20 mg L−1 Foliar Selenium | ||||||
| Peat moss + 0 | 455 b | 47.4 a | 2126 c | 294.4 b | 2997 ab | 172.3 b |
| Peat moss + 10 | 510 a | 52.9 a | 2885 a | 402.1 a | 3148 a | 217.6 a |
| Peat moss + 20 | 535 a | 55.0 a | 2624 b | 366.8 a | 2840 b | 178.1 b |
| ANOVA significance | ||||||
| Treatment | * | ns | ** | ** | ns | ** |
| Irrigation Experiment 0, 2.5, and 5 mg L−1 Selenium in the Fertilizer Solution Substrate + Se (mg L−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Organ | Minerals | S + 0 | S + 2.5 | S + 5 | P + 0 | P + 2.5 | P + 5 | ANOVA significance | ||
| SB | C | SB × C | ||||||||
| Root | N | 26.6 b † | 32.1 a | 26.2 b | 24.5 b | 25.6 b | 21.1 c | ** | ** | ns |
| P | 1.9 b | 2.0 b | 1.8 b | 4.0 a | 4.0 a | 4.0 a | ** | ns | ns | |
| K | 17 a | 16 a | 20 a | 19 a | 20 a | 17 a | ns | ns | ns | |
| Ca | 11 a | 10 ab | 9 ab | 12 a | 7 b | 7 b | ns | * | ns | |
| Mg | 0.9 ab | 0.8 ab | 0.8 ab | 1.4 a | 0.5 ab | 0.3 b | ns | ns | ns | |
| Shoot | N | 26.8 b | 32.2 a | 26.3 b | 24.7 b | 25.3 b | 21.2 c | ** | ** | * |
| P | 3.6 c | 4.7 b | 3.3 c | 4.2 bc | 7.5 a | 8.1 a | ** | ** | ** | |
| K | 17 abc | 21 a | 20 ab | 21 a | 15.6 bc | 15.2 c | ns | ns | * | |
| Ca | 21 a | 20 a | 21 a | 21 a | 23 a | 12 b | ns | ns | ns | |
| Mg | 1.0 b | 0.9 b | 1.0 b | 1.0 b | 1.6 a | 0.6 b | ns | * | * | |
| Fruit | N | 26.9 b | 32.4 a | 26.4 b | 24.8 b | 25.4 b | 21.3 c | ** | ** | * |
| P | 4.5 a | 4.6 a | 4.7 a | 4.8 a | 5.5 a | 5.5 a | ns | ns | ns | |
| K | 29 ab | 29 ab | 20 c | 34 a | 28 ab | 22 bc | ns | ** | ns | |
| Ca | 2.8 b | 2.7 b | 2.6 b | 3.3 ab | 2.6 b | 3.9 a | ns | ns | ns | |
| Mg | 0.2 a | 0.2 a | 0.2 a | 0.2 a | 0.2 a | 0.1 b | ** | ** | ** | |
| Foliar Application Experiment 0, 10, and 20 mg L−1 Foliar Selenium Substrate + Se (mg L−1) | |||||
|---|---|---|---|---|---|
| Organ | Minerals | PM + 0 | PM + 10 | PM + 20 | ANOVA significance |
| Root | N | 27 a † | 26 a | 24 a | ns |
| P | 2.4 a | 2.6 a | 2.4 a | ns | |
| K | 18 a | 23 a | 22 a | ns | |
| Ca | 25 a | 27 a | 25 a | ns | |
| Mg | 0.7 a | 0.7 a | 0.7 a | ns | |
| Shoot | N | 29 a | 25 a | 27 a | ns |
| P | 3.3 b | 2.9 b | 4.0 a | ** | |
| K | 21 a | 24 a | 24 a | ns | |
| Ca | 30 a | 32 a | 30 a | ns | |
| Mg | 0.7 ab | 1.0 a | 0.6 b | * | |
| Fruit | N | 29 a | 25 a | 27 a | ns |
| P | 5.6 a | 5.3 a | 5.8 a | ns | |
| K | 27 a | 26 a | 30 a | ns | |
| Ca | 5.8 a | 5.4 a | 6.8 a | ns | |
| Mg | 0.1 a | 0.1 a | 0.1 a | ns | |
| Treatments Substrate + Se (mg L−1) | Root | Shoot | Fruit | Extraction by Plant |
|---|---|---|---|---|
| µg g−1 | µg g−1 | µg g−1 | µg plant−1 | |
| Irrigation Experiment (0, 2.5, and 5 mg L−1 Selenium in Fertilizer Solution) | ||||
| Soil + 0 | 8.30 d † | 9.06 d | 5.38 e | 3939 bc |
| Soil + 2.5 | 66.56 c | 15.39 b | 10.50 d | 9938 a |
| Soil + 5 | 93.18 b | 18.30 a | 17.44 b | 9168 a |
| Perlite + 0 | 9.80 d | 5.66 e | 6.22 e | 2934 c |
| Perlite + 2.5 | 101.77 b | 12.36 c | 14.11 c | 10,180 a |
| Perlite + 5 | 148.90 a | 9.48 d | 20.74 a | 4419 b |
| ANOVA significance | ||||
| Substrate | ** | ** | ** | ** |
| Concentration | ** | ** | ** | ** |
| S × C | ** | ** | ns | ** |
| Foliar Application Experiment (0, 10, and 20 mg L−1 Foliar Selenium) | ||||
| Peat moss + 0 | 15.60 b | 11.30 a | 3.87 b | 4725 b |
| Peat moss + 10 | 15.13 b | 20.43 a | 9.83 a | 11,331 a |
| Peat moss + 20 | 26.90 a | 20.33 a | 11.33 a | 10,964 a |
| ANOVA significance | ||||
| Treatment | * | ns | ** | * |
| Irrigation Experiment (0, 2.5, and 5 mg L−1 Selenium in Fertilizer Solution) | Foliar Application Experiment (0, 10, and 20 mg L−1 Foliar Selenium) | ||
|---|---|---|---|
| Treatment Substrate + Se (mg L−1) | Total Antioxidant Capacity (mM mg−1) | Treatment Substrate + Se (mg L−1) | Total Antioxidant Capacity (mM mg−1) |
| Soil + 0 | 2.88 d † | Peat moss + 0 | 2.85 c |
| Soil + 2.5 | 3.68 ab | Peat moss + 10 | 3.26 b |
| Soil + 5 | 3.98 a | Peat moss + 20 | 3.86 a |
| Perlite + 0 | 3.25 c | ||
| Perlite + 2.5 | 3.44 bc | ||
| Perlite + 5 | 3.53 bc | ||
| ANOVA significance | |||
| Substrate (S) | ns | ||
| Concentration (C) | ** | ||
| S × C | ** | Treatment | ** |
| Irrigation Experiment (0, 2.5 and 5 mg L−1 Se in Fertilizer Solution) | Foliar Application Experiment (0, 10 and 20 mg L−1 Foliar Se) | ||||||
|---|---|---|---|---|---|---|---|
| Variables | Se-Root | Se-Shoot | Se-Fruit | Se-Root | Se-Shoot | Se-Fruit | |
| Root | N | −0.44 | 0.40 | −0.44 | −0.22 | −0.64 | −0.77 * |
| P | 0.33 | −0.47 | 0.19 | −0.22 | −0.08 | 0.28 | |
| K | 0.23 | 0.19 | 0.26 | 0.12 | 0.28 | 0.37 | |
| Ca | −0.64 | −0.05 | −0.59 | −0.55 | 0.04 | −0.27 | |
| Mg | −0.79 ** | 0.04 | −0.64 | 0.08 | −0.13 | −0.43 | |
| Se | 1 | 0.33 | 0.91 ** | 1 | 0.13 | 0.55 | |
| Shoot | N | −0.44 | 0.39 | −0.46 | −0.22 | −0.64 | −0.77 * |
| P | 0.59 | −0.22 | 0.52 | 0.55 | 0.39 | 0.47 | |
| K | −0.41 | 0.12 | −0.25 | 0.17 | 0.11 | 0.03 | |
| Ca | −0.32 | 0.23 | −0.31 | −0.08 | −0.13 | −0.05 | |
| Mg | −0.26 | 0.06 | −0.33 | −0.61 | 0.26 | −0.02 | |
| Se | 0.33 | 1 | 0.40 | 0.13 | 1 | 0.74 * | |
| Fruit | N | −0.41 | 0.43 | −0.45 | −0.15 | −0.79 * | −0.78 * |
| P | 0.54 | −0.07 | 0.36 | 0.43 | −0.31 | 0.0 | |
| K | −0.60 | −0.33 | −0.66 | 0.15 | −0.13 | 0.35 | |
| Ca | 0.20 | −0.50 | 0.07 | 0.51 | −0.21 | 0.15 | |
| Mg | −0.65 | 0.27 | −0.65 | 0.09 | 0.83 ** | 0.69 | |
| Se | 0.91 ** | 0.40 | 1 | 0.55 | 0.74 * | 1 | |
| ANTIOX | 0.62 | 0.75 ** | 0.70 ** | 0.67 | 0.54 | 0.88 * | |
| Biomass | RFW | −0.48 | −0.25 | −0.56 | 0.58 | 0.21 | 0.70 * |
| RDW | −0.50 | −0.24 | −0.56 | 0.58 | 0.15 | 0.72 * | |
| SFW | −0.77 ** | 0.12 | −0.75 ** | 0.07 | 0.69 | 0.52 | |
| SDW | −0.75 ** | 0.14 | −0.74 ** | 0.03 | 0.71 * | 0.58 | |
| FFW | −0.67 | −0.28 | −0.69 | −0.40 | 0.24 | 0.13 | |
| FDW | −0.78 ** | −0.30 | −0.76 ** | −0.23 | 0.33 | 0.22 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narváez-Ortiz, W.A.; Becvort-Azcurra, A.A.; Fuentes-Lara, L.O.; Benavides-Mendoza, A.; Valenzuela-García, J.R.; González-Fuentes, J.A. Mineral Composition and Antioxidant Status of Tomato with Application of Selenium. Agronomy 2018, 8, 185. https://doi.org/10.3390/agronomy8090185
Narváez-Ortiz WA, Becvort-Azcurra AA, Fuentes-Lara LO, Benavides-Mendoza A, Valenzuela-García JR, González-Fuentes JA. Mineral Composition and Antioxidant Status of Tomato with Application of Selenium. Agronomy. 2018; 8(9):185. https://doi.org/10.3390/agronomy8090185
Chicago/Turabian StyleNarváez-Ortiz, Willian Alfredo, Armando Ariel Becvort-Azcurra, Laura Olivia Fuentes-Lara, Adalberto Benavides-Mendoza, Jesús Rodolfo Valenzuela-García, and José Antonio González-Fuentes. 2018. "Mineral Composition and Antioxidant Status of Tomato with Application of Selenium" Agronomy 8, no. 9: 185. https://doi.org/10.3390/agronomy8090185