Using Plant Phenomics to Exploit the Gains of Genomics

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Advent of Phenomics
3. Phenotyping Bottlenecks
4. High-Precision Phenotyping and Automation
5. Phenotyping for Important Traits
5.1. Root Growth and Functions
5.2. Seedling Vigour
5.3. Plant Architecture
5.4. Leaf Area and Senescence
5.5. Leaf Water Potential
5.6. Chlorophyll Content
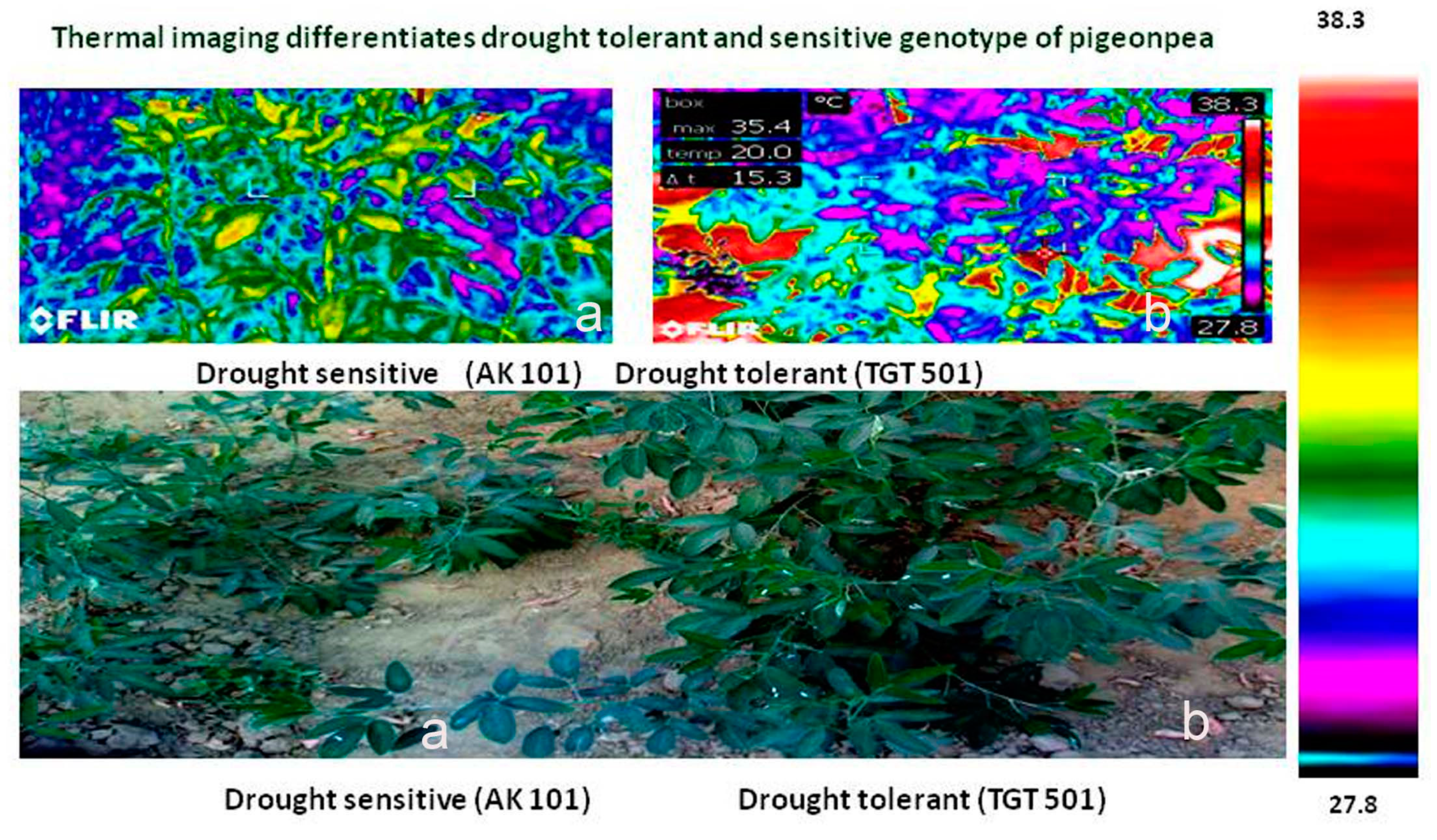
5.7. Canopy Temperature
5.8. Stomatal Conductance
5.9. Pollen Traits
5.10. Fruit Color
6. Plant Phenotyping Platforms
7. High Throughput Plant Phenotyping Platforms (HTPPS)
8. Connecting Genomics to Phenomics
9. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Sticklen, M.B. Feedstock crop genetic engineering for alcohol fuels. Crop Sci. 2007, 47, 2238–2248. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A. Predicting the impact of changing CO2 on crop yields: Some thoughts on food. New Phytol. 2007, 175, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Pieruschka, R.; Poorter, H. Phenotyping plants: Genes, phenes and machines. Funct. Plant Biol. 2012, 39, 813–820. [Google Scholar] [CrossRef]
- Chapman, C.S.; Merz, T.; Chan, A.; Jackway, P.; Hrabar, S.; Dreccer, F.M.; Holland, E.; Zheng, B.; Ling, J.T.; Jimenez-Berni, J. Pheno-Copter: A low-altitude, autonomous remote-sensing robotic helicopter for high-throughput field-based phenotyping. Agronomy 2014, 4, 279–301. [Google Scholar] [CrossRef]
- Cobb, J.N.; Declerck, G.; Greenberg, A.; Clark, R.; McCouch, S. Next-generation phenotyping: Requirements and strategies for enhancing our understanding of genotype-phenotype relationships and its relevance to crop improvement. Theor. Appl. Genet. 2013, 126, 867–887. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Czauderna, T.; Hoffmann, R.; Stein, N.; Schreiber, F. HTPheno: An image analysis pipeline for high-throughput plant phenotyping. BMC Bioinformatics 2011, 12, 148. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Duan, L.; Chen, G.; Xiong, L.; Liu, Q. Plant phenomics and high-throughput phenotyping: Accelerating rice functional genomics using multidisciplinary technologies. Curr. Opin. Plant Biol. 2013, 16, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.B.; Cheng, R.; Sirault, X.R.R.; Rungrat, T.; Murray, K.D.; Trtilek, M.; Furbank, R.T.; Badger, M.; Pogson, B.J.; Borevitz, J.O. TraitCapture: Genomic and environment modelling of plant phenomic data. Curr. Opin. Plant Biol. 2014, 18, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Furbank, R.; von Caemmerer, S.; Sheehy, J.; Edwards, G. C4 rice: A challenge for plant phenomics. Funct. Plant Biol. 2009, 36, 845–856. [Google Scholar] [CrossRef]
- Berger, B.; Parent, B.; Tester, M. High-throughput shoot imaging to study drought responses. J. Exp. Bot. 2010, 61, 3519–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehan, M.A.; Kellogg, E.A. High-throughput phenotyping. Am. J. Bot. 2017, 104, 505–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soule, M. Phenetics of natural populations I. Phenetic relationships of insular populations of the side-blotched lizard. Evolution 1967, 21, 584–591. [Google Scholar]
- Kumar, J.; Pratap, A.; Kumar, S. Plant phenomics: An overview. In Phenomics in Crop Plants: Trends, Options and Limitations; Kumar, J., Pratap, A., Kumar, S., Eds.; Springer: New Delhi, India, 2015; pp. 1–10. ISBN 978-81-322-2226-2. [Google Scholar]
- Johannsen, W. The genotype conception of heredity. Int. J. Epidemiol. 2014, 43, 989–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, A.; Liebisch, F.; Hund, A. Plant phenotyping: From bean weighing to image analysis. Plant Methods 2015, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. Statistical methods for research workers. In Breakthroughs in Statistics: Methodology and Distribution; Kotz, S., Johnson, N.L., Eds.; Springer: New York, NY, USA, 1992; pp. 66–70. ISBN 978-1-4612-4380-9. [Google Scholar]
- Pearson, C.H.; Ernst, S.M.; Barbarick, K.A.; Hatfield, J.L.; Peterson, G.A.; Buxton, D.R. Agronomy journal turns one hundred 1 manuscript tracker is an online, wed-based system for electronically submitting and reviewing manuscripts. Manuscript tracker is used by authors, reviewers, and editors. 2 HighWire Press (http://highwire.stanford.edu). Agron. J. 2008, 100, 1–8. [Google Scholar] [CrossRef]
- Fiorani, F.; Schurr, U. Future scenarios for plant phenotyping. Annu. Rev. Plant Biol. 2013, 64, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Swanson, C.P. Experimental studies on the nature of species III. Environmental responses of climatic races of Achillea. Jens Clausen, David D. Keck, William M. Hiesey. Q. Rev. Biol. 1949, 24, 144. [Google Scholar] [CrossRef]
- Griffiths, A.J.; Miller, J.H.; Suzuki, D.T.; Lewontin, R.C.; Gelbart, W.M. An Introduction to Genetic Analysis, 7th ed.; W. H. Freeman: Oxford, UK, 2000; ISBN 10: 0-7167-3520-2. [Google Scholar]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecol. Monogr. 1992, 62, 365–392. [Google Scholar] [CrossRef]
- Grime, J.P.; Hunt, R. Relative growth-rate: Its range and adaptive significance in a local flora. J. Ecol. 1975, 63, 393–422. [Google Scholar] [CrossRef]
- Poorter, H.; Remkes, C.; Lambers, H. Carbon and nitrogen economy of 24 wild species differing in relative growth rate. Plant Physiol. 1990, 94, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.S.; Srivastava, M.; Singh, P.; Porwal, P.; Kant, R.; Singh, J. High-precision phenotyping under controlled versus natural environments. In Phenomics in Crop Plants: Trends, Options and Limitations; Kumar, J., Pratap, A., Kumar, S., Eds.; Springer: New Delhi, India, 2015; pp. 27–40. ISBN 978-81-322-2226-2. [Google Scholar]
- Masuka, B.; Araus, J.L.; Das, B.; Sonder, K.; Cairns, J.E. Phenotyping for abiotic stress tolerance in maizeF. J. Integr. Plant Biol. 2012, 54, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Pratap, A.; Tomar, R.; Kumar, J.; Pandey, V.R.; Mehandi, S.; Katiyar, P.K. High-throughput plant phenotyping platforms. In Phenomics in Crop Plants: Trends, Options and Limitations; Kumar, J., Pratap, A., Kumar, S., Eds.; Springer: New Delhi, India, 2015; pp. 285–296. ISBN 978-81-322-2226-2. [Google Scholar]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot size matters: A meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef]
- White, J.W.; Andrade-Sanchez, P.; Gore, M.A.; Bronson, K.F.; Coffelt, T.A.; Conley, M.M.; Feldmann, K.A.; French, A.N.; Heun, J.T.; Hunsaker, D.J.; et al. Field-based phenomics for plant genetics research. F. Crop. Res. 2012, 133, 101–112. [Google Scholar] [CrossRef]
- Rahaman, M.M.; Chen, D.; Gillani, Z.; Klukas, C.; Chen, M. Advanced phenotyping and phenotype data analysis for the study of plant growth and development. Front. Plant Sci. 2015, 6, 619. [Google Scholar] [CrossRef] [PubMed]
- Nasarudin, N.E.M.; Helmi, S. Development and utilization of urban spectral library for remote sensing of urban environment. J. Urban Environ. Eng. 2011, 5, 44–56. [Google Scholar] [CrossRef] [Green Version]
- Fender, F.; Hanneken, M.; Der Stroth, S.I.; Linz, A.; Ruckelshausen, A. Sensor fusion meets gps: Individual plant detection. In Proceedings of the CIGR EurAgEng/VDI-MEG, Bonn, Germany, 3–7 September 2006; pp. 279–280. [Google Scholar]
- Pask, A.; Pietragalla, J.; Mullan, D.; Reynolds, M. Physiological Breeding II: A Field Guide to Wheat Phenotyping, 1st ed.; CIMMYT Mexico: Texcoco, Mexico, 2012; ISBN 978-970-648-182-5. [Google Scholar]
- Tsaftaris, S.; Noutsos, C. Plant phenotyping with low cost digital cameras and image analytics. In Information Technologies in Environmental Engineering; Springer: Berlin/Heidelberg, Germany, 2009; pp. 238–251. [Google Scholar]
- Lin, Y. LiDAR: An important tool for next-generation phenotyping technology of high potential for plant phenomics? Comput. Electron. Agric. 2015, 119, 61–73. [Google Scholar] [CrossRef]
- Kjaer, K.H.; Ottosen, C.-O. 3D Laser triangulation for plant phenotyping in challenging environments. Sensors 2015, 15, 13533–13547. [Google Scholar] [CrossRef] [PubMed]
- Dornbusch, T.; Hawkesford, M.; Jansen, M.; Nagel, K.; Niehaus, B.; Paulus, S.; Radermacher, M.; Sabermanseh, K.; Sadeghi-Tehran, P.; Schardt, C.; et al. Digital Field Phenotyping by LemnaTec. 2015; Unpublished work. [Google Scholar] [CrossRef]
- Engineers Make Wearable Sensors for Plants, Enabling Measurement of Water Use in Crops. Available online: https://shar.es/1N2bDU (accessed on 5 March 2019).
- Cohen, Y.; Alchanatis, V.; Meron, M.; Saranga, Y.; Tsipris, J. Estimation of leaf water potential by thermal imagery and spatial analysis*. J. Exp. Bot. 2005, 56, 1843–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozzani, R.; Iyer-Pascuzzi, A. Postembryonic control of root meristem growth and development. Curr. Opin. Plant Biol. 2014, 17, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J. Root Architecture and Plant Productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armengaud, P.; Zambaux, K.; Hills, A.; Sulpice, R.; Pattison, R.J.; Blatt, M.R.; Amtmann, A. EZ-Rhizo: Integrated software for the fast and accurate measurement of root system architecture. Plant J. 2009, 57, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 65, 6155–6166. [Google Scholar] [CrossRef] [PubMed]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [PubMed]
- Tracy, S.R.; Black, C.R.; Roberts, J.A.; Mooney, S.J. Soil compaction: A review of past and present techniques for investigating effects on root growth. J. Sci. Food Agric. 2011, 91, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Klepper, B.; Kaspar, T. Rhizotrons: Their Development and Use in Agricultural Research. Agron. J. 1994, 86, 745–753. [Google Scholar] [CrossRef]
- Iyer-Pascuzzi, A.; Simpson, J.; Herrera-Estrella, L.; Benfey, P.N. Functional genomics of root growth and development in Arabidopsis. Curr. Opin. Plant Biol. 2009, 12, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Yan, X.; Liao, H. 3D reconstruction and dynamic modeling of root architecture in situ and its application to crop phosphorus research. Plant J. 2009, 60, 1096–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.T.; MacCurdy, R.B.; Jung, J.K.; Shaff, J.E.; McCouch, S.R.; Aneshansley, D.J.; Kochian, L.V. Three-dimensional root phenotyping with a novel imaging and software platform. Plant Physiol. 2011, 156, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, M.; Sadashiva, A.T.; Laxmanan, V. Impact of water stress on root architecture in tomato (Solanum lycopersicum Mill). Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2095–2103. [Google Scholar] [CrossRef]
- Carretero, R.; Bert, F.E.; Podestá, G. Maize root architecture and water stress tolerance: An approximation from crop models. Agron. J. 2014, 106, 2287–2295. [Google Scholar] [CrossRef]
- Herridge, R.P.; Day, R.C.; Baldwin, S.; Macknight, R.C. Rapid analysis of seed size in Arabidopsis for mutant and QTL discovery. Plant Methods 2011, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, P.M.B.; Brunel-Muguet, S.; Dürr, C.; Huguet, T.; Demilly, D.; Wagner, M.-H.; Teulat-Merah, B. QTL analysis of seed germination and pre-emergence growth at extreme temperatures in Medicago truncatula. Theor. Appl. Genet. 2011, 122, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Blanco Romía, M.; Alcalà Bernàrdez, M. Chapter 3—Multivariate calibration for quantitative analysis. In Infrared Spectroscopy for Food Quality Analysis and Control; Sun, D.-W., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 51–82. ISBN 978-0-12-374136-3. [Google Scholar]
- Borisjuk, L.; Rolletschek, H.; Neuberger, T. Surveying the plant’s world by magnetic resonance imaging. Plant J. 2012, 70, 129–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolon, Y.-T.; Haun, W.J.; Xu, W.W.; Grant, D.; Stacey, M.G.; Nelson, R.T.; Gerhardt, D.J.; Jeddeloh, J.A.; Stacey, G.; Muehlbauer, G.J.; et al. Phenotypic and genomic analyses of a fast neutron mutant population resource in soybean. Plant Physiol. 2011, 156, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Tilly, N.; Aasen, H.; Bareth, G. Fusion of plant height and vegetation indices for the estimation of barley biomass. Remote Sens. 2015, 7, 11449–11480. [Google Scholar] [CrossRef]
- Berry, P.M.; Spink, J.H.; Foulkes, M.J.; Wade, A. Quantifying the contributions and losses of dry matter from non-surviving shoots in four cultivars of winter wheat. F. Crop. Res. 2003, 80, 111–121. [Google Scholar] [CrossRef]
- Blonquist, J.M.; Norman, J.M.; Bugbee, B. Automated measurement of canopy stomatal conductance based on infrared temperature. Agric. For. Meteorol. 2009, 149, 1931–1945. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Waldhoff, G.; Korres, W.; Curdt, C.; Bareth, G. Crop height variability detection in a single field by multi-temporal terrestrial laser scanning. Precis. Agric. 2016, 17, 296–312. [Google Scholar] [CrossRef]
- Tubana, B.; Girma, K.; Holtz, S.; Kanke, Y.; Lawles, K.; Edmonds, D.; Dotson, C.; England, B.; Daft, C.; Arnall, D.; et al. Indirect measurement of crop plant height. In Proceedings of the ASA-CSSA-SSAA, International Annual Meeting, New Orleans, LA, USA, 4–8 November 2007. [Google Scholar]
- Schima, R.; Mollenhauer, H.; Grenzdörffer, G.; Merbach, I.; Lausch, A.; Dietrich, P.; Bumberger, J. Imagine all the plants: Evaluation of a light-field camera for on-site crop growth monitoring. Remote Sens. 2016, 8, 823. [Google Scholar] [CrossRef]
- Madec, S.; Baret, F.; de Solan, B.; Thomas, S.; Dutartre, D.; Jezequel, S.; Hemmerlé, M.; Colombeau, G.; Comar, A. High-throughput phenotyping of plant height: Comparing unmanned aerial vehicles and ground LiDAR estimates. Front. Plant Sci. 2017, 8, 2002. [Google Scholar] [CrossRef] [PubMed]
- Bendig, J.; Bolten, A.; Bareth, G. UAV-based imaging for multi-temporal, very high resolution crop surface models to monitor crop growth variability. Photogramm. - Fernerkundung - Geoinf. 2013, 6, 551–562. [Google Scholar] [CrossRef]
- Remondino, F.; Spera, M.G.; Nocerino, E.; Menna, F.; Nex, F. State of the art in high density image matching. Photogramm. Rec. 2014, 29, 144–166. [Google Scholar] [CrossRef]
- Xu, R.; Dai, J.; Luo, W.; Yin, X.; Li, Y.; Tai, X.; Han, L.; Chen, Y.; Lin, L.; Li, G.; et al. A photothermal model of leaf area index for greenhouse crops. Agric. For. Meteorol. 2010, 150, 541–552. [Google Scholar] [CrossRef]
- Singh, B.; Reddy, K.R.; Redoña, E.D.; Walker, T. Screening of rice cultivars for morpho-physiological responses to early-season soil moisture stress. Rice Sci. 2017, 24, 322–335. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with drought: Stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 2015, 6, 542. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Bandopadhyay, P.; Bera, A.K. Identification of leaf based physiological markers for drought susceptibility during early seedling development of mungbean. Am. J. Plant Sci. 2016, 07, 1921–1936. [Google Scholar] [CrossRef]
- Mula, M.G.; Patil, S.B.; Aden, J.; Rathore, A.; Anil Kumar, V.; Kumar, R.V. Screening of pigeonpea genotypes for drought stress at early vegetative phase in Vertisol and Alfisol. Green Farming 2016, 7, 507–511. [Google Scholar]
- Belachew, K.Y.; Stoddard, F.L. Screening of faba bean (Vicia faba L.) accessions to acidity and aluminium stresses. PeerJ 2017, 5, e2963. [Google Scholar] [CrossRef] [PubMed]
- Howarth, C.J.; Gay, A.P.; Draper, J.; Powell, W. Development of high throughput plant phenotyping facilities at Aberystwyth. In Proceedings of the Phenomics Workshop, San Diego, CA, USA, 9 January 2011. [Google Scholar]
- Leport, L.; Musse, M.; Cambert, M.; De Franscesci, L.; Le Caherec, F.; Burel, A.; Mariette, F.; Bouchereau, A. Oilseed Rape leaf senescence phenotyping and identification of subcellular structural and metabolic changes using NMR tool. In Proceedings of the 2nd International Plant Phenotyping Symposium, Jülich, Germany, 5–7 September 2011. [Google Scholar]
- Waring, R.; Cleary, B.D. Plant moisture stress: Evaluation by pressure bomb. Science 1967, 155, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- De Bei, R.; Cozzolino, D.; Sullivan, W.; Cynkar, W.; Fuentes, S.; Dambergs, R.; Pech, J.; Tyerman, S. Non-destructive measurement of grapevine water potential using near infrared spectroscopy. Aust. J. Grape Wine Res. 2011, 17, 62–71. [Google Scholar] [CrossRef]
- Zakaluk, R.; Sri Ranjan, R. Predicting the leaf water potential of potato plants using RGB reflectance. Can. Biosyst. Eng. Le Genie des Biosyst. au Canada 2008, 50, 7.1–7.12. [Google Scholar]
- Patanè, C.; Scordia, D.; Testa, G.; Cosentino, S.L. Physiological screening for drought tolerance in Mediterranean long-storage tomato. Plant Sci. 2016, 249, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, S.; Grzesiak, M.T.; Filek, W.; Stabryła, J. Evaluation of physiological screening tests for breeding drought resistant triticale (x Triticosecale wittmack). Acta Physiol. Plant. 2003, 25, 29–37. [Google Scholar] [CrossRef]
- Puangbut, D.; Jogloy, S.; Vorasoot, N.; Akkasaeng, C.; Kesmalac, T.; Patanothai, A. Variability in yield responses of peanut (Arachis hypogaea L.) genotypes under early season drought. Asian J. Plant Sci. 2009, 8. [Google Scholar]
- Songsri, P.; Jogloy, S.; Holbrook, C.C.; Kesmala, T.; Vorasoot, N.; Akkasaeng, C.; Patanothai, A. Association of root, specific leaf area and SPAD chlorophyll meter reading to water use efficiency of peanut under different available soil water. Agric. Water Manag. 2009, 96, 790–798. [Google Scholar] [CrossRef]
- Samdur, M.Y.; Singh, A.; Mathur, R.; Manivel, P.; Chikani, B.M.; Gor, H.; Khan, M.A. Field evaluation of chlorophyll meter for screening groundnut (Arachis hypogaea L.) genotypes tolerant to iron-deficiency chlorosis. Curr. Sci. 2000, 79, 211–214. [Google Scholar]
- Vollmann, J.; Walter, H.; Sato, T.; Schweiger, P. Digital image analysis and chlorophyll metering for phenotyping the effects of nodulation in soybean. Comput. Electron. Agric. 2011, 75, 190–195. [Google Scholar] [CrossRef]
- Dwyer, L.; Tollenaar, M.; Houwing, L. A nondestructive method to monitor leaf greenness in corn. Can. J. Plant Sci. 1991, 71, 505–509. [Google Scholar] [CrossRef] [Green Version]
- Arunyanark, A.; Jogloy, S.; Vorasoot, N.; Chutipong, A.; Kesmala, T.; Patanothai, A. Stability of relationship between chlorophyll density and soil plant analysis development chlorophyll meter readings in peanut across different drought stress conditions. Asian J. Plant Sci. 2009, 8, 102–110. [Google Scholar]
- Dhyani, K.; Ansari, M.W.; Rao, Y.R.; Verma, R.S.; Shukla, A.; Tuteja, N. Comparative physiological response of wheat genotypes under terminal heat stress. Plant Signal. Behav. 2013, 8, e24564. [Google Scholar] [CrossRef] [PubMed]
- Jangpromma, N.; Songsri, P.; Thammasirirak, S.; Jaisil, P. Rapid assessment of chlorophyll content in sugarcane using a SPAD chlorophyll meter across different water stress conditions. Asian J. Plant Sci. 2010, 9, 368–374. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Identification of high-temperature tolerant lentil (Lens culinaris Medik.) genotypes through leaf and pollen traits. Front. Plant Sci. 2017, 8, 744. [Google Scholar] [CrossRef] [PubMed]
- Flood, P.J.; Kruijer, W.; Schnabel, S.K.; van der Schoor, R.; Jalink, H.; Snel, J.F.H.; Harbinson, J.; Aarts, M.G.M. Phenomics for photosynthesis, growth and reflectance in Arabidopsis thaliana reveals circadian and long-term fluctuations in heritability. Plant Methods 2016, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Yendrek, C.R.; Tomaz, T.; Montes, C.M.; Cao, Y.; Morse, A.M.; Brown, P.J.; McIntyre, L.M.; Leakey, A.D.B.; Ainsworth, E.A. High-Throughput phenotyping of maize leaf physiological and biochemical traits using hyperspectral reflectance. Plant Physiol. 2017, 173, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Van Bezouw, R.F.H.M.; Keurentjes, J.J.B.; Harbinson, J.; Aarts, M.G.M. Converging phenomics and genomics to study natural variation in plant photosynthetic efficiency. Plant J. 2019, 97, 112–133. [Google Scholar] [PubMed]
- Yadav, B.; Kumawat, C.; Verma, A.K.; Yadav, D.K.; Yadav, P. Fluorescence imaging for crop stress monitoring: A review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 568–575. [Google Scholar]
- Costa, J.M.; Grant, O.M.; Chaves, M.M. Thermography to explore plant–environment interactions. J. Exp. Bot. 2013, 64, 3937–3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deery, D.M.; Rebetzke, G.J.; Jimenez-Berni, J.A.; James, R.A.; Condon, A.G.; Bovill, W.D.; Hutchinson, P.; Scarrow, J.; Davy, R.; Furbank, R.T. Methodology for high-throughput field phenotyping of canopy temperature using airborne thermography. Front. Plant Sci. 2016, 7, 1808. [Google Scholar] [CrossRef] [PubMed]
- O’Shaughnessy, S.A.; Hebel, M.A.; Evett, S.R.; Colaizzi, P.D. Evaluation of a wireless infrared thermometer with a narrow field of view. Comput. Electron. Agric. 2011, 76, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Zia, S.; Romano, G.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Infrared thermal imaging as a rapid tool for identifying water-stress tolerant maize genotypes of different phenology. J. Agron. Crop Sci. 2013, 199, 75–84. [Google Scholar] [CrossRef]
- Alchanatis, V.; Cohen, Y.; Cohen, S.; Moller, M.; Sprinstin, M.; Meron, M.; Tsipris, J.; Saranga, Y.; Sela, E. Evaluation of different approaches for estimating and mapping crop water status in cotton with thermal imaging. Precis. Agric. 2010, 11, 27–41. [Google Scholar] [CrossRef]
- Orbegozo, H.O. Application of Thermography for the Assessment of Vineyard Water Status; Universidad De La Rioja: La Rioja, Spain, 2012. [Google Scholar]
- Wang, D.; Gartung, J. Infrared canopy temperature of early-ripening peach trees under postharvest deficit irrigation. Agric. Water Manag. 2010, 97, 1787–1794. [Google Scholar] [CrossRef]
- Kashiwagi, J.; L, K.; Upadhyaya, H.; Gaur, P. Rapid screening technique for canopy temperature status and its relevance to drought tolerance improvement in chickpea. J. SAT Agric. Res. 2008, 6, 104–105. [Google Scholar]
- Munns, R.; James, R.A.; Sirault, X.R.R.; Furbank, R.T.; Jones, H.G. New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J. Exp. Bot. 2010, 61, 3499–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macková, H.; Hronková, M.; Dobrá, J.; Turečková, V.; Novák, O.; Lubovská, Z.; Motyka, V.; Haisel, D.; Hájek, T.; Prášil, I.T.; et al. Enhanced drought and heat stress tolerance of tobacco plants with ectopically enhanced cytokinin oxidase/dehydrogenase gene expression. J. Exp. Bot. 2013, 64, 2805–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahnama, A.; James, R.A.; Poustini, K.; Munns, R. Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Funct. Plant Biol. 2010, 37, 255–263. [Google Scholar] [CrossRef]
- Jiang, Q.; Roche, D.; Monaco, T.; Hole, D. Stomatal conductance is a key parameter to assess limitations to photosynthesis and growth potential in barley genotypes. Plant Biol. (Stuttg.) 2006, 8, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Murtaza, N.; Shah, K.; Qayyum, A.; Ullah, I.; Malik, W. Genetic variation for stomatal conductance in upland cotton as influenced by heat-stressed and non-stressed growing regimes. Acta Agron. Hungarica 2008, 56, 11–19. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Diaz-Espejo, A.; Perez-Martin, A.; Hernández-Santana, V. Role of hydraulic and chemical signals in leaves, stems and roots in the stomatal behaviour of olive trees under water stress and recovery conditions. Tree Physiol. 2015, 35, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Heslop-Harrison, J.; Heslop-Harrison, Y.; Shivanna, K.R. The evaluation of pollen quality, and a further appraisal of the fluorochromatic (FCR) test procedure. Theor. Appl. Genet. 1984, 67, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Pressman, E.; Peet, M.M.; Pharr, D.M. The effect of heat stress on tomato pollen characteristics is associated with changes in carbohydrate concentration in the developing anthers. Ann. Bot. 2002, 90, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Heidmann, I.; Schade-Kampmann, G.; Lambalk, J.; Ottiger, M.; Di Berardino, M. Impedance flow cytometry: A novel technique in pollen analysis. PLoS ONE 2016, 11, e0165531. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, M.; Brewer, M.; Anderson, C.; Sullivan, D.; Gray, S.; Knaap, E. Tomato fruit shape analysis using morphometric and morphology attributes implemented in tomato analyzer software program. J. Am. Soc. Hortic. Sci. 2009, 134, 77–87. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Fukino, N. Image-based phenotyping: Use of colour signature in evaluation of melon fruit colour. Euphytica 2009, 171, 409. [Google Scholar] [CrossRef]
- Granier, C.; Aguirrezabal, L.; Chenu, K.; Cookson, S.J.; Dauzat, M.; Hamard, P.; Thioux, J.-J.; Rolland, G.; Bouchier-Combaud, S.; Lebaudy, A.; et al. PHENOPSIS, an automated platform for reproducible phenotyping of plant responses to soil water deficit in Arabidopsis thaliana permitted the identification of an accession with low sensitivity to soil water deficit. New Phytol. 2006, 169, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Harbinson, J.; Prinzenberg, A.E.; Kruijer, W.; Aarts, M.G.M. High throughput screening with chlorophyll fluorescence imaging and its use in crop improvement. Curr. Opin. Biotechnol. 2012, 23, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Deikman, J.; Petracek, M.; Heard, J.E. Drought tolerance through biotechnology: Improving translation from the laboratory to farmers’ fields. Curr. Opin. Biotechnol. 2012, 23, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Ruixiu, S.; Wilkerson, J.B.; Wilhelm, L.R.; Tompkins, F.D. A microcomputer-based morphometer for bush-type plants. Comput. Electron. Agric. 1989, 4, 43–58. [Google Scholar] [CrossRef]
- Montes, J.M.; Melchinger, A.E.; Reif, J.C. Novel throughput phenotyping platforms in plant genetic studies. Trends Plant Sci. 2007, 12, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Mccarthy, C.; Hancock, N.; Raine, S. Apparatus and infield evaluations of a prototype machine vision system for cotton plant internode length measurement. J. Cotton Sci. 2010, 14, 221–232. [Google Scholar]
- White, J.W.; Conley, M.M. A Flexible, Low-Cost Cart for Proximal Sensing. Crop Sci. 2013, 53, 1646–1649. [Google Scholar] [CrossRef]
- Andrade-Sanchez, P.; Gore, M.A.; Heun, J.T.; Thorp, K.R.; Carmo-Silva, A.E.; French, A.N.; Salvucci, M.E.; White, J.W. Development and evaluation of a field-based high-throughput phenotyping platform. Funct. Plant Biol. 2013, 41, 68–79. [Google Scholar] [CrossRef]
- Rascher, U.; Blossfeld, S.; Fiorani, F.; Jahnke, S.; Jansen, M.; Kuhn, A.; Matsubara, S.; Märtin, L.L.; Merchant, A.; Metzner, R.; et al. Non-invasive approaches for phenotyping of enhanced performance traits in bean. Funct. Plant Biol. 2011, 38, 968–983. [Google Scholar] [CrossRef] [Green Version]
- Losos, J.B.; Arnold, S.J.; Bejerano, G.; Brodie, E.D., III; Hibbett, D.; Hoekstra, H.E.; Mindell, D.P.; Monteiro, A.; Moritz, C.; Orr, H.A.; et al. Evolutionary biology for the 21st century. PLoS Biol. 2013, 11, e1001466. [Google Scholar] [CrossRef] [PubMed]
- De Diego, N.; Fürst, T.; Humplík, J.F.; Ugena, L.; Podlešáková, K.; Spíchal, L. An automated method for high-throughput screening of arabidopsis rosette growth in multi-well plates and its validation in stress conditions. Front. Plant Sci. 2017, 8, 1702. [Google Scholar] [CrossRef] [PubMed]
- Trachsel, S.; Kaeppler, S.M.; Brown, K.M.; Lynch, J.P. Shovelomics: High throughput phenotyping of maize (Zea mays L.) root architecture in the field. Plant Soil 2011, 341, 75–87. [Google Scholar] [CrossRef]
- Bai, G.; Ge, Y.; Hussain, W.; Baenziger, P.S.; Graef, G. A multi-sensor system for high throughput field phenotyping in soybean and wheat breeding. Comput. Electron. Agric. 2016, 128, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Pumphrey, M.; Zhou, J.; Gao, H.; Zhang, Q.; Sankaran, S. Development of Automated High-Throughput Phenotyping System for Controlled Environment Studies. In Proceedings of the ASABE Annual International Meeting, Spokane, WA, USA, 16–19 July 2017. [Google Scholar] [CrossRef]
- Yang, Y. Early detection of rice blast (Pyricularia) at seedling stage in Nipponbare rice variety using near-infrared hyper-spectral image. African J. Biotechnol. 2012, 11, 6809–6817. [Google Scholar] [CrossRef]
- Golzarian, M.R.; Frick, R.A.; Rajendran, K.; Berger, B.; Roy, S.; Tester, M.; Lun, D.S. Accurate inference of shoot biomass from high-throughput images of cereal plants. Plant Methods 2011, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LemnaTec. Available online: http://www.lemnatec.com/plant-phenotyping/ (accessed on 5 March 2019).
- KeyGene The Digital Phenotype. Available online: http://www.keygene.com/products-tech/digital-phenotyping/ (accessed on 5 March 2019).
- International Plant Phenotyping Network. Available online: https://www.plant-phenotyping.org/ (accessed on 5 March 2019).
- Jülich Plant Phenotyping Center. Available online: http://www.fz-juelich.de/ibg/ibg-2/EN/_organisation/JPPC/JPPC_node.html (accessed on 5 March 2019).
- Montpellier Plant Phenotyping Platform. Available online: https://www6.montpellier.inra.fr/lepse/Presentation-generale/Montpellier-Plant-Phenotyping-Platform-M3P (accessed on 5 March 2019).
- PPHD-INRA Dijon. Available online: http://www.dijon.inra.fr/Plateformes-Dispositifs/Plateformes-techniques/PPHD (accessed on 5 March 2019).
- La plate-forme PHENOPSIS. Available online: http://bioweb.supagro.inra.fr/phenopsis/InfoBDD.php (accessed on 5 March 2019).
- PhenoFab. Available online: https://www.keygene.com/technology/2-the-digital-phenotype/ (accessed on 5 March 2019).
- Biotron. Available online: https://www.uwo.ca/sci/research/biotron/ (accessed on 5 March 2019).
- Bresson, J.; Vasseur, F.; Dauzat, M.; Labadie, M.; Varoquaux, F.; Touraine, B.; Vile, D. Interact to survive: Phyllobacterium brassicacearum improves Arabidopsis tolerance to severe water deficit and growth recovery. PLoS ONE 2014, 9, e107607. [Google Scholar] [CrossRef] [PubMed]
- Clauw, P.; Coppens, F.; De Beuf, K.; Dhondt, S.; Van Daele, T.; Maleux, K.; Storme, V.; Clement, L.; Gonzalez, N.; Inzé, D. Leaf responses to mild drought stress in natural variants of Arabidopsis. Plant Physiol. 2015, 167, 800–816. [Google Scholar] [CrossRef] [PubMed]
- Tisné, S.; Serrand, Y.; Bach, L.; Gilbault, E.; Ben Ameur, R.; Balasse, H.; Voisin, R.; Bouchez, D.; Durand-Tardif, M.; Guerche, P.; et al. Phenoscope: An automated large-scale phenotyping platform offering high spatial homogeneity. Plant J. 2013, 74, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Gilmer, F.; Biskup, B.; Nagel, K.; Rascher, U.; Fischbach, A.; Briem, S.; Dreissen, G.; Tittmann, S.; Braun, S.; et al. Simultaneous phenotyping of leaf growth and chlorophyll fluorescence via GROWSCREENFLUORO allows detection of stress tolerance in. Funct. Plant Biol. 2009, 36, 902–914. [Google Scholar] [CrossRef]
- Nagel, K.; Putz, A.; Gilmer, F.; Kathrin, H.; Fischbach, A.; Pfeifer, J.; Marc, F.; Blossfeld, S.; Michaela, E.; Dimaki, C.; et al. GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 2012, 39, 891. [Google Scholar] [CrossRef]
- Reuzeau, C.; Pen, J.; Frankard, V.; De Wolf, J.; Peerbolte, R.; Broekaert, W.; Van Camp, W. TraitMill: A Discovery Engine for Identifying Yield-enhancement Genes in Cereals. Plant Gene Trait 2010, 1. [Google Scholar] [CrossRef]
- Sirault, X.; Fripp, J.; Paproki, A.; Kuffner, P.; Nguyen, C.; Li, R.; Daily, H.; Guo, J.; Furbank, R. PlantScan: A three-dimensional phenotyping platform for capturing the structural dynamic of plant development and growth. In Proceedings of the 7th International Conference on Functional-Structural Plant Models, Saariselkä, Finland, 9–14 June 2013. [Google Scholar]
- Neumann, K.; Klukas, C.; Friedel, S.; Rischbeck, P.; Chen, D.; Entzian, A.; Stein, N.; Graner, A.; Kilian, B. Dissecting spatiotemporal biomass accumulation in barley under different water regimes using high-throughput image analysis. Plant. Cell Environ. 2015, 38, 1980–1996. [Google Scholar] [CrossRef] [PubMed]
- Vadez, V.; Kholová, J.; Hummel, G.; Zhokhavets, U.; Gupta, S.K.; Hash, C.T. LeasyScan: A novel concept combining 3D imaging and lysimetry for high-throughput phenotyping of traits controlling plant water budget. J. Exp. Bot. 2015, 66, 5581–5593. [Google Scholar] [CrossRef] [PubMed]
- Pereyra Irujo, G.; Gasco, E.D.; Peirone, L.; Aguirrezábal, L. GlyPh: A low-cost platform for phenotyping plant growth and water use. Funct. Plant Biol. 2012, 39, 905. [Google Scholar] [CrossRef]
- Busemeyer, L.; Mentrup, D.; Möller, K.; Wunder, E.; Alheit, K.; Hahn, V.; Maurer, P.H.; Reif, C.J.; Würschum, T.; Müller, J.; et al. BreedVision — A multi-sensor platform for non-destructive field-based phenotyping in plant breeding. Sensors 2013, 13, 2830–2847. [Google Scholar] [CrossRef] [PubMed]
- Humplík, J.F.; Lazár, D.; Fürst, T.; Husičková, A.; Hýbl, M.; Spíchal, L. Automated integrative high-throughput phenotyping of plant shoots: A case study of the cold-tolerance of pea (Pisum sativum L.). Plant Methods 2015, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Phenospex: Smart Plant Analysis. Available online: https://phenospex.com/ (accessed on 5 March 2019).
- Burridge, J.; Jochua, C.N.; Bucksch, A.; Lynch, J.P. Legume shovelomics: High—Throughput phenotyping of common bean (Phaseolus vulgaris L.) and cowpea (Vigna unguiculata subsp, unguiculata) root architecture in the field. F. Crop. Res. 2016, 192, 21–32. [Google Scholar] [CrossRef]
- Sadok, W.; Naudin, P.; Boussuge, B.; Muller, B.; Welcker, C.; Tardieu, F. Leaf growth rate per unit thermal time follows QTL-dependent daily patterns in hundreds of maize lines under naturally fluctuating conditions. Plant. Cell Environ. 2007, 30, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.; Juan, J.; Chang, L.; Zhang, J.; Huang, D. Discrimination of plant root zone water status in greenhouse production based on phenotyping and machine learning techniques. Sci. Rep. 2017, 7, 8303. [Google Scholar] [CrossRef] [PubMed]
- Klukas, C.; Chen, D.; Pape, J.-M. Integrated analysis platform: An open-source information system for high-throughput plant phenotyping. Plant Physiol. 2014, 165, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Bylesjö, M.; Segura, V.; Soolanayakanahally, R.Y.; Rae, A.M.; Trygg, J.; Gustafsson, P.; Jansson, S.; Street, N.R. LAMINA: A tool for rapid quantification of leaf size and shape parameters. BMC Plant Biol. 2008, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- De Vylder, J.; Vandenbussche, F.; Hu, Y.; Philips, W.; Van Der Straeten, D. Rosette Tracker: An open source image analysis tool for automatic quantification of genotype effects. Plant Physiol. 2012, 160, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Weight, C.; Parnham, D.; Waites, R. TECHNICAL ADVANCE: LeafAnalyser: A computational method for rapid and large-scale analyses of leaf shape variation. Plant J. 2008, 53, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Bucksch, A.; Burridge, J.; York, L.M.; Das, A.; Nord, E.; Weitz, J.S.; Lynch, J.P. Image-based high-throughput field phenotyping of crop roots. Plant Physiol. 2014, 166, 470–486. [Google Scholar] [CrossRef] [PubMed]
- Galkovskyi, T.; Mileyko, Y.; Bucksch, A.; Moore, B.; Symonova, O.; Price, C.A.; Topp, C.N.; Iyer-Pascuzzi, A.S.; Zurek, P.R.; Fang, S.; et al. GiA Roots: Software for the high throughput analysis of plant root system architecture. BMC Plant Biol. 2012, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Green, J.M.; Appel, H.; Rehrig, E.M.; Harnsomburana, J.; Chang, J.-F.; Balint-Kurti, P.; Shyu, C.-R. PhenoPhyte: A flexible affordable method to quantify 2D phenotypes from imagery. Plant Methods 2012, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Pound, M.P.; French, A.P.; Atkinson, J.A.; Wells, D.M.; Bennett, M.J.; Pridmore, T. RootNav: Navigating images of complex root architectures. Plant Physiol. 2013, 162, 1802–1814. [Google Scholar] [CrossRef] [PubMed]
- Tanabata, T.; Shibaya, T.; Hori, K.; Ebana, K.; Yano, M. SmartGrain: High-throughput phenotyping software for measuring seed shape through image analysis. Plant Physiol. 2012, 160, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Lobet, G.; Pagès, L.; Draye, X. A Novel image-analysis toolbox enabling quantitative analysis of root system architecture. Plant Physiol. 2011, 157, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Le Bot, J.; Serra, V.; Fabre, J.; Draye, X.; Adamowicz, S.; Pagès, L. DART: A software to analyse root system architecture and development from captured images. Plant Soil 2010, 326, 261–273. [Google Scholar] [CrossRef]
- Mir, R.R.; Choudhary, N.; Singh, B.; Khandy, I.A.; Bawa, V.; Sofi, P.; Wani, A.; Kumari, S.; Jain, S.; Kumar, A. Harnessing genomics through phenomics. In Phenomics in Crop Plants: Trends, Options and Limitations; Kumar, J., Pratap, A., Kumar, S., Eds.; Springer: New Delhi, India, 2015; pp. 273–283. ISBN 978-81-322-2226-2. [Google Scholar]
- Furbank, R.T.; Tester, M. Phenomics–technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, R. What proportion of declared QTL in plants are false? Theor. Appl. Genet. 2004, 109, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Heffner, E.L.; Sorrells, M.E.; Jannink, J.-L. Genomic selection for crop improvement. Crop Sci. 2009, 49. [Google Scholar] [CrossRef]
- Jansen, R.C.; Van Ooijen, J.W.; Stam, P.; Lister, C.; Dean, C. Genotype-by-environment interaction in genetic mapping of multiple quantitative trait loci. Theor. Appl. Genet. 1995, 91, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topp, C.N.; Iyer-Pascuzzi, A.S.; Anderson, J.T.; Lee, C.-R.; Zurek, P.R.; Symonova, O.; Zheng, Y.; Bucksch, A.; Mileyko, Y.; Galkovskyi, T.; et al. 3D phenotyping and quantitative trait locus mapping identify core regions of the rice genome controlling root architecture. Proc. Natl. Acad. Sci. USA 2013, 110, E1695–E1704. [Google Scholar] [CrossRef] [PubMed]
- Shorinola, O.; Kaye, R.; Golan, G.; Peleg, Z.; Kepinski, S.; Uauy, C. Isolation and characterisation of mutants with altered seminal root numbers in hexaploid wheat. bioRxiv 2018, 364018. [Google Scholar]
- Oakley, C.G.; Savage, L.; Lotz, S.; Larson, G.R.; Thomashow, M.F.; Kramer, D.M.; Schemske, D.W. Genetic basis of photosynthetic responses to cold in two locally adapted populations of Arabidopsis thaliana. J. Exp. Bot. 2018, 69, 699–709. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Silva, F.M.; Lichtenstein, G.; Alseekh, S.; Rosado-Souza, L.; Conte, M.; Suguiyama, V.F.; Lira, B.S.; Fanourakis, D.; Usadel, B.; Bhering, L.L.; et al. The genetic architecture of photosynthesis and plant growth-related traits in tomato. Plant. Cell Environ. 2018, 41, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Borrill, P.; Harrington, S.A.; Uauy, C. Applying the latest advances in genomics and phenomics for trait discovery in polyploid wheat. Plant J. 2019, 97, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Battenfield, S.D.; Guzmán, C.; Gaynor, R.C.; Singh, R.P.; Peña, R.J.; Dreisigacker, S.; Fritz, A.K.; Poland, J.A. Genomic Selection for processing and end-use quality traits in the CIMMYT spring bread wheat breeding program. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.; Mohan, S.M.; Gaur, P.; Chamarthi, S.; Singh, V.; Samineni, S.; Swapna, N.; Sharma, M.; Pande, S.; Singh, S.; et al. Marker-assisted backcrossing to introgress resistance to fusarium wilt race 1 and Ascochyta blight in C 214, an elite cultivar of chickpea. Plant Genome 2014, 7, 1–11. [Google Scholar] [CrossRef]
- Pratap, A.; Chaturvedi, S.K.; Tomar, R.; Rajan, N.; Malviya, N.; Thudi, M.; Saabale, P.R.; Prajapati, U.; Varshney, R.K.; Singh, N.P. Marker-assisted introgression of resistance to fusarium wilt race 2 in Pusa 256, an elite cultivar of desi chickpea. Mol. Genet. Genomics 2017, 292, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Bosquet, L.; Crossa, J.; von Zitzewitz, J.; Serret, M.D.; Luis Araus, J. High-throughput phenotyping and genomic selection: The frontiers of crop breeding converge. J. Integr. Plant Biol. 2012, 54, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Heslot, N.; Yang, H.-P.; Sorrells, M.E.; Jannink, J.-L. Genomic selection in plant breeding: A comparison of models. Crop Sci. 2012, 52, 146–160. [Google Scholar] [CrossRef]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic selection in plant breeding: Methods, models, and perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Elías, M.J.; García-Arata, M.I.; Munoz, V.; Santos, I.; Sanz-Moreno, J.; Abraira, V.; Arribas, J.; González García, J.; Moreno, A.; Dronda, F.; et al. Phenotype or virtual phenotype for choosing antiretroviral therapy after failure: A prospective, randomized study. Antivir. Ther. 2003, 8, 577–584. [Google Scholar] [PubMed]
- Tsukor, V.; Klose, R.; Ruckelshausen, A. Multi-sensor simulation method for outdoor plant phenotyping based on autonomous field robots. In Proceedings of the CIGR-Agicultural Enineering Conference, Valencia, Spain, 8–12 July 2012. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Name | Target Plant Organ | Parameters | Description | References |
|---|---|---|---|---|
| PHENOPSIS | Leaf | Plant growth parameters | An automated platform for reproducible phenotyping of plant responses to soil water deficit in Arabidopsis thaliana | [114,139] |
| WIWAM | Leaf | Growth parameters | Used to impose stress early during leaf development | [140] |
| PHENOSCOPE | Shoots | Vegetative growth and homogeneity | An integrated device, allowing a simultaneous culture of individual Arabidopsis plants and high-throughput acquisition, storage, and analysis of quality phenotypes | [141] |
| GROWSCREEN | Leaf | 3D surface area of leaf discs | Platform to study plant leaf growth fluorescence and root architecture from seedling under control conditions in Arabidopsis thaliana, barley and maize | [142,143] |
| TraitMill | Flowers, grains, etc. | Growth and yield parameters | Automated high resolution phenotypic platform, uniquely placed to identify genes that improve the yield of cereals | [144] |
| PlantScan | Whole plant | Vegetative growth parameters | Automated high-resolution phenomic center providing non-invasive analysis of plant structure, morphology and function in Gossypium, wheat and maize | [145] |
| LemnaTec | Leaf | Growth and yield parameters | Visualize and analyze 2D/3D non-destructive high-throughput imaging, monitor plant growth and behavior under fully controlled conditions | [146] |
| LeasyScan | Leaf, whole plant | Canopy traits | Phenotyping for traits controlling plant water use with precision in pearl millet | [147] |
| HRPF | Whole plant | Growth and yield parameters | High-throughput rice phenotyping facility | [128] |
| GlyPh (self-construction) | Whole plant | Soil water content and growth estimation | Low-cost platform for phenotyping plant growth and water use under a broad range of conditions | [148] |
| BreedVision | Whole plant | Growth and physiological parameters | Measures various agronomic traits and leads to non-destructive phenotyping for crop improvement and plant genetic studies | [149] |
| PlantScreenTM | Shoot | Chlorophyll fluorescence imaging and non-imaging chlorophyll fluorescence, growth parameters | Evaluates various parameters of chlorophyll fluorescence obtained from kinetic chlorophyll fluorescence imaging | [150] |
| OloPhen | Whole plant | Rosette area, growth and survival rate | Suitable for analysis of rosette growth in multi-well plates, suitable to evaluate plant stress tolerance. | [124] |
| Color eye (RBG scanner) | Leaf | Leaf greenness, lesions | Data can be overlayed over laser triangulation data obtained by plant eye | [151] |
| LabVIEW | Canopy | Growth parameters | Low-cost, accurate, and high-throughput phenotyping system with custom algorithms | [126,127] |
| Shovelomics | Root | Root growth parameters | Identification and selection of useful root architectural phenotypes for annual legume or dicotyledonous crops. | [152] |
| Phenodyn/Phenoarch | Leaf | Leaf elongation rate | Follows QTL-dependent daily patterns in maize lines under naturally fluctuating conditions, located in INRA, France | [153] |
| LemnaGrid | Root and leaf | Plant and root growth parameters | Compares growth behaviors of different genotypes, discriminates plant root zone water status | [154] |
| Integrated Analysis Platform (IAP) | Leaf | Plant leaf orientation | Provides user-friendly interfaces with highly adaptable core functions, supports image data transfer from different acquisition environments and large-scale image analysis | [155] |
| LAMINA | Leaf | Leaf parameters | Tool for automated analysis of images of leaves, designed to provide classical indicators of leaf structure | [156] |
| Rosette Tracker | Shoot | Area, perimeter diameter stockiness | Allows to simultaneously quantify plant growth, photosynthesis, and leaf temperature-related parameters | [157] |
| Leaf Analyser | Leaf | Leaf architecture | Provides a high-throughput method to evaluate leaf shape variation in higher-dimensional phenotypic space | [158] |
| Self-construction | Root | Root growth parameters | Algorithms allow the automatic extraction of many root traits in a high-throughput fashion | [159] |
| Phenovator | Leaf | Photosynthesis | High-throughput phenotyping facility for photosynthesis developed at Wageningen University and Research | [91] |
| Name of the Software | Target Plant Organ | Parameters | Description | References |
|---|---|---|---|---|
| MATLAB | Leaf | Leaf architecture | Uses image processing algorithms for high-throughput analysis of images for estimating phenotypes/traits associated with tested plants | [127] |
| HTPheno | Shoot | Height, width and shoot area | Analyzes colour images of plants and different phenotypical parameters for each plant | [8] |
| GiaRoots | Root | Morpho-geometric parameters | Semi-automated software tool for high-throughput analysis of root system images | [160] |
| RootReader 3D | Roots | Root types and phenotypic root traits | Imaging and software platform for HTP of 3-D root traits during seedling development | [52] |
| PhenoPhyte | Leaf | Leaf and plant growth parameters | Tool to analyze the non-destructive imaging of plants can be used in suboptimal imaging conditions also | [161] |
| RootNav | Root | Root system architecture | Image analysis tool for semi-automated quantification of complex root system architecture in a range of plant species | [162] |
| SmartGrain | Seed | Seed structure parameters | Software for high-throughput measurement of seed shape, makes possible to distinguish between lines with small differences in seed shape | [163] |
| SmartRoot | Root | Root system architecture | Operating system-independent freeware and relies on cross-platform standards for communication with data-analysis software | [164] |
| DART | Root | Root system architecture | Uses human vision tracing to avoid analytical biases | [165] |
| Tomato analyzer | Fruit | Fruit colour | Analyzes tomato fruit colour | [112] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pratap, A.; Gupta, S.; Nair, R.M.; Gupta, S.K.; Schafleitner, R.; Basu, P.S.; Singh, C.M.; Prajapati, U.; Gupta, A.K.; Nayyar, H.; et al. Using Plant Phenomics to Exploit the Gains of Genomics. Agronomy 2019, 9, 126. https://doi.org/10.3390/agronomy9030126
Pratap A, Gupta S, Nair RM, Gupta SK, Schafleitner R, Basu PS, Singh CM, Prajapati U, Gupta AK, Nayyar H, et al. Using Plant Phenomics to Exploit the Gains of Genomics. Agronomy. 2019; 9(3):126. https://doi.org/10.3390/agronomy9030126
Chicago/Turabian StylePratap, Aditya, Sanjeev Gupta, Ramakrishnan Madhavan Nair, S. K. Gupta, Roland Schafleitner, P. S. Basu, Chandra Mohan Singh, Umashanker Prajapati, Ajeet Kumar Gupta, Harsh Nayyar, and et al. 2019. "Using Plant Phenomics to Exploit the Gains of Genomics" Agronomy 9, no. 3: 126. https://doi.org/10.3390/agronomy9030126
APA StylePratap, A., Gupta, S., Nair, R. M., Gupta, S. K., Schafleitner, R., Basu, P. S., Singh, C. M., Prajapati, U., Gupta, A. K., Nayyar, H., Mishra, A. K., & Baek, K.-H. (2019). Using Plant Phenomics to Exploit the Gains of Genomics. Agronomy, 9(3), 126. https://doi.org/10.3390/agronomy9030126