Marker Assisted Transfer of Stripe Rust and Stem Rust Resistance Genes into Four Wheat Cultivars

 ,
,
Abstract
:1. Introduction
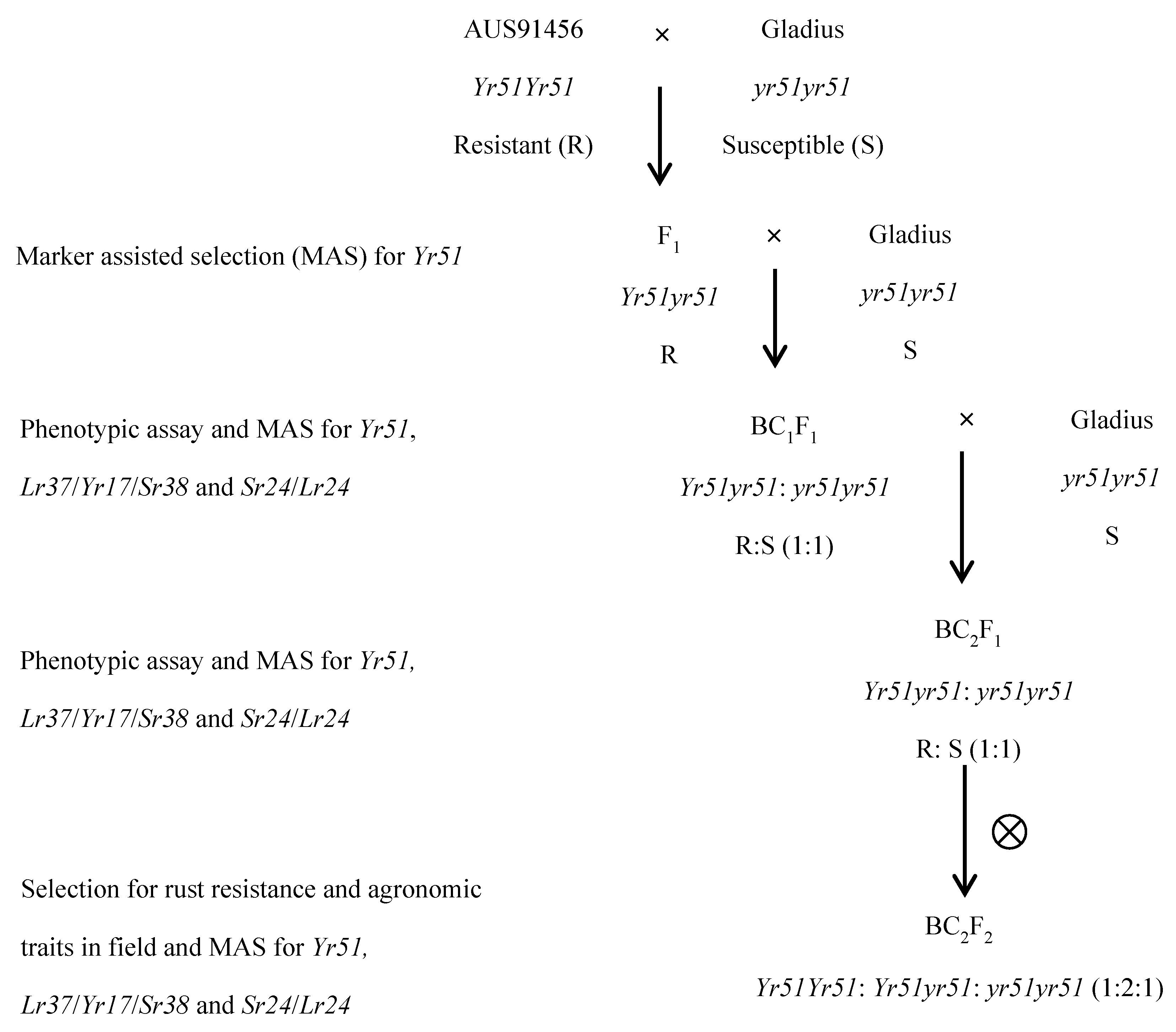
2. Materials and Methods
2.1. Plant Materials
2.2. Pathogen Materials
2.3. Greenhouse Tests
2.4. Field Tests
2.5. DNA Isolation and Quantification
2.6. PCR Amplification and Gel Electrophoresis
2.7. Marker Assisted Selection
2.8. Chi-Squared Analyses
3. Results
3.1. Marker Assisted Transfer
3.2. Yr51
3.3. Yr57
3.4. Sr22
3.5. Sr26
3.6. Sr50
3.7. Marker Assisted Selection of Gene Combinations Among BCF2 Families
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watson, I.A.; Butler, F.C. Wheat Rust Control in Australia. In Proceedings of the National Conferences and Other Initiatives and Developments, Sydney, Australia; 1984; p. 88. [Google Scholar]
- Hills, A.; Dickie, R.; Loughman, R. Wheat rust management begins now. Aust. Grain 1999, 9, 3–5. [Google Scholar]
- Wellings, C.R.; Wright, D.G.; Keiper, F.; Loughman, R. First detection of wheat stripe rust in Western Australia: Evidence for a foreign incursion. Aust. Plant Pathol. 2003, 32, 321–322. [Google Scholar] [CrossRef]
- Wellings, C.R. Puccinia striiformis in Australia: A review of the incursion, evolution, and adaptation of stripe rust in the period 1979-2006. Aust. J. Agric. Res. 2007, 58, 567–575. [Google Scholar] [CrossRef]
- Biffen, R.H. Mendel’s laws of inheritance and wheat breeding. J. Agric. Sci. 1905, 1, 4–48. [Google Scholar] [CrossRef]
- Brennan, J.P.; Murray, G.M. Australian wheat diseases—Assessing their economic importance. Agric. Sci. 1988, 1, 26–35. [Google Scholar]
- Watson, I.W.; Singh, D. The future of rust resistant wheat in Australia. J. Aust. Inst. Agric. Sci. 1952, 18, 190–197. [Google Scholar]
- Paterson, A.H.; Tanksley, S.D.; Sorrells, M.E. DNA markers in plant improvement. Adv. Agron. 1991, 46, 39–90. [Google Scholar]
- Gupta, P.K.; Varshney, R.K.; Sharma, P.C.; Ramesh, B. Molecular markers and their applications in wheat breeding. Plant Breed. 1999, 118, 369–390. [Google Scholar] [CrossRef]
- Bariana, H.S.; Bansal, U.K.; Basandrai, D.; Chhetri, M. Application of genomics to breed disease-resistant crop varieties. In Genomics and Breeding for Climate-Resilient Crops; Kole, C., Ed.; Springer: Berlin, Heidelberg, Germany, 2013; Volume 2, pp. 291–314. [Google Scholar]
- Randhawa, M.; Bansal, U.; Valárik, M.; Klocova, B.; Dolezel, J.; Bariana, H. Molecular mapping of stripe rust resistance gene Yr51 in chromosome 4AL of wheat. Theor. Appl. Genet. 2014, 127, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, M.; Bansal, U.; Mago, R.; Bariana, H. Mapping of a new stripe rust resistance locus Yr57 on chromosome 3BS of wheat. Mol. Breed. 2015, 35, 1–8. [Google Scholar] [CrossRef]
- Periyannan, S.K.; Bansal, U.K.; Bariana, H.S.; Pumphrey, M.; Lagudah, E.S. A robust molecular marker for the detection of shortened introgressed segment carrying the stem rust resistance gene Sr22 in common wheat. Theor. Appl. Genet. 2011, 122, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mago, R.; Bariana, H.S.; Dundas, I.S.; Spielmeyer, W.; Lawrence, G.J.; Pryor, A.J.; Ellis, G. Development of PCR markers for the selection of wheat stem rust resistance genes Sr24 and Sr26 in diverse wheat germplasm. Theor. Appl. Genet. 2005, 111, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Mago, R.; Zhang, P.; Vautrin, S.; Šimková, H.; Bansal, U.; Luo, M.C.; Rouse, M.; Karaoglu, H.; Periyannan, S.; Kolmer, J.; et al. The wheat Sr50 gene reveals rich diversity at a cereal disease resistance locus. Nature Plants 2015, 1, 15186. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.A.; Singh, P.K.; Singh, S.; Velu, G. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Ann. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Bariana, H.S.; McIntosh, R.A. Cytogenetic studies in wheat. XV. Location of rust resistance genes in VPM1 and their genetic linkage with other disease resistance genes in chromosome 2A. Genome 1993, 36, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Bansal, U.K.; Hayden, M.J.; Gill, M.B.; Bariana, H.S. Chromosomal location of an uncharacterised stripe rust resistance gene in wheat. Euphytica 2010, 171, 121–127. [Google Scholar] [CrossRef]
- Lagudah, E.S.; McFadden, H.; Singh, R.P.; Huerta-Espino, J.; Bariana, H.S.; Spielmeyer, W. Molecular genetic characterization of the Lr34/Yr18 slow rusting resistance gene region in wheat. Theor. Appl. Genet. 2006, 114, 21–30. [Google Scholar] [CrossRef]
- Helguera, M.; Khan, I.A.; Kolmer, J.; Lijavetzky, D.; Zhong-qi, L.; Dubcovsky, J. PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop Sci. 2003, 43, 1839–1847. [Google Scholar] [CrossRef]
- Mago, R.; Brown-Guedira, G.; Dreisigacker, S.; Breen, J.; Jin, Y.; Singh, R.; Appels, R.; Lagudah, E.S.; Ellis, J.; Spielmeyer, W. An accurate DNA marker assay for stem rust resistance gene Sr2 in Wheat. Theor. Appl. Genet. 2011, 122, 735–744. [Google Scholar] [CrossRef]
- Bariana, H.S.; McIntosh, R.A. Genetics of adult plant stripe rust resistance in four Australian wheat cultivars and French cultivar Hybride-de-Bersee. Plant Breed. 1995, 114, 485–491. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Rajaram, S. Achieving near immunity to leaf and stripe rusts in wheat by combining slow rusting resistance genes. Acta Phytopathol. Entomol. Hung. 2000, 35, 135–139. [Google Scholar]
- Singh, R.P.; Huerta-Espino, J. The use of ‘single backcross, selected bulk’ breeding approach for transferring minor genes based rust resistance into adapted cultivars. In Proceedings of the 54th Australian Cereal Chemistry Conference and 11th Wheat Breeders Assembly, Canberra, Australia, 21–24 September 2004; Black, C.K., Panozzo, J.F., Rebetzke, G.J., Eds.; Cereal Chemistry Division, Royal Australian Chemical Institute: Melbourne, Australia, 2004; pp. 48–51. [Google Scholar]
- Kuchel, H.; Fox, R.; Reinheimer, J.; Mosioek, L.; Willey, N.; Bariana, H.; Jefferies, S. The successful application of marker-assisted wheat breeding strategy. Mol. Breed. 2007, 20, 295–308. [Google Scholar] [CrossRef]
- Koebner, R.M.D.; Summers, W. 21st century wheat breeding: Plot selection or plate detection? Trends Biotechnol. 2003, 21, 59–63. [Google Scholar] [CrossRef]
- Bonnet, D.; Hyles, J.; Rebetzke, G. Efficient integration of molecular and conventional breeding methodologies. In Wheat Production in Stressed Environments; Buck, H.T., Nisi, J.E., Salomon, N., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 12, pp. 746–753. [Google Scholar]
- Bansal, U.; Bariana, H. Advances in identification and mapping of rust resistance genes in wheat. Methods Mol. Biol. 2017, 1659, 151–162. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Recurrent Parents | Donor Parents | Target Gene | Backcross Progenies |
|---|---|---|---|
| Gladius, Livingston, PBW550, DBW17 | AUS91456 | Yr51 | BC2F2 |
| AUS91463 | Yr57 | BC2F2 | |
| Sr22/3*K441 | Sr22 | BC1F2 | |
| Sr26 WA1 | Sr26 | BC1F2 | |
| Dra-1/Chinese Spring ph1b/2/3* Gabo | Sr50 | BC1F2 |
| Gene | Chromosome | Marker | Dominant/Co-Dominant | Base Pairs/Allele | Source | |
|---|---|---|---|---|---|---|
| Donor Parent | Recurrent Parent | |||||
| Lr34 | 7DS | csLV34 | Co-dominant | 150 | 229 | Lagudah et al. [19] |
| Lr37 | 2AS | VENTRIUP-LN2 | Dominant | 252 | Null | Helguera et al. [20] |
| Yr51 | 4AL | sun104 | Dominant | 225 | Null | Randhawa et al. [11] |
| Yr57 | 3BS | gwm389 | Co-dominant | 150 | 145 or 147 | Randhawa et al. [12] |
| Yr57 | 3BS | BS00062676 | Co-dominant | A:A | G:G | Randhawa et al. [12] |
| Sr2 | 3BS | csSr2 | Co-dominant | 172, 112, 53 | 225,112 | Mago et al. [21] |
| Sr22 | 7AL | cssu22 | Co-dominant | 237 | 335 | Periyannan et al. [13] |
| Sr24 | 3DL | Sr24#12 | Dominant | 500 | Null | Mago et al. [14] |
| Sr26 | 6AL | Sr26#43 | Dominant | 270 | Null | Mago et al. [14] |
| Sr50 | 1DL.1RS | Sr50-5p-F3, R2 | Dominant | 470 | Null | Mago et al. [15] |
| Genotype | Pedigree | Infection type | |
|---|---|---|---|
| 134 E 16A+Yr17+Yr27+ | 34-1,2,3,4,5,6,7 | ||
| Donor parent | |||
| AUS91456 | AUS27858/Westonia RIL #5515-1 | ;n-1n | - |
| AUS91463 | AUS27858/Westonia RIL #5474-6 | 0; | - |
| Sr22/3*K441 | - | - | ;1+ |
| Sr26 WA1 | - | - | ;1+c |
| Dra-1/Chinese Spring ph1b/2/3* Gabo | - | - | ;1++c |
| Recurrent parent | |||
| Gladius | (DH)Rac-875/Krichauff//Excalibur /Kukri/3/RAC-875/Krichauff/4/RAC-875//Excalibur/Kukri | 3+ | ;1 |
| Livingston | SUN-129-A/Sunvale | 3+ | 23c |
| PBW550 | WH-594/RAJ-3856//W-485 | 3+ | ;1− |
| DBW17 | CMH-79-A-95/3*Ciano79//RAJ-3777 | 3+ | ;1 |
| BC1F1 | BC2F1 | |||||||
| Cross | Yr51yr51 | yr51yr51 | χ2 (1:1) | p-value | Yr51yr51 | yr51yr51 | χ2 (1:1) | p-value |
| Yr51/Gladius | 11 | 8 | 0.47 | 0.49 | 22 | 14 | 1.78 | 0.18 |
| Yr51/Livingston | 9 | 8 | 0.06 | 0.81 | 14 | 11 | 0.36 | 0.55 |
| Yr51/PBW550 | 11 | 7 | 0.89 | 0.35 | 9 | 4 | 1.92 | 0.16 |
| Yr51/DBW17 | 11 | 9 | 0.20 | 0.65 | 6 | 14 | 3.20 | 0.07 |
| Yr57yr57 | yr57yr57 | Yr57yr57 | yr57yr57 | |||||
| Yr57/Gladius | 10 | 10 | 0.00 | 1.00 | 11 | 11 | 0.00 | 1.00 |
| Yr57/Livingston | 14 | 6 | 3.20 | 0.07 | 14 | 6 | 3.20 | 0.07 |
| Yr57/PBW550 | 6 | 5 | 0.09 | 0.76 | 8 | 9 | 0.06 | 0.80 |
| Yr57/DBW17 | 10 | 9 | 0.05 | 0.82 | 7 | 4 | 0.82 | 0.37 |
| BC1F1 | BC1F2 | |||||||
| Sr22sr22 | sr22sr22 | Sr22sr22 | sr22sr22 | χ2 (3:1) | ||||
| Sr22/Gladius | 4 | 6 | 0.40 | 0.53 | 9 | 7 | 3.00 | 0.08 |
| Sr22/Livingston | 5 | 4 | 0.11 | 0.74 | 13 | 3 | 0.33 | 0.56 |
| Sr22/PBW550 | 3 | 5 | 0.50 | 0.48 | 10 | 6 | 1.33 | 0.25 |
| Sr22/DBW17 | 7 | 5 | 0.33 | 0.56 | 13 | 3 | 0.33 | 0.56 |
| Sr26sr26 | Sr26sr26 | Sr26sr26 | sr26sr26 | |||||
| Sr26/Gladius | 5 | 5 | 0.00 | 1.00 | 13 | 3 | 0.33 | 0.56 |
| Sr26/Livingston | 5 | 5 | 0.00 | 1.00 | 10 | 6 | 1.33 | 0.25 |
| Sr26/PBW550 | 4 | 5 | 0.11 | 0.74 | 14 | 2 | 1.33 | 0.25 |
| Sr26/DBW17 | 3 | 9 | 3.00 | 0.08 | 12 | 4 | 0.00 | 1.00 |
| Sr50sr50 | sr50sr50 | Sr50sr50 | sr50sr50 | |||||
| Sr50/Gladius | 7 | 3 | 1.60 | 0.20 | 9 | 7 | 3.00 | 0.08 |
| Sr50/Livingston | 4 | 6 | 0.40 | 0.53 | 10 | 6 | 1.33 | 0.25 |
| Sr50/PBW550 | 4 | 6 | 0.40 | 0.53 | 11 | 5 | 0.33 | 0.56 |
| Sr50/DBW17 | 6 | 6 | 0.00 | 1.00 | 15 | 1 | 3.00 | 0.08 |
| Recurrent Parents | Background Gene (s) | Gene Combinations | Number of Selected Plants |
|---|---|---|---|
| Gladius | Lr37/Yr17/Sr38, Lr24/Sr24 | Yr51 + Lr37/Yr17/Sr38+ Lr24/Sr24 | 4 |
| Yr57 + Lr37/Yr17/Sr38 + Lr24/Sr24 | 7 | ||
| Sr22 + Lr37/Yr17/Sr38 + Lr24/Sr24 | 7 | ||
| Sr26 + Lr37/Yr17/Sr38 + Lr24/Sr24 | 13 | ||
| Sr50 + Lr37/Yr17/Sr38 + Lr24/Sr24 | 8 | ||
| Livingston | Lr34/Yr18/Sr57, Lr37/Yr17/Sr38, Sr2 | Yr51 + Lr34/Yr18/Sr57 + Lr37/Yr17/Sr38 | 4 |
| Yr57 + Lr34/Yr18/Sr57 + Lr37/Yr17/Sr38 | 11 | ||
| Sr22 + Lr34/Yr18/Sr57+ Lr37/Yr17/Sr38 | 12 | ||
| Sr26 + Lr34/Yr18/Sr57 + Lr37/Yr17/Sr38 | 2 | ||
| Sr50 + Lr34/Yr18/Sr57 + Lr37/Yr17/Sr38 | 4 | ||
| PBW550 | Lr34/Yr18/Sr57 | Yr51 + Lr34/Yr18/Sr57 | 3 |
| Yr57 + Lr34/Yr18/Sr57 | 3 | ||
| Sr22 + Lr34/Yr18/Sr57 | 10 | ||
| Sr26 + Lr34/Yr18/Sr57 | 4 | ||
| Sr50 + Lr34/Yr18/Sr57 | 4 | ||
| DBW17 | Lr34/Yr18/Sr57 | Yr51 + Lr34/Yr18/Sr57 | 3 |
| Yr57 + Lr34/Yr18/Sr57 | 2 | ||
| Sr22 + Lr34/Yr18/Sr57 | 9 | ||
| Sr26 + Lr34/Yr18/Sr57 | 3 | ||
| Sr50 + Lr34/Yr18/Sr57 | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randhawa, M.S.; Bains, N.S.; Sohu, V.S.; Chhuneja, P.; Trethowan, R.M.; Bariana, H.S.; Bansal, U. Marker Assisted Transfer of Stripe Rust and Stem Rust Resistance Genes into Four Wheat Cultivars. Agronomy 2019, 9, 497. https://doi.org/10.3390/agronomy9090497
Randhawa MS, Bains NS, Sohu VS, Chhuneja P, Trethowan RM, Bariana HS, Bansal U. Marker Assisted Transfer of Stripe Rust and Stem Rust Resistance Genes into Four Wheat Cultivars. Agronomy. 2019; 9(9):497. https://doi.org/10.3390/agronomy9090497
Chicago/Turabian StyleRandhawa, Mandeep S., Navtej S. Bains, Virinder S. Sohu, Parveen Chhuneja, Richard M. Trethowan, Harbans S. Bariana, and Urmil Bansal. 2019. "Marker Assisted Transfer of Stripe Rust and Stem Rust Resistance Genes into Four Wheat Cultivars" Agronomy 9, no. 9: 497. https://doi.org/10.3390/agronomy9090497