Genetic Association between Amyotrophic Lateral Sclerosis and Cancer



1
Department of Physics, Chuo University, 1-13-27 Kasuga, Bunky-ku, Tokyo 112-8551, Japan
2
Institute of Statistics, National Chiao Tung University, Hsinchu 30010, Taiwan
*
Author to whom correspondence should be addressed.
Genes 2017, 8(10), 243; https://doi.org/10.3390/genes8100243
Submission received: 12 August 2017
/
Revised: 15 September 2017
/
Accepted: 22 September 2017
/
Published: 27 September 2017
(This article belongs to the Special Issue Statistical Resources for the Interpretation and Integration of Human Genetic Association Studies)
Abstract
:Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease. An ALS drug, Riluzole, has been shown to induce two different anticancer effects on hepatocellular carcinoma (HCC). In light of this finding, we explore the relationship between ALS and cancer, especially for HCC, from the molecular biological viewpoint. We establish biomarkers that can discriminate between ALS patients and healthy controls. A principal component analysis (PCA) based unsupervised feature extraction (FE) is used to find gene biomarkers of ALS based on microarray gene expression data. Based on this method, 101 probes were selected as biomarkers for ALS with 95% high accuracy to discriminate between ALS patients and controls. Most of the genes corresponding to these probes are shown to be related to various cancers. These findings might provide a new insight for developing new therapeutic options or drugs for both ALS and cancer.
1. Introduction
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease, which is caused by degeneration of upper motor neurons of the motor cortex and corticospinal tract and lower motor neurons of the brainstem and spinal cord. Eventually, it can affect the ability to control the muscles for movement, leading to death. Despite many studies to investigate this disease, no effective therapeutics have been identified to prevent or stop neuronal death in patients so far. Most of the cases are caused by unknown reasons. Only 5–10% of ALS cases have a family history of the disease [1]. Familial cases have been linked to mutations in a number of genes, including Cu/Zn superoxide dismutase 1 (SOD1), TAR DNA binding protein (TARDBP), fused in sarcoma (FUS), and chromosome 9 open reading frame 72 repeat expansions (C9ORF72) [2,3].
The comorbidity of ALS with other disorders has been discussed in previous cohort studies. Increasing evidence suggests that ALS is inversely related to cancer [4]. An increased risk of ALS was observed during the first year after cancer diagnosis, and in contrast, a lower risk of cancer was observed in ALS patients after diagnosis compared with ALS-free individuals [5]. Most discussions of the comorbidity of ALS with cancer are based on cohort studies, case reports of individual observations, or sequence mutation analysis. Genetic speculation through the microarray data with respect to the relationship between ALS and cancer has not been much explored in the literature. In this study, we aim to explore the potential genetic link between ALS and cancer, especially for hepatocellular carcinoma (HCC) and pancreatic adenocarcinoma (PAAD), which are primary tumors of the liver and pancreas.
The clinical evidence of the association between ALS and cancer in the literature includes the effect of ALS drugs in cancer therapy and the association between paraneoplastic antibodies and ALS. An ALS drug, Riluzole, was shown to induce two different anti-cancer effects on HCC [5]. A male patient was reported with C9ORF72 familial ALS and paraneoplastic antibodies (anti-Ma2/Ta) [6]. Manganese superoxide dismutase (MnSOD) can be induced to protect against pro-oxidant insults resulting from certain tumors and ALS [7]. Reports have shown increased MnSOD protein or activity in ALS patient samples [8,9]. However, the role of MnSOD in cancer is still disputable as some studies have reported that tumor cells/tissues contain decreased MnSOD activity whereas some describe an increase of MnSOD protein in tumor cells/tissues [10,11,12].
To find the biological significance of the association between ALS and cancer, we analyze microarray gene expression data for ALS and healthy control samples. All samples were from human biopsies, either from healthy muscles or from the muscle of patients who were clearly diagnosed as having ALS [13]. The findings in this study enhance the biological evidence connecting ALS with cancer and might provide a new insight for developing new therapeutic options for both diseases, and a new perspective for understanding the pathophysiology of ALS.
2. Materials and Methods
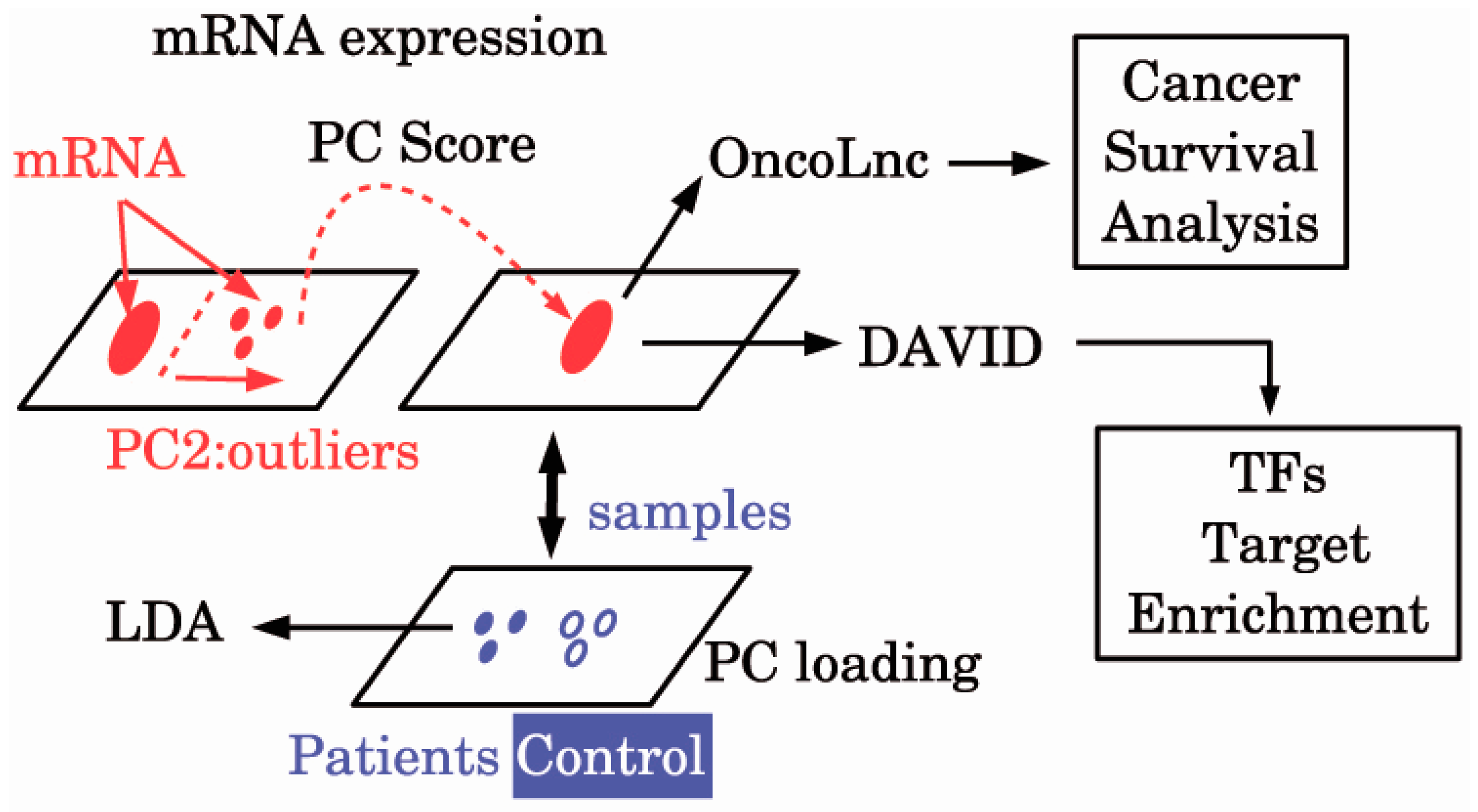
In order to clarify the relationship between ALS and cancers from the molecular biological viewpoint, we first establish gene biomarkers that can discriminate between ALS patients and healthy controls, and then we investigate whether these biomarkers are functional/dysfunctional in a cancer-specific manner. For this purpose, we applied the principal component analysis (PCA) based unsupervised feature extraction (FE) [14,15] to gene microarray expression profiles E-MEXP-3260 [13] to discover potential ALS probes.
Gene expression profiles used in this study were downloaded from ArrayExpress using the accession number E-MEXP-3260. It includes 10 normal samples (01a to 10a) and nine ALS samples (11a to 19a). The 10 normal samples include five females and five males; the nine ALS samples are all males. The ALS diagnosis of the nine patients with probable or definite ALS is based on the revised El Escorial criteria [16] by performing an open biopsy in the middle portion of deltoid muscle on these patients [13]. All patients had sporadic ALS, and presented with symptoms of limb onset. All of them underwent a complete needle electromyography investigation. The normal control samples were obtained from 10 subjects without any significant neurological history, when undergoing a shoulder orthopedic surgery. The samples were processed and stored using standard procedures [17]. The gender and age of the samples can be found in Table 1 of [13]. The average age of the 10 normal samples and the nine ALS samples are 52.1 ± 7.8 and 48.5 ± 15.9 years, respectively. The time from onset of symptoms to biopsy, the revised ALS functional rating scale and other information of the nine patients can be obtained from that table.
To analyze the data, the obtained 19 CHP format files, which contains probe set analysis results generated from Affymetrix software, were loaded into R (R Core Team, 2015) using the ‘readChp’ function in the package ‘affxparser’. Then, the component ‘QuantificationEntries$Signal’ of the output of ‘readChp’ was used for further analyses. Gene expression profiles were normalized with each sample (i.e., each column was normalized).
Suppose is the gene expression of the th gene in the th sample. In PCA based unsupervised FE, not samples but genes are embedded into low dimensional space. Thus, principal component (PC) scores attributed to genes are computed as a component of eigenvector, , of , where is the matrix of and is its transposed matrix
where is the eigenvalues (). Principal component loading attributed to samples are the components of the vector that is the eigenvector of because
Since we could identify that the second PC loading is distinct between healthy controls and ALS patients with a p-value 9.4 × 10−7 by the Student’s t-test, we decided to select genes with the second PC score. p-Values were attributed to genes with assuming that the test statistic follows a distribution where is the th component of and is the standard deviation of . p-Values were further adjusted by Benjamini and Hochberg criterion [18] and genes associated with adjusted p-values less than 0.01 were selected.
In order to examine whether the selected probes can discriminate between ALS patients and healthy controls, PC loading was recomputed using only the selected probes and was attributed to 19 samples. Then samples were discriminated using linear discriminate analysis (LDA) with the second PC loading. Leave one out cross-validation was employed.
Then we identified the genes which are associated with the selected probes by the Database for Annotation, Visualization, and Integrated Discovery (DAVID) gene ID converter [19]. Probe ID (GENBANK accession) was converted to gene symbol. Single GENBANK accession was often converted to multiple gene symbols. The obtained genes symbols were uploaded to OncoLnc [20] one by one, and cancers associated with false discovery rate (FDR) of less than 0.05 were identified as cancers associated with significant survival probabilities for the investigated genes.
All of the statistical analyses were performed using R codes. Gene expression profiles were normalized by the function ‘scale’ and PCA was performed by the function ‘prcomp’. Student’s t-test was performed by the function ‘t.test’ and LDA was performed by the function ‘lda’ in R package ‘MASS’.
3. Results
A schematic diagram of the analyses is provided in Figure 1. Then, 101 probes in E-MEXP-3260 were selected (Supplementary Table S1). Using these 101 probes, we can discriminate between ALS patients and healthy controls with the accuracy of 95% (Table 1) by applying the LDA. Thus, we could successfully identify biomarkers that can discriminate between ALS patients and healthy controls with high accuracy.
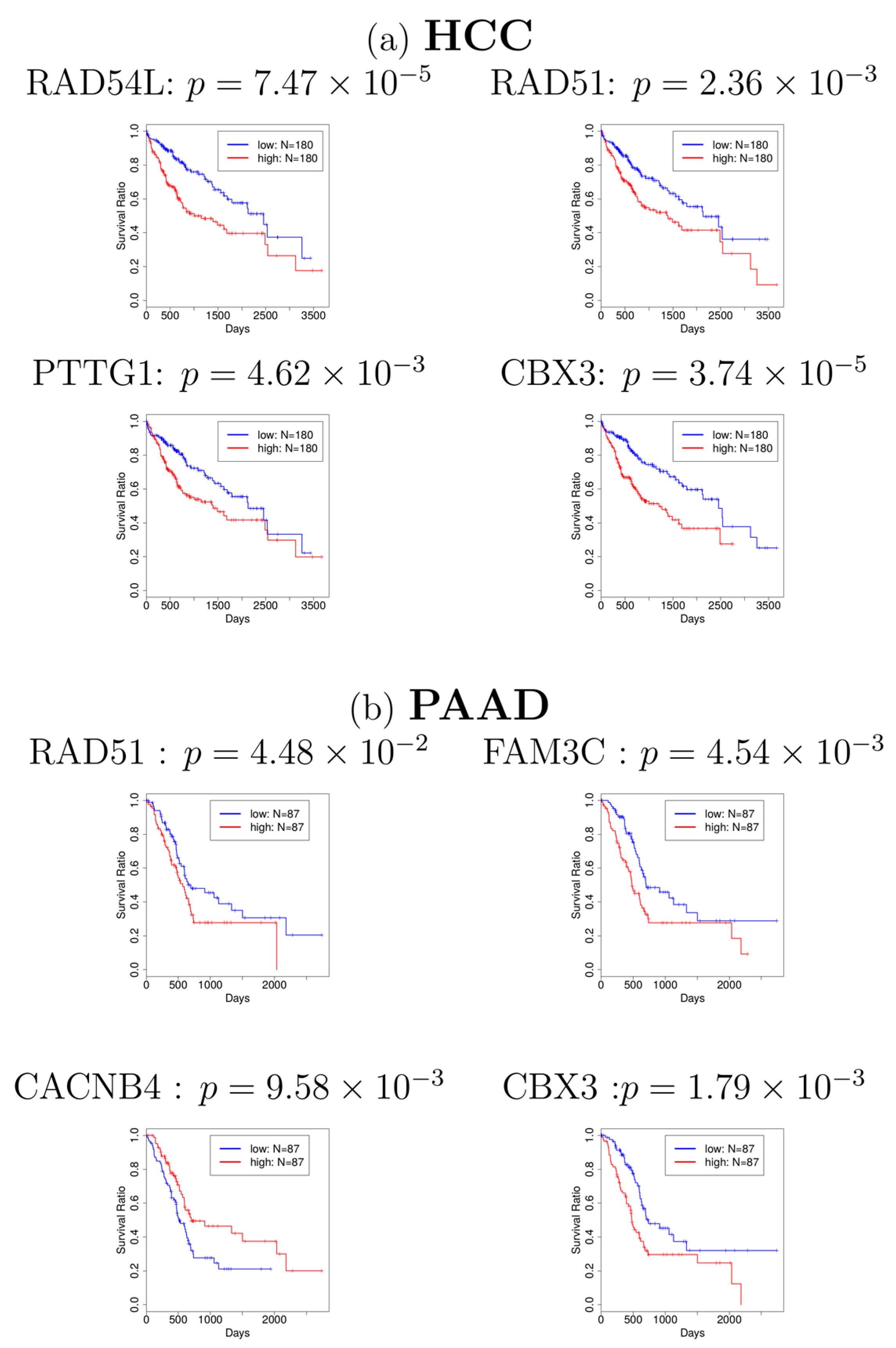
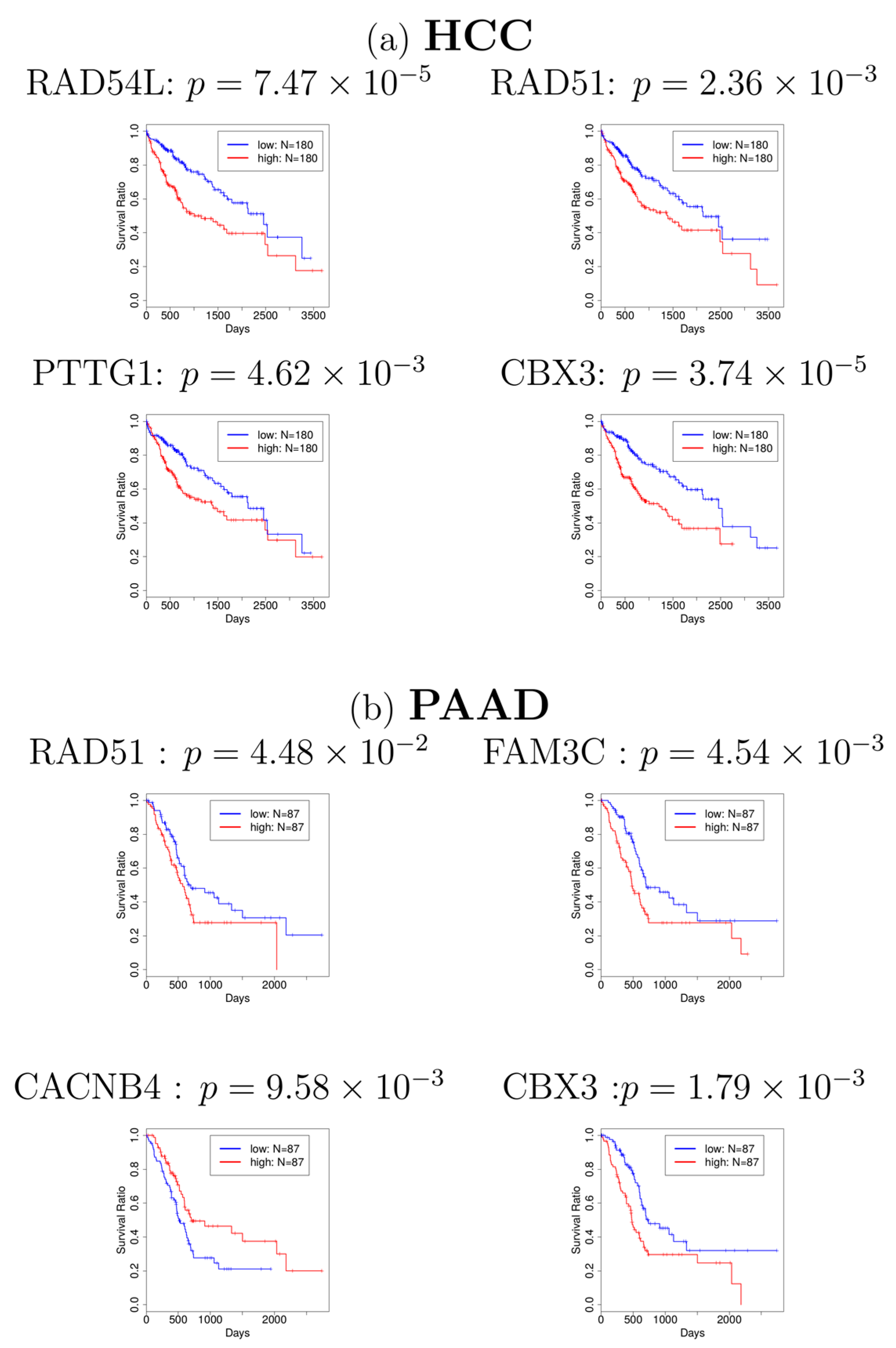
Next, in order to investigate whether the genes associated with these 101 probes have a significant relationship with cancer, we first identified the genes which are associated with 101 probes. The identified genes are shown in Supplementary Table S2. Many of them have been reported to be related to cancers. For example, RAD51 was reported to be related to HCC and PAAD [21,22]. CBX3, also known as HP1γ, was also reported to be related to HCC [23] and PAAD [24]. In addition to these findings from literature review, to investigate whether most of the genes in Supplementary Table S2 are related to cancers, we adopted the database OncoLnc [20] to find the association between these genes and cancers, which is a pre-computed database for multiple cancers and has been used in calculating survival probability in several studies [25,26]. The survival analyses for genes in Supplementary Table S2 are provided in Supplementary Table S3, which shows that most of the genes identified are related to various cancers. Especially, four genes (RAD54L, RAD51, PTTG1, and CBX3) and four genes (RAD51, FAM3C, CACNB4, and CBX3) are significantly related to HCC and PAAD in the survival analyses (see Figure 2), respectively, which corresponds with some of the previous studies. Finally, we selected 68 genes (Supplementary Table S4) that turn out to be significantly associated with cancer survival probability. To our knowledge, this is the first report that relates gene expression profiles of ALS to cancer survival analysis
4. Discussion
Although we could successfully identify ALS biomarkers that are related to cancer from the survival analyses, more detailed biological consideration of these biomarker genes might provide an understanding of the biological background shared between ALS and cancer. In order to clarify this point, we uploaded the 68 genes (Supplementary Table S4) to DAVID to perform pathway enrichment analysis. Multiple Reactome pathway enrichment was observed (Supplementary Table S5), which included many terms associated with cancer. For example, the terms, R-HSA-975956 and R-HSA-975957, were included in the nonsense-mediated decay (NMD) pathway, and NMD was known to be related to cancers. Wang et al. [27] reported that inhibition of NMD by the tumor microenvironment promotes tumorigenesis. Ionov et al. [28] found that manipulation of NMD identifies gene mutations in colon cancer cells with microsatellite instability. Nonsense-mediated decay was also suggested to play critical roles in implications for tumorigenesis [29]. On the other hand, NMD is known to block antisense oligonucleotides (ASOs), and blocking ASOs could reduce SOD1 in the central nervous system [30]. SOD1 is the famous ALS-causing gene. Li et al. [31] showed that endoplasmic reticulum (ER) stress compounded TAR DNA-binding protein 43 (TDP-43) depletion in the upregulation of NMD isoforms that had been implicated in the pathogenic mechanisms of ALS. TDP-43-associated cryptic exons was also reported to lead to NMD [32]. TDP-43 is the main component of ubiquitinated protein aggregates found in sporadic ALS patients, not only familial ALS [33]. In addition to this, expansion of hexanucleotide repeats in C9ORF72 increases the susceptibility for pathological alteration of TDP-34 [34]. The worldwide frequency of the C9ORF72 repeat expansion is now estimated to more than 30% for familial ALS, which represents more than 5% of the sporadic ALS cases [35]. Thus, these findings suggested that ALS is related to cancers. Second, signal recognition particle (SRP) related term R-HSA-1799339 in Supplementary Table S5, was enriched. Signal recognition particle is suggested to be related to cancer histology [36] and the SRP protein is also detected in colon cancer [37]. On the other hand, myopathy associated with anti-signal recognition particle (anti-SRP) is a severe necrotizing immune-mediated disease [38]. Anti-SRP myopathy also differs from other immune-mediated myopathies by its characteristically poor responsiveness to steroid monotherapy and conventional immunosuppressive therapies [39]. Moreover, there it has been reported that the enhancement of nerve inflammation causes nerve cell death [40]. As a result, SRP can associate ALS with cancers.
More interestingly, in some cases, NMD and SRP were detected together in cancer tissues. Simões et al. [41] had identified the enrichment of “SRP-dependent cotranslational protein targeting to membrane” and “nonsense-mediated decay” in the MCF-7 cell line by investigating an interactome of endogenous and overexpressed ER in MCF-7 cell line generated from breast cancer. Durmaz et al. [42] identified enrichment of “SRP-dependent cotranslational protein targeting to membrane” and “nonsense-mediated decay” infrequent subgraph mining of personalized signaling pathway networks analysis applied to breast cancer as well as glioblastoma multiforme patients. This strengthens the feasibility that NMD and SRP were identified together in the present research when cancer-related genes were sought among those related to ALS.
In addition to pathway enrichments, multiple transcription factor (TF) target genes were enriched (Supplementary Table S5). These TFs were P300, E47, AHR, HMX1, AREB6, GATA2, and MEIS1AHOXA9. As for the relationship between these TFs and ALS, E47, also known as TCF3, was reported as one of 23 candidate causal master regulators of neurodegeneration in an in vitro model of ALS [43]. Butovsky et al. [44] found that AHR was significantly upregulated in Ly6Chi monocytes in SOD1 mice. AREB6, also known as ZEB1, was suggested to be included in a regulatory circuit related to ALS pathogenesis [45]. P300, also known as EP300, also has a role in the etiology of ALS [46]. Thus, not all but the majority of these TFs were reported to be related to ALS. On the other hand, as for the relationship between cancers and these TFs, mutations truncating EP300 acetylase was reported in human cancers [47]. ZEB1 turns into a transcriptional activator by interacting with YAP1 in aggressive cancer types [48]. AHR is also known to be deeply related to cancer biogenesis [49]. Breast cancer growth and initiation were controlled by TCF3 [50]. GATA is also a well- known cancer-related TF [51]. Therefore, all of the discussions here suggested that genes that can distinguish between ALS and healthy controls can biologically bridge between ALS and cancer biogenesis.
5. Conclusions
In this paper, we have identified genes whose expression profiles can discriminate between ALS patients and healthy controls. Most of these gene expression profiles turned out to be related to various cancers using survival analysis, pathway enrichment analysis, and TF enrichment analysis. Furthermore, biological consideration of these genes can help us to understand the molecular biological background that ALS patients can have distinct cancer pathogenesis from other patients. Specifically, NMD and SRP, as well as enhanced TF binding protein genes, are promising candidates that relate to ALS and cancer. These findings suggested that our approach is useful for understanding the relationship between cancers and ALS, especially from a molecular biology point of view
Supplementary Materials
The following are available online at www.mdpi.com/2073-4425/8/10/243/s1. Table S1: 101 probes identified by PCA based unsupervised FE, Table S2: Gene symbols associated with 101 probes identified by DAVID gene ID converter, Table S3: Cancer survival probabilities identified by OncoLnc for genes in Table S2, Table S4: Genes associated with significant cancer survival probabilities, Table S5: REACTOME and TFBS enrichment by DAVID for genes in Table S4.
Author Contributions
Y.T. and H.W. conceived and designed the experiments; H.W. performed the experiments; Y.T. analyzed the data; Y.T. and H.W. wrote the paper.
Acknowledgments
This work has been supported by Ministry of Science and Technology 105-2118-M-009-001 -MY2, Taiwan.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Kiernan, M.C.; Vucic, S.; Cheah, B.C.; Turner, M.R.; Eisen, A.; Hardiman, O.; Burrell, J.R.; Zoing, M.C. Amyotrophic lateral sclerosis. Lancet 2011, 377, 942–955. [Google Scholar] [CrossRef]
- Westeneng, H.J.; Walhout, R.; Straathof, M.; Schmidt, R.; Hendrikse, J.; Veldink, J.H.; van den Heuvel, M.P.; van den Berg, L.H. Widespread structural brain involvement in ALS is not limited to the C9orf72 repeat expansion. J. Neurol. Neurosurg. Psychiatry 2016, 87, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Bergen, J.M.; Park, I.K.; Horner, P.J.; Pun, S.H. Nonviral approaches for neuronal delivery of nucleic acids. Pharm. Res. 2008, 25, 983–998. [Google Scholar] [CrossRef] [PubMed]
- Freedman, D.M.; Curtis, R.E.; Daugherty, S.E.; Goedert, J.J.; Kuncl, R.W.; Tucker, M.A. The association between cancer and amyotrophic lateral sclerosis. Cancer Cause Control 2013, 24, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Al-Chalabi, A.; Ronnevi, L.O.; Turner, M.R.; Wirdefeldt, K.; Kamel, F.; Ye, W. Amyotrophic lateral sclerosis and cancer: A register-based study in Sweden. Amyotroph. Lateral Scler. Front. Degener. 2013, 14, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Seol, H.S.; Lee, S.E.; Song, J.S.; Lee, H.Y.; Park, S.; Kim, I.; Singh, S.R.; Chang, S.; Jang, S.J. Glutamate release inhibitor, Riluzole, inhibited proliferation of human hepatocellular carcinoma cells by elevated ROS production. Cancer Lett. 2016, 382, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Geevasinga, N.; Burrell, J.R.; Hibbert, M.; Vucic, S.; Ng, K. C9ORF72 familial motor neuron disease—Frontotemporal dementia associated with lung adenocarcinoma and anti-Ma2/Ta antibodies: A chance association? Eur. J. Neurol. 2014, 21, e31–e33. [Google Scholar] [CrossRef] [PubMed]
- Macmillan-Crow, L.A.; Cruthirds, D.L. Invited review: Manganese superoxide dismutase in disease. Free Radic. Res. 2001, 34, 325–336. [Google Scholar] [CrossRef] [PubMed]
- McEachern, G.; Kassovska-Bratinova, S.; Raha, S.; Tarnopolsky, M.A.; Turnbull, J.; Bourgeois, J.; Robinson, B. Manganese superoxide dismutase levels are elevated in a proportion of amyotrophic lateral sclerosis patient cell lines. Biochem. Biophys. Res. Commun. 2000, 273, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Shaw, I.C.; Fitzmaurice, P.S.; Mitchell, J.D.; Lynch, P.G. Studies on cellular free radical protection mechanisms in the anterior horn from patients with amyotrophic lateral sclerosis. Neurodegeneration 1995, 4, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Oberley, L.W.; Buettner, G.R. Role of superoxide dismutase in cancer: A review. Cancer Res. 1979, 39, 1141–1149. [Google Scholar] [PubMed]
- Zhong, W.; Yan, T.; Lim, R.; Oberley, L.W. Expression of superoxide dismutases, catalase, and glutathione peroxidase in glioma cells. Free Radic. Biol. Med. 1999, 27, 1334–1345. [Google Scholar] [CrossRef]
- Pradat, P.F.; Dubourg, O.; de Tapia, M.; di Scala, F.; Dupuis, L.; Lenglet, T.; Bruneteau, G.; Salachas, F.; Lacomblez, L.; Corvol, J.C.; et al. Muscle gene expression is a marker of amyotrophic lateral sclerosis severity. Neurodegener. Dis. 2012, 9, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Kubo, S.; Tamori, A.; Itami, S.; Kawamura, E.; Iwaisako, K.; Ikeda, K.; Kawada, N.; Ochiya, T.; Taguchi, Y.H. Comprehensive analysis of transcriptome and metabolome analysis in intrahepatic cholangiocarcinoma and hepatocellular carcinoma. Sci. Rep. 2015, 5, 16294. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, Y.H. Identification of more feasible microRNA-mRNA interactions within multiple cancers using principal component analysis based unsupervised feature extraction. Int. J. Mol. Sci. 2016, 17, 696. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Miller, R.G.; Swash, M.; Munsat, T.L. El Escorial revisited: Revised criteria for the diagnosis of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2000, 1, 293–299. [Google Scholar] [CrossRef]
- Dubowitz, V.; Sewry, C.A.; Oldfors, A.; Lane, R.J.M. Muscle Biopsy: A Practical Approach; Saunders: Oxford, UK, 2013; Available online: https://www.clinicalkey.com/dura/browse/bookChapter/3-s2.0-C2009063539X (accessed on 1 April 2013).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J. OncoLnc: Linking TCGA survival data to mRNAs, miRNAs, and lncRNAs. PeerJ Comput. Sci. 2016, 2, e674. [Google Scholar] [CrossRef]
- Han, H.Y.; Bearss, D.J.; Browne, L.W.; Calaluce, R.; Nagle, R.B.; Von Hoff, D.D. Identification of differentially expressed genes in pancreatic cancer cells using cDNA microarray. Cancer Res. 2002, 62, 2890–2896. [Google Scholar] [PubMed]
- Maacke, H.; Jost, K.; Opitz, S.; Miska, S.; Yuan, Y.; Hasselbach, L.; Lüttges, J.; Kalthoff, H.; Stürzbecher, H.W. DNA repair and recombination factor Rad51 is over-expressed in human pancreatic adenocarcinoma. Oncogene 2000, 19, 2791–2795. [Google Scholar] [CrossRef] [PubMed]
- Dighe, N.; Khoury, M.; Mattar, C.; Chong, M.; Choolani, M.; Chen, J.; Antoniou, M.N.; Chan, J.K. Long-term reproducible expression in human fetal liver hematopoietic stem cells with a UCOE-based lentiviral vector. PLoS ONE. 2014, 9, e104805. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, S.; Glesel, E.; Singh, G.; Chen, N.M.; Reutlinger, K.; Zhang, J.; Billadeau, D.D.; Fernandez–Zapico, M.E. Restricted heterochromatin formation Links NFATc2 repressor activity with growth promotion in pancreatic cancer. Gastroenterology 2012, 142, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Leucci, E.; Coe, E.A.; Marine, J.C.; Vance, K.W. The emerging role of long non-coding RNAs in cutaneous melanoma. Pigment Cell Melanoma Res. 2016, 29, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Tong, G.; Zhang, Y.; Liang, S.; Tang, K.; Yang, Q. PGK1 drives hepatocellular carcinoma metastasis by enhancing metabolic process. Int. J. Mol. Sci. 2017, 18, 1630. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zavadil, J.; Martin, L.; Parisi, F.; Friedman, E.; Levy, D.; Harding, H.; Ron, D.; Gardner, L.B. Inhibition of nonsense-mediated RNA decay by the tumor microenvironment promotes tumorigenesis. Mol. Cell. Biol. 2011, 31, 3670–3680. [Google Scholar] [CrossRef] [PubMed]
- Ionov, Y.; Nowak, N.; Perucho, M.; Markowitz, S.; Cowell, J.K. Manipulation of nonsense mediated decay identifies gene mutations in colon cancer cells with microsatellite instability. Oncogene 2004, 23, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Gardner, L.B. Nonsense-mediated RNA decay regulation by cellular stress: Implications for tumorigenesis. Mol. Cancer Res. 2010, 8, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.J.; Norrbom, M.; Chun, S.; Bennett, C.F.; Rigo, F. Nonsense-mediated decay as a terminating mechanism for antisense oligonucleotides. Nucleic Acids Res. 2014, 42, 5871–5879. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Vuong, J.K.; Zhang, M.; Stork, C.; Zheng, S. Inhibition of nonsense-mediated RNA decay by ER stress. RNA 2016, 23, 378–394. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.P.; Pletnikova, O.; Troncoso, J.C.; Wong, P.C. TDP-43 repression of nonconserved cryptic exons is compromised in ALS-FTD. Science 2015, 349, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Farrawell, N.E.; Lambert-Smith, I.A.; Warraich, S.T.; Blair, I.P.; Saunders, D.N.; Hatters, D.M.; Yerbury, J.J. Distinct partitioning of ALS associated TDP-43, FUS and SOD1 mutants into cellular inclusions. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Onodera, O.; Sugai, A.; Konno, T.; Tada, M.; Koyama, A.; Nishizawa, M. What is the key player in TDP-43 pathology in ALS: Disappearance from the nucleus or inclusion formation in the cytoplasm? Neurol. Clin. Neurosci. 2013, 1, 11–17. [Google Scholar] [CrossRef]
- Van Blitterswijk, M.; DeJesus-Hernandez, M.; Rademakers, R. How do C9ORF72 repeat expansions cause amyotrophic lateral sclerosis and frontotemporal dementia: Can we learn from other noncoding repeat expansion disorders? Curr. Opin. Neurol. 2012, 25, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Apiwattanakul, M.; Milone, M.; Pittock, S.J.; Kryzer, T.J.; Fryer, J.P.; O’Toole, O.; Mckeon, A.; Lennon, V.A. Signal recognition particle immunoglobulin g detected incidentally associates with autoimmune myopathy. Muscle Nerve 2016, 53, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.H.; Qin, S.; Wang, J.Y.; Roehrl, M.H. Proteomic expression analysis of surgical human colorectal cancer tissues: Up-regulation of PSB7, PRDX1, and SRP9 and hypoxic adaptation in cancer. J. Proteome Res. 2008, 7, 2959–2972. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, J.; Harty, L.; Mayer, N.; Critcher, V.; Ryan, J. Immune-mediated necrotizing myopathy, associated with antibodies to signal recognition particle, together with lupus nephritis: Case presentation and management. J. Clin. Med. Res. 2015, 7, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Nishikawa, A.; Kuwana, M.; Nishimura, H.; Watanabe, Y.; Nakahara, J.; Hayashi, Y.K.; Suzuki, N.; Nishino, I. Inflammatory myopathy with anti-signal recognition particle antibodies: Case series of 100 patients. Orphanet J. Rare Dis. 2015, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Rumbaugh, J.A.; Nath, A. Neuronal Cell Death and Inflammation. In Encyclopedia of Neuroscience; Springer: Berlin/Heidelberg, Germany, 2009; pp. 2772–2776. [Google Scholar]
- Simoes, J.; Amado, F.M.; Vitorino, R.; Helguero, L.A. A meta-analysis to evaluate the cellular processes regulated by the interactome of endogenous and over-expressed estrogen receptor alpha. Oncoscience 2015, 2, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Durmaz, A.; Henderson, T.A.; Brubaker, D.; Bebek, G. Frequent subgraph mining of personalized signaling pathway networks groups patients with frequently dysregulated disease pathways and predicts prognosis. In Proceedings of the Pacific Symposium on Biocomputing Pacific Symposium on Biocomputing, Kohala Coast, HI, USA, 4–8 January 2016; Volume 22, pp. 402–413. [Google Scholar]
- Ikiz, B.; Alvarez, M.J.; Re, D.B.; Le Verche, V.; Politi, K.; Lotti, F.; Phani, S.; Pradhan, R.; Yu, C.; Croft, G.F.; et al. The regulatory machinery of neurodegeneration in in vitro models of amyotrophic lateral sclerosis. Cell Rep. 2015, 12, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Siddiqui, S.; Gabriely, G.; Lanser, A.J.; Dake, B.; Murugaiyan, G.; Doykan, C.E.; Wu, P.M.; Gali, R.R.; Iyer, L.K.; et al. Modulating inflammatory monocytes with a unique microRNA gene signature ameliorates murine ALS. J. Clin. Investig. 2012, 122, 3063–3087. [Google Scholar] [CrossRef] [PubMed]
- Dini Modigliani, S.; Morlando, M.; Errichelli, L.; Sabatelli, M.; Bozzoni, I. An ALS-associated mutation in the FUS 3’-UTR disrupts a microRNA-FUS regulatory circuitry. Nat. Commun. 2014, 5, 4335. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Filippakopoulos, P.; Knapp, S. Bromodomains as therapeutic targets. Exp. Rev. Mol. Med. 2011, 13, e29. [Google Scholar] [CrossRef] [PubMed]
- Gayther, S.A.; Batley, S.J.; Linger, L.; Bannister, A.; Thorpe, K.; Chin, S.F.; Daigo, Y.; Russell, P.; Wilson, A.; Sowter, H.M.; et al. Mutations truncating the EP300 acetylase in human cancers. Nat. Genet. 2000, 24, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, W.; Mossmann, D.; Kleemann, J.; Mock, K.; Meisinger, C.; Brummer, T.; Herr, R.; Brabletz, S.; Stemmler, M.P.; Brabletz, T. ZEB1 turns into a transcriptional activator by interacting with YAP1 in aggressive cancer types. Nat. Commun. 2016, 7, 10498. [Google Scholar] [CrossRef] [PubMed]
- Murray, I.A.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat. Rev. Cancer 2014, 14, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Slyper, M.; Shahar, A.; Bar-Ziv, A.; Granit, R.Z.; Hamburger, T.; Maly, B.; Peretz, T.; Ben-Porath, I. Control of breast cancer growth and initiation by the stem cell-associated transcription factor TCF3. Cancer Res. 2012, 72, 5613–5624. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Blobel, G.A. GATA Transcription Factors and Cancer. Genes Cancer 2010, 1, 1178–1188. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The diagram of the analysis. DAVID: Database for Annotation, Visualization, and Integrated Discovery; LDA: Linear discriminate analysis; mRNA: Messenger RNA; PC: Principal component; TFs: Transcription factors.
Figure 1.
The diagram of the analysis. DAVID: Database for Annotation, Visualization, and Integrated Discovery; LDA: Linear discriminate analysis; mRNA: Messenger RNA; PC: Principal component; TFs: Transcription factors.
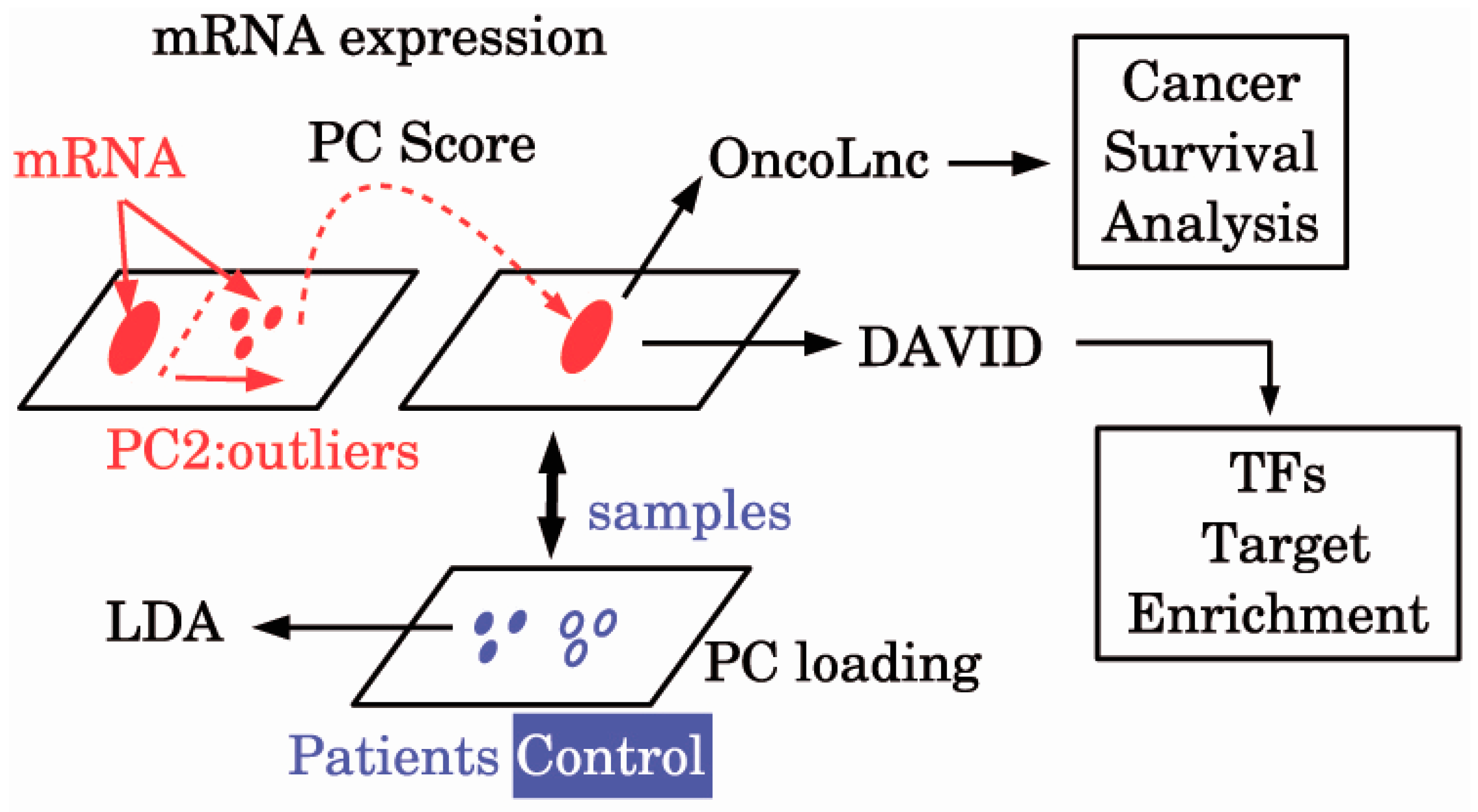
Figure 2.
Kaplan–Meier plots for (a) hepatocellular carcinoma (HCC) and (b) pancreatic adenocarcinoma (PAAD). Blue: patients with lower gene expression (lower half, N = 180 for HCC, N = 87 for PAAD). Red: patients with higher gene expression (upper half, N = 180 for HCC, N = 87 for PAAD). Most of the figures show that patients with lower gene expression (blue line) have a higher survival probability.
Figure 2.
Kaplan–Meier plots for (a) hepatocellular carcinoma (HCC) and (b) pancreatic adenocarcinoma (PAAD). Blue: patients with lower gene expression (lower half, N = 180 for HCC, N = 87 for PAAD). Red: patients with higher gene expression (upper half, N = 180 for HCC, N = 87 for PAAD). Most of the figures show that patients with lower gene expression (blue line) have a higher survival probability.

{kind=link}
{kind=link}
Table 1.
Confusion table of LDA between amyotrophic lateral sclerosis (ALS) and normal control.
| True | |||
|---|---|---|---|
| Normal | ALS | ||
| Prediction | Normal | 10 | 1 |
| ALS | 0 | 8 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Taguchi, Y.-h.; Wang, H. Genetic Association between Amyotrophic Lateral Sclerosis and Cancer. Genes 2017, 8, 243. https://doi.org/10.3390/genes8100243
AMA Style
Taguchi Y-h, Wang H. Genetic Association between Amyotrophic Lateral Sclerosis and Cancer. Genes. 2017; 8(10):243. https://doi.org/10.3390/genes8100243
Chicago/Turabian StyleTaguchi, Y-h., and Hsiuying Wang. 2017. "Genetic Association between Amyotrophic Lateral Sclerosis and Cancer" Genes 8, no. 10: 243. https://doi.org/10.3390/genes8100243
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.