
Soil Slope Exposure Affects Physico-Chemical and Microbiological Properties in Soil Aggregate Size Fractions

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
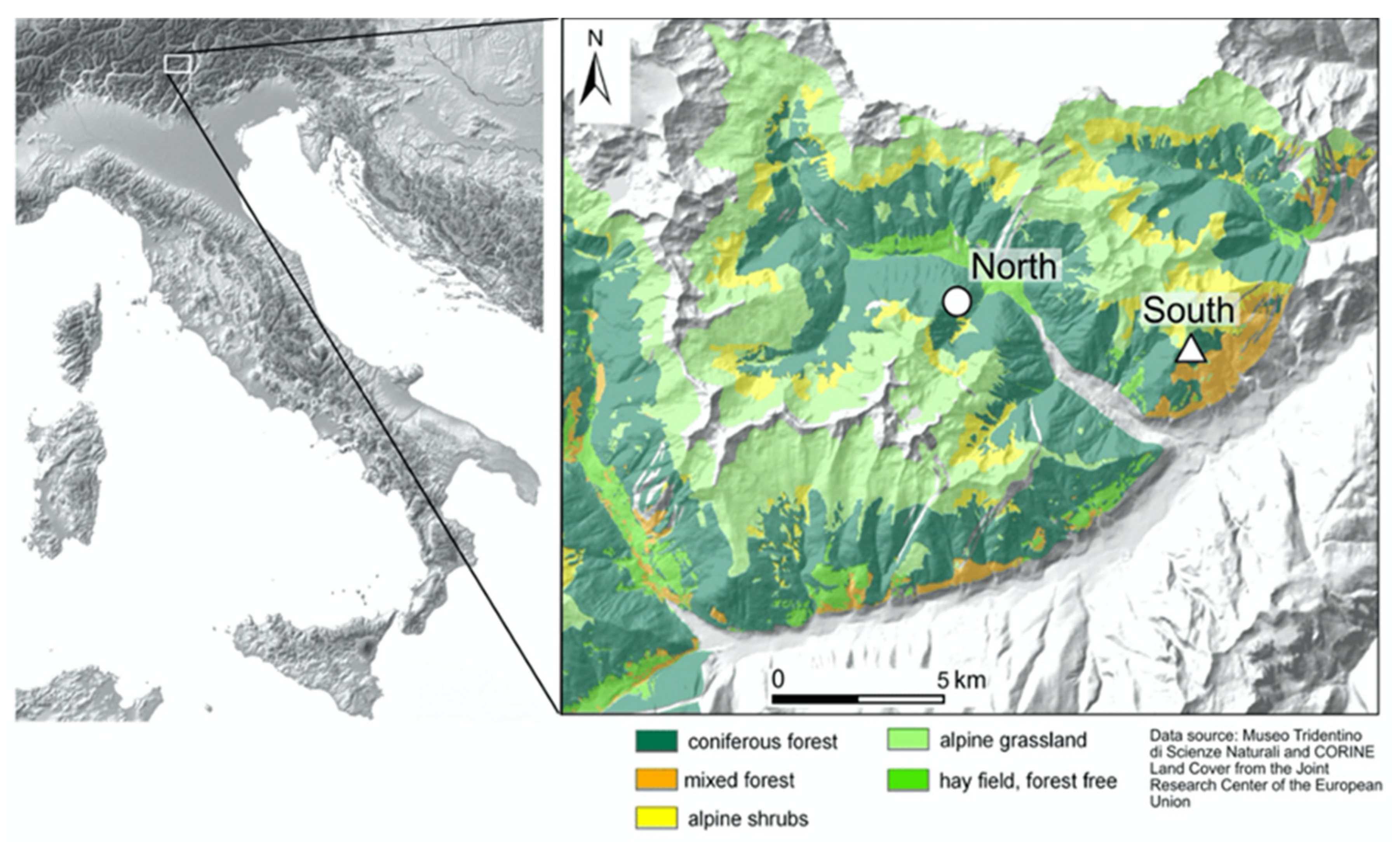
2.1. Study Sites and Experimental Design
2.2. Physico-Chemical Analyses
2.3. Microbial Biomass Index (dsDNA Yield)
2.4. Sequential DNA Extraction
2.5. Potential Enzyme Activities
2.6. Statistical Analyses
3. Results
3.1. Overview of the Aggregate Size Fractions as a Function of Exposure and Soil Horizon
3.2. Overview of the Physico-Chemical and Microbiological Properties in the of Horizon
3.3. Overview of the Physico-Chemical and Microbiological Properties in the AE Horizon
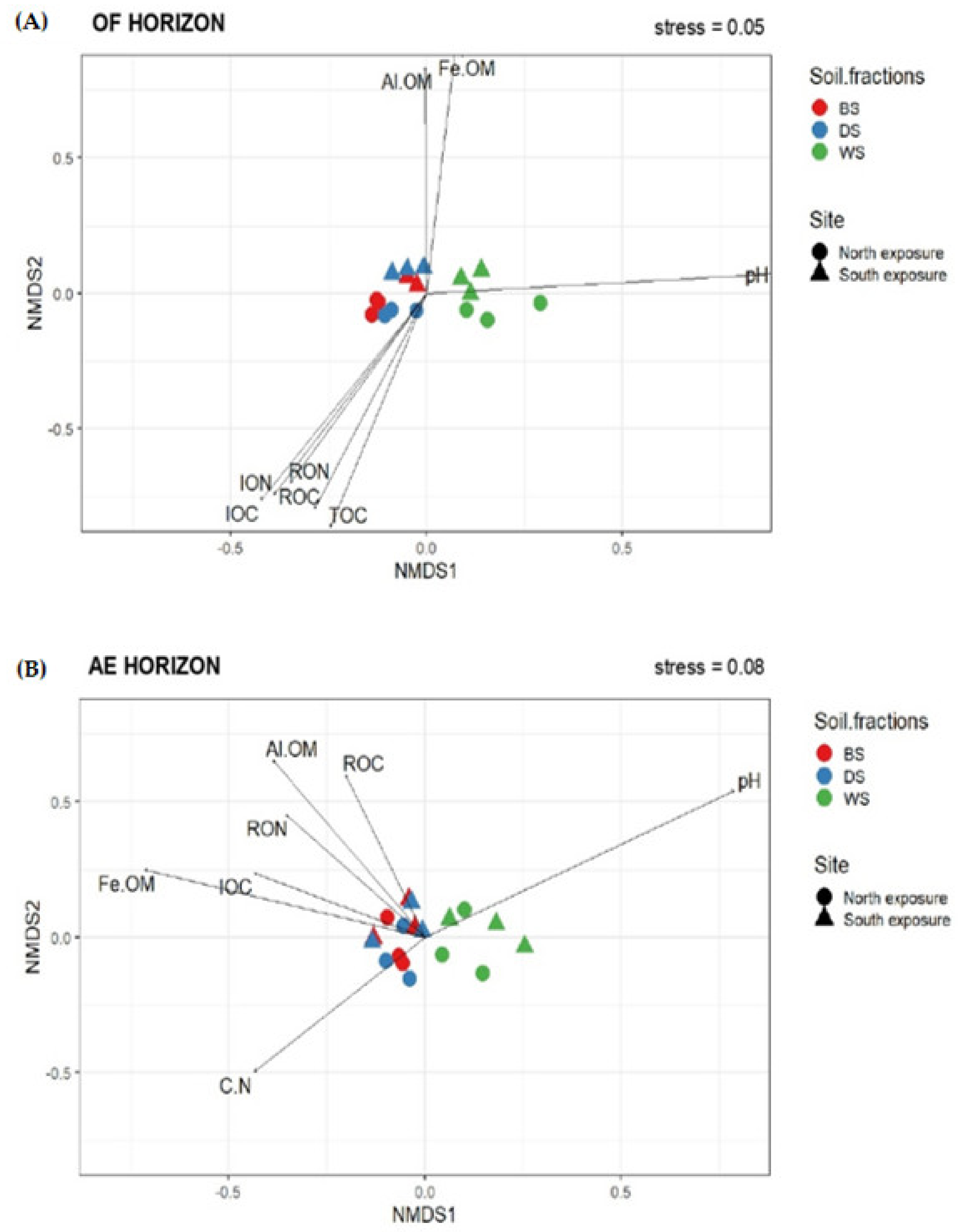
3.4. Non-Metric Multidimensional Scaling (NMDS) Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Six, J.; Paustian, K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool. Soil Biol. Biochem. 2014, 68, A4–A9. [Google Scholar] [CrossRef]
- Kim, H.; Nunan, N.; Dechesne, A.; Juarez, S.; Grundmann, G.L. The spatial distribution of exoenzyme activities across the soil micro-landscape, as measured in micro-and macro-aggregates, and ecosystem processes. Soil Biol. Biochem. 2015, 91, 258–267. [Google Scholar] [CrossRef]
- Dinakaran, J.; Mohammad, H.; Archana, M.; Rao, K.S. The chronological advancement of soil organic carbon sequestration research: A review. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2014, 84, 487–504. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; Ge, J.; He, N.; Wang, Q.; Wang, D. The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain. Appl. Soil Ecol. 2015, 86, 19–29. [Google Scholar] [CrossRef]
- Hillel, D. Fundamentals of Soil Physics; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Sarker, T.C.; Incerti, G.; Spaccini, R.; Piccolo, A.; Mazzoleni, S.; Bonanomi, G. Linking organic matter chemistry with soil aggregate stability: Insight from 13C NMR spectroscopy. Soil Biol. Biochem. 2018, 117, 175–184. [Google Scholar] [CrossRef]
- Voltolini, M.; Tas, N.; Wang, S.; Brodie, E.L.; Ajo-Franklin, J.B. Quantitative characterization of soil micro-aggregates: New opportunities from sub-micron resolution synchrotron X-ray microtomography. Geoderma 2017, 305, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Xue, K.; Wang, F.; Liu, S.; Bai, S.; Sun, B.; Zhou, J.; Yang, Y. Microbial mediation of biogeochemical cycles revealed by simulation of global changes with soil transplant and cropping. ISME J. 2014, 8, 2045–2055. [Google Scholar] [CrossRef] [Green Version]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Alagöz, Z.; Yilmaz, E. Effects of different sources of organic matter on soil aggregate formation and stability: A laboratory study on a Lithic Rhodoxeralf from Turkey. Soil Tillage Res. 2009, 103, 419–424. [Google Scholar] [CrossRef]
- Elliott, E.T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 1986, 5, 627–633. [Google Scholar] [CrossRef]
- Gunina, A.; Kuzyakov, Y. Pathways of litter C by formation of aggregates and SOM density fractions: Implications from 13C natural abundance. Soil Biol. Biochem. 2014, 71, 95–104. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Dohrmann, A.B.; Christensen, B.T.; Tebbe, C.C. Bacterial preferences for specific soil particle size fractions revealed by community analyses. Front. Microbiol. 2018, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil aggregate microbial communities: Towards understanding microbiome interactions at biologically relevant scales. Appl. Environ. Microbiol. 2019, 85, e00324-e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, R.; Li, J.; Lin, Z.; Li, Z.; Li, Y.; Yang, X.; Zhang, J.; Zhao, B. Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl. Soil Ecol. 2016, 102, 36–45. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Zühlke, D.; Becher, D.; Riedel, K.; Baldrian, P. Cellulose and hemicellulose decomposition by forest soil bacteria proceeds by the action of structurally variable enzymatic systems. Sci. Rep. 2016, 6, 25279. [Google Scholar] [CrossRef]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. Relationship between soil enzyme activities, nutrient cycling and soil fungal communities in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 795–803. [Google Scholar] [CrossRef]
- Wang, R.; Dorodnikov, M.; Yang, S.; Zhang, Y.; Filley, T.R.; Turco, R.F.; Jiang, Y. Responses of enzymatic activities within soil aggregates to 9-year nitrogen and water addition in a semi-arid grassland. Soil Biol. Biochem. 2015, 201881, 159–167. [Google Scholar] [CrossRef]
- Bhattacharyya, R.; Rabbi, S.M.; Zhang, Y.; Young, I.M.; Jones, A.R.; Dennis, P.G.; Dalal, R.C. Soil organic carbon is significantly associated with the pore geometry, microbial diversity and enzyme activity of the macro-aggregates under different land uses. Sci. Total Environ. 2021, 778, 146286. [Google Scholar] [CrossRef]
- Lagomarsino, A.; Grego, S.; Kandeler, E. Soil organic carbon distribution drives microbial activity and functional diversity in particle and aggregate-size fractions. Pedobiologia 2012, 55, 101–110. [Google Scholar] [CrossRef]
- Nie, M.; Pendall, E.; Bell, C.; Wallenstein, M.D. Soil aggregate size distribution mediates microbial climate change feedbacks. Soil Biol. Biochem. 2014, 68, 357–365. [Google Scholar] [CrossRef]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Carolan, R.; Crawley, M.J.; Clément, J.C.; Lavorel, S. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils. Soil Biol. Biochem. 2016, 96, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Turner, B.L.; Wei, K.; Tian, J.; Chen, Z.; Lü, X.; Wang, C.; Chen, L. Divergent composition and turnover of soil organic nitrogen along a climate gradient in arid and semiarid grasslands. Geoderma 2018, 327, 36–44. [Google Scholar] [CrossRef]
- Egli, M.; Sartori, G.; Mirabella, A.; Giaccai, D.; Favilli, F. Effect of north and south exposure on organic matter in high Alpine soils. Geoderma 2009, 149, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Egli, M.; Hafner, S.; Derungs, C.; Ascher-Jenull, J.; Camin, F.; Sartori, G.; Raab, G.; Paolini, M.; Bontempo, L.; Ziller, L.; et al. Decomposition and stabilisation of Norway spruce needle-derived material in Alpine soils using a 13C-labelling approach in the field. Biogeochemistry 2016, 13, 321–338. [Google Scholar] [CrossRef]
- Fravolini, G.; Egli, M.; Derungs, C.; Cherubini, P.; Ascher-Jenull, J.; Gómez-Brandón, M.; Bardelli, T.; Tognetti, R.; Lombardi, F.; Marchetti, M. Soil attributes and microclimate are important drivers of initial deadwood decay in sub-alpine Norway spruce forests. Sci. Total Environ. 2016, 569–570, 1064–1076. [Google Scholar] [CrossRef] [Green Version]
- Ascher, J.; Sartori, G.; Graefe, U.; Thornton, B.; Ceccherini, M.T.; Pietramellara, G.; Egli, M. Are humus forms, mesofauna and microflora in subalpine forest soils sensitive to thermal conditions? Biol. Fertil. Soils 2012, 48, 709–725. [Google Scholar] [CrossRef] [Green Version]
- Bardelli, T.; Gómez-Brandón, M.; Fornasier, F.; Arfaioli, P.; Egli, M.; Pietramellara, G.; Ceccherini, M.T.; Insam, H.; Ascher-Jenull, J. Chemical and microbiological changes in Norway spruce deadwood during the early stage of decomposition as a function of exposure in an Alpine setting. Arct. Antarct. Alpine Res. 2018, 50, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bardelli, T.; Ascher-Jenull, J.; Burkia-Stocker, E.; Fornasier, F.; Arfaioli, P.; Fravolini, G.; Alves Medeiros, L.R.; Egli, M.; Pietramellara, G.; Insam, H.; et al. Impact of slope exposure on chemical and microbiological properties of Norway spruce deadwood and underlying soil during early stages of decomposition in the Italian Alps. Catena 2018, 167, 110–115. [Google Scholar] [CrossRef]
- Probst, M.; Gómez-Brandón, M.; Bardelli, T.; Egli, M.; Insam, H.; Ascher-Jenull, J. Bacterial communities of decaying Norway spruce follow distinct slope exposure and time-dependent trajectories. Environ. Microbiol. 2018, 20, 3657–3670. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Probst, M.; Siles, J.A.; Peintner, U.; Bardelli, T.; Egli, M.; Insam, H.; Ascher-Jenull, J. Fungal communities and their association with nitrogen-fixing bacteria affect early decomposition of Norway spruce deadwood. Sci. Rep. 2020, 10, 8025. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Asher-Jenull, J.; Bardelli, T.; Fornasier, F.; Sartori, G.; Pietramellara, G.; Arfaioli, P.; Egli, M.; Beylich, A.; Insam, H.; et al. Ground cover and slope exposure effects on micro- and mesobiota in forest soils. Ecol. Indic. 2017, 80, 174–185. [Google Scholar] [CrossRef]
- Egli, M.; Mirabella, A.; Sartori, G.; Zanelli, R.; Bischof, S. Effect of north and south exposure on weathering rates and clay mineral formation in Alpine soils. Catena 2006, 67, 155–174. [Google Scholar] [CrossRef]
- Bardelli, T.; Gómez-Brandón, M.; Ascher-Jenull, J.; Fornasier, F.; Arfaioli, P.; Francioli, D.; Egli, M.; Sartori, G.; Insam, H.; Pietramellara, G. Effects of slope exposure on soil physico-chemical and microbiological properties along an altitudinal climosequence in the Italian Alps. Sci. Total Environ. 2017, 575, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Sboarina, C.; Cescatti, A. Il Clima Del Trentino—Distribuzione Spaziale Delle Principali Variabili Climatiche [The Climate of Trentino—Spatial Distribution of the Principal Climatic Variables]; Report 33; Centro di Ecologia Alpina Monte Bondone: Trento, Italy, 2004; p. 20. [Google Scholar]
- Diaz-Zorita, M.; Perfect, E.; Grove, G.H. Disruptive methods for assessing soil structure. Soil Till. Res. 2002, 64, 3–22. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenau, R.C. Aggregate stability and size distribution. In Methods of Soil Analysis, Part 1. Physical and Mineralogical Methods; Klute, A., Ed.; Society of Agronomy/Soil Science Society of America: Madison, WI, USA, 1986; pp. 425–442. [Google Scholar]
- Van Bavel, C.H.M. Mean Weight-Diameter of Soil Aggregates as a Statistical Index of Aggregation. Soil Sci. Soc. Am. J. 1949, 14, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Leifeld, J.; Abiven, S.; Schmidt, M.W.I.; Fuhrer, J. Sodium hypochlorite separates an older soil organic matter fraction than acid hydrolysis. Geoderma 2007, 139, 171–179. [Google Scholar] [CrossRef]
- Barral, M.T.; Arias, M.; Guérif, J. Effects of iron and organic matter on the porosity and structural stability of soil aggregates. Soil Tillage Res. 1998, 46, 261–272. [Google Scholar] [CrossRef]
- Fornasier, F.; Ascher, J.; Ceccherini, M.T.; Tomat, E.; Pietramellara, G. A simplified rapid, low-cost and versatile DNA-based assessment of soil microbial biomass. Ecol. Indic. 2014, 45, 75–82. [Google Scholar] [CrossRef]
- Ascher, J.; Ceccherini, M.T.; Pantani, O.L.; Agnelli, A.; Borgogni, F.; Guerri, G.; Nannipieri, P.; Pietramellara, G. Sequential extraction and genetic fingerprinting of a forest soil metagenome. Appl. Soil Ecol. 2009, 42, 176–181. [Google Scholar] [CrossRef]
- Fornasier, F.; Margon, A. Bovine serum albumin and Triton X-100 greatly increase phosphomonoesterases and arylsulphatase extraction yield from soil. Soil. Biol. Biochem. 2007, 39, 2682–2684. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; Simpson, G.; Sólymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package—R Package Version 1.15–2 2008. Available online: http://cran.r-project.org/ (accessed on 19 December 2013).
- Li, C.; Cao, Z.; Chang, J.; Zhang, Y.; Zhu, G.; Zong, N.; He, Y.; Zhang, J.; He, N. Elevational gradient affect functional fractions of soil organic carbon and aggregates stability in a Tibetan alpine meadow. Catena 2017, 156, 139–148. [Google Scholar] [CrossRef]
- Vignozzi, N.; Agnelli, A.E.; Brandi, G.; Gagnarli, E.; Goggioli, D.; Lagomarsino, A.; Pellegrini, S.; Simoncini, S.; Simoni, S.; Valboa, G.; et al. Soil ecosystem functions in a high-density olive orchard managed by different soil conservation practices. Appl. Soil Ecol. 2019, 134, 64–76. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, C.; Wang, J.; Meng, Q.; Yuan, Y.; Ma, X.; Liu, X.; Zhu, Y.; Ding, G.; Zhang, J.; et al. Soil aggregates stability and storage of soil organic carbon respond to cropping systems on Black Soils of Northeast China. Sci. Rep. 2020, 10, 265. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Yan, X.; Zhou, H.; Zhang, Y.Z.; Sun, H. Assessing the Contributions of sesquioxides and soil organic matter to aggregation in an Ultisol under long-term fertilization. Soil Tillage Res. 2015, 146, 89–98. [Google Scholar] [CrossRef]
- Egli, M.; Sartori, G.; Mirabella, A.; Giaccai, D.; Favilli, F.; Scherrer, D.; Krebs, R.; Delbos, E. The influence of weathering and organic matter on heavy metals lability in silicatic, Alpine soils. Sci. Total Environ. 2010, 408, 931–946. [Google Scholar] [CrossRef] [Green Version]
- Hayes, M.H.; Mylotte, R.; Swift, R.S. Humin: Its composition and importance in soil organic matter. Adv. Agron. 2017, 143, 47–138. [Google Scholar]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Nagler, M.; Podmirseg, S.M.; Griffith, G.W.; Insam, H.; Ascher-Jenull, J. The use of extracellular DNA as a proxy for specific microbial activity. Appl. Microbiol. Biot. 2018, 102, 2885–2898. [Google Scholar] [CrossRef] [Green Version]
- Nagler, M.; Insam, H.; Pietramellara, G.; Ascher-Jenull, J. Extracellular DNA in natural environments: Features, relevance and applications. Appl. Microbiol. Biot. 2018, 102, 6343–6356. [Google Scholar] [CrossRef] [Green Version]
- Pathan, S.I.; Arfaioli, P.; Ceccherini, M.T.; Ascher-Jenull, J.; Nannipieri, P.; Pietramellara, G. Physical protection of extracellular and intracellular DNA in soil aggregates against simulated natural oxidative processes. Appl Soil Ecol. 2021, 165, 104002. [Google Scholar] [CrossRef]
- Bardelli, T.; Rossetti, L.; Zago, M.; Carminati, D.; Giraffa, G.; Tidona, F. Extracellular and intracellular DNA for bacterial profiling of long-ripened cheeses. FEMS Microbiol. Lett. 2020, 367, 13. [Google Scholar] [CrossRef] [PubMed]
- Ceccherini, M.T.; Ascher, J.; Agnelli, A.; Borgogni, F.; Pantani, O.L.; Pietramellara, G. Experimental discrimination and molecular characterization of the extracellular soil DNA fraction. Anton Leeuw. Antonie Van Leeuwenhoek 2009, 96, 653–657. [Google Scholar]
- Nielsen, K.M.; Johnsen, P.J.; Bensasson, D.; Daffonchio, D. Release and persistence of extracellular DNA in the environment. Environ. Res. 2007, 6, 37–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietramellara, G.; Ascher, J.; Borgogni, F.; Ceccherini, M.T.; Guerri, G.; Nannipieri, P. Extracellular DNA in soil and sediment: Fate and ecological relevance. Biol. Fertil. Soils 2009, 45, 219–235. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Theerthankar, D.; Sharma, P.K.; Busscher, H.J.; van der Mei, H.C.; Krom, B.P. Role of extracellular DNA in initial bacterial adhesion and surface aggregation. Appl. Environ. Microbiol. 2010, 76, 3405–3408. [Google Scholar]
- Costa, O.Y.; Raaijmakers, J.M.; Kuramae, E.E. Microbial extracellular polymeric substances: Ecological function and impact on soil aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef] [Green Version]
- Fraser, T.D.; Lynch, D.H.; Bent, E.; Entz, M.H.; Dunfield, K.E. Soil bacterial phoD gene abundance and expression in response to applied phosphorus and long-term management. Soil Biol. Biochem. 2015, 88, 137–147. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure | Soil Fractions | pH | Fe-OM (mg g−1 dw) | Al-OM (mg g−1 dw) | Mn-OM (mg g−1 dw) | TOC (%) | ROC (%) | LOC (%) | IOC (%) | TON (%) | RON (%) | ION (%) | C/N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| North | BS | 5.1 (0.2) c | 570 (385.2) b | 461 (88.8) c | 8.4 (7.3) b | 41.6 (2.4) a | 28.8 (3.6) a | 12.8 (1.8) a | 27.7 (2.7) a | 1.2 (0.01) a | 2.8 (0.5) a | 0.4 (0.1) a | 35.5 (2.3) ab |
| DS | 5.4 (0.4) b | 533 (179.6) b | 453 (56.0) c | 8.1 (6.6) b | 41.0 (2.3) a | 24.3 (8.1) ab | 16.7 (5.8) a | 26.9 (1.7) ab | 1.4 (0.1) a | 2.5 (0.9) a | 0.4 (0.01) a | 28.6 (0.8) bc | |
| WS | 6.4 (0.4) a | 553 (371.8) b | 561 (238.7) c | 17.4 (11.4) ab | 37.9 (7.2) ab | 22.9 (6.8) ab | 14.9 (1.9) a | 19.6 (6.3) bc | 1.0 (0.3) a | 2.3 (0.7) a | 0.3 (0.1) ab | 40.4 (8.9) a | |
| South | BS | 5.2 (0.3) c | 1583 (119.2) a | 2538 (629.1) a | 23.4 (6.6) a | 29.4 (3.1) bc | 13.5 (2.4) bc | 16.0 (1.6) a | 18.2 (2.5) c | 0.9 (0.3) a | 1.6 (0.5) ab | 0.2 (0.1) b | 35.1 (8.1) ab |
| DS | 5.6 (0.3) b | 1313 (65.4) a | 1739 (470.4) b | 26.1 (6.0) a | 29.7 (5.2) bc | 17.0 (7.8) bc | 12.7 (3.0) a | 17.7 (6.3) c | 1.1 (0.3) a | 2.1 (1.0) ab | 0.3 (0.1) ab | 27.1 (3.4) bc | |
| WS | 6.5 (0.2) a | 1445 (204.6) a | 1562 (249.0) b | 16.2 (7.6) ab | 25.7 (7.9) c | 8.2 (4.9) c | 17.5 (3.7) a | 5.7 (3.4) d | 1.1 (0.3) a | 0.8 (0.5) b | 0.01 (0.1) c | 22.9 (2.3) c |
| Exposure | Soil Fractions | Alfagluc | Betagluc | Cell | xylo | uroni | ester_nona | chit | leu | acP | alkP | bisP | piroP | aryS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| North | BS | 18.2 (4.8) a | 281.4 (158.2) a | 61.8 (42.8) b | 40.6 (14.5) a | 23.8 (7.3) ab | 3053 (662.6) ab | 231.3 (88.4) a | 339 (172.7) a | 1351 (68.8) a | 255 (70.5) c | 55.9 (15.2) c | 11.1 (5.1) b | 192 (26.7) ab |
| DS | 24.5 (4.9) a | 179.5 (35.6) ab | 54.5 (40.1) b | 35.9 (16.6) a | 23.2 (4.9) abc | 4086 (534.0) a | 291.2 (27.4) a | 542 (224.1) a | 1437 (204.0) a | 410 (260.4) bc | 82.0 (40.1) bc | 18.6 (15.8) b | 237 (64.3) a | |
| WS | 26.8 (8.7) a | 72.3 (2.6) c | 12.4 (4.4) b | 22.2 (5.2) ab | 12.3 (4.7) bc | 2107 (53.9) b | 110.9 (43.4) b | 367 (191.1) a | 294 (106.2) b | 2102 (1763.0) a | 233 (135.7) a | 84.9 (51.3) a | 77.4 (31.0) c | |
| South | BS | 18.4 (7.1) a | 261.3 (133.3) ab | 70.4 (46.2) b | 36.5 (14.2) a | 22.0 (8.6) abc | 3590 (1730.0) ab | 111.6 (63.8) b | 403 (145.6) a | 2977 (1642.2) a | 462 (139.5) bc | 159 (59.0) ab | 60.3 (14.3) a | 195 (50.4) ab |
| DS | 21.0 (1.7) a | 265.1 (49.8) ab | 134.4 (44.7) a | 35.7 (8.2) a | 30.5 (6.9) a | 5021 (2649.7) a | 124.8 (55.2) b | 406 (100.4) a | 2209 (1389.8) a | 467 (49.5) bc | 110 (19.5) abc | 54.0 (34.5) a | 205 (32.1) ab | |
| WS | 21.0 (6.9) a | 99.2 (11.1) bc | 19.6 (2.1) b | 12.8 (1.9) b | 11.8 (1.6) c | 2070 (438.8) b | 78.0 (15.3) b | 314 (129.1) a | 340 (59.3) b | 592 (58.1) b | 116 (30.6) abc | 73.3 (53.7) a | 125 (42.6) bc |
| Slope Exposure (North vs. South) | Sample Type (BS vs. DS vs. WS) | Interaction | ||||
|---|---|---|---|---|---|---|
| OF | F | p | F | OF | F | p |
| pH | 1.90 | ns | 47.81 | pH | 1.90 | ns |
| Fe-OM | 57.19 | *** | 0.56 | Fe-OM | 57.19 | *** |
| Al-OM | 76.50 | *** | 2.85 | Al-OM | 76.50 | *** |
| Mn-OM | 9.39 | ** | 0.04 | Mn-OM | 9.39 | ** |
| TOC | 23.60 | *** | 0.99 | TOC | 23.60 | *** |
| ROC | 19.56 | *** | 1.61 | ROC | 19.56 | *** |
| LOC | 0.15 | ns | 0.53 | LOC | 0.15 | ns |
| IOC | 29.66 | *** | 11.12 | IOC | 29.66 | *** |
| TON | 1.83 | ns | 1.76 | TON | 1.83 | ns |
| RON | 9.27 | * | 2.22 | RON | 9.27 | * |
| ION | 27.44 | *** | 8.36 | ION | 27.44 | *** |
| C/N | 6.69 | * | 2.92 | C/N | 6.69 | * |
| exDNA | 51.82 | *** | 74.77 | exDNA | 51.82 | *** |
| iDNA | 8.43 | * | 7.11 | iDNA | 8.43 | * |
| exDNA/iDNA | 60.99 | *** | 17.69 | exDNA/iDNA | 60.99 | *** |
| dsDNA | 11.95 | ** | 1.83 | dsDNA | 11.95 | ** |
| alfagluc | 1.30 | ns | 1.68 | alfagluc | 1.30 | ns |
| betagluc | 0.01 | ns | 8.61 | betagluc | 0.01 | ns |
| cell | 4.9 | * | 10.12 | cell | 4.9 | * |
| xylo | 0.70 | ns | 6.02 | xylo | 0.70 | ns |
| uroni | 0.34 | ns | 9.49 | uroni | 0.34 | ns |
| ester_nona | 0.14 | ns | 7.20 | ester_nona | 0.14 | ns |
| chit | 25.7 | *** | 10.2 | chit | 25.7 | *** |
| leu | 0.34 | ns | 1.26 | leu | 0.34 | ns |
| acP | 4.32 | ns | 39.38 | acP | 4.32 | ns |
| alkP | 0.11 | ns | 10.48 | alkP | 0.11 | ns |
| bisP | 2.01 | ns | 3.72 | bisP | 2.01 | ns |
| piroP | 10.29 | ** | 5.32 | piroP | 10.29 | ** |
| aryS | 0.09 | ns | 12.69 | aryS | 0.09 | ns |
| Exposure | Soil Fractions | pH | Fe-OM (mg g−1 dw) | Al-OM (mg g−1 dw) | Mn-OM (mg g−1 dw) | TOC (%) | ROC (%) | LOC (%) | IOC (%) | TON (%) | RON (%) | ION (%) | C/N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| North | BS | 5.1 (0.2) c | 5293 (1508.3) a | 3616 (780.1) ab | 20.1 (11.5) a | 5.6 (1.2) b | 1.7 (0.4) c | 3.8 (0.8) c | 1.0 (0.2) b | 0.2 (0.1) b | 0.4 (0.1) ab | udl | 44.5 (21.8) ab |
| DS | 4.9 (0.5) c | 5180 (884.6) a | 2801 (526.8) bc | 17.6 (9.8) a | 7.2 (2.1) b | 1.6 (0.4) c | 5.5 (1.7) abc | 2.5 (2.2) b | 0.1 (0.1) b | 0.2 (0.1) b | udl | 55.8 (10.5) a | |
| WS | 5.8 (0.4) b | 4320 (937.8) a | 2607 (671.1) c | 18.5 (9.8) a | 5.8 (2.1) b | 1.3 (0.2) c | 4.5 (1.9) bc | 0.7 (0.2) b | 0.3 (0.1) ab | 0.2 (0.1) b | udl | 19.3 (3.1) c | |
| South | BS | 5.3 (0.5) bc | 4608 (1439.7) a | 4040 (259.4) a | 12.6 (7.7) a | 12.9 (3.3) a | 4.4 (0.9) a | 8.5 (2.5) ab | 4.8 (0.6) a | 0.5 (0.1) a | 0.6 (0.3) a | udl | 28.6 (2.6) bc |
| DS | 5.3 (0.4) bc | 5432 (597.2) a | 4054 (188.6) a | 10.3 (4.8) a | 12.5 (3.5) a | 2.9 (0.7) b | 9.6 (3.3) a | 2.1 (0.1) b | 0.4 (0.2) a | 0.4 (0.01) ab | udl | 35.8 (6.1) bc | |
| WS | 6.8 (0.1) a | 3206 (1076.1) a | 3024 (191.3) bc | 9.6 (4.2) a | 8.6 (2.3) ab | 2.0 (0.2) bc | 6.6 (2.2) abc | 1.1 (0.1) b | 0.4 (0.1) a | 0.2 (0.1) b | udl | 23.2 (2.2) c |
| Exposure | Soil fractions | Alfagluc | Betagluc | Cell | xylo | uroni | ester_nona | chit | leu | acP | alkP | bisP | piroP | aryS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| North | BS | 3.2 (1.2) a | 27.5 (7.4) b | 3.9 (3.0) a | 4.5 (1.0) bc | 4.5 (3.8) ab | 346 (103.0) ab | 19.9 (5.5) bc | 49.7 (19.3) b | 622 (495.8) ab | 48.9 (47.7) a | 32.9 (27.2) ab | 11.5 (8.7) ab | 310 (166.1) ab |
| DS | 3.6 (2.0) a | 27.6 (9.9) b | 2.1 (0.5) a | 4.6 (1.7) bc | 4.2 (2.8) ab | 358 (96.4) ab | 21.8 (4.5) bc | 52.5 (35.2) b | 575 (554.5) ab | 47.6 (45.5) a | 31.0 (27.9) b | 13.1 (8.3) ab | 272 (180.6) ab | |
| WS | 4.3 (2.7) a | 15.1 (3.4) b | 1.5 (1.3) a | 2.4 (0.8) c | 4.1 (2.5) ab | 478 (132.3) ab | 15.5 (3.9) bc | 97.6 (42.8) ab | 195 (111.0) bc | 115 (108.8) a | 40.3 (39.6) ab | 13.5 (11.2) ab | 162 (76.4) ab | |
| South | BS | 7.8 (3.8) a | 60.8 (33.7) ab | 10.6 (7.7) a | 11.6 (4.9) a | 11.6 (5.5) a | 867 (617.7) a | 38.0 (11.4) ab | 165.2 (79.1) a | 1498 (400.4) a | 208 (187.8) a | 110 (55.8) a | 75.2 (64.3) a | 386 (186.3) a |
| DS | 7.2 (2.6) a | 46.6 (10.4) ab | 7.6 (3.5) a | 9.1 (2.6) ab | 10.1 (3.0) ab | 781 (323.3) a | 50.4 (12.0) a | 140.1 (65.6) ab | 1280 (503.7) a | 151 (131.2) a | 97.9 (41.2) a | 63.0 (55.7) a | 321 (145.4) ab | |
| WS | 4.0 (0.4) a | 14.8 (15.1) b | 1.9 (2.3) a | 2.6 (3.2) c | 4.0 (4.7) b | 430 (217.6) a | 13.1 (9.6) c | 90.2 (52.2) ab | 99.6 (90.3) c | 106 (62.4) a | 43.5 (15.2) ab | 9.1 (5.3) b | 70.3 (34.9) b |
| Slope Exposure (North- vs. South) | Sample Type (BS vs. DS vs. WS) | Interaction | ||||
|---|---|---|---|---|---|---|
| AE | F | p | F | p | F | p |
| pH | 11.06 | ** | 20.24 | *** | 2.07 | ns |
| Fe-OM | 0.98 | ns | 3.20 | ns | 0.60 | ns |
| Al-OM | 9.02 | * | 6.43 | * | 1.43 | ns |
| Mn-OM | 3.97 | ns | 0.16 | ns | 0.02 | ns |
| TOC | 18.24 | ** | 1.74 | ns | 1.66 | ns |
| ROC | 37.48 | *** | 10.41 | ** | 5.32 | * |
| LOC | 12.39 | ** | 1.39 | ns | 0.58 | ns |
| IOC | 8.23 | * | 7.20 | ** | 8.36 | ** |
| TON | 15.77 | ** | 0.92 | ns | 1.69 | ns |
| RON | 4.93 | * | 8.15 | ** | 1.1 | ns |
| ION | na | na | na | na | na | na |
| C/N | 4.74 | * | 8.59 | ** | 2.28 | ns |
| exDNA | 13.63 | ** | 20.53 | *** | 9.44 | ** |
| iDNA | 4.97 | * | 5.54 | * | 1.20 | ns |
| exDNA/iDNA | 0.61 | ns | 0.57 | ns | 0.54 | ns |
| dsDNA | 4.81 | * | 1.26 | ns | 3.96 | * |
| alfagluc | 5.52 | * | 0.58 | ns | 1.72 | ns |
| betagluc | 4.93 | * | 5.12 | * | 1.56 | ns |
| cell | 0.01 | ns | 1.34 | ns | 0.78 | ns |
| xylo | 9.11 | * | 6.78 | * | 2.38 | ns |
| uroni | 5.45 | * | 1.77 | ns | 1.49 | ns |
| ester_nona | 5.11 | * | 0.26 | ns | 2.34 | ns |
| chit | 6.47 | * | 3.34 | ns | 2.16 | ns |
| leu | 6.87 | * | 0.11 | ns | 2.23 | ns |
| acP | 1.24 | ns | 15.20 | *** | 3.91 | * |
| alkP | 4.23 | ns | 0.29 | ns | 0.89 | ns |
| bisP | 9.15 | * | 0.40 | ns | 1.10 | ns |
| piroP | 7.17 | * | 0.29 | ns | 1.44 | ns |
| aryS | 0.02 | ns | 4.34 | * | 0.59 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardelli, T.; Pathan, S.I.; Arfaioli, P.; Vignozzi, N.; Pellegrini, S.; Fornasier, F.; Egli, M.; Gómez-Brandón, M.; Insam, H.; Pietramellara, G.; et al. Soil Slope Exposure Affects Physico-Chemical and Microbiological Properties in Soil Aggregate Size Fractions. Land 2022, 11, 750. https://doi.org/10.3390/land11050750
Bardelli T, Pathan SI, Arfaioli P, Vignozzi N, Pellegrini S, Fornasier F, Egli M, Gómez-Brandón M, Insam H, Pietramellara G, et al. Soil Slope Exposure Affects Physico-Chemical and Microbiological Properties in Soil Aggregate Size Fractions. Land. 2022; 11(5):750. https://doi.org/10.3390/land11050750
Chicago/Turabian StyleBardelli, Tommaso, Shamina Imran Pathan, Paola Arfaioli, Nadia Vignozzi, Sergio Pellegrini, Flavio Fornasier, Markus Egli, María Gómez-Brandón, Heribert Insam, Giacomo Pietramellara, and et al. 2022. "Soil Slope Exposure Affects Physico-Chemical and Microbiological Properties in Soil Aggregate Size Fractions" Land 11, no. 5: 750. https://doi.org/10.3390/land11050750