
Multicropping Pattern Reveals Human Adaptation at the Wanbei Site (ca. 5.7–4.4 ka cal. BP) in the Middle and Lower Huai River Valley, China
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
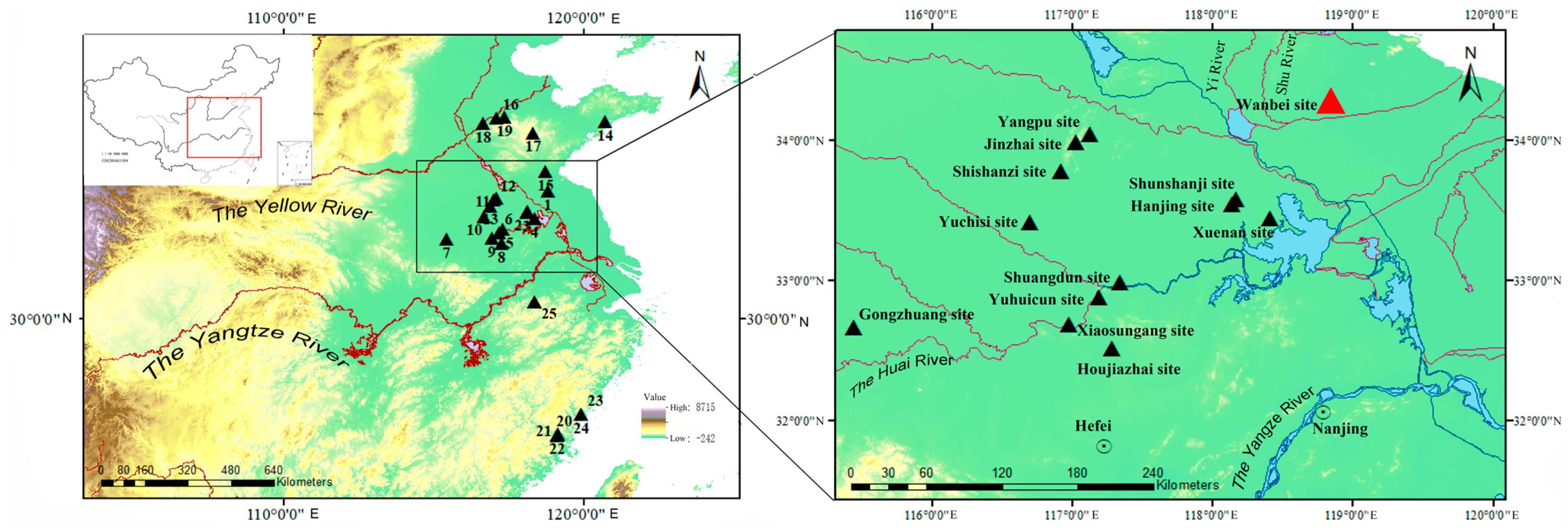
:1. Introduction
2. Materials and Methods
3. Results
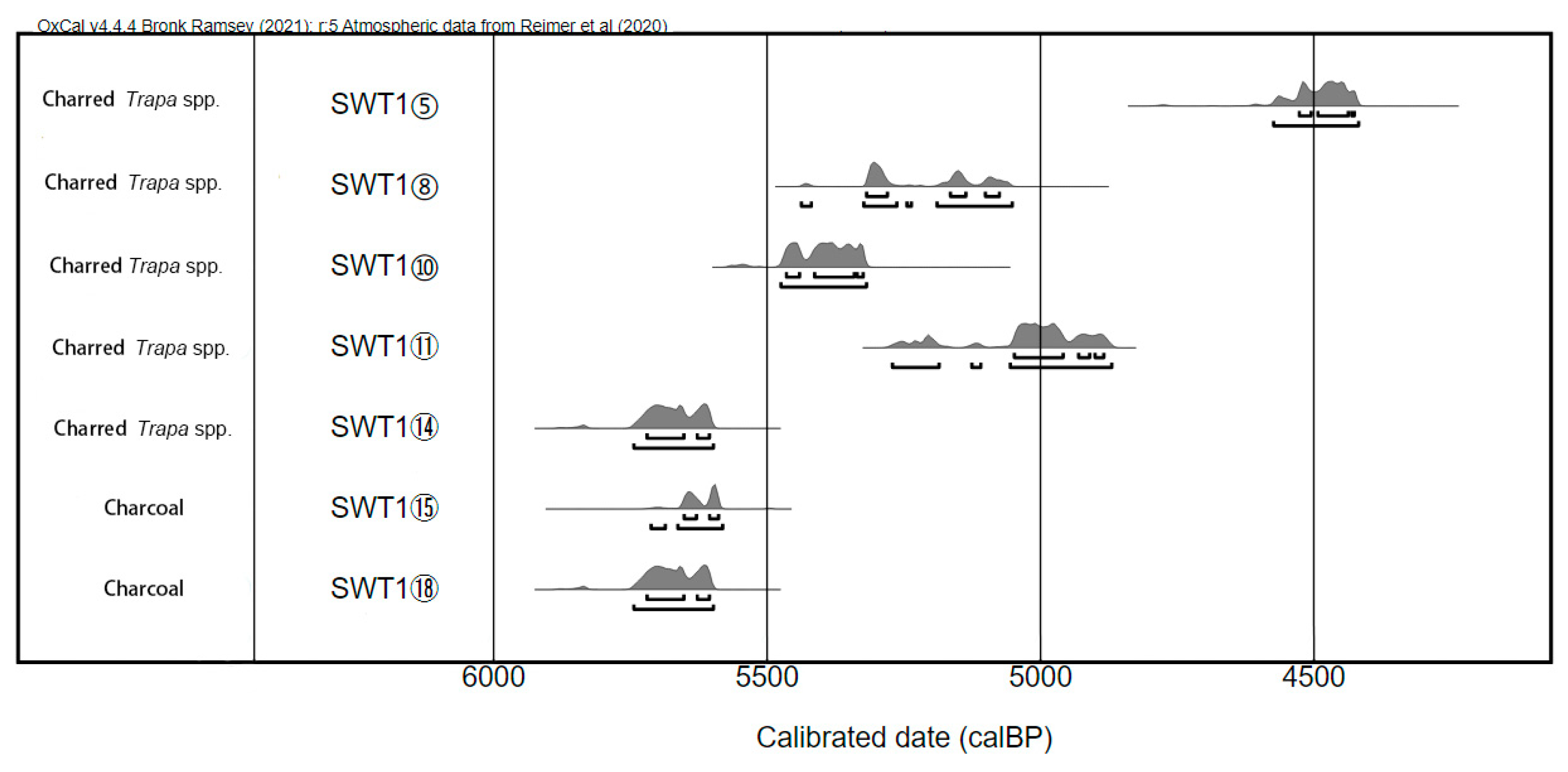
3.1. Chronology
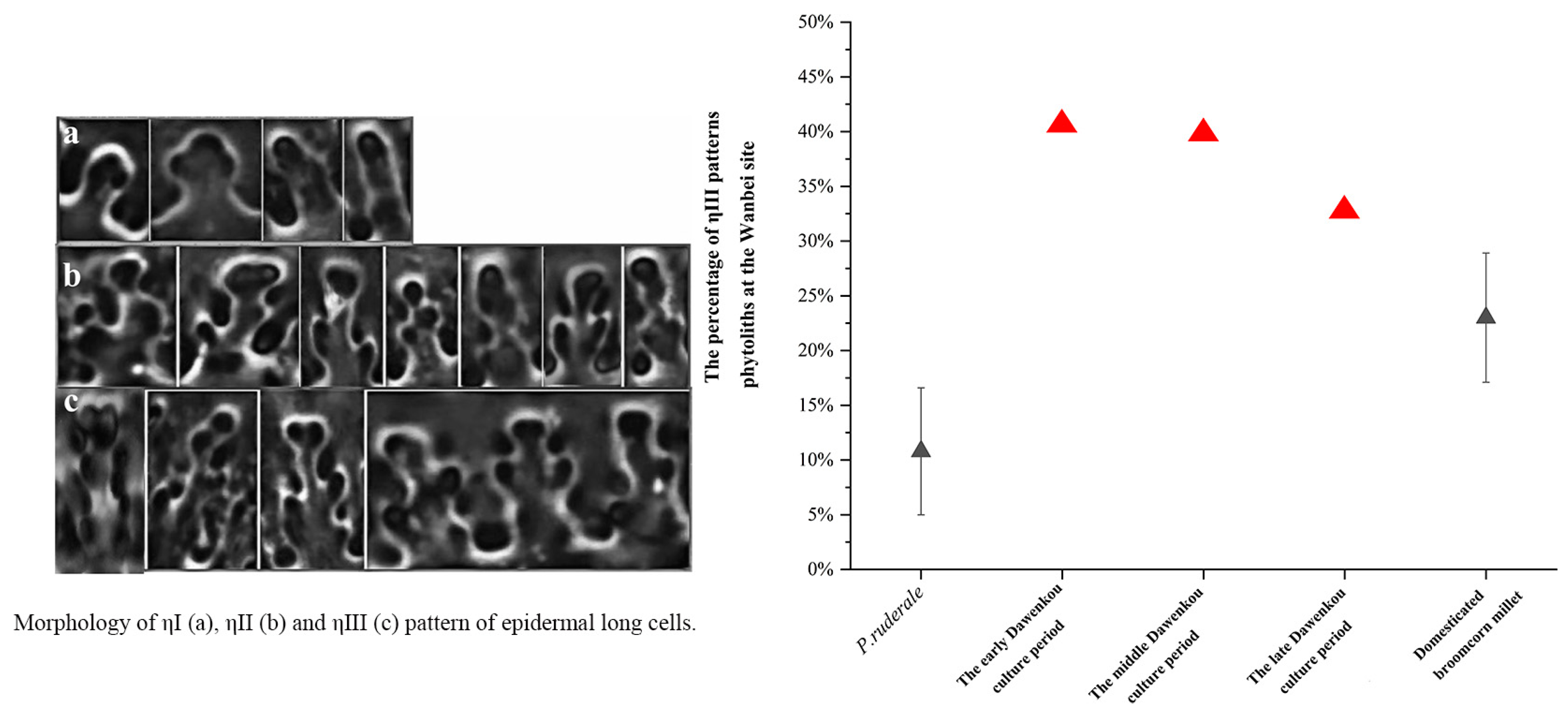
3.2. Phytolith Results
3.2.1. Phytoliths from Ash Pits
3.2.2. Phytoliths from the Profile
4. Discussion
4.1. Multicropping Pattern at Wanbei
4.2. Domestication of Rice and Millets
4.3. Cropping Pattern Response to Climatic Environment and Cultural Exchange at Wanbei
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, F.; Dong, G.; Zhang, D.; Liu, X.; Jia, X.; An, C.-B.; Ma, M.; Xie, Y.; Barton, L.; Ren, X. Agriculture Facilitated Permanent Human Occupation of the Tibetan Plateau after 3600 BP. Science 2015, 347, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z. New Archaeobotanic Data for the Study of the Origins of Agriculture in China. Curr. Anthropol. 2011, 52, S295–S306. [Google Scholar] [CrossRef]
- Li, R.; Lv, F.; Yang, L.; Liu, F.; Liu, R.; Dong, G. Spatial–Temporal Variation of Cropping Patterns in Relation to Climate Change in Neolithic China. Atmosphere 2020, 11, 677. [Google Scholar] [CrossRef]
- Gupta, A.K. Origin of Agriculture and Domestication of Plants and Animals Linked to Early Holocene Climate Amelioration. Curr. Sci. 2004, 87, 54–59. [Google Scholar]
- Yang, X.; Wu, W.; Perry, L.; Ma, Z.; Bar-Yosef, O.; Cohen, D.J.; Zheng, H.; Ge, Q. Critical Role of Climate Change in Plant Selection and Millet Domestication in North China. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, J.; Liu, K.; Wu, N.; Li, Y.; Zhou, K.; Ye, M.; Zhang, T.; Zhang, H.; Yang, X. Earliest Domestication of Common Millet (Panicum Miliaceum) in East Asia Extended to 10,000 Years Ago. Proc. Natl. Acad. Sci. USA 2009, 106, 7367–7372. [Google Scholar] [CrossRef]
- Zhao, Z. Plant Archaeological Research on the Origin of Agriculture and Civilization. Manag. Rev. Soc. Sci. 2005, 2, 82–91. (In Chinese) [Google Scholar]
- Zuo, X.; Lu, H.; Jiang, L.; Zhang, J.; Yang, X.; Huan, X.; He, K.; Wang, C.; Wu, N. Dating Rice Remains through Phytolith Carbon-14 Study Reveals Domestication at the Beginning of the Holocene. Proc. Natl. Acad. Sci. USA 2017, 114, 6486–6491. [Google Scholar] [CrossRef]
- Jones, M.K.; Liu, X. Origins of Agriculture in East Asia. Science 2009, 324, 730–731. [Google Scholar] [CrossRef]
- Zhao, Z. The Middle Yangtze Region in China Is One Place Where Rice Was Domesticated: Phytolith Evidence from the Diaotonghuan Cave, Northern Jiangxi. Antiquity 1998, 72, 885–897. [Google Scholar]
- Zhao, Y.; Yu, Z.; Chen, F.; Zhang, J.; Yang, B. Vegetation Response to Holocene Climate Change in Monsoon-Influenced Region of China. Earth-Sci. Rev. 2009, 97, 242–256. [Google Scholar] [CrossRef]
- He, Y.; Theakstone, W.H.; Zhang, Z.; Zhang, D.; Yao, T.; Chen, T.; Shen, Y.; Pang, H. Asynchronous Holocene Climatic Change across China. Quat. Res. 2004, 61, 52–63. [Google Scholar] [CrossRef]
- Yang, M.; Fan, X.; Sun, B.; Chen, C.; Lang, J.; Ko, Y.-C.; Tsang, C.; Chiu, H.; Wang, T.; Bao, Q. Ancient DNA Indicates Human Population Shifts and Admixture in Northern and Southern China. Science 2020, 369, 282–288. [Google Scholar] [CrossRef]
- He, K.; Lu, H.; Zhang, J.; Wang, C.; Huan, X. Prehistoric Evolution of the Dualistic Structure Mixed Rice and Millet Farming in China. Holocene 2017, 27, 1885–1898. [Google Scholar] [CrossRef]
- Zhou, C. Forming and Development of Pre-History Culture in Huai River Basin. J. Anhui Univ. (Philos. Soc.Sci.) 1999, 23, 6–12. (In Chinese) [Google Scholar]
- Yang, Y.; Cheng, Z.; Li, W.; Yao, L.; Li, Z.; Luo, W.; Yuan, Z.; Zhang, J.; Zhang, J. The Emergence, Development and Regional Differences of Mixed Farming of Rice and Millet in the Upper and Middle Huai River Valley, China. Sci. China Earth Sci. 2016, 46, 1037–1050. (In Chinese) [Google Scholar] [CrossRef]
- Deng, Z.; Hung, H.; Fan, X.; Huang, Y.; Lu, H. The Ancient Dispersal of Millets in Southern China: New Archaeological Evidence. Holocene 2018, 28, 34–43. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhuang, L.; Lin, L. Phytolith Evidence for Rice Domestication of the Hanjing Site in Sihong, Jiangsu Province and the Related Issues. Southeast Cult. 2018, 2, 71–80. (In Chinese) [Google Scholar]
- Institute of archaeology of Nanjing Museum; Sihong County Museum. The Excavation of the Shunshanji Site of Neolithic Age in Sihong County, Jiangsu. Acta Archaeol. Sin. 2014, 4, 519–562. (In Chinese) [Google Scholar]
- Institute of Archaeology of Nanjing Museum; Sihong County Museum. Shunshanji: An Archaeological Excavation Report of Neolithic Site in Sihong County; Science Press: Beijing, China, 2016; pp. 1–421. (In Chinese) [Google Scholar]
- Yang, Y.; Li, W.; Yao, L.; Cheng, Z.; Luo, W.; Zhang, J.; Lin, L.; Gan, H.; Yan, L. Plant Food Sources and Stone Tools’ Function at the Site of Shunshanji Based on Starch Grain Analysis. Sci. China Earth Sci. 2016, 59, 1574–1582. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhuang, L.; Lin, L. A Preliminary Study on Plant Resources and Environmental Landscape of the Xuenan Site in Sihong, Jiangsu Province. J. Natl. Mus. China 2021, 8, 24–41. (In Chinese) [Google Scholar]
- Luo, W.; Yang, Y.; Yao, L.; Chen, Z.; Li, J.; Yin, C.; Zhang, J.; Lin, L.; Gan, H. Phytolith Records of Rice Agriculture during the Middle Neolithic in the Middle Reaches of Huai River Region, China. Quat. Int. 2016, 426, 133–140. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M. More Study of the Original Rice Remains at the Longqiuzhuang Site in Gaoyou. Agric. Archaeol. 1998, 1, 172–181. (In Chinese) [Google Scholar]
- Cheng, Z.; Yang, Y.; Zhang, J.; Fang, F.; Yu, J.; Chen, B.; Chen, C.; Zhang, H.; Gong, X. Research on Charred Plant Remains from The Xiaosungang Site in Huainan City, Anhui Province. Quat. Sci. 2016, 36, 302–311. (In Chinese) [Google Scholar]
- Luo, W.; Xuan, H.; Yao, L. Starch Grain Evidence of Utilizing Plant in Phase II of the Houjiazhai Site in Dingyuan County, Anhui Province. Acta Anthropol. Sin. 2020, 39, 292–305. (In Chinese) [Google Scholar]
- Luo, W.; Gu, C.; Yang, Y.; Zhang, D.; Liang, Z.; Li, J.; Huang, C.; Zhang, J. Phytoliths Reveal the Earliest Interplay of Rice and Broomcorn Millet at the Site of Shuangdun (ca. 7.3–6.8 Ka BP) in the Middle Huai River Valley, China. J. Archaeol. Sci. 2019, 102, 26–34. [Google Scholar] [CrossRef]
- Li, W.; Luo, W.; Yao, L.; Xuan, H.; Yi, W.; Tian, W.; Zhang, D.; Sun, Y.; Kan, X.; Zhang, J. Pottery Use and Starchy Foods During the Shuangdun Culture (ca. 7.3–6.8 Ka BP) in the Middle Catchment of the Huai River, China. Front. Earth Sci. 2023, 10, 886179. [Google Scholar] [CrossRef]
- Gu, C.; Luo, W.; Zhang, D.; Yang, Y. Phytolith Evidence for the Agricultural Development during Shuangdun Culture Period from the Yuhuicun Site, Anhui Province. Acta Anthropol. Sin. 2023, 42, 110. (In Chinese) [Google Scholar]
- Luo, W.; Yang, Y.; Zhang, D.; Liang, Z.; Fang, F.; Huang, C. Phytolith Evidence for the Development of Agriculture between Early Dawenkou and Longshan Cultures at the Gongzhuang Site in Linquan County, Anhui Province. Acta Micropaleontol. Sin. 2018, 35, 370–380. (In Chinese) [Google Scholar]
- Cheng, Z.; Yang, Y.; Yuan, Z.; Zhang, J.; Yu, J.; Chen, B.; Zhang, H.; Gong, X. A Study on Charred Plant Remains in Yangpu Site of Suzhou, Anhui. Jianghan Archaeol. 2016, 1, 95–103. (In Chinese) [Google Scholar]
- Yang, F.; Zhang, X.; Jin, G. Research on Plant Remains from the Jinzhai Site (2016) in Xiaoxian County of Anhui Province. Agric. Archaeol. 2018, 4, 426–433. (In Chinese) [Google Scholar]
- Yang, F.; Duan, Y.; Zhang, X.; Jin, G. Research on Plant Remains from the Jinzhai Site (2017) in Xiaoxian County of Anhui Province. Southeaset Cult. 2020, 3, 112–121. (In Chinese) [Google Scholar]
- Zhang, J.; Yang, Y.; Zhang, Y.; Cheng, Z.; Zhang, Z.; Zhang, J. Research on the Charred Plant Remains from the Diaoyutai Site in Bengbu City, Anhui Province. Quat. Sci. 2018, 38, 393–405. (In Chinese) [Google Scholar]
- Tang, L.; Sun, J.; Zhang, M.; Li, M. The Primitive Rice Cultivation at Longqiuzhuang Site of Gaoyou. Acta Agron. Sin. 1996, 22, 608–612. (In Chinese) [Google Scholar]
- Zhang, Y.; Zhang, D.; Yang, Y.; Wu, X. Pollen and Lipid Analysis of Coprolites from Yuhuicun and Houtieying, China: Implications for Human Habitats and Diets. J. Archaeol.Sci. Rep. 2020, 29, 102135. [Google Scholar] [CrossRef]
- Zhao, Z. Analysis Report on Flotation Results of Yuchisi Site in Mengcheng, Anhui Province. In Paleoethnobotany: Theories, Methods and Practices; Zhao, Z., Ed.; Chinese Science Publishing: Beijing, China, 2010; pp. 1–1300. (In Chinese) [Google Scholar]
- Cheng, Z.; Yang, Y.; Gan, H.; Lin, L.; Zhang, J. The Analysis of the Charred Plant Remains from Wanbei Site, Shuyang City, Jiangsu Province. Agric. Hist. China 2020, 39, 33–42. (In Chinese) [Google Scholar]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Altamira Press: Lanham, ML, USA, 2006; pp. 1–238. [Google Scholar]
- Zhang, J.; Lu, H.; Liu, M.; Diao, X.; Shao, K.; Wu, N. Phytolith Analysis for Differentiating between Broomcorn Millet (Panicum Miliaceum) and Its Weed/Feral Type (Panicum Ruderale). Sci. Rep. 2018, 8, 13022. [Google Scholar] [CrossRef]
- Ball, T.; Chandler-Ezell, K.; Dickau, R.; Duncan, N.; Hart, T.C.; Iriarte, J.; Lentfer, C.; Logan, A.; Lu, H.; Madella, M. Phytoliths as a Tool for Investigations of Agricultural Origins and Dispersals around the World. J. Archaeol. Sci. 2016, 68, 32–45. [Google Scholar] [CrossRef]
- Pearsall, D.M. Paleoethnobotany: A Handbook of Procedures; Left Coast Press: New York, NY, USA, 2015; pp. 1–700. [Google Scholar]
- Huan, X.; Lu, H.; Wang, C.; Tang, X.; Zuo, X.; Ge, Y.; He, K. Bulliform Phytolith Research in Wild and Domesticated Rice Paddy Soil in South China. PLoS ONE 2015, 10, e0141255. [Google Scholar] [CrossRef]
- Lu, H.; Wu, N.; Lu, B. Recognition of Rice Phytoliths. In Estado Actual de Los Estudios de Ftolitos En Suelos y Plantas (the Current State of Studies on Phytoliths in Soils and Plants). Monografas Del Centro de Ciencas Medioambientales 4; Pinilla, A., Juan-Tresserras, J., Machado, M.J., Eds.; Centro de Ciencas Medioambi Entales: Madrid, Spain, 1997; pp. 159–174. [Google Scholar]
- Gu, Y.; Zhao, Z.; Pearsall, D.M. Phytolith Morphology Research on Wild and Domesticated Rice Species in East Asia. Quat. Int. 2013, 287, 141–148. [Google Scholar] [CrossRef]
- Zhao, Z.; Pearsall, D.M.; Benfer, R.A., Jr.; Piperno, D.R. Distinguishing Rice (Oryza Sativa Poaceae) from Wild Oryza Species through Phytolith Analysis, II: Finalized Method. Econ. Bot. 1998, 52, 134–145. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, H.; Wu, N.; Yang, X.; Diao, X. Phytolith Analysis for Differentiating between Foxtail Millet (Setaria Italica) and Green Foxtail (Setaria Viridis). PLoS ONE 2011, 6, e19726. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lu, H.; Wu, N.; Li, F.; Yang, X.; Wang, W.; Ma, M.; Zhang, X. Phytolith Evidence of Millet Agriculture during about 6000–2100 a BP in the Guanzhong Basin, China. Quat. Sci. 2010, 30, 287–297. (In Chinese) [Google Scholar]
- Lu, H.; Zhang, J.; Wu, N.; Liu, K.; Xu, D.; Li, Q. Phytoliths Analysis for the Discrimination of Foxtail Millet (Setaria Italica) and Common Millet (Panicum Miliaceum). PLoS ONE 2009, 4, e4448. [Google Scholar] [CrossRef]
- Ge, H.; Chen, S.; Wang, L.; Wang, Z.; Zhang, J.; Ge, M. Investigation and Reflection on the Comprehensive Development and Utilization of Forest Resources in Shuyang County. Prot. For. Sci. Technol. 2008, 6, 36–38. (In Chinese) [Google Scholar]
- Nanjing Museum. Brief report on Excavation of Neolithic Remains at Wanbei site in Shuyang, Jiangsu Province. Southeast Cult. 1992, 01, 124–133. (In Chinese) [Google Scholar]
- Gu, J.; Yin, Z. Preliminary Results of Excavation of Wanbei Site in Shuyang, Jiangsu Province. Southeast Cult. 1988, 2, 49–50. (In Chinese) [Google Scholar]
- Lin, X.; Gan, H. Wanbei Site in Shuyang, Jiangsu Province. Pop. Archeol. 2016, 9, 12–13. (In Chinese) [Google Scholar]
- Tang, L.; Li, M.; Shen, C. Spore and Pollen Identification Report of Wanbei Site in Shuyang, Jiangsu Province. Southeast Cult. 1991, Z1, 190–192. (In Chinese) [Google Scholar]
- Li, M. Identification Report on Animal Skeletons of Wanbei Neolithic Site in Shuyang, Jiangsu Province. Southeast Cult. 1991, Z1, 183–189. (In Chinese) [Google Scholar]
- Piperno, D.R. Phytolith Analysis: An Archaeological and Geological Perspective; Academic Press: San Diego, CA, USA, 1990; pp. 1–280. [Google Scholar]
- Wang, Y.; Lv, H. The Study of Phytolith and Its Application; Ocean Press: Beijing, China, 1993; pp. 1–278. (In Chinese) [Google Scholar]
- Madella, M.; Alexandre, A.; Ball, T. International Code for Phytolith Nomenclature 1.0. Ann. Bot. 2005, 96, 253–260. [Google Scholar] [CrossRef]
- Reimer, P.J.; Austin, W.E.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 Cal KBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Luan, F.; Wagner, M. The Chronology and Basic Developmental Sequence of Archaeological Cultures in the Haidai Region. In Chinese Archaeology and Palaeoenvironment I: Prehistory at the Lower Reaches of the Yellow River, The Haidai Region; Luan, F., Ed.; Verlag Philipp von Zabern: Mainz, German, 2009; pp. 1–15. (In Chinese) [Google Scholar]
- Luan, F. Research on Dawenkou Culture (Dawenkou Wenhua yanjiu). In Haidaidiqu Kaogu Yanjiu (Archaeology Research of the Haidai Region); Luan, F., Ed.; Shandong University Press: Jinan, China, 1997; pp. 69–113. (In Chinese) [Google Scholar]
- Jin, G.; Wu, W.; Zhang, K.; Wang, Z.; Wu, X. 8000-Year Old Rice Remains from the North Edge of the Shandong Highlands, East China. J. Archaeol. Sci. 2014, 51, 34–42. [Google Scholar] [CrossRef]
- Crawford, G.W.; Chen, X.; Luan, F.; Wang, J. People and Plant Interaction at the Houli Culture Yuezhuang Site in Shandong Province, China. Holocene 2016, 26, 1594–1604. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, H.; Gu, W.; Wu, N.; Zhou, K.; Hu, Y.; Xin, Y.; Wang, C. Early Mixed Farming of Millet and Rice 7800 Years Ago in the Middle Yellow River Region, China. PLoS ONE 2012, 7, e52146. [Google Scholar] [CrossRef]
- Bestel, S.; Bao, Y.; Zhong, H.; Chen, X.; Liu, L. Wild Plant Use and Multi-Cropping at the Early Neolithic Zhuzhai Site in the Middle Yellow River Region, China. Holocene 2018, 28, 195–207. [Google Scholar] [CrossRef]
- Wang, C.; Lu, H.; Gu, W.; Zuo, X.; Zhang, J.; Liu, Y.; Bao, Y.; Hu, Y. Temporal Changes of Mixed Millet and Rice Agriculture in Neolithic-Bronze Age Central Plain, China: Archaeobotanical Evidence from the Zhuzhai Site. Holocene 2018, 28, 738–754. [Google Scholar] [CrossRef]
- Wang, C.; Lu, H.; Gu, W.; Wu, N.; Zhang, J.; Zuo, X.; Li, F.; Wang, D.; Dong, Y.; Wang, S. The Development of Yangshao Agriculture and Its Interaction with Social Dynamics in the Middle Yellow River Region, China. Holocene 2019, 29, 173–180. [Google Scholar] [CrossRef]
- Jin, G.; Wagner, M.; Tarasov, P.E.; Wang, F.; Liu, Y. Archaeobotanical Records of Middle and Late Neolithic Agriculture from Shandong Province, East China, and a Major Change in Regional Subsistence during the Dawenkou Culture. Holocene 2016, 26, 1605–1615. [Google Scholar] [CrossRef]
- Dai, J.; Cai, X.; Jin, J.; Ge, W.; Huang, Y.; Wu, W.; Xia, T.; Li, F.; Zuo, X. Earliest Arrival of Millet in the South China Coast Dating Back to 5,500 Years Ago. J. Archaeol. Sci. 2021, 129, 105356. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Zhang, G.; Zhang, G.; Wu, Y. The History of Agriculture in the Mountainous Areas of the Lower Yangtze River since the Late Neolithic. Front. Earth Sci. 2022, 16, 809–818. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, X.; Huan, X.; Gao, Y.; Wang, W.; Li, Z.; Ma, Z.; Perry, L.; Sun, G.; Jiang, L. Multiple Indicators of Rice Remains and the Process of Rice Domestication: A Case Study in the Lower Yangtze River Region, China. PLoS ONE 2018, 13, e0208104. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Crawford, G.W.; Jiang, L.; Chen, X. Rice Domestication Revealed by Reduced Shattering of Archaeological Rice from the Lower Yangtze Valley. Sci. Rep. 2016, 6, 28136. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Piperno, D.R. Late Pleistocene/Holocene Environments in the Middle Yangtze River Valley, China and Rice (Oryza Sativa L.) Domestication: The Phytolith Evidence. Geoarchaeology 2000, 15, 203–222. [Google Scholar] [CrossRef]
- Huan, X.; Lv, H.; Wang, C.; Zhang, J. Progress of Rice Bulliform Phytolith Research on Wild–Domesticated Characteristics. Acta Palaeontol. Sin. 2020, 59, 467–478. (In Chinese) [Google Scholar]
- Ma, Y.; Yang, X.; Huan, X.; Wang, W.; Ma, Z.; Li, Z.; Sun, G.; Jiang, L.; Zhuang, Y.; Lu, H. Rice Bulliform Phytoliths Reveal the Process of Rice Domestication in the Neolithic Lower Yangtze River Region. Quat. Int. 2016, 426, 126–132. [Google Scholar] [CrossRef]
- Qiu, Z. Some Thoughts on the Study of Rice Phytolith in Shangshan Culture. Archaeology 2021, 9, 109–120. (In Chinese) [Google Scholar]
- Yang, X.; Wan, Z.; Perry, L.; Lu, H.; Wang, Q.; Zhao, C.; Li, J.; Xie, F.; Yu, J.; Cui, T. Early Millet Use in Northern China. Proc. Natl. Acad. Sci. USA 2012, 109, 3726–3730. [Google Scholar] [CrossRef]
- Liu, C.; Kong, Z. Morphological Comparison of Foxtail Millet and Broomcorn Millet Seeds and Its Significance in Archaeological Identification. Archaeology 2004, 443, 76–83. (In Chinese) [Google Scholar]
- Liu, X.; Jones, P.J.; Matuzeviciute, G.M.; Hunt, H.V.; Lister, D.L.; An, T.; Przelomska, N.; Kneale, C.J.; Zhao, Z.; Jones, M.K. From Ecological Opportunism to Multi-Cropping: Mapping Food Globalisation in Prehistory. Quat. Sci. Rev. 2019, 206, 21–28. [Google Scholar] [CrossRef]
- Jones, H.; Lister, D.L.; Cai, D.; Kneale, C.J.; Cockram, J.; Peña-Chocarro, L.; Jones, M.K. The Trans-Eurasian Crop Exchange in Prehistory: Discerning Pathways from Barley Phylogeography. Quat. Int. 2016, 426, 26–32. [Google Scholar] [CrossRef]
- Fuller, D.Q. Finding Plant Domestication in the Indian Subcontinent. Curr. Anthropol. 2011, 52, S347–S362. [Google Scholar] [CrossRef]
- Li, H.; Cui, Y.; James, N.; Ritchey, M.; Liu, F.; Zhang, J.; Ma, M.; Dong, G. Spatiotemporal Variation of Agricultural Patterns in Different Geomorphologic and Climatic Environments in the Eastern Loess Plateau, North-Central China during the Late Neolithic and Bronze Ages. Sci. China Earth Sci. 2022, 65, 934–948. [Google Scholar] [CrossRef]
- Tang, L.; Shen, C.; Zhao, X.; Xiao, J.; Yu, G.; Han, H. Vegetation and Climate in Qingfeng Section of Jianhu, Jiangsu Province in the Past 10 thousand years. Sci. China (Ser. B) 1993, 23, 637–643. (In Chinese) [Google Scholar]
- Qin, X.; Zhang, L.; Mu, Y. The Holocene Climatic Changes of the Huaihe River Semi-humid Region in the North and South Transition Zone of the Eastern China. Quat. Sci. 2015, 35, 1509–1524. (In Chinese) [Google Scholar]
- Qiu, Z.; Jiang, H.; Ding, L.; Shang, X. Late Pleistocene-Holocene Vegetation History and Anthropogenic Activities Deduced from Pollen Spectra and Archaeological Data at Guxu Lake, Eastern China. Sci. Rep. 2020, 10, 9306. [Google Scholar] [CrossRef]
- Jiang, S.; Luo, W.; Tu, L.; Yu, Y.; Fang, F.; Liu, X.; Zhan, T.; Fang, L.; Zhang, X.; Zhou, X. The Holocene Optimum (HO) and the Response of Human Activity: A Case Study of the Huai River Basin in Eastern China. Quat. Int. 2018, 493, 31–38. [Google Scholar] [CrossRef]
- Crawford, G.; Underhill, A.; Zhao, Z.; Lee, G.; Feinman, G.; Nicholas, L.; Luan, F.; Yu, H.; Fang, H.; Cai, F. Late Neolithic Plant Remains from Northern China: Preliminary Results from Liangchengzhen, Shandong. Curr. Anthropol. 2005, 46, 309–317. [Google Scholar] [CrossRef]
- Sun, B.; Wagner, M.; Zhao, Z.; Li, G.; Wu, X.; Tarasov, P.E. Archaeological Discovery and Research at Bianbiandong Early Neolithic Cave Site, Shandong, China. Quat. Int. 2014, 348, 169–182. [Google Scholar] [CrossRef]
- Wu, W.; Wang, X.; Wu, X.; Jin, G.; Tarasov, P.E. The Early Holocene Archaeobotanical Record from the Zhangmatun Site Situated at the Northern Edge of the Shandong Highlands, China. Quat. Int. 2014, 348, 183–193. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, W.; Luo, W.; Yang, Y.; Gan, H.; Cheng, Z.; Sun, Y.; Zhang, D.; Lin, L.; Zhang, J. Multicropping Pattern Reveals Human Adaptation at the Wanbei Site (ca. 5.7–4.4 ka cal. BP) in the Middle and Lower Huai River Valley, China. Land 2023, 12, 1158. https://doi.org/10.3390/land12061158
Tian W, Luo W, Yang Y, Gan H, Cheng Z, Sun Y, Zhang D, Lin L, Zhang J. Multicropping Pattern Reveals Human Adaptation at the Wanbei Site (ca. 5.7–4.4 ka cal. BP) in the Middle and Lower Huai River Valley, China. Land. 2023; 12(6):1158. https://doi.org/10.3390/land12061158
Chicago/Turabian StyleTian, Weixin, Wuhong Luo, Yuzhang Yang, Huiyuan Gan, Zhijie Cheng, Yajie Sun, Dailing Zhang, Liugen Lin, and Juzhong Zhang. 2023. "Multicropping Pattern Reveals Human Adaptation at the Wanbei Site (ca. 5.7–4.4 ka cal. BP) in the Middle and Lower Huai River Valley, China" Land 12, no. 6: 1158. https://doi.org/10.3390/land12061158