
Where Will Threatened Aegle marmelos L., a Tree of the Semi-Arid Region, Go under Climate Change? Implications for the Reintroduction of the Species

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
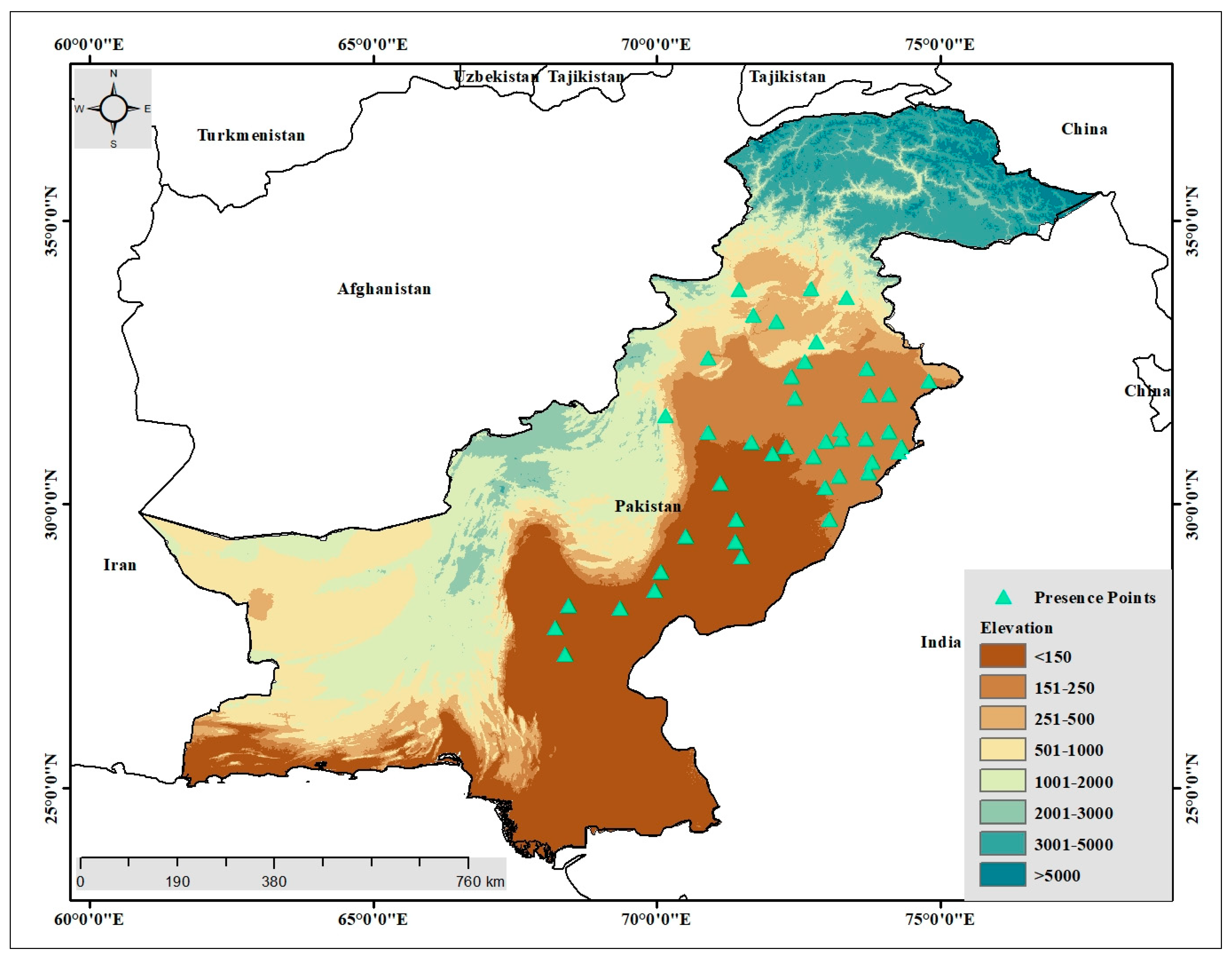
2.1. Aegle Marmelos and Occurrence Data
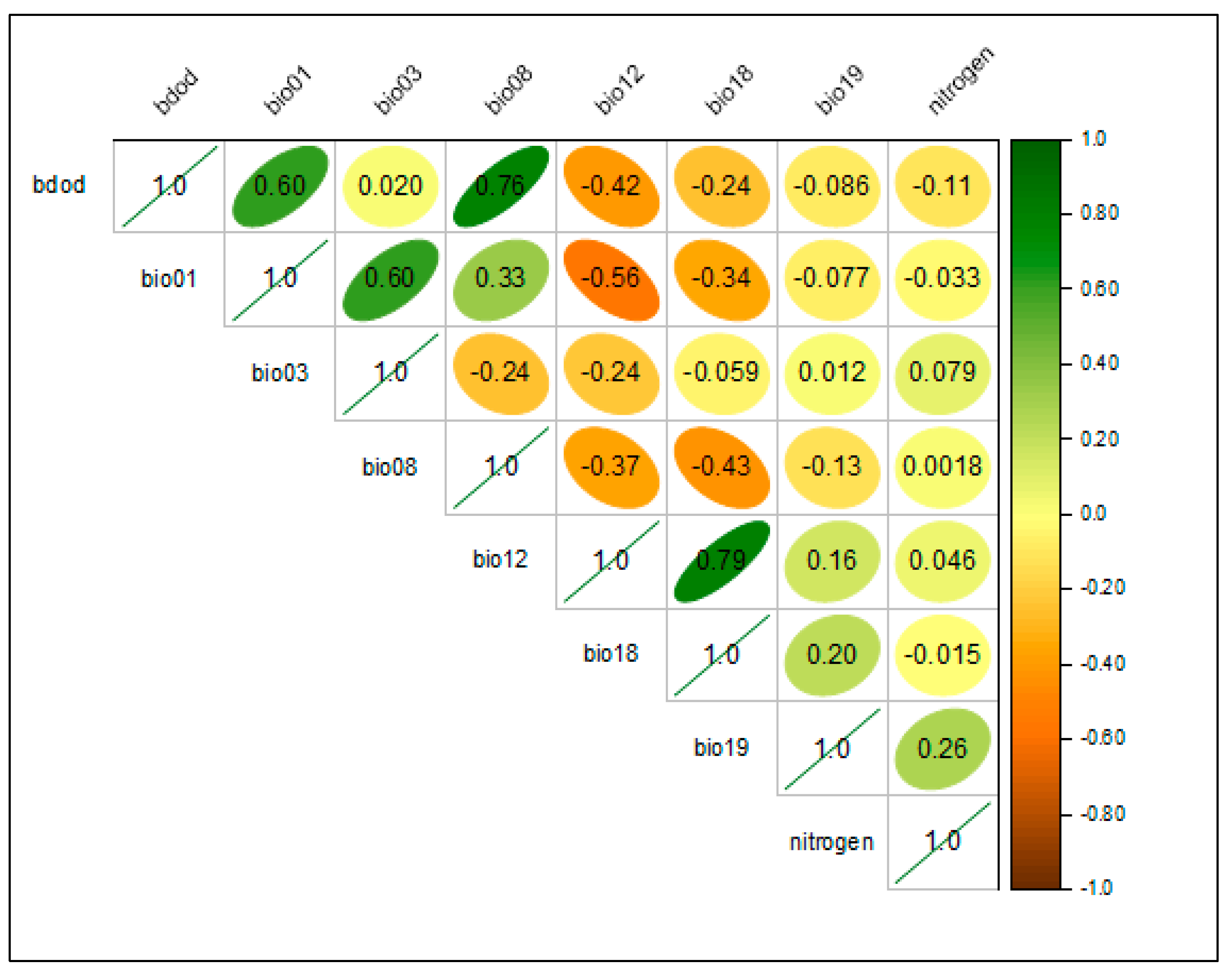
2.2. Environmental Data Collection and Variable Selection
2.3. Preliminary Variables Processing
2.4. Model Calibration and Optimization
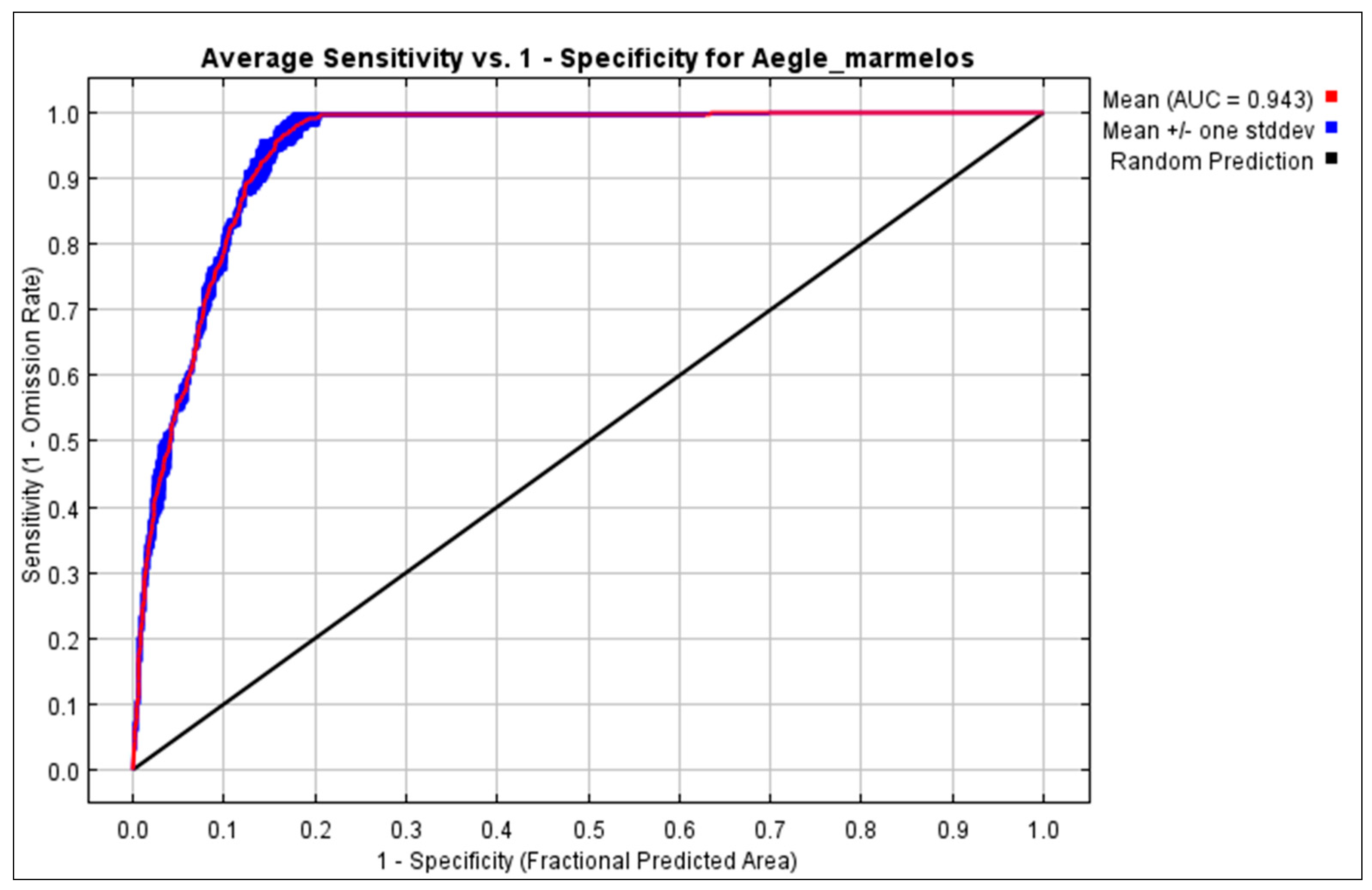
2.5. Reclassification of Predictions and Model Evaluation
3. Results
3.1. Key Environmental Variables
3.2. Current Distribution
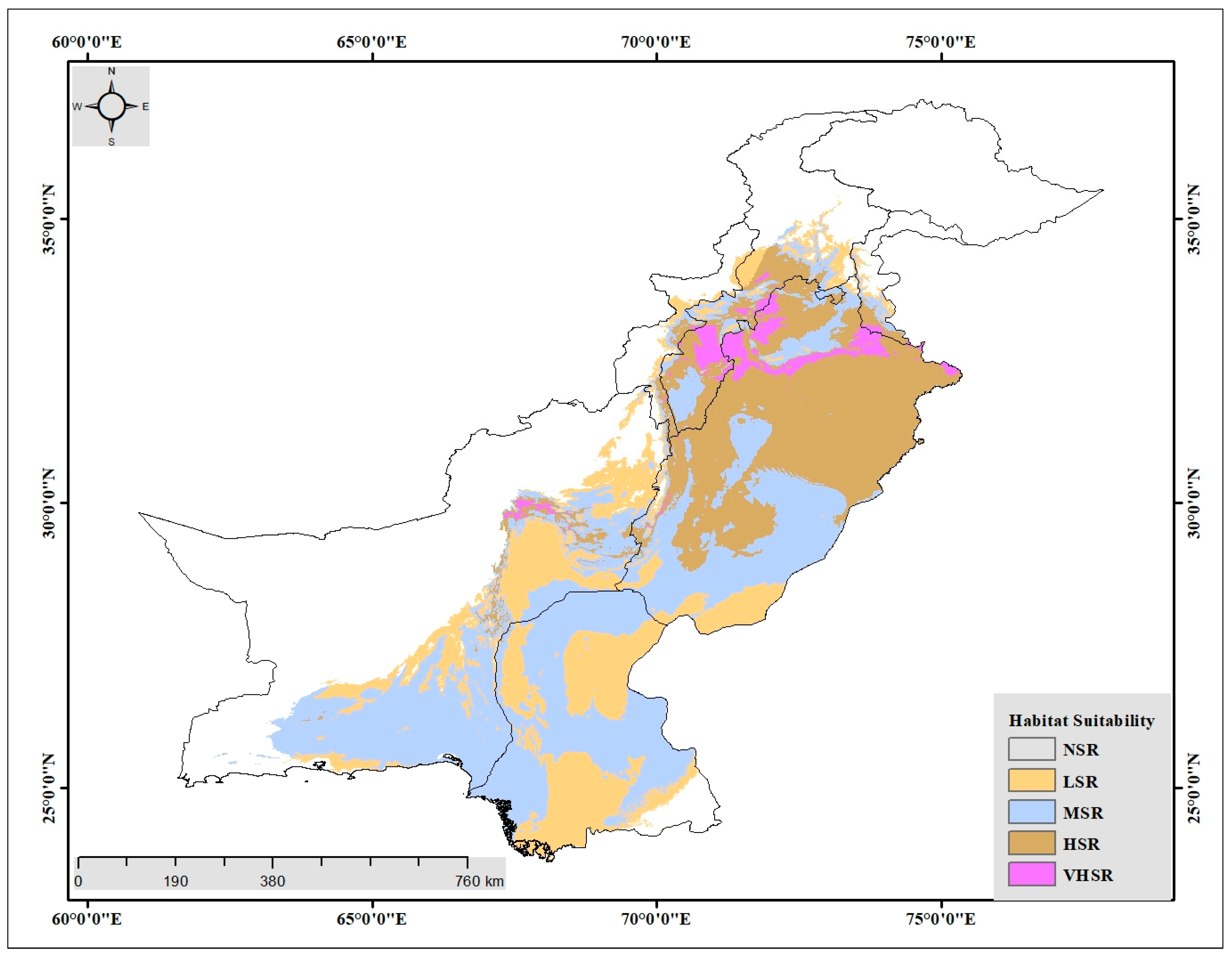
3.3. Potential Habitat Suitability under the Future Climate Change Scenario
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamann, E.; Blevins, C.; Franks, S.J.; Jameel, M.I.; Anderson, J.T. Climate change alters plant–herbivore interactions. New Phytol. 2021, 229, 1894–1910. [Google Scholar] [CrossRef] [PubMed]
- Elsen, P.R.; Saxon, E.C.; Simmons, B.A.; Ward, M.; Williams, B.A.; Grantham, H.S.; Kark, S.; Levin, N.; Perez-Hammerle, K.V.; Reside, A.E.; et al. Accelerated shifts in terrestrial life zones under rapid climate change. Glob. Change Biol. 2022, 28, 918–935. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.M.; Waheed, M.; Ahmad, R.; Bussmann, R.W.; Arshad, F.; Khan, A.M.; Casini, R.; Alataway, A.; Dewidar, A.Z.; Elansary, H.O. Climate change and human activities, the significant dynamic drivers of Himalayan Goral Distribution (Naemorhedus goral). Biology 2023, 12, 610. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; He, H.S.; Liang, Y.; Hawbaker, T.J.; Henne, P.D.; Xu, W.; Gong, P.; Zhu, Z. The changes in species composition mediate direct effects of climate change on future fire regimes of boreal forests in northeastern China. J. Appl. Ecol. 2021, 58, 1336–1345. [Google Scholar] [CrossRef]
- Schmeller, D.S.; Courchamp, F.; Killeen, G. Biodiversity loss, emerging pathogens and human health risks. Biodivers. Conserv. 2020, 29, 3095–3102. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Z.; Abdukeyum, N.; Ling, Y. Potential geographical distribution of medicinal plant Ephedra sinica Stapf. under climate change. Forests 2022, 13, 2149. [Google Scholar] [CrossRef]
- Zhang, Y.; Tariq, A.; Hughes, A.C.; Hong, D.; Wei, F.; Sun, H.; Sardans, J.; Peñuelas, J.; Perry, G.; Qiao, J.; et al. Challenges and solutions to biodiversity conservation in arid lands. Sci. Total Environ. 2023, 857, 159695. [Google Scholar] [CrossRef]
- Williams, J.N.; Seo, C.; Thorne, J.; Nelson, J.K.; Erwin, S.; O’Brien, J.M.; Schwartz, M.W. Using species distribution models to predict new occurrences for rare plants. Divers. Distrib. 2009, 15, 565–576. [Google Scholar] [CrossRef]
- Briscoe, N.J.; Elith, J.; Salguero-Gómez, R.; Lahoz-Monfort, J.J.; Camac, J.S.; Giljohann, K.M.; Holden, M.H.; Hradsky, B.A.; Kearney, M.R.; McMahon, S.M.; et al. Forecasting species range dynamics with process-explicit models: Matching methods to applications. Ecol. Lett. 2019, 22, 1940–1956. [Google Scholar] [CrossRef] [Green Version]
- Jinga, P.; Liao, Z.; Nobis, M.P. Species distribution modeling that overlooks intraspecific variation is inadequate for proper conservation of marula (Sclerocarya birrea, Anacardiaceae). Glob. Ecol. Conserv. 2021, 32, e01908. [Google Scholar] [CrossRef]
- Smith, J.N.; Kelly, N.; Renner, I.W. Validation of presence-only models for conservation planning and the application to whales in a multiple-use marine park. Ecol. Appl. 2021, 31, e02214. [Google Scholar] [CrossRef]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CABI Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
- Waheed, M.; Arshad, F.; Majeed, M.; Haq, S.M.; Aziz, R.; Bussmann, R.W.; Ali, K.; Subhan, F.; Jones, D.A.; Zaitouny, A. Potential distribution of a noxious weed (Solanum viarum Dunal), current status, and future invasion risk based on MaxEnt modeling. Geol. Ecol. Landsc. 2023, 12, 1–16. [Google Scholar] [CrossRef]
- Stewart, S.B.; Fedrigo, M.; Kasel, S.; Roxburgh, S.H.; Choden, K.; Tenzin, K.; Allen, K.; Nitschke, C.R. Predicting plant species distributions using climate-based model ensembles with corresponding measures of congruence and uncertainty. Divers. Distrib. 2022, 28, 1105–1122. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Chauvier, Y.; Descombes, P.; Guéguen, M.; Boulangeat, L.; Thuiller, W.; Zimmermann, N.E. Resolution in species distribution models shapes spatial patterns of plant multifaceted diversity. Ecography 2022, 10, e05973. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Stanton-Geddes, J. Climate change, genetic markers and species distribution modelling. J. Biogeogr. 2015, 42, 1577–1585. [Google Scholar] [CrossRef]
- Mod, H.K.; Scherrer, D.; Luoto, M.; Guisan, A. What we use is not what we know: Environmental predictors in plant distribution models. J. Veg. Sci. 2016, 27, 1308–1322. [Google Scholar] [CrossRef] [Green Version]
- Falk, W.; Mellert, K.H. Species distribution models as a tool for forest management planning under climate change: Risk evaluation of Abies alba in Bavaria. J. Veg. Sci. 2011, 22, 621–634. [Google Scholar] [CrossRef]
- Grenouillet, G.; Buisson, L.; Casajus, N.; Lek, S. Ensemble modelling of species distribution: The effects of geographical and environmental ranges. Ecography 2011, 34, 9–17. [Google Scholar] [CrossRef]
- Arshad, F.; Waheed, M.; Fatima, K.; Harun, N.; Iqbal, M.; Fatima, K.; Umbreen, S. Predicting the suitable current and future potential distribution of the native endangered tree Tecomella undulata (Sm.) Seem. in Pakistan. Sustainability 2022, 14, 7215. [Google Scholar] [CrossRef]
- Albert, C.H.; Rayfield, B.; Dumitru, M.; Gonzalez, A. Applying network theory to prioritize multispecies habitat networks that are robust to climate and land-use change. Biol. Conserv. 2017, 31, 1383–1396. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Feijoo, R.J.; Torres, B.; López-Tobar, R.; Tipán-Torres, C.; Toulkeridis, T.; Heredia-R, M.; Mateo, R.G. Modelling climatically suitable areas for Mahogany (Swietenia macrophylla King) and their shifts across Neotropics: The role of Protected Areas. Forests 2023, 14, 385. [Google Scholar] [CrossRef]
- Ferraz, K.M.P.M.D.B.; Morato, R.G.; Bovo, A.A.A.; da Costa, C.O.R.; Ribeiro, Y.G.G.; de Paula, R.C.; Desbiez, A.L.J.; Angelieri, C.S.C.; Traylor-Holzer, K. Bridging the gap between researchers, conservation planners, and decision makers to improve species conservation decision-making. Conserv. Sci. Pract. 2021, 3, e330. [Google Scholar] [CrossRef]
- UCN. The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org/ (accessed on 10 December 2022).
- Hassan-ud-Din, G.; Ghazanfar, S. Rutaceae. Flora Pak. 1980, 132, 10–15. [Google Scholar]
- Kress, W.J.; DeFilipps, R.A.; Farr, E.; Kyi, D.Y.Y. A checklist of the trees, shrubs, herbs, and climbers of Myanmar. Natl. Herbarium 2003, 45, 1–590. [Google Scholar]
- Pandey, R.P.; Diwakar, P.O. An integrated check-list flora of Andaman and Nicobar Islands, India. J. Econ. Taxon. Bot. 2008, 32, 403–500. [Google Scholar]
- Neeraj, V.B.; Johar, V. Bael (Aegle marmelos) extraordinary species of India: A review. Int. J. Curr. Microbiol. Appl. 2017, 6, 1870–1887. [Google Scholar]
- Vasava, D.; Kher, M.M.; Nataraj, M.; Teixeira da Silva, J.A. Bael tree (Aegle marmelos (L.) Corrêa): Importance, biology, propagation, and future perspectives. Trees 2018, 32, 1165–1198. [Google Scholar] [CrossRef]
- Waheed, M.; Arshad, F.; Majeed, M.; Fatima, S.; Mukhtar, N.; Aziz, R.; Mangrio, W.M.; Almohamad, H.; Al Dughairi, A.A.; Al-Mutiry, M.; et al. Community structure and distribution pattern of woody vegetation in response to soil properties in semi-arid lowland District Kasur Punjab, Pakistan. Land 2022, 11, 2145. [Google Scholar] [CrossRef]
- Pande, H.K.; Arora, S. India’s Fifth National Report to the Convention on Biological Diversity; Ministry of Environment and Forests, Government of India, New Delhi: Delhi, India, 2014; Volume 142. [Google Scholar]
- Porfirio, L.L.; Harris, R.M.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.L.; Mackey, B. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, V.; Singh, R.; Pandey, A.K. Aegle marmelos: Pharmacological, medicinal importance, and conservation in India. J. Exp. Zool. India 2018, 21, 1–9. [Google Scholar]
- Rahman, S.; Parvin, R. Therapeutic potential of Aegle marmelos (L.)—An overview. Asian Pac. J. Trop. Dis. 2014, 4, 71–77. [Google Scholar] [CrossRef]
- Maity, P.; Hansda, D.; Bandyopadhyay, U.; Mishra, D.K. Biological activities of crude extracts and chemical constituents of Bael, Aegle marmelos (L.) Corr. Indian J. Exp. Biol. 2009, 47, 849–861. [Google Scholar]
- Mujeeb, F.; Bajpai, P.; Pathak, N. Phytochemical evaluation, antimicrobial activity, and determination of bioactive components from leaves of Aegle marmelos. BioMed. Res. Int. 2014, 2014, 497606. [Google Scholar] [CrossRef] [Green Version]
- Rajan, S.; Gokila, M.; Jency, P.; Brindha, P.; Sujatha, R.K. Antioxidant and phytochemical properties of Aegle marmelos fruit pulp. Int. J. Curr. Pharm. Res. 2011, 3, 65–70. [Google Scholar]
- Waheed, M.; Haq, S.M.; Arshad, F.; Bussmann, R.W.; Iqbal, M.; Bukhari, N.A.; Hatamleh, A.A. Grasses in semi-arid lowlands—Community composition and spatial dynamics with special regard to the influence of edaphic factors. Sustainability 2022, 14, 14964. [Google Scholar] [CrossRef]
- Bosso, L.; Di Febbraro, M.; Cristinzio, G.; Zoina, A.; Russo, D. Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean basin. Biol. Invasions 2016, 18, 1759–1768. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. A J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.P.; Gonzalez, I., Jr. Species-specific tuning increases robustness to sampling bias in models of species distributions: An implementation with Maxent. Ecol. Modell. 2011, 222, 2796–2811. [Google Scholar] [CrossRef]
- Bao, R.; Li, X.; Zheng, J. Feature tuning improves MAXENT predictions of the potential distribution of Pedicularis longiflora Rudolph and its variant. PeerJ 2022, 10, e13337. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Bai, D.F.; Chen, P.J.; Atzeni, L.; Cering, L.; Li, Q.; Shi, K. Assessment of habitat suitability of the snow leopard (Panthera uncia) in Qomolangma National Nature Reserve based on MaxEnt modeling. Zool. Res. 2018, 39, 373. [Google Scholar] [PubMed]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Summers, D.M.; Bryan, B.A.; Crossman, N.D.; Meyer, W.S. Species vulnerability to climate change: Impacts on spatial conservation priorities and species representation. Glob. Change Biol. 2012, 18, 2335–2348. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Diver. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Gassó, N.; Thuiller, W.; Pino, J.; Vilà, M. Potential distribution range of invasive plant species in Spain. NeoBiota 2012, 12, 25. [Google Scholar]
- Khan, A.M.; Li, Q.; Saqib, Z.; Khan, N.; Habib, T.; Khalid, N.; Majeed, M.; Tariq, A. MaxEnt modelling and impact of climate change on habitat suitability variations of economically important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests 2022, 13, 715. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papes, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Chu, J.; Li, Y.; Zhang, L.; Li, B.; Gao, M.; Tang, X.; Ni, J.; Xu, X. Potential distribution range and conservation strategies for the endangered species Amygdalus pedunculata. Biodiver. Sci. 2017, 25, 799. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.L.; He, Y.L.; Deng, F.Y.; Li, T.Y.; Yu, L. Assessment of alpine mean response to climate change in Southwest China based on MaxEnt model. Acta Ecol. Sin. 2019, 39, 9033–9043. [Google Scholar]
- Venne, S.; Currie, D.J. Can habitat suitability estimated from MaxEnt predict colonizations and extinctions? Divers. Distrib. 2021, 27, 873–886. [Google Scholar] [CrossRef]
- Zengli, L.I.U.; Lile, H.U. Prediction of potential distribution and climate change of rare species Cephalotaxus oliveri. For. Resour. Manag. 2022, 1, 35. [Google Scholar]
- Cotto, O.; Wessely, J.; Georges, D.; Klonner, G.; Schmid, M.; Dullinger, S.; Thuiller, W.; Guillaume, F. A dynamic eco-evolutionary model predicts slow response of alpine plants to climate warming. Nat. Commun. 2017, 8, 15399. [Google Scholar] [CrossRef] [Green Version]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Midolo, G.; Wellstein, C. Plant performance and survival across transplant experiments depend upon temperature and precipitation change along elevation. J. Ecol. 2020, 108, 2107–2120. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2022, 41, 1–31. [Google Scholar] [CrossRef]
- Li, Y.; Shao, W.; Jiang, J. Predicting the potential global distribution of Sapindus mukorossi under climate change based on MaxEnt modelling. Environ. Sci. Poll. Res. 2021, 29, 21751–21768. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Tang, L.; He, H.; Yang, F.; Tao, J.; Wang, W. Assessing the impact of climate change on the distribution of Osmanthus fragrans using Maxent. Environ. Sci. Poll. Res. 2021, 28, 34655–34663. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Li, J.; Wu, S. MaxEnt Modeling to estimate the impact of climate factors on distribution of Pinus densiflora. Forests 2022, 13, 402. [Google Scholar] [CrossRef]
- Thakur, K.K.; Bhat, P.; Kumar, A.; Ravikanth, G.; Saikia, P. Distribution mapping of Bauhinia vahlii Wight & Arn. in India using ecological niche modelling. Trop. Ecol. 2022, 63, 286–299. [Google Scholar]
- Austin, M.P.; Van Niel, K.P. Improving species distribution models for climate change studies: Variable selection and scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Wu, B.; Zhou, L.; Qi, S.; Jin, M.; Hu, J.; Lu, J. Effect of habitat factors on the understory plant diversity of Platycladus orientalis plantations in Beijing mountainous areas based on MaxEnt model. Ecol. Indic. 2021, 129, 107917. [Google Scholar] [CrossRef]
- Cantón, Y.; Del Barrio, G.; Solé-Benet, A.; Lázaro, R. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain. Catena 2004, 55, 341–365. [Google Scholar] [CrossRef]
- Sharath, A.A.; Ghosh, S.N.; Das, B.C. Integrated nutrient management in bael (Aegle marmelos Corr.) in New Alluvial soil. IJMFM AP 2016, 2, 41–46. [Google Scholar]
- Rajpoot, R.; Adhikari, D.; Verma, S.; Saikia, P.; Kumar, A.; Grant, K.R.; Dayanandan, A.; Kumar, A.; Khare, P.K.; Khan, M.L. Climate models predict a divergent future for the medicinal tree Boswellia serrata Roxb. in India. Glob. Ecol. Conserv. 2020, 23, e01040. [Google Scholar] [CrossRef]
- Priti, H.; Aravind, N.A.; Shaanker, R.U.; Ravikanth, G. Modeling impacts of future climate on the distribution of Myristicaceae species in the Western Ghats, India. Ecol. Eng. 2016, 89, 14–23. [Google Scholar] [CrossRef]
- Ahmad, R.; Khuroo, A.A.; Hamid, M.; Charles, B.; Rashid, I. Predicting invasion potential and niche dynamics of Parthenium hysterophorus (Congress grass) in India under projected climate change. Biodiv. Conserv. 2019, 28, 2319–2344. [Google Scholar] [CrossRef]
- Purohit, S.; Rawat, N. MaxEnt modeling to predict the current and future distribution of Clerodendrum infortunatum L. under climate change scenarios in Dehradun district, India. Model. Earth Sys. Environ. 2022, 8, 2051–2063. [Google Scholar] [CrossRef]
- Mahmoodi, S.; Heydari, M.; Ahmadi, K.; Khwarahm, N.R.; Karami, O.; Almasieh, K.; Naderi, B.; Bernard, P.; Mosavi, A. The current and future potential geographical distribution of Nepeta crispa Willd., an endemic, rare and threatened aromatic plant of Iran: Implications for ecological conservation and restoration. Ecol. Indic. 2022, 137, 108752. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Code | Percent Contribution |
|---|---|---|
| Bulk Density | bdod | 74.3 |
| Isothermality (Bio2/Bio7) (×100) | Bio03 | 9.2 |
| Precipitation of the Warmest Quarter | Bio18 | 6.9 |
| Mean Temperature of the Wettest Quarter | Bio08 | 5.2 |
| Precipitation of the Coldest Quarter | Bio19 | 4.2 |
| Annual Mean Temperature | Bio01 | 1 |
| Annual Precipitation | Bio12 | 1.7 |
| Total Nitrogen | nitrogen | 1.5 |
| Climate Change Scenario | NS | LS | MS | HS | VHS | Total Suitable Land Area (km2) |
|---|---|---|---|---|---|---|
| (p ≤ 0.2) | (p 0.21–0.4) | (p 0.41–0.6) | (p 0.61–0.8) | (p ≥ 0.81) | ||
| Current | 484,918 | 128,961 | 102,161 | 110,455 | 55,292 | 396,869 |
| SSPs_245_2050 | 515,774 | 121,145 | 94,268 | 94,430 | 56,170 | 366,013 |
| Rate of change (%) | 3.5 | −1.3 | −0.3 | −2.3 | 0.4 | −3.5 |
| SSPs_585_2050 | 538,774 | 131,145 | 85,268 | 84,430 | 52,170 | 343,013 |
| Rate of change (%) | 6.1 | −2.3 | −1.3 | −3.4 | −0.1 | −6.1 |
| SSPs_245_2070 | 526,774 | 118,045 | 92,568 | 90,230 | 54,170 | 355,013 |
| Rate of change (%) | 4.7 | −1.6 | −0.5 | −2.8 | 0.1 | −4.7 |
| SSPs_585_2070 | 555,774 | 101,145 | 88,268 | 87,430 | 49,170 | 326,013 |
| Rate of change (%) | 8.0 | −3.5 | −1.0 | −3.1 | −0.4 | −8.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waheed, M.; Haq, S.M.; Arshad, F.; Jameel, M.A.; Siddiqui, M.H.; Bussmann, R.W.; Manshoor, N.; Alamri, S. Where Will Threatened Aegle marmelos L., a Tree of the Semi-Arid Region, Go under Climate Change? Implications for the Reintroduction of the Species. Land 2023, 12, 1433. https://doi.org/10.3390/land12071433
Waheed M, Haq SM, Arshad F, Jameel MA, Siddiqui MH, Bussmann RW, Manshoor N, Alamri S. Where Will Threatened Aegle marmelos L., a Tree of the Semi-Arid Region, Go under Climate Change? Implications for the Reintroduction of the Species. Land. 2023; 12(7):1433. https://doi.org/10.3390/land12071433
Chicago/Turabian StyleWaheed, Muhammad, Shiekh Marifatul Haq, Fahim Arshad, Muhammad Azhar Jameel, Manzer H. Siddiqui, Rainer W. Bussmann, Nabeel Manshoor, and Saud Alamri. 2023. "Where Will Threatened Aegle marmelos L., a Tree of the Semi-Arid Region, Go under Climate Change? Implications for the Reintroduction of the Species" Land 12, no. 7: 1433. https://doi.org/10.3390/land12071433