
A 20-Year Ecotone Study of Pacific Northwest Mountain Forest Vulnerability to Changing Snow Conditions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
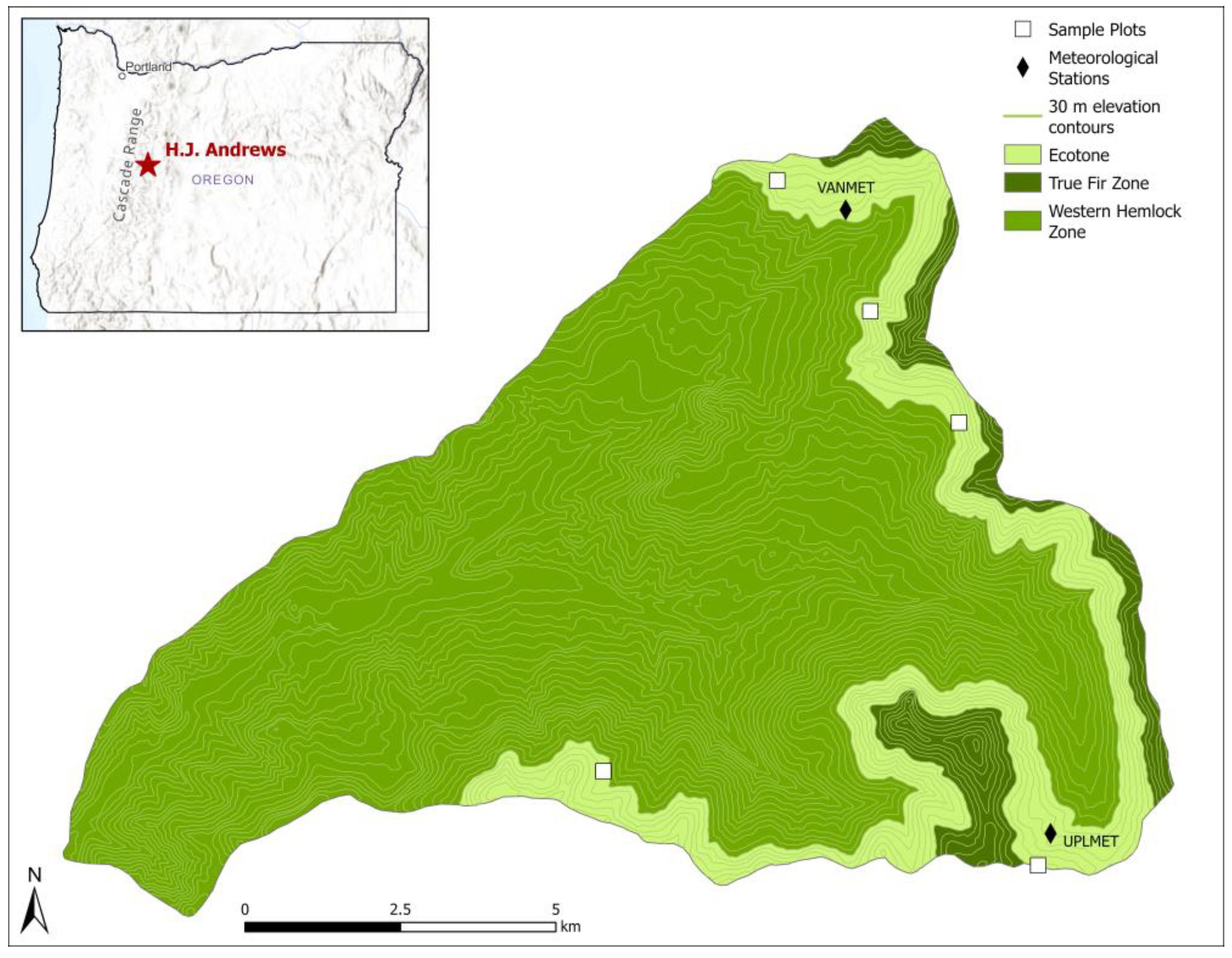
2.1. Study Area
2.2. Data
2.3. Analyses
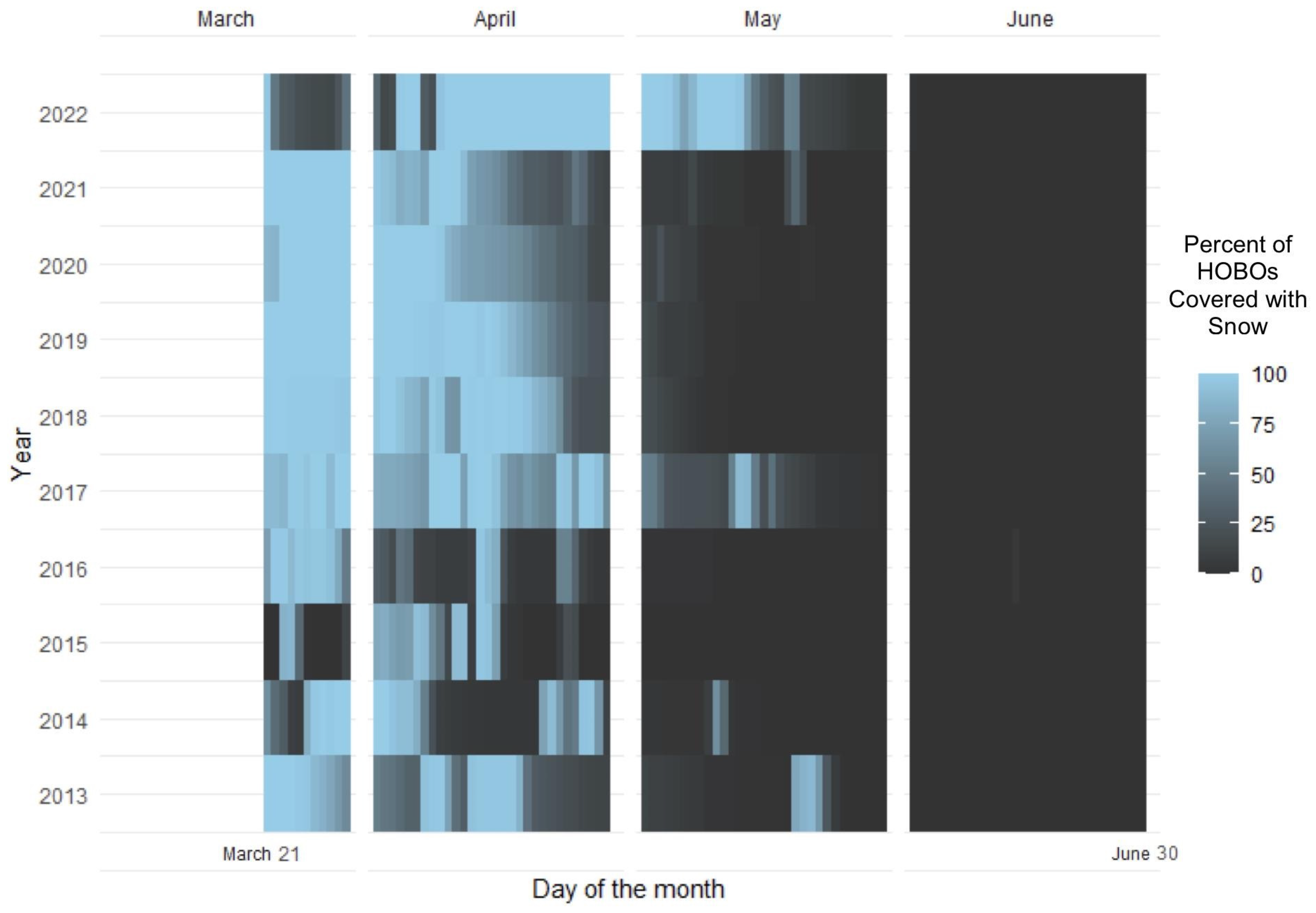
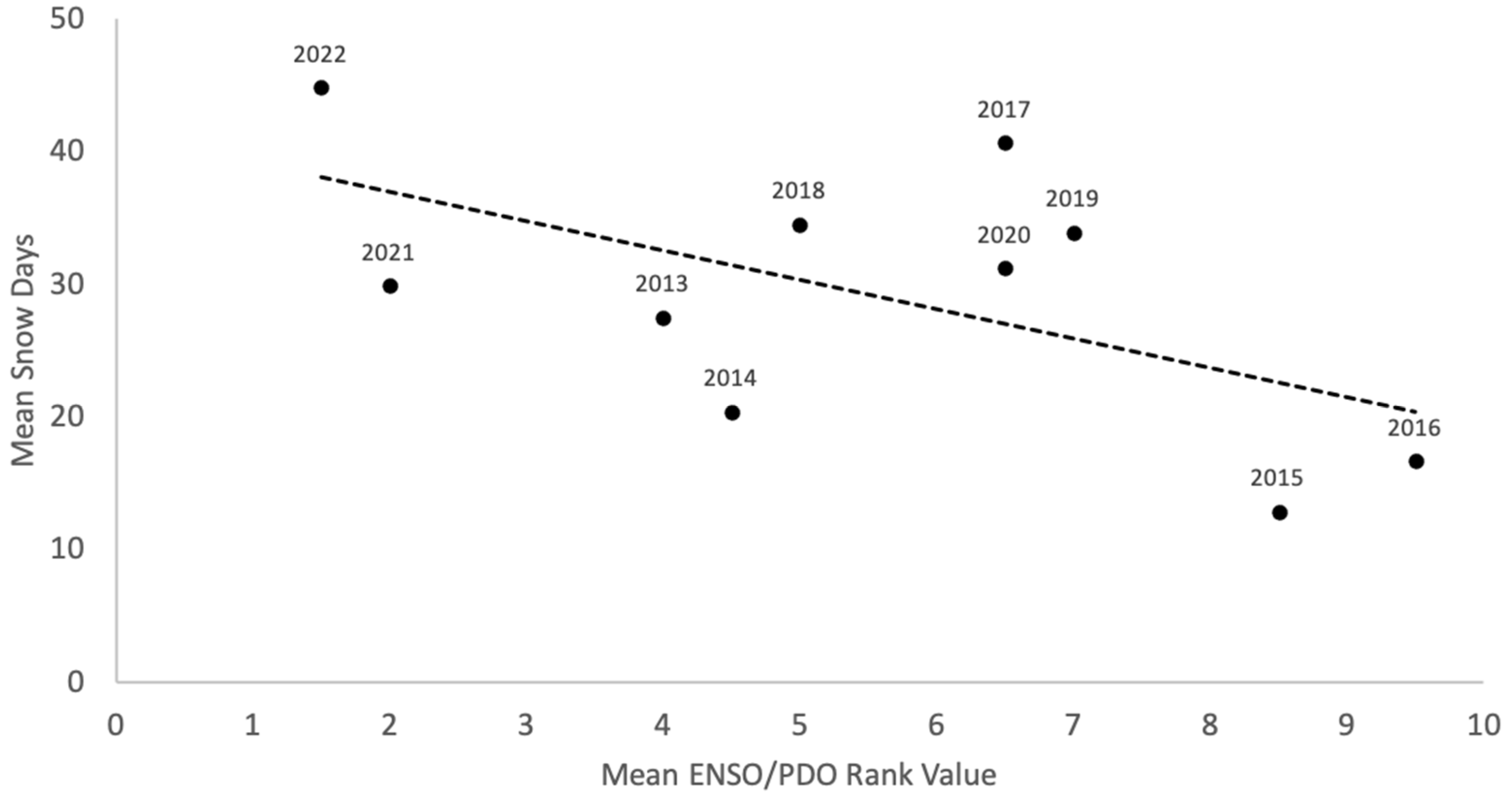
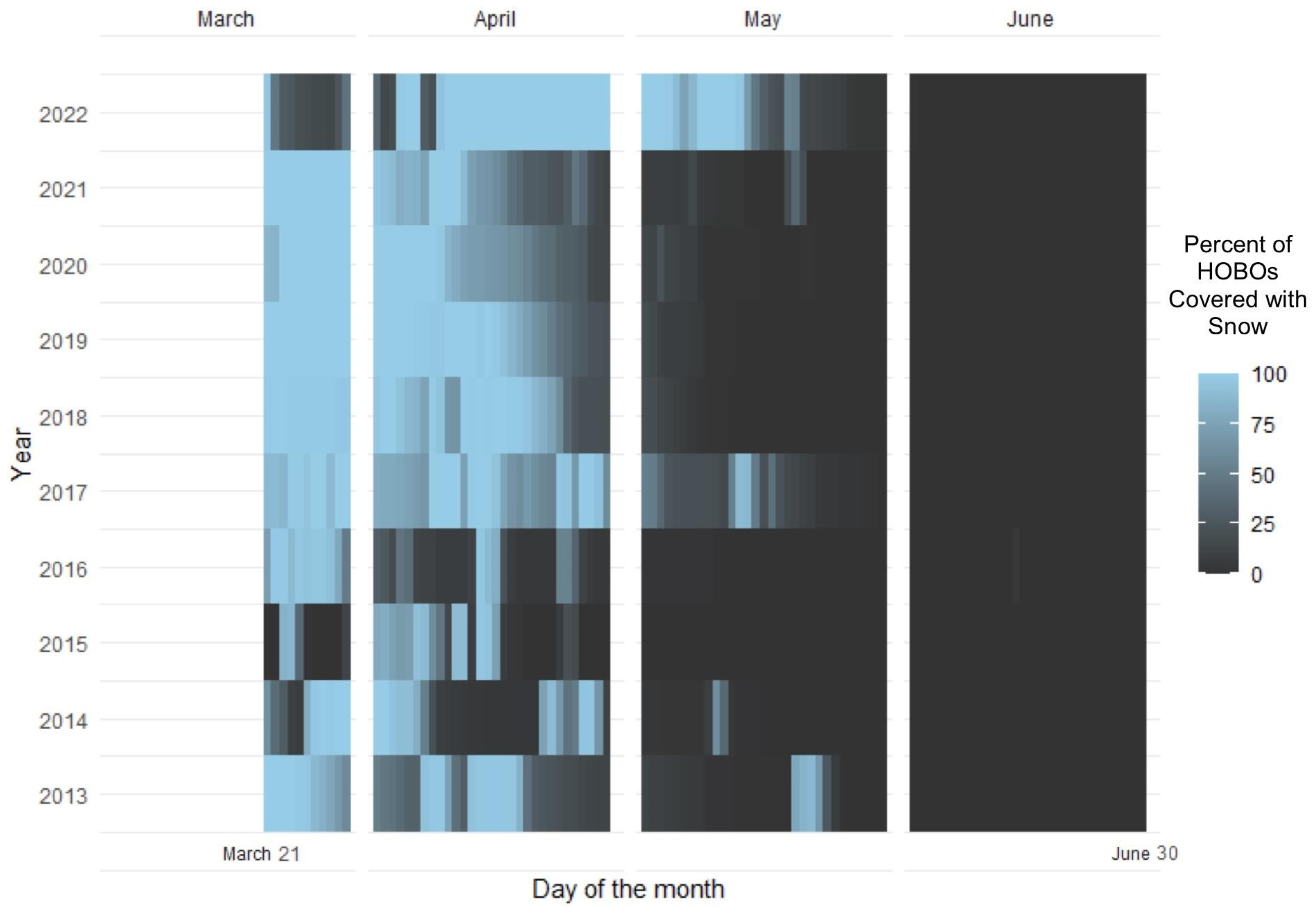
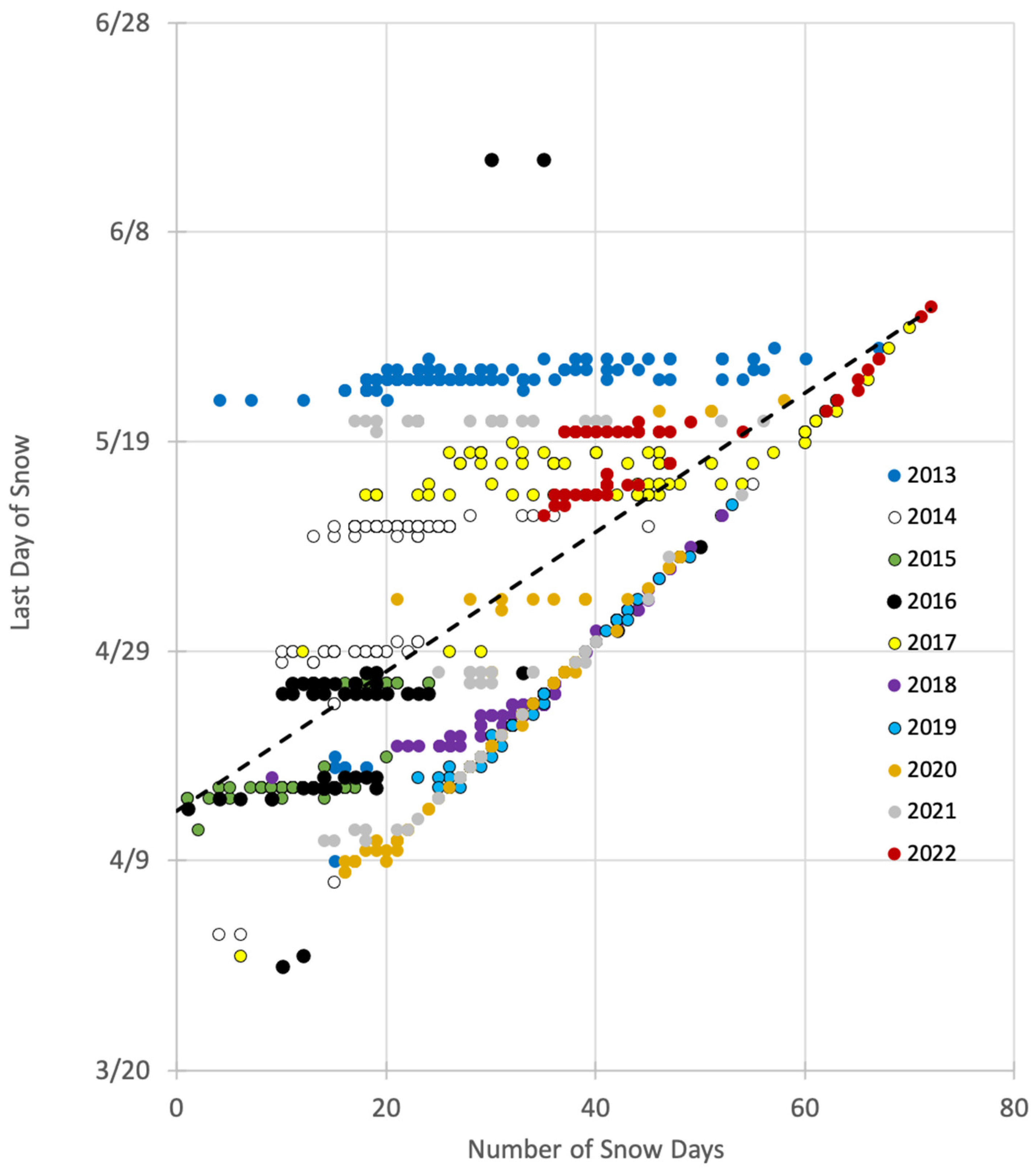
2.3.1. Snow Variability
- Number of snow days—the total number of days during the snow depletion season that the data logger was covered by snow;
- Last day of snow—the last day of the water year (which ends on 30 September) in which the data logger was covered by snow.
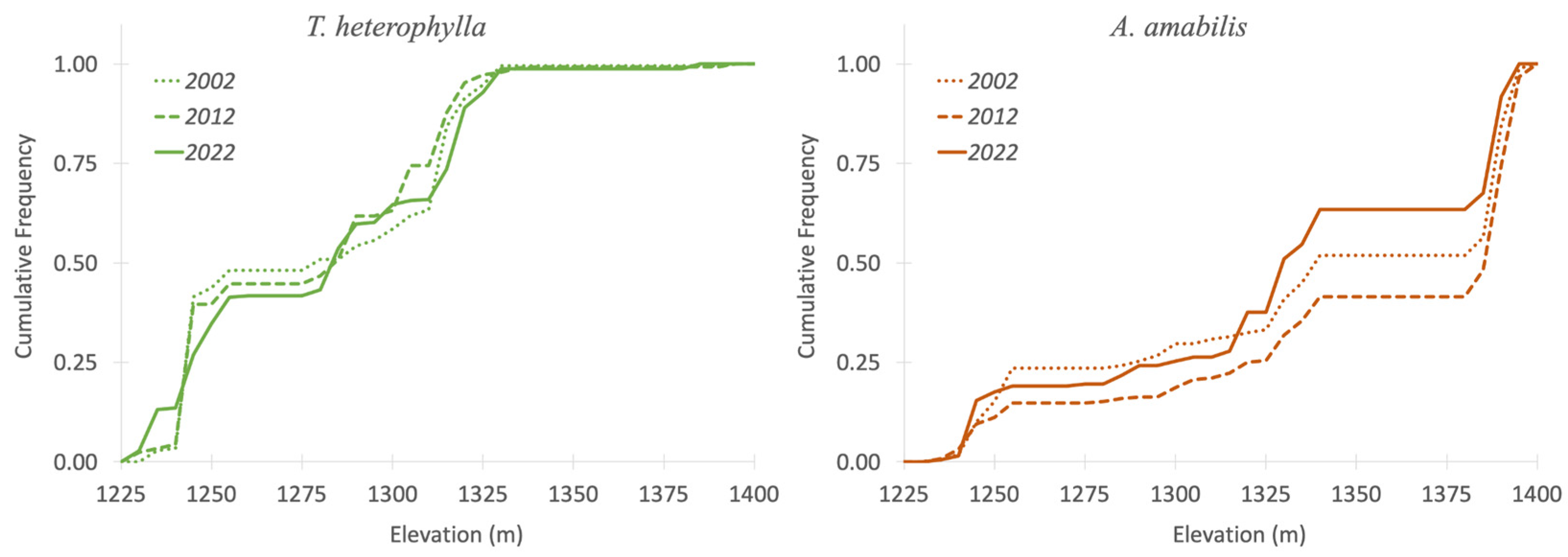
2.3.2. Forest Regeneration
3. Results
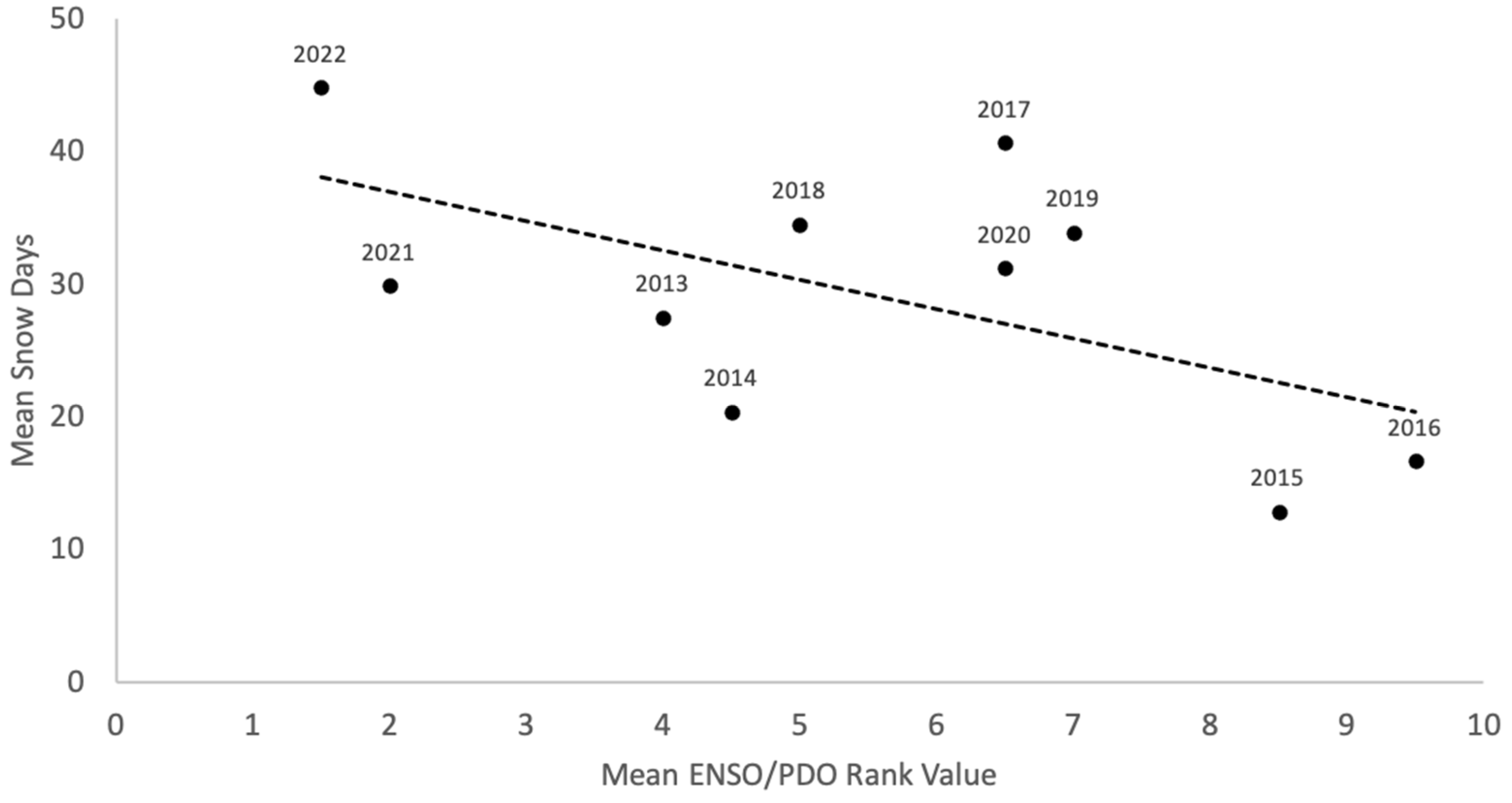
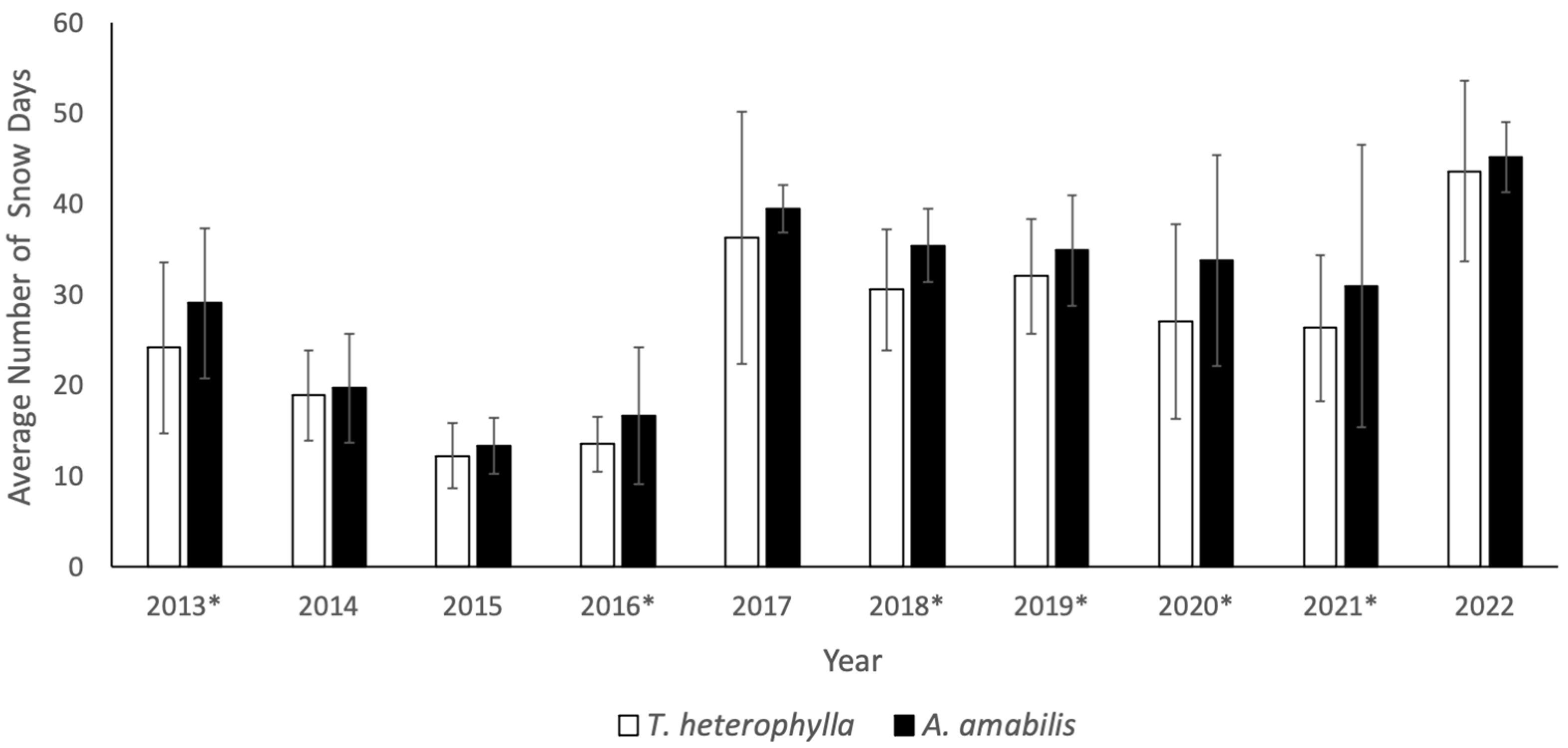
3.1. Snow Variability
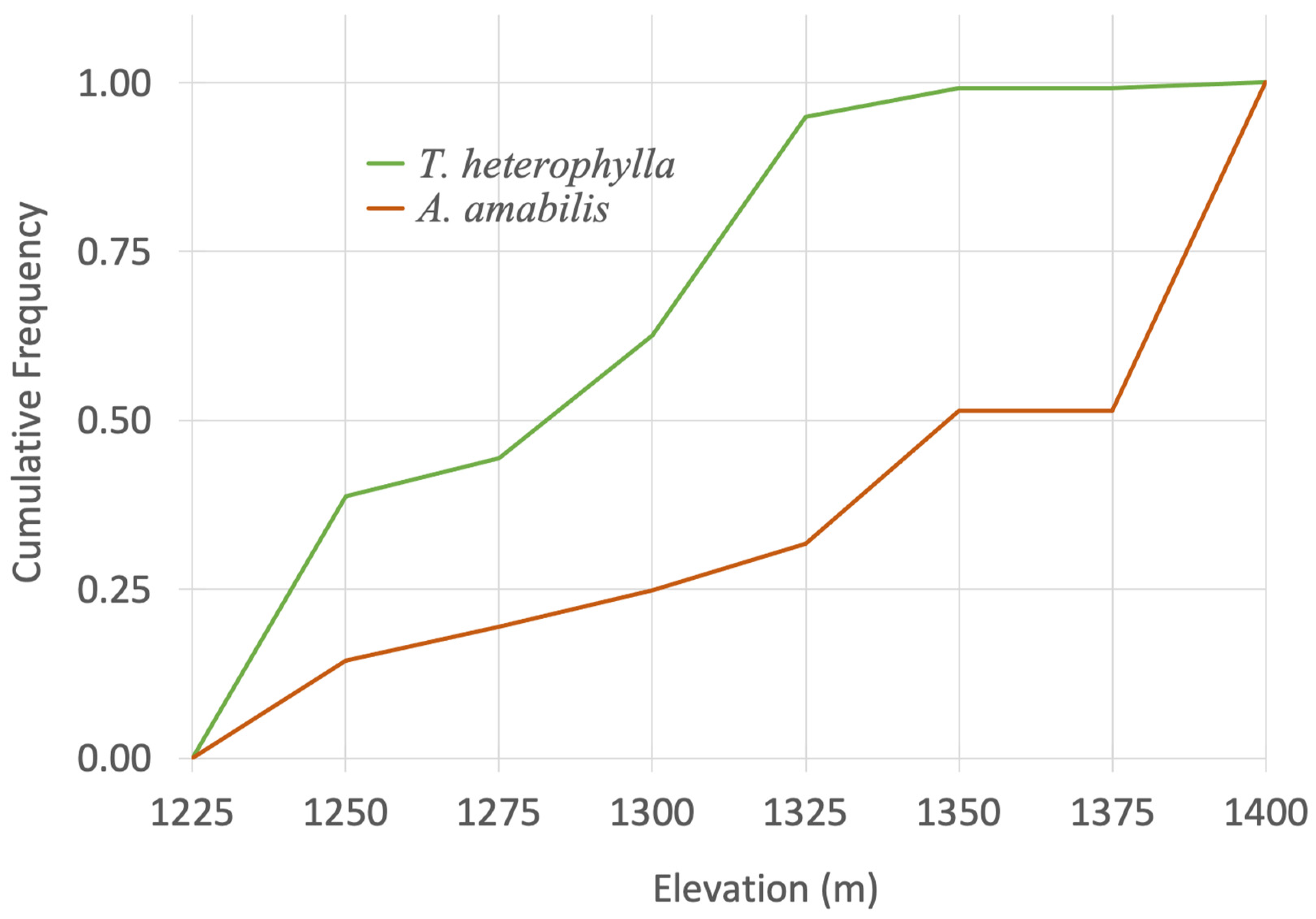
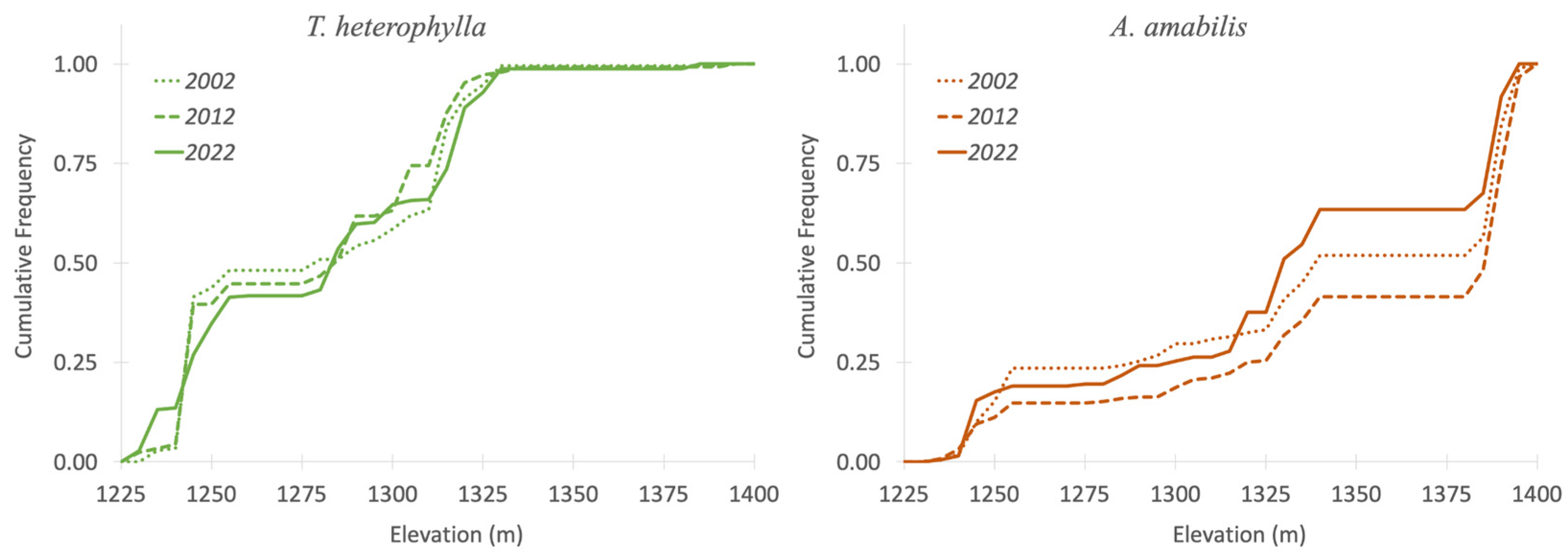
3.2. Forest Regeneration
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierce, D.; Cayan, D. The uneven response of different snow measures to human-induced climate warming. J. Clim. 2013, 26, 4148–4167. [Google Scholar] [CrossRef]
- Rhoades, A.M.; Jones, A.D.; Ullrich, P.A. The changing character of the California Sierra Nevada as a natural reservoir. Geophys. Res. Lett. 2018, 45, 8–13. [Google Scholar] [CrossRef]
- Ikeda, K.; Rasmussen, R.; Liu, C.; Newman, A.; Chen, F.; Barlage, M.; Gutmann, E.; Dudhia, J.; Dai, A.; Luce, C.; et al. Snowfall and snowpack in the Western U.S. as captured by convection permitting climate simulations: Current climate and pseudo global warming future climate. Clim. Dyn. 2021, 57, 2191–2215. [Google Scholar] [CrossRef]
- Beniston, M. Climatic change in mountain regions: A review of possible impacts. Clim. Change 2003, 59, 5–31. [Google Scholar] [CrossRef]
- Sproles, E.A.; Nolin, A.W.; Rittger, K.; Painter, T.H. Climate change impacts on maritime mountain snowpack in the Oregon Cascades. Hydrol. Earth Syst. Sci. 2013, 17, 2581–2597. [Google Scholar] [CrossRef]
- Mazzotti, G.; Webster, C.; Quéno, L.; Cluzet, B.; Jonas, T. Canopy structure, topography, and weather are equally important drivers of small-scale snow cover dynamics in sub-alpine forests. Hydrol. Earth Syst. Sci. 2023, 27, 2099–2121. [Google Scholar] [CrossRef]
- Dickerson-Lange, S.E.; Gersonde, R.F.; Hubbart, J.A.; Link, T.E.; Nolin, A.W.; Perry, G.H.; Roth, T.R.; Wayand, N.E.; Lundquist, J.D. Snow disappearance timing is dominated by forest effects on snow accumulation in warm winter climates of the Pacific Northwest, United States. Hydrol. Process. 2017, 31, 1846–1862. [Google Scholar] [CrossRef]
- Mood, B.J.; Coulthard, B.; Smith, D.J. Three hundred years of snowpack variability in southwestern British Columbia reconstructed from tree-rings. Hydrol. Process. 2020, 34, 5123–5133. [Google Scholar] [CrossRef]
- Jump, A.S.; Mátyás, C.; Peñuelas, J. The altitude-for-latitude disparity in the range retractions of woody species. Trends Ecol. Evol. 2009, 24, 694–701. [Google Scholar] [CrossRef]
- Breshears, D.D.; Huxman, T.E.; Adams, H.D.; Davison, J.E. Vegetation synchronously leans upslope as climate warms. Proc. Natl. Acad. Sci. USA 2008, 105, 11591–11592. [Google Scholar] [CrossRef]
- Vitasse, Y.; Ursenbacher, S.; Klein, G.; Bohnenstengel, T.; Chittaro, Y.; Delestrade, A.; Monnerat, C.; Rebetez, M.; Rixen, C.; Strebel, N.; et al. Phenological and elevational shifts of plants, animals and fungi under climate change in the European Alps. Biol. Rev. Camb. Philos. Soc. 2021, 96, 1816–1835. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.G.; Lee-Yaw, J.A.; Sunday, J.M.; Hargreaves, A.L. Expanding, shifting, and shrinking: The impact of global warming on species’ elevational distributions. Glob. Ecol. Biogeogr. 2018, 27, 1268–1276. [Google Scholar] [CrossRef]
- Spies, T.A.; Giesen, T.W.; Swanson, F.J.; Franklin, J.F.; Lach, D.; Johnson, K.N. Climate change adaptation strategies for federal forests of the Pacific Northwest, USA: Ecological, policy, and socio-economic perspectives. Landsc. Ecol. 2010, 25, 1185–1199. [Google Scholar] [CrossRef]
- Hagedorn, B.; Flower, A. Conifer establishment and encroachment on subalpine meadows around Mt. Baker, WA, USA. Forests 2021, 12, 1390. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring snowmelt increase western US forest fire activity. Science 2006, 313, 949–952. [Google Scholar] [CrossRef] [PubMed]
- Contosta, A.R.; Casson, N.J.; Garlick, S.; Nelson, S.J.; Ayres, M.P.; Burakowski, E.A.; Campell, J.; Creed, I.; Eimers, C.; Evans, C.; et al. Northern forest winters have lost cold, snowy conditions that are important for ecosystems and human communities. Ecol. Appl. 2019, 29, e01974. [Google Scholar] [CrossRef] [PubMed]
- Kueppers, L.M.; Conlisk, E.; Castanha, C.; Moyes, A.B.; Germino, M.J.; de Valpine, P.; Torn, M.S.; Mitton, J.B. Warming and provenance limit tree recruitment across and beyond the elevation range of subalpine forest. Glob. Change Biol. 2016, 23, 2383–2395. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.M.; Young, D.J.N.; Safford, H.D.; Young, T.P. Decreased snowpack and warmer temperatures reduce the negative effects of interspecific competitors on regenerating conifers. Oecologia 2019, 191, 731–743. [Google Scholar] [CrossRef]
- Ford, K.R.; HilleRisLambers, J. Soil alters seedling establishment responses to climate. Ecol. Lett. 2019, 23, 140–148. [Google Scholar] [CrossRef]
- Kark, S. Ecotones and ecological gradients. In Ecological Systems; Leemans, R., Ed.; Springer: New York, NY, USA, 2012; pp. 147–160. [Google Scholar]
- Wasson, K.; Woolfolk, A.; Fresquez, C. Ecotones as indicators of changing environmental conditions: Rapid migration of salt marsh-upland boundaries. Estuaries Coasts 2013, 36, 654–664. [Google Scholar] [CrossRef]
- Nelson, K.N.; O’Dean, E.; Knapp, E.E.; Parker, A.J.; Bisbing, S.M. Persistent yet vulnerable: Resurvey of an Abies ecotone reveals few differences but vulnerability to climate change. Ecology 2021, 102, e03525. [Google Scholar] [CrossRef] [PubMed]
- Dearborn, K.D.; Danby, R.K. Spatial analysis of forest-tundra ecotones reveals the influence of topography and vegetation on alpine treeline patterns in the subarctic. Ann. Am. Assoc. Geogr. 2019, 110, 18–35. [Google Scholar] [CrossRef]
- Gleason, K.E.; Nolin, A.W.; Roth, T.R. Developing a representative snow-monitoring network in a forested mountain watershed. Hydrol. Earth Syst. Sci. 2017, 21, 1137–1147. [Google Scholar] [CrossRef]
- Kostadinov, T.S.; Lookingbill, T.R. Snow cover variability in a forest ecotone of the Oregon Cascades via MODIS Terra products. Remote Sens. Environ. 2015, 164, 155–169. [Google Scholar] [CrossRef]
- Kostadinov, T.S.; Schumer, R.; Hausner, M.; Bormann, K.J.; Gaffney, R.; McGwire, K.; Painter, T.H.; Tyler, S.; Harpold, A.A. Watershed-scale mapping of fractional snow cover under conifer forest canopy using lidar. Remote Sens. Environ. 2019, 222, 34–49. [Google Scholar] [CrossRef]
- De Frenne, P.; Zellweger, F.; Rodríguez-Sánchez, F.; Scheffers, B.R.; Hylander, K.; Luoto, M.; Vellend, M.; Verheyen, K.; Lenoir, J. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Aulló-Maestro, I.; Gómez, C.; Hernández, L.; Camarero, J.J.; Sánchez-González, M.; Cañellas, I.; de la Cueva, A.V.; Montes, F. Monitoring montane-subalpine forest ecotone in the Pyrenees through sequential forest inventories and Landsat imagery. Ann. For. Sci. 2023, 80, 32. [Google Scholar] [CrossRef]
- Franklin, J.F.; Dyrness, C.T. Natural Vegetation of Oregon and Washington; Oregon State University Press: Corvallis, OR, USA, 1988. [Google Scholar]
- Fleishman, E. (Ed.) Sixth Oregon Climate Assessment; Oregon Climate Change Research Institute, Oregon State University: Corvallis, OR, USA, 2023; Available online: https://blogs.oregonstate.edu/occri/oregon-climate-assessments (accessed on 30 June 2023).
- Lookingbill, T.R.; Urban, D.L. Spatial estimation of air temperature differences for landscape-scale studies in montane environments. Agric. For. Meteorol. 2003, 114, 141–151. [Google Scholar] [CrossRef]
- Lookingbill, T.; Urban, D. An empirical approach towards improved spatial estimates of soil moisture for vegetation analysis. Landsc. Ecol. 2004, 19, 417–433. [Google Scholar] [CrossRef]
- Pierce, K.B.; Lookingbill, T.; Urban, D. A simple method for estimating potential relative radiation (PRR) for landscape-scale vegetation analysis. Landsc. Ecol. 2005, 20, 137–147. [Google Scholar] [CrossRef]
- Lookingbill, T.R.; Urban, D.L. Gradient analysis, the next generation: Towards more plant-relevant explanatory variables. Can. J. For. Res. 2005, 35, 1744–1753. [Google Scholar] [CrossRef]
- Lookingbill, T.R.; Rocca, M.E.; Urban, D.L. Focused assessment of scale-dependent vegetation pattern. In Predictive Species and Habitat Modeling in Landscape Ecology; Drew, C.A., Wiersma, Y., Huettmann, F., Eds.; Springer: New York, NY, USA, 2010; pp. 111–138. [Google Scholar]
- Grubb, P.J. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. Camb. Philos. Soc. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Serra-Diaz, J.M.; Franklin, J.; Sweet, L.C.; McCullough, I.M.; Syphard, A.D.; Regan, H.M.; Flint, L.E.; Flint, A.L.; Dingman, J.R.; Moritz, M.A.; et al. Averaged 30 year climate change projections mask opportunities for species establishment. Ecography 2016, 39, 844–845. [Google Scholar] [CrossRef]
- Hall, D.K.; Foster, J.L.; DiGirolamo, N.; Riggs, G.A. Snow cover, snowmelt timing and stream power in the Wind River Range, Wyoming. Geomorphology 2012, 137, 87–93. [Google Scholar] [CrossRef]
- Wolter, K.; Timlin, M.S. Monitoring ENSO in COADS with a seasonally adjusted principal component index. In Proceedings of the 17th Climate Diagnostics Workshop, Norman, OK, USA, 18–23 October 1992; pp. 52–57. [Google Scholar]
- Wolter, K.; Timlin, M.S. Measuring the strength of ENSO events: How does 1997/1998 rank? R. Meteorol. Soc. 1998, 53, 315–324. [Google Scholar]
- Mantua, N.J.; Hare, S.R.; Zhang, Y.; Wallace, J.M.; Francis, R.C. A pacific interdecadal climate oscillation with impacts on salmon production. Bull. Am. Meteorol. Soc. 1997, 78, 1069–1080. [Google Scholar] [CrossRef]
- Zhang, Y.; Wallace, J.M.; Battisti, D.S. ENSO-like interdecadal variability. J. Clim. 1997, 10, 1004–1020. [Google Scholar] [CrossRef]
- Tyler, S.W.; Selker, J.S.; Hausner, M.B.; Hatch, C.E.; Torgersen, T.; Thodal, C.E.; Schladow, S.G. Environmental temperature sensing using Raman spectra DTS fiber-optic methods. Water Resour. Res. 2009, 45, W00D23. [Google Scholar] [CrossRef]
- Raleigh, M.S.; Rittger, K.; Moore, C.E.; Henn, B.; Lutz, J.A.; Lundquist, J.D. Ground-based testing of MODIS fractional snow cover in subalpine meadows and forests of the Sierra Nevada. Remote Sens. Environ. 2013, 128, 44–57. [Google Scholar] [CrossRef]
- Tutton, R.J.; Way, R.G. A low-cost method for monitoring snow characteristics at remote field sites. Cryosphere 2021, 15, 1–15. [Google Scholar] [CrossRef]
- Thornburgh, D.A. Dynamics of the True Fir-Hemlock Forests of the West Slope of the Washington Cascade Range; University of Washington: Seattle, WA, USA, 1969. [Google Scholar]
- Mori, A.S.; Mizumachi, E.; Sprugel, D.G. Morphological acclimation to understory environments in Abies amabilis, a shade- and snow-tolerant conifer species of the Cascade Mountains, Washington, USA. Tree Physiol. 2008, 28, 815–824. [Google Scholar] [CrossRef]
- Jones, J.A.; Creed, I.F.; Hatcher, K.L.; Warren, R.J.; Adams, M.B.; Benson, M.H.; Boose, E.; Brown, W.A.; Campell, J.L.; Covich, A.; et al. Ecosystem processes and human influences regulate streamflow response to climate change at long-term ecological research sites. BioScience 2012, 62, 390–404. [Google Scholar] [CrossRef]
- Weatherhead, E.C.; Reinsel, G.C.; Tiao, G.C.; Meng, X.; Choi, D.; Cheang, W.; Keller, T.; DeLuisi, J.; Wuebbles, D.J.; Kerr, J.B.; et al. Factors affecting the detection of trends: Statistical considerations and applications to environmental data. J. Geophys. Res. 1998, 103, 17149–17161. [Google Scholar] [CrossRef]
- Beaulieu, C.; Henson, S.A.; Sarmiento, J.L.; Dunne, J.P.; Doney, S.C.; Rykaczewski, R.R.; Bopp, L. Factors challenging our ability to detect long-term trends in ocean chlorophyll. Biogeosciences 2013, 10, 2711–2724. [Google Scholar] [CrossRef]
- Brown, C.D.; Vellend, M. Non-climatic constraints on upper elevational plant range expansion under climate change. R. Soc. Biol. 2014, 281, 20141779. [Google Scholar] [CrossRef] [PubMed]
- Hannah, L.; Flint, L.; Syphard, A.D.; Moritz, M.A.; Buckley, L.B.; McCullough, I.M. Fine-grain modeling of species’ response to climate change: Holdouts, stepping-stones, and microrefugia. Trends Ecol. Evol. 2014, 29, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Bonannella, C.; Parente, L.; de Bruin, S.; Herold, M. Multi-decadal trend analysis and forest disturbance assessment of European tree species: Concerning signs of a subtle shift. For. Ecol. Manag. 2024, 554, 121652. [Google Scholar] [CrossRef]
- Smith, A.L.; Blair, D.; McBurney, L.; Banks, S.C.; Barton, P.S.; Blanchard, W.; Driscoll, D.A.; Gill, A.M.; Lindenmayer, D.B. Dominant drivers of seedling establishment in a fire-dependent obligate seeder: Climate or fire regimes? Ecosystems 2014, 17, 258–270. [Google Scholar] [CrossRef]
- Liang, S.; Hurteau, M.D. Novel climate-fire-vegetation interactions and their influence on forest ecosystems in the western USA. Funct. Ecol. 2023, 37, 2126–2142. [Google Scholar] [CrossRef]
- Lippok, D.; Beck, S.G.; Renison, D.; Hensen, I.; Apaza, A.E.; Schleuning, M. Topography and edge effects are more important than elevation as drivers of vegetation patterns in a neotropical montane forest. J. Veg. Sci. 2013, 25, 724–733. [Google Scholar] [CrossRef]
- Alatalo, J.M.; Ferrarini, A. Braking effect of climate and topography on global change-induced upslope forest expansion. Int. J. Biometeorol. 2016, 61, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.N.; Spies, T.A. Microsite controls on tree seedling establishment in conifer forest canopy gaps. Ecology 1997, 78, 2458–2473. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 2004, 34, 59–234. [Google Scholar]
- Halofsky, J.E.; Peterson, D.L. Climate change vulnerabilities and adaptation options for forest vegetation management in the Northwestern USA. Atmosphere 2016, 7, 46. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cameras | |||
|---|---|---|---|
| Snow-Covered | Snow-Free | ||
| Data Loggers | Snow-Covered | 3496 (28.3%) | 186 (1.5%) |
| Snow-Free | 141 (1.1%) | 8534 (69.1%) | |
| Species | Total | Percentage on Nurse Logs |
|---|---|---|
| Western hemlock (T. heterophylla) | 829 | 80.9% |
| Silver fir (A. amabilis) | 734 | 4.3% |
| Grand fir (A. grandis) | 10 | 0.0% |
| Noble fir (A. procera) | 277 | 10.1% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lookingbill, T.R.; DuPuy, J.; Jacobs, E.; Gonzalez, M.; Kostadinov, T.S. A 20-Year Ecotone Study of Pacific Northwest Mountain Forest Vulnerability to Changing Snow Conditions. Land 2024, 13, 424. https://doi.org/10.3390/land13040424
Lookingbill TR, DuPuy J, Jacobs E, Gonzalez M, Kostadinov TS. A 20-Year Ecotone Study of Pacific Northwest Mountain Forest Vulnerability to Changing Snow Conditions. Land. 2024; 13(4):424. https://doi.org/10.3390/land13040424
Chicago/Turabian StyleLookingbill, Todd R., Jack DuPuy, Ellery Jacobs, Matteo Gonzalez, and Tihomir S. Kostadinov. 2024. "A 20-Year Ecotone Study of Pacific Northwest Mountain Forest Vulnerability to Changing Snow Conditions" Land 13, no. 4: 424. https://doi.org/10.3390/land13040424
APA StyleLookingbill, T. R., DuPuy, J., Jacobs, E., Gonzalez, M., & Kostadinov, T. S. (2024). A 20-Year Ecotone Study of Pacific Northwest Mountain Forest Vulnerability to Changing Snow Conditions. Land, 13(4), 424. https://doi.org/10.3390/land13040424