Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth


1
Department of Inorganic and Organic Chemistry, University of Barcelona, Institute of Cosmos Science (UB-EEC), c. Martí i Franquès 1, 08028-Barcelona, Catalonia, Spain
2
Centro de Astrobiología (CSIC-INTA), Department of Molecular Evolution, Carretera Ajalvir Kilómetro 4, 28850 Torrejón de Ardoz, Madrid, Spain
*
Author to whom correspondence should be addressed.
Symmetry 2019, 11(6), 814; https://doi.org/10.3390/sym11060814
Submission received: 31 May 2019
/
Revised: 14 June 2019
/
Accepted: 18 June 2019
/
Published: 20 June 2019
(This article belongs to the Special Issue Possible Scenarios for Homochirality on Earth)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Spontaneous mirror symmetry breaking (SMSB), a phenomenon leading to non-equilibrium stationary states (NESS) that exhibits biases away from the racemic composition is discussed here in the framework of dissipative reaction networks. Such networks may lead to a metastable racemic non-equilibrium stationary state that transforms into one of two degenerate but stable enantiomeric NESSs. In such a bifurcation scenario, the type of the reaction network, as well the boundary conditions, are similar to those characterizing the currently accepted stages of emergence of replicators and autocatalytic systems. Simple asymmetric inductions by physical chiral forces during previous stages of chemical evolution, for example in astrophysical scenarios, must involve unavoidable racemization processes during the time scales associated with the different stages of chemical evolution. However, residual enantiomeric excesses of such asymmetric inductions suffice to drive the SMSB stochastic distribution of chiral signs into a deterministic distribution. According to these features, we propose that a basic model of the chiral machinery of proto-life would emerge during the formation of proto-cell systems by the convergence of the former enantioselective scenarios.
1. Introduction
The origin of biological homochirality (BH) [1,2,3,4] is often described as a scientific mystery [5] and the lack of a scientific explanation of the BH phenomenon is often used as proof against the theory of evolution [6]. However, it is possible nowadays to give a rigorous basic physico-chemical justification for the origin of BH. There are indisputable experimental reports on spontaneous mirror symmetry breaking (SMSB) [7,8,9] as well as many theoretical reports. SMSB can be interpreted within the framework of entropy production and dissipative processes [10,11]. This means that BH belongs to the same physico-chemical scenarios as those of open chemical networks proposed for the description of minimal self-reproducing proto-cells, primordial biochemical cycles, and the chemical thermodynamics of the reaction networks that maintain the phenomenon of life [12,13,14,15,16,17,18,19,20,21,22,23,24,25,26]. The paradox is that all of these reports eschew the enantioselective character of biological replicators, organocatalysis and autocatalytic reaction networks. Here we report that, in spite of the current mainstream ideas that dominate the possible explanations of BH, there are currently well-established theoretical and experimental chemical reports supporting chemical abiotic scenarios, which show a resilience to racemization and a preference for homochiral reaction outcomes with respect to the racemic ones, i.e. these are the very trends defining BH. We discuss these points with the aim to close the gap in the understanding of SMSB and of its place within the differentiated and hierarchical stages of chemical evolution.
The advances made in asymmetric induction during the 20th Century dominate the mainstream opinion on chiral methods in chemistry. Consequently, historically the emergence of homochirality has been justified mainly on the basis of the asymmetric induction of enantioselective reactions by physical chiral forces [2,27,28], in conjunction with non-linear asymmetric inductions leading to chiral amplifications [29,30,31] and the kinetic trapping of the final chiral species. To assume this as the basis for BH implies that during the evolutionary stages (see Figure 1) corresponding to the condensation reactions leading to the homochiral functional polymers and chiral replicators, homochirality is achieved by starting from enantiopure pools of homochiral building blocks (amino acids and carbohydrates). From a chemical point of view, this is an unlikely assumption that places SMSB at one of the earlier stages of chemical evolution as a singular event. In this regard, the low probability of such a scenario has been previously reported by some authors. Root-Bernstein proposed [32] the simultaneous origin of BH and the genetic code. Moreover, the relationship between the emergence of autocatalytic replication and homochirality has been reported, as well as the cooperative effect of external chiral polarizations [33]. However, despite their seminal significance, in our opinion, these proposals lack a description of their physico-chemical basis. Without this, the acceptance of such models by applied chemists is not possible. Furthermore, the design and experimental research in the field of artificial life cannot progress without the assumption of the chemical relationship between homochirality and autocatalysis. In this report, we describe: a) What type of enantioselective reaction networks may or may not lead to racemic biases; b) the general physico-chemical basis of how such enantioselective autocatalytic reaction networks may or may not lead to racemic biases; and c) how these SMSB systems (reaction networks plus open dissipative systems) depend on external chiral polarizations for the deterministic emergence in chemical evolution of one of the two chiral signs.
2. Racemization and Racemic Mixtures
Asymmetric synthesis of simple chiral organic compounds taking place under the effect of chiral forces and during the early stages of chemical evolution, i.e. in astrophysical scenarios, is an hypothesis that is being experimentally confirmed [35,36]. In such physical scenarios, strong chiral polarization forces can be expected to be operative. The chiral compounds obtained in such asymmetric synthesis must undergo racemization processes, which in spite of their low rates due to the low temperatures of interstellar space, would most likely be significant on long time scales. Therefore, in the currently and widely accepted hypothesis of the bombardment of Earth by planetesimal objects containing organic compounds [37], the enantiomeric excess (ee) values could be expected to be on the order of a few percent. Furthermore, on Earth, the increase of complexity towards supra and macromolecular compounds could only begin at the latter stages of the Hadean Era, when the Earth’s temperature decreased low enough so as to not decompose organic compounds. This means the complexity increase could not have occurred before the beginning of the zircon chronology. This also means that the starting temperatures of chemical evolution on Earth should be high enough to lead to significant racemization rates. Furthermore, condensation polymer synthesis probably requires experimental conditions favoring racemization processes [38]. In summary, it is chemically reasonable to expect an ee ≠ 0 during the stage of the formation of condensation polymers, but certainly much lower than the homochirality required for obtaining homochiral polymers (Figure 1) from pure enantiomeric pools of monomers. Note also that when assuming the first formation of polymers was not the result of a simple direct synthesis yet occurred under heterogeneous catalysis as mediated for example by inorganic materials such as clays [39], the existence of non-racemizing enantiopure pools of amino acids and sugars is highly unlikely.
A key point for starting a discussion of SMSB is the chemical meaning of a racemic mixture. Mislow discussed this concept in a Socratic-like method by proposing the question of whether a racemic mixture is composed by an exact number of molecules of both enantiomers [40]. In fact, older reports [41] had already established that there is an unavoidable statistical deviation from the ideal racemic composition as expressed by:
where N is the number of molecules. Notice that this statistical ee deviation (1) from the ideal racemic composition increases when the concentration decreases, so that for extremely dilute solutions there are low but statistically significant ee ≠ 0 values. Extremely low concentrations of racemic mixtures occur, for example, when we consider the chirality arising by isotopic element substitution according to natural isotopic compositions [42], or in the case of polymers of non-identical building blocks [43], such as those of the RNA-world and proteins, where random condensation leads to a sufficiently high number of isomers such that their concentrations are low enough to show detectable ee values [44]. However, chiral amplifications of such stochastically distributed initial ee’s in reversible reactions cannot take place via asymmetric induction reactions. This is because both at thermodynamic equilibrium and in non-equilibrium stationary states (NESS) in the linear regime of non-equilibrium thermodynamics, the racemic composition represents a stable potential well even in the presence of the statistical fluctuations about the ideal racemic composition.
Chemical evolution is not supported by irreversible reaction pathways and the existence of certain racemization rates must be assumed to occur. Therefore, despite the fluctuations about the definition of the ideal/mathematical racemic mixture, the real meaning of the racemic mixture is the existence of a thermodynamic potential minimum for the ideal racemic composition. This means that for any compositional bias from the ideal racemic composition, such as those associated with the statistical ee values (1), the system will tend to return to an ee = 0. Obviously, there is a statistical chiral fluctuation or chiral noise about the potential minimum of the pure 1:1 enantiomeric mixture, but the system tends to return to ee = 0. This is a consequence of the energy degeneracy between the enantiomers and of the law of large numbers acting in the thermodynamic limit [45]. In summary, chiral statistical fluctuations around a stable racemic configuration cannot be amplified.
In contrast to asymmetric synthesis, absolute asymmetric synthesis (AAS), in the absence of any chiral polarization other than that between the very enantiomeric species of the reaction network, is possible for some reaction networks having non-linear kinetics of the enantiomer concentrations. When the system is maintained far from thermodynamic equilibrium, it is due to the imposed boundary conditions. The final reaction states, provided the boundary conditions are maintained, are non-equilibrium stationary states (NESS). These types of reactions, or simply the deracemization of racemic NESS to scalemic or homochiral NESS, are called spontaneous mirror symmetry breaking (SMSB). It is worth noting that in a SMSB scenario the chiral statistical fluctuations about the ideal racemic composition are now necessary to expose the metastability of the racemic NESS, which does not correspond to a potential well topology but to that of a saddle point. It is important to stress that the role of the fluctuations is not that of an activation energy and that their effect at the SMSB event is not at all an amplification of chirality.
3. Stable and Unstable Non-Equilibrium Stationary States (NESS) in Enantioselective Reactions
Reaction networks that are able to lead to SMSB have focused the research interest on the topic [3,46]. The fact that such reaction networks may exhibit permanent SMSB as non-equilibrium stationary states (NESS) and only when operative in open or some special closed systems is assumed [47], but they are rarely analyzed within the thermodynamic framework of dissipative reaction systems. Moreover, previous seminal reports on dissipative reaction networks actually hamper, rather than facilitate, the chemical understanding of such systems. This is because the erroneous use of the kinetic approximation of clamped concentrations for the species exchanged with the environment overlooked the role of the boundary conditions of open systems for the correct description of the NESSs. This error originates from the first seminal reports on dissipative chemical reactions (e.g. [10]) and was most likely a consequence, at that time, of the mathematical difficulties of studying the sets of coupled non-linear differential equation describing reaction networks. Nowadays, this simplification cannot be excused when simple and powerful computational tools for solving such equations are readily available. The use of the clamped concentration approximation has even led to the refutation [48] of Prigogine’s theorem on minimum entropy production for the linear thermodynamic regime and to cast doubts on the validity of the General Evolution Criterion (GEC) valid for the non-linear thermodynamic regime. Further discussion concerning the correct thermodynamic description of models for the simulation of SMSB as NESS can be found in references [49,50].
3.1. Entropy Production and Balance in Open Systems
The boundary conditions (either systems open to matter exchange or closed systems unable to equilibrate energy with their surroundings) can keep the reaction network operating far from thermodynamic equilibrium. By increasing the “distance” from thermodynamic equilibrium, the internal entropy production of the reaction network increases [51]: dSi ≥ 0, where the equality corresponds to the thermodynamic equilibrium state. Depending how far the reaction network is driven from equilibrium, a NESS can result, forming the so-called thermodynamic branch (for example the black trace of Figure 2). However, in a real chemical system, above a critical value of the entropy production, instability of the NESS can occur. This is because taking the system very far from equilibrium leads to an increase of the entropy production, which is obtained by increasing the chemical affinities (forces) and the absolute reaction rates (fluxs/currents). This in turn corresponds to high concentrations, which can lead to increased viscosities, and so to a breakdown of the mean field approximation of chemical kinetics. Therefore, the reaction rates become diffusion-controlled and inhomogeneous distributions of the reaction species and the temperature takes place. This may lead to either chaotic distributions or to organized structures (oscillatory or stationary). The ordered structures arising as a consequence of the increased entropy production are called “dissipative” because they are maintained thanks to the entropy dissipated by the reaction network. However, for some autocatalytic reaction networks, and before such a spatial instability occurs, the thermodynamic branch can become unstable for homogeneous distributions of matter and energy that yield other types of NESS, or even oscillatory compositional states, distinct from the single NESS of the thermodynamic branch. It was recognized early on that this could occur only in the case of autocatalytic reactions [11]. However, such chemical reactions have received much less attention than the study of oscillating reactions or the formation of macroscopic dissipative spatial structures. Surprisingly, the enantioselective character of the autocatalytic reaction networks significant in the chemical machinery of life and in consequence the possible relationship with SMSB processes and BH has been mostly overlooked: only some reports considerer SMSB in the framework of entropy production and dissipative systems ([52,53,54,55]).
The entropy production (dSi) of a chemical reaction is always positive definite:
where the equality corresponds to thermodynamic equilibrium. In an open system at any stationary state the balance between the internal (reaction) entropy production (dSi) and the exchange entropy production (dSe) must be zero:
At thermodynamic equilibrium dS = 0 and dSi = 0, hence there is no exchange entropy with the exterior dSe = 0. At a NESS dS = 0, this means that the exchange entropy production is negative, allowing a more ordered internal configuration, than that corresponding to thermodynamic equilibrium: as occurs for example in SMSB. Non-equilibrium states of the system fulfilling equation (2) may be unstable, metastable or stable. States that do not have positive time derivative of the second variation of entropy may become unstable, but it is not a sufficient condition for instability [10,11,50]. Jacobian linear stability analysis distinguishes metastable from the true stable states [53]. Notice that there are non-stationary states with positive entropy production dSi > 0 obeying dS = 0. Non-stationary states with dS = 0 and d2S ≠ 0 correspond to states where the local equilibrium concept can be applied. These states may form continuous pathways between NESS and, therefore, represent the existence of thermodynamic reversible routes between different NESS, i.e. those NESSs that are not located in kinetic traps.
SMSB, and in the case of one asymmetric center, leads to a racemic NESS of saddle point topology (similar to that of transition state), which is able to evolve through reversible paths (while maintaining dS = 0) to one of two degenerate enantiomeric NESS located in an entropy production well dSi > 0 (the entropy production is minimized). More details about this will be published elsewhere. A schematic outline of this is shown in Figure 2, wherein the open system consists of a linear flow reactor where the achiral resources enter with a zero-order rate constant (as a molecular pump) in a well-mixed solution that has the same volume exiting the reactor, i.e. the exit of all species under the law of mass action. This open system at the final NESSs have constant volume and the same chemical mass ([A]O) for both the racemic and the homochiral NESS, which simplifies the evaluation of the associated entropy productions.
It is worth noting that the NESSs are thermodynamically defined by equality (3), referring to the entropy production balance between the internal reactions and the imposed boundary conditions, which is entirely overlooked under the clamped concentration approximation of SMSB commonly used to describe reaction networks in open systems.
The calculation of the entropy production (equal to the product of the forces and the currents that these forces lead to) is a chemical reaction that is relatively straightforward [52]. Furthermore, the way the contribution to the entropy production is calculated from the fluxes exchanging matter with surroundings and on the basis of the chemical potentials of the species can be found, for example, in reference [49]. From the point of view of chemical evolution, SMSB is subject to thermodynamic constraints and boundary conditions similar to those of the evolutionary stages of chemical networks.
3.2. Potential Reaction Networks Able to Yield SMSB.
The direct synthesis
fulfils the entropy balance (3) along the thermodynamic branch of racemic stationary states. However, enantioselective autocatalysis may also fulfill condition (3) for several different compositions.
Autocatalytic reaction networks capable of yielding species/enantiomer selection have been previously reviewed [3,46,47,56,57,58]. In addition to the networks cited/discussed therein, we must include the recently reported enantioselective hypercycle [50,59], which is highly significant due to its coincidence with the replicators of the nucleic acid and protein domain [60].
These reaction networks for SMSB contain a central mechanism in the first order (quadratic) enantioselective catalysis:
where A is achiral and D and L are enantiomers. Reaction (4) may also represent a simplification for the formation of chiral polymers from achiral building blocks. Reaction (5) can form homochiral NESS in the case that the initial conditions contain only one of the enantiomers. However, in any model concerning chemical evolution, the model should include the direct synthesis (4), albeit for a much slower reaction rate than for the enantioselective autocatalysis (5). This is because the autocatalytic functionality of (5) can only emerge if reaction (4) is operative. Reaction (4) can start for initial conditions without any D and L.
The first order autocatalysis (5), also called quadratic autocatalysis, cannot lead to species selection [56] (between D and L in SMSB), i.e., the entropy production needed to achieve the instability of the racemic branch is not high enough. This is because the kinetic/dynamic sigmoidal curve of growth implicit in autocatalytic processes (n > 0; see the meaning of n in Figure 2) is needed, as well as a super-exponential growth (n > 1) is necessary for species selection. Of course, this is not the case of cubic autocatalysis (n = 2), but in this case, high concentrations are necessary for the reaction progress in open system scenarios. Furthermore, to our knowledge, no natural replicators (autocatalytic systems) have been detected having a reaction order higher than the quadratic. Therefore, the reaction networks proposed for SMSB are based on first order autocatalysis, which through the coupling of other enantioselective reactions (Figure 3) yields, autocatalytic kinetic/dynamic signatures such as those expected for values of n > 1 necessary for achieving the instability of the thermodynamic branch and for certain reaction parameters and boundary conditions.
3.2.1. Frank-like Models
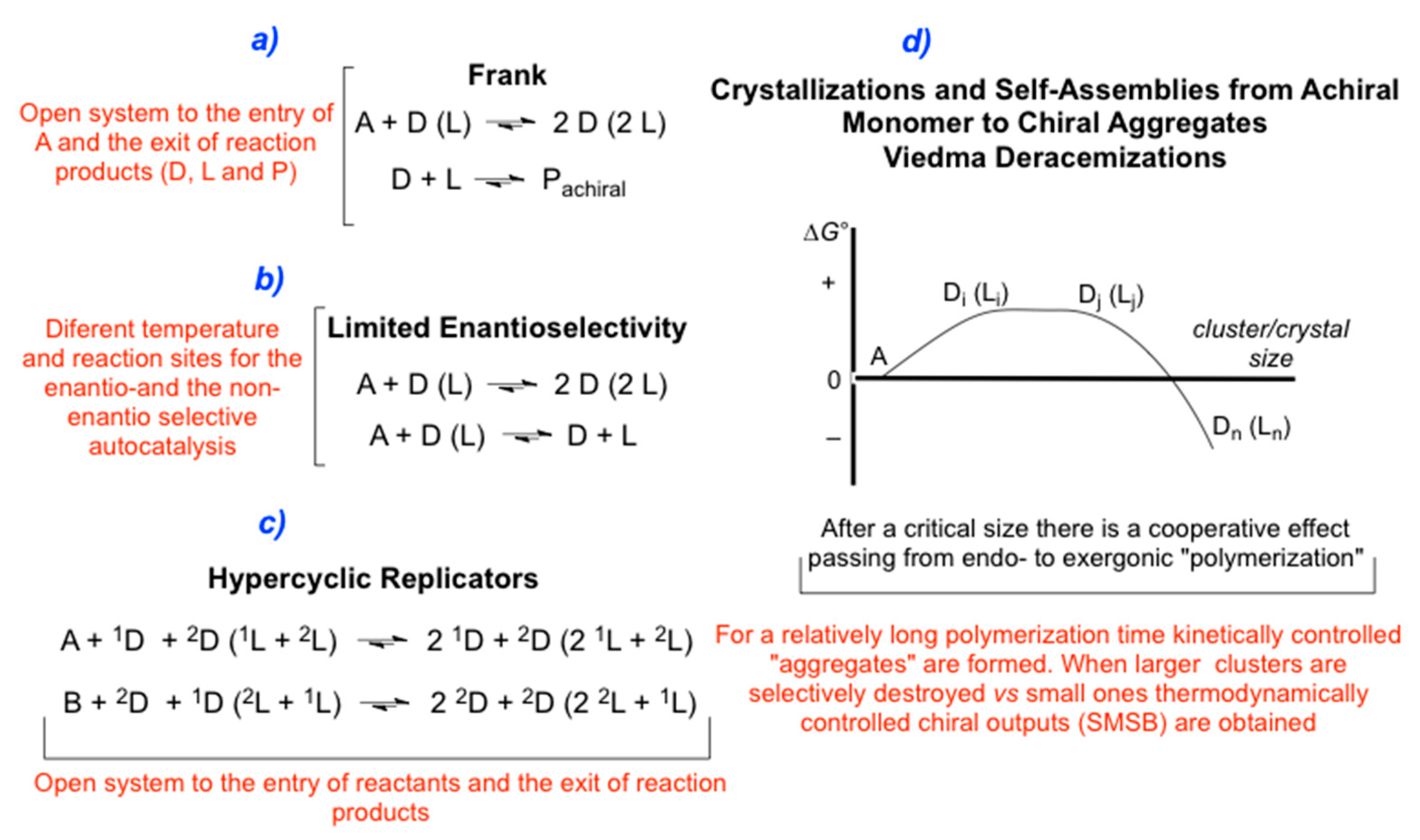
The first theoretical proposal of a reaction network able to lead to SMSB was the so-called Frank model (1953) [61] (Figure 3a). This model has its historical precedent in the Lotka-Volterra model for the competitive exclusion between two species [59], and when we take into account that enantiomerism is the limiting case of species distinguishability. Frank-like models (Figure 3a) contain the homochiral reaction between enantiomers as the additional chiral recognition step that allows for the increase in the dynamic of growth allowing enantiomer selection. All evidence indicates that the experimental SMSB of the Soai reaction [7,8] is a complex reaction network that basically fits into a Frank-like reaction network [62,63,64].
3.2.2. Limited Enantioselective Model
The enantioselectivity model (Figure 3b) proposed by Goldanskii and Avetisov [65] is based on the reasonable assumption of the presence of the corresponding non-enantioselective autocatalysis. However, thermodynamic constraints [66] determine that LES only makes thermodynamic sense when the enantioselective and the non-enantioselective autocatalytic steps are located at different points in the reaction domain and held at quite different temperatures [67,68,69]. There are no experimental examples for this model, but it adapts well to SMSB for the abiotic scenario of deep ocean thermal vents [68].
3.2.3. Enantioselective Hypercyclic Replicators
SMSB in hypercyclic replicators [59] is a consequence of the increase of non-linearity of the quadratic autocatalysis when cross-catalysis appears. In fact, the hypercycle model [14] was developed to solve the problem of selection and Darwian evolution for low autocatalytic orders. Although the implications of this model cannot be an ultimate explanation in systems biology, its raison d’etre is during the chemical evolution stages of compartmentalization of the RNA-world and the nucleic acid/ protein domain. Notice that this implies that the formation of systems able to self-reproduce while transmitting functionalities at the same time could be the origin of the emergence of BH.
Notice that if autocatalysis is present, the emergence of cross catalysis for the same family of compounds and chemical processes, for example polymerization of oligonucleotides or peptides, is quite reasonable from a point of view of chemical reactivity and structure. It is quite reasonable that autocatalytic and cross-catalytic functionalities could emerge simultaneously.
3.2.4. SMSB in Enantioselective Autocatalytic Polymerization/Depolymerization
In our opinion, the Viedma deracemization of chiral racemic conglomerates [9] (Figure 3c) in the presence of achiral or racemizing building blocks gives unprecedented hints on reaction networks able to lead to SMSB in the formation of condensation polymers. There are still controversial interpretations of the Viedma deracemization. In the following, we will discuss this according to our experimental and theoretical reports. Viedma deracemization can be interpreted, both experimentally and theoretically, as a phenomenon taking place within the framework of dissipative systems [70] which we summarize as follows.
A far from equilibrium system is created by mechanical grinding or by establishing temperature gradients [71,72] in solutions of crystals of racemic conglomerates made up from of either achiral or racemic monomers. These constitute closed systems unable to achieve energy equilibrium (energy balance) with their surroundings because:
- (a)
- In spite of being a closed system, the energy input is given selectively to only some of the species of the system. In other words, the mechanical grinding (the energy input) affects only the largest crystals.
- (b)
- The higher solubility of the smaller crystals obtained by grinding creates supersaturated solutions for the larger crystals and in consequence, a constant and permanent cycle of solubilization and crystal growth is maintained.
- (c)
- Homochiral cluster-to-cluster growth (this is equivalent—through its dynamic signature—to first order enantioselective autocatalysis). Notice that in saturated solutions monomer-to-cluster exchange between solution and crystal/cluster is not autocatalytic and therefore is unable to lead to any chiral amplification, nor to racemization, of the crystal mixture ee value.
- (d)
- Additional growth dynamics to first order autocatalysis of the cluster-to-cluster growth in (c) that increase the dynamic growth signature are provided by a mutualistic effect in the growth of homochiral material; each cluster coming from the fragmentation of large crystals can react with crystals of many different sizes leading to the formation of larger cluster/crystals of many different sizes, but of the same chiral sign. Furthermore, the free energy profile of the polymer formation, which is endergonic during the first “polymerization” steps, with cooperative growth beyond a critical size as well the intermediate sizes of the clusters coming from the breakage of the large ones, are decisive features for the achievement of the growth dynamics leading to SMSB [70].
From a synthetic chemical point of view, it is paradoxical how the resulting homochiral mixture does not racemize when the experimental conditions a) and b) above are halted. This is simply due to the fact that the solid chiral species at saturated conditions do not show solid-to-solid interactions. Therefore, in the absence of chiral recognition between enantiomeric phases, they are thermodynamically identical (for a detailed discussion of this, see refs. [73,74]). Furthermore, there are results pointing to the absence of ripening mechanisms of the larger crystals at the expense of the smaller ones in Viedma deracemization [75]. This suggests that the former results on the homochiral biases reported earlier by Kondepudi, in the formation of strong racemic biases in such types of racemic conglomerates [76,77], could belong to this type of SMSB mechanism. Regarding a generalization of the Viedma deracemization, the increasing number of reports on the spontaneous formation of strong bias from the racemic composition in the self-assembly of achiral building blocks towards chiral aggregates or liquid crystals, is surely significant [78,79,80].
The lessons learned from Viedma deracemization, make SMSB scenarios reasonable for the formation of primordial homochiral biopolymers based on cluster-to-cluster growth, as for example by template mechanisms in conjunction with the catalyzed depolymerization of the larger polymer chains to intermediate oligomer sizes. This type of SMSB polymerization has been previously theoretically studied [81]. There are experimental reports on oligomer formation/depolymerization cycles of peptides [82] that open prospects on the translation of the former speculations into experimental research [47].
3.2.5. On the Detection of SMSB in Low Exergonic Reactions in Solution
Most of the examples on SMSB correspond to phase transitions: crystals, self-assembly aggregates, e.g. Viedma-like deracemizations (e.g. [9,76,77,78,79,80]), that is, cases for which when the boundary conditions disappear allowing the system to equilibrate with its surroundings, racemization cannot occur. The only indisputable example of SMSB in solution is the Soai reaction [7]. However, the Soai reaction corresponds to a high exergonic reaction (organozinc addition to a carbaldehyde) that allows a reaction workup withouth significant racemization. By contrast, prebiotic chemistry involves many low exergonic reactions [83].
The lack of experimental results on the emergence of natural optical activity in autocatalytic reaction networks in solution, for example in the formose reaction (see below) in the carbohydrate domain, is hampered by the lack of synthetic methods and techniques for the direct study of the final stationary states in open systems. SMSB in solution should be performed in open systems under strength-controlled parameters, in contrast to the usual synthetic experiments carried out in the flask. Furthermore, the emergence of natural optical activity in low exergonic reactions in these open systems should be performed in real time, because racemization may occur in the common reaction workups when the open system conditions disappear. However, open system reactors at the bench scale are today possible thanks to microfluidic instrumentation. In addition, detection of natural optical activity for specific compounds, or families of compounds, could be carried out by circular dichroism (CD) spectroscopy, direct observation of reactor content, or of its effluents.
4. BH Based on SMSB Requires Previous Asymmetric Synthetic Scenarios: Stochastic vs. Deterministic Chiral Signs in SMSB
We assume that the decisive stage in the formation of proto-cells is the compartmentalization [84] of similar autocatalytic sets in a myriad of abiotic reactors [12,13]. Therefore, a hypothesis on a BH based on SMSB for the autocatalytic sets reveals the paradox that the stochastic chiral sign distribution in the different compartments would lead to a set of racemic outcomes when averaged over all the compartments. The hypothesis of the emergence of two competing enantiomeric worlds is in part contradictory to the hypothesis that by the exchange of species showing similar functions between compartmentalized autocatalytic sets, life emerges as a cooperative phenomenon [12,13,14,15,16,17,18,19,85].
However, the change from a stochastic to a deterministic distribution of chiral signs in SMSB is a remarkable property of SMSB. Under the effect of very weak polarizations, sufficiently weak so that such polarizations have no chemical consequence in common asymmetric synthesis, the stochastic distribution of chiral signs in SMSB changes to a single deterministic chiral sign. There are simulations of this [86,87], but the Soai reaction show dramatic examples, such as the deterministic distribution of chiral signs between experiments by such weak polarizations as those due to cryptochiral isotopic enantiomers [8].
Asymmetric induction by the action of natural physical chiral forces, active during the first stages of chemical evolution, can be considered as proven, despite of the diverse possible origins and organic compounds. The unavoidable slow racemization times (Section 2) do not prevent these compounds from arriving to the stage of the formation of instructed polymers and compartmentalization, showing small residual ee values. These low ee values of the organic compounds, constituting the autocatalytic replicators, would convert the stochastic SMSB chiral sign into a deterministic one.
According to this, BH would belong to a process developed through all the stages of chemical evolution. The selection of a deterministic chiral sign in the formation of instructed/functional polymers would constitute a first Darwinian selection, namely that of the phenotypic replicator able to survive by adopting the boundary chiral polarizations (the entry of resources into the compartment).
An interesting question is that the emergence of order in the evolutionary stage of compartmentalization arises because of a cooperative effect between autocatalytic networks and quasi-species. This information exchange has been proposed to arise thanks to similar catalytic functionalities of the exchanged species. This overlooks the chirality of the exchanged species. However, the chirality sign of similar functionalized polymers shows a chemical recognition of chirality based on a digitized +/− type of signal.
5. Hypothesis on the Emergence of BH
The former sections correspond to an attempt to describe selected concepts from different scientific fields, which are essential for the understanding of the emergence of chirality in chemical evolution and for the description of the basic principles necessary for the understanding of BH. In our opinion, the former discussion allows for a reasonable speculation of how, and in which stages of chemical evolution, the present chiral machinery of BH has been formed.
5.1. Is Carbohydrate Synthesis the Third Leg of a Tripodal SMSB Scenario?
Cross-catalysis between peptide and oligonucleotide hypercyclic replicators justifies a SMSB driving force to one enantiomer class in nucleic acids and in protein polymers [59]. Although weak this would require the chiral induction, coming from the asymmetric inductions transformations at previous stages of chemical evolution. However, the formation of primordial nucleotide-like compounds requires the selection of the adequate chiral carbohydrate monomer to achieve the structural functionalities of the nucleic acid chains [44,88]. Therefore, chirality in the domain of carbohydrate synthesis should be an essential element along the path leading to BH. In this respect, assuming SMSB as the central part of the emergence of BH, the question arises if the selection of one enantiomer of the carbohydrate family of compounds occurs by enantioselective selection at the formation of the nucleotides, or if it originated from its own SMSB process during the prebiotic stage of carbohydrate synthesis.
Prebiotic carbohydrate formation is based on aldol-like reactions integrating formaldehyde units to carbohydrate chains [89,90,91]. The synthetic integration of formaldehyde is the expected chemical pathway towards pentoses and hexoses: This was shown already by Butlerov (1861) in the so-called formose reaction. This reaction leads to a huge diversity of compounds, but the significant point with respect to SMSB is that it is composed by a family of interconnected autocatalytic cycles [92]. This strongly suggests that the necessary, but not sufficient, condition of autocatalysis for SMSB may occur at the prebiotic carbohydrate synthesis. However, there are no experimental data on the detection of spontaneous natural optical activity in the integrative synthesis of sugars, but we believe that this a consequence of a lack of methods to detect SMSB in solutions in open systems as discussed above.
The difficulty for theoretical studies of SMSB, and for future research experimental work in this topic in the carbohydrate domain, compared to those described in Section 3.2., is that, and with the exception of the first chiral compound of the family (glyceraldehyde or glycerol-1-phosphate), carbohydrates have more than one asymmetric center. This means that because of epimerization processes, i.e, the existence of chiral diastereoisomers, when the instability of the thermodynamic branch occurs, oscillatory final states may arise instead of the simple NESSs (saddle-point racemic NESS and two chiral NESS at minimum well potentials) described above. In this respect, it is surely significant that oscillations are an experimental behavior of glycolysis [90], and that in theoretical models implying compounds with two asymmetric centers, oscillatory behavior between epimers has been reported [93,94,95].
The difficulty for experimental studies of formose-like reactions is that, for the synthetic chemist, the final outcome is a “messy” mixture of compounds. However, from the point of view of the origin of life topic, it leads to a “high diversity” of carbohydrates. Both points of view are true, but diversity is probably a characteristic of the first stages of chemical evolution (see for example the thousands of compounds detected in a carbonaceous meteorite [96]). The first stages of chemical evolution are characterized by the formation of a very high diversity of organic compounds of the structural families that will constitute the chemical support of the phenomenon of life. Later, the increase of complexity, by forming interconnected reaction networks, the diversity would be thinned out towards a lower number of structural families and compounds. Synthetic chemical research in the origin of life is needed to discern which transformations are possible and which reaction mechanisms and interactions would have be at work to discover the reaction pathways that justify chemical evolution. However, abiotic scenarios need to be based on “impure” mixtures of compounds of related structural families to be reliable [97].
Reaction networks that show enantioselective autocatalysis are rare in chemistry. However, in the protein and nucleic acid domain the presence of replicators showing template mechanisms and the mutual catalytic activities between the RNA-world and peptide synthesis justify the connections between SMSB in these two domains. Yet the connection of these two with a possible SMSB at the carbohydrate metabolisms is more speculative. In the carbohydrate domain the question of BH is not only related to the supply of a specific pentose to the pre-RNA- or RNA-world, but also to membrane formation. In this respect, such a lateral connection is shown in the different starting chiral sign of the glycerol-1-phosphate building block to form the membrane lipids either in archeobacteria or in bacteria: chiral configuration L (sn-glycerol-1-phosphate) for archeobacteria and the chiral configuration D (sn-glycerol-3-phosphate) for bacteria [98].
5.2. Enantioselective Linear Reactions in the Chiral Machinery of Life
Regarding the chemical connection between chiral nitrogen-containing compounds and chiral hydroxy organic compounds, i.e. between the simple compounds of the amino acid and the carbohydrate domain (for example in the prebiotic chemical cycles proposed by Eschenmoser [83]), chirality can only imply simple asymmetric inductions and chirality transfer in linear kinetic dependences between enantiomers.
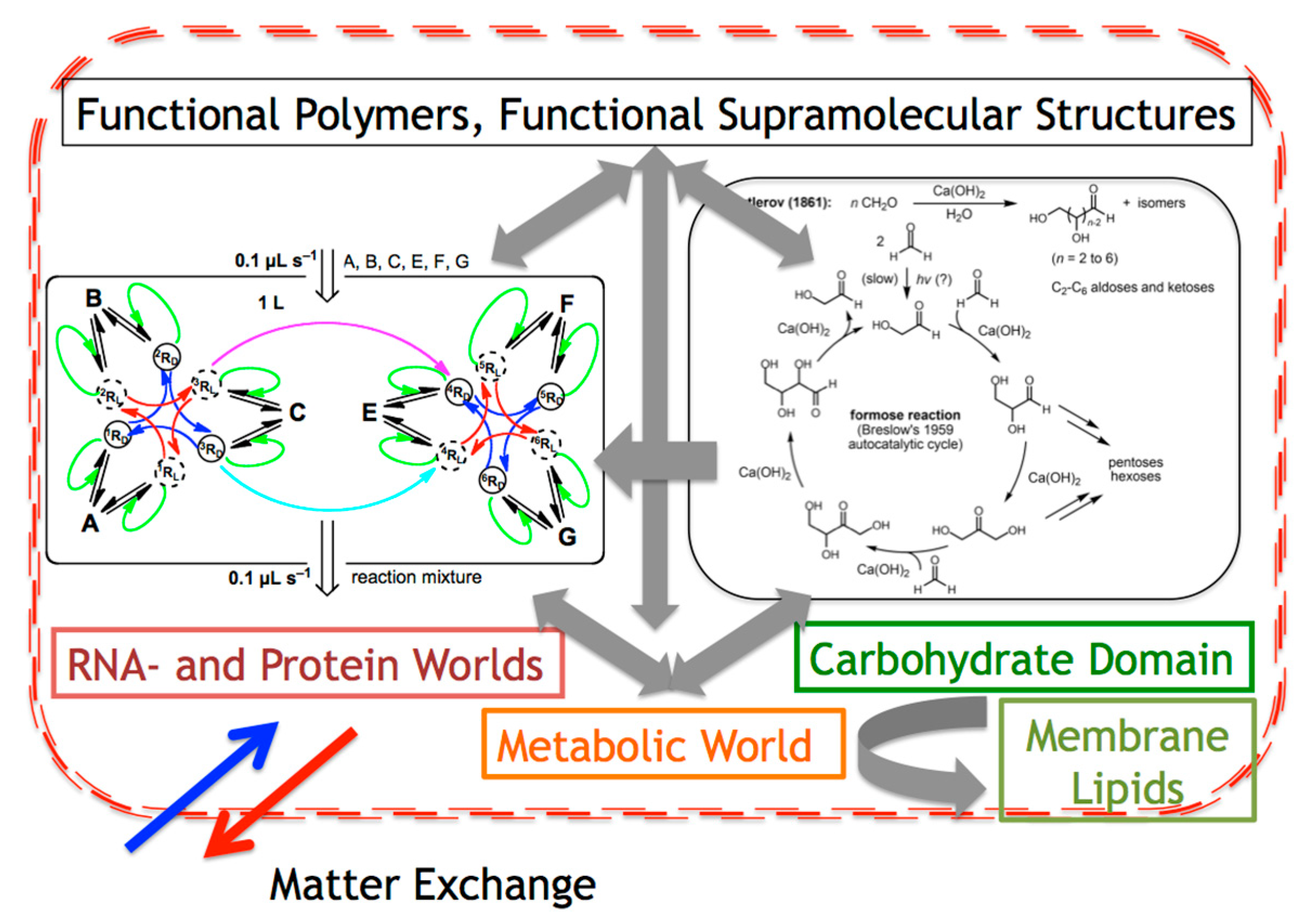
Metabolic cycles are necessary for the activation of low exergonic reactions, for example by substrate phosphorylation. Without the energy provided by metabolic cycles it is not possible for species activation to undergo endergonic or low exergonic reactions with higher yields and rates. It is surely significant that all indicate a minor role of enantiomerism in the metabolic world proposals, for example, the citric acid cycle, independent of the enzymes (enantioslective catalysis) that currently catalyze the cycle, chirality appears for only a few compounds and apparently as a mere accident that does not have an effect on the energetic function of the cycle. Chirality in metabolic cycles probably had appeared because of its interconnection with other worlds where SMSB had emerged. A reasonable hypothesis on the formation of proto-cells is to avoid the term “first” for the proposed models on the origin of life and consider the proto-cell emergence as a consequence of the mutualistic effects originating in the compartmentalization of the RNA-, protein-, carbohydrate-, lipid-, and metabolic-worlds (Figure 4). There, homochirality would be preserved by the SMSB autocatalysis of the interconnected RNA/protein and carbohydrate domains, which additionally would drive the rest of the enantioselective reactions and chiral transfer transformations to a common chiral sign.
6. Conclusions
- (a)
- BH has its physico-chemical basis in enantioselective autocatalytic reaction networks operating in open dissipative systems. This thermodynamic scenario is similar to that proposed for the compartmentalization stages of chemical evolution where the question of chirality has been avoided up to the present. Moreover, the reaction networks proposed for the formation of pre-RNA worlds and peptides showing catalytic and autocatalytic functionalities and able to develop Darwinian evolution, also possess the ability for SMSB, i.e. of the selection between enantiomers.
- (b)
- Asymmetric inductions originating through natural chiral forces during previous stages of chemical evolution, as for example in astrophysical scenarios, could provide the necessary chiral polarization to transform, at later stages of chemical evolution, the stochastic chiral sign outcome of the SMSB processes into a deterministic one. This is in spite of the unavoidable racemization processes acting during the long-time interval of chemical evolution.
- (c)
- SMSB autocatalytic cycles provide the resilience against racemization characteristic of BH and would drive the rest of the enantioselective reactions towards a definite chiral sign.
- (d)
- Such a general scenario of BH may be applied to all current models, which aim to find the links between prebiotic chemistry and biological chemistry [97]. The emergence of BH is concurrent with the emergence of autocatalytic sets, proto-cells, etc., and it probably represents an energetic advantage (lower entropy production) [51], with respect to the racemic outcome.
- (e)
- Homochirality has been considered as an inherent property of matter [98]. From a chemical point of view, it is probably also a necessary condition for life because of the informational entropy advantage of asymmetry in molecular recognition, that is, in the emergence of catalytic and autocatalytic functionalities. Once the reasonable existence of SMSB in the decisive stages of chemical evolution is recognized, in our opinion, then the models concerning the emergence of autocatalytic sets, proto-cells, etc., cannot avoid the question of the enantioselectivity of the processes involved. Note that this also has direct consequences for the applied biotechnological fields of new metabolic cycles and of artificial cells.
Author Contributions
J.M.R. and D.H. contributed equally to the present work.
Funding
This research was funded by MINECO (Spain), coordinated grant numbers CTQ2017-87864-C2-1-P and –C2-2-P.
Acknowledgments
We have benefitted from fruitful discussions with J. Crusats, Z. El-Hachemi and A. Moyano on the topic of this report, and in the framework of the coordinated project cited above.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guijarro, L.; Yus, M. The Origin of Chirality in the Molecules of Life: A Revision from Awareness to the Current; RSC Publishing: Cambridge, UK, 2009. [Google Scholar]
- Gal, J.; Cintas, P. Early History of the Recognition of Molecular Biochirality. In Biochirality; Springer: Berlin/Heidelberg, Germany, 2013; Volume 333, pp. 1–40. [Google Scholar]
- Blackmond, D.G. The origin of biological homochirality. Cold Spring Harb Perspect. Biol. 2010, 2, a002147. [Google Scholar] [CrossRef]
- Davankov, V.A. Biological homochirality on the earth, or in the universe? A selective review. Symmetry 2018, 10, 749. [Google Scholar] [CrossRef]
- An Evolutionary Mystery: Mirror Asymmetry in Life and in Space. Available online: https://www.acs.org/content/acs/en/acs-webinars/popular-chemistry/homochirality.html (accessed on 20 February 2019).
- Origin of Life. The Chirality Problem. Available online: https://creation.com/origin-of-life-the-chirality-problem (accessed on 20 February 2019).
- Soai, K.; Kawasaki, T. Asymmetric autocatalysis with amplification of chirality. In Amplification of Chirality; Springer: Berlin/Heidelberg, Germany, 2008; Volume 284, pp. 1–33. [Google Scholar]
- Kawasaki, T.; Matsumura, Y.; Tsutsumi, T.; Suzuki, K.; Ito, M.; Soai, K. Asymmetric autocatalysis triggered by carbon Isotope (13C/12C) chirality. Science 2009, 324, 492–495. [Google Scholar] [CrossRef]
- Viedma, C. Chiral symmetry breaking during crystallization: Complete chiral purity induced by non-linear autocatalysis. Phys. Rev. Lett. 2005, 94, 065504. [Google Scholar] [CrossRef]
- Glansdorff, P.; Prigogine, I. Thermodynamic Theory of Structure, Stability and Fluctuations; Wiley-Interscience: London, UK, 1971. [Google Scholar]
- Nicolis, G.; Prigogine, I. Self-Organization in Nonequilibrium Systems; Wiley: New York, NY, USA, 1977. [Google Scholar]
- Ruiz-Mirazo, K.; Briones, C.; de la Escosura, A. Prebiotic systems chemistry: New perspectives for the origins of life. Chem. Rev. 2014, 114, 285–366. [Google Scholar] [CrossRef]
- Kauffman, S. Autocatalytic sets of proteins. J. Theor. Biol. 1986, 119, 1–24. [Google Scholar] [CrossRef]
- Eigen, M.; Schuster, P. The Hypercycle: A Principle of Natural Self-Organization; Springer: Berlin, Germany, 1979. [Google Scholar]
- Hordijk, W.; Hein, J.; Steel, M. Autocatalytic sets and the origin of life. Entropy 2010, 12, 1733–1742. [Google Scholar] [CrossRef]
- Lifson, S. On the crucial stages in the origin of animate matter. J. Molec. Evol. 1997, 44, 1–8. [Google Scholar] [CrossRef]
- Schneider, E.D.; Kay, J.J. Life as manifestation of the second law of thermodynamics. Math. Comp. Model. 1994, 19, 25–48. [Google Scholar] [CrossRef]
- Mavelli, F.; Ruiz-Mirazo, K. Stochastic simulations of minimal self-reproducting cellular systems. Phil Trans. Royal Soc.: Biol. Sci. 2007, 362. [Google Scholar] [CrossRef]
- Rasmussen, S.; Chen, L.; Deamer, D.; Krakauer, D.C.; Packard, N.H.; Stadler, P.F.; Bedau, M.A. Transitions from Nonliving to Living Matter. Science 2004, 303, 963–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.; Beard, D.A. Thermodynamic of stoichiometric biochemical networks in living system far from equilibrium. Biophys. Chem. 2005, 114, 213–220. [Google Scholar] [CrossRef]
- Schmiedl, T.; Seifert, U. Stochastic thermodynamics of chemical reaction networks. J. Chem. Phys. 2007, 126, 044101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaelian, K. Thermodynamic dissipation theory for the origin of life. Earth Syst. Dynam. 2011, 2, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Mirazo, K.; Briones, C.; de la Escosura, A. Chemial roots of biological evolution: The origins of life as a process of development of autonomous functional systems. Open Biol. 2017, 7, 170050. [Google Scholar] [CrossRef] [PubMed]
- Schuster, P.; Stadler, P.F. Networks in chemical evolution. Complexity 2001, 8, 34–42. [Google Scholar] [CrossRef]
- Polettini, M.; Esposito, M. Irreversible thermodynamics of open chemical networks. J. Chem. Phys. 2014, 141, 024117. [Google Scholar] [CrossRef]
- Eschenmoser, A.; Kisakürek, M.V. Chemistry and the Origin of Life. Helv. Chim. Acta 1996, 79, 1249–1259. [Google Scholar] [CrossRef]
- Barron, L.D. Symmetry and molecular chirality. Chem. Soc. Rev. 1986, 15, 189–223. [Google Scholar] [CrossRef]
- Avalos, M.; Babiano, R.; Cintas, P.; Jimenez, J.L.; Palacios, J.C.; Barron, L. Absolute asymmetric synthesis under physical fields: Facts and fictions. Chem. Rev. 1998, 98, 2391–2404. [Google Scholar] [CrossRef]
- Feringa, B.L.; van Delden, R.A. Absolute asymmetric synthesis: The origin, control, and amplification of chirality. Angew. Chem. Int. Ed. 1999, 38, 3418–3438. [Google Scholar] [CrossRef]
- Gu, H.; Nakamura, Y.; Sato, T.; Teramoto, A.; Green, M.M.; Jha, S.K.; Andreola, C.; Reidy, M.P.; Mark, H.F. Optical Rotation of Random Copolyisocyanates of Chiral and Achiral Monomers: Sergeant and Soldier Copolymers. Macromolecules 1998, 31, 6362–6368. [Google Scholar] [CrossRef]
- Guillaneux, D.; Zhao, S.H.; Samuel, O.; Rainford, D.; Kagan, E.B. Nonlinear Effects in Asymmetric Catalysis. J. Am. Chem. Soc. 1994, 116, 9430–9439. [Google Scholar] [CrossRef]
- Root-Bernstein, R. Simultaneous origin of homochirality, the genetic code and its directionality. BioEssays 2007, 29, 689–698. [Google Scholar] [CrossRef]
- Wu, M.; Walker, S.I.; Higgs, P.G. Autocatalytic replication and homochirality in biopolymers: Is homochirality a requierment of life or a result of it? Astrobiology 2012, 12, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Schuster, P. Germany-Japan round table. Heidelberg. 2011. Available online: https://www.tbi.univie.ac.at/~pks/ (accessed on 30 May 2019).
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoric amino acids. Science 1997, 275, 951–955. [Google Scholar] [CrossRef]
- McGuire, A.B.; Carroll, P.B.; Loomis, R.A.; Finneran, I.A.; Jewell, P.R.; Remijan, A.J.; Blake, G.A. Discovery of the interstellar chiral molecule propylene oxide (CH3CHCH2O). Science 2016, 352, 1449–1452. [Google Scholar] [CrossRef]
- Oró, J.; Mills, T.; Lazcano, A. Comets and the formation of biochemical compounds on the primitive Earth–A review. Orig. Life Evol. Biosph. 1991, 21, 267–277. [Google Scholar] [CrossRef]
- Kempe, S.; Kazmierczak, J. Biogenesis and early life on Earth and Europa: Favored by an alkaline ocean? Astrobiology 2002, 2, 123–130. [Google Scholar] [CrossRef]
- Brack, A. Clay minerals and the origin of life. In Handbook of Clay Science, Developments in Clay Science; Bergaya, F., Theng, B.K.G., Lagaly, G., Eds.; Elsevier: Oxford, UK, 2006; Volume 1, pp. 379–391. [Google Scholar]
- Mislow, K. Absolute asymmetric synthesis: A commentary. Collect. Czech. Chem. Commun. 2003, 68, 849–864. [Google Scholar] [CrossRef]
- Mills, W.H. Some aspects of stereochemsitry. Chem. Ind. (Lond.) 1932, 51, 750–759. [Google Scholar] [CrossRef]
- Barabás, B.; Kurdi, R.; Pályi, G. Natural abundance isotopic chirality in the reagents of the soai reaction. Symmetry 2016, 8, 2. [Google Scholar] [CrossRef]
- Bolli, M.; Micura, R.; Eschenmoser, A. Pyranosyl-RNA: Chiroselective self-assembly of base sequences by ligative oligomerization of tetranucleotide-2′,3′-cyclophosphates (with a commentary concerning the origin of biomolecular homochirality). Chem. Biol. 1997, 4, 309–320. [Google Scholar] [CrossRef]
- Ben-Naim, B. Entropy Demystified: The Second Law of Thermodynamics Reduced to Plain Common Sense; World Scientific: Singapore, 2015. [Google Scholar]
- Plasson, R.; Kondepudi, D.K.; Bersini, H.; Commeyras, A.; Asakura, K. Emergence of homochirality in far-from-equilibrium systems: Mechanisms and role in prebiotic chemistry. Chirality 2007, 19, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Ribo, J.M.; Blanco, C.; Crusats, J.; El-Hachemi, Z.; Hochberg, D.; Moyano, A. Absolute asymmetric synthesis in enantioselective autocatalytic reaction networks: Theoretical games, speculations on chemical evolution and perhaps a synthetic option. Chem. Eur. J. 2014, 20, 17250–17271. [Google Scholar] [CrossRef]
- Ross, J.; Vlad, M. Exact solutions for the entropy production rate of several irreversible processes. J. Phys. Chem. A 2005, 109, 10607–10612. [Google Scholar] [CrossRef]
- Hochberg, D.; Ribó, J.M. Stochiometric network analysis of entropy production in reaction networks. Phys. Chem. Chem. Phys. 2018, 20, 23726–23739. [Google Scholar] [CrossRef]
- Hochberg, D.; Ribó, J.M. Entropic analysis of mirror symmetry breaking in chiral hypercycles. Life 2019, 9, 28. [Google Scholar] [CrossRef]
- Werth, H. Über Irreversibilität, Naturprozesse und Zeitstrukture. In Offene Systeme I; Weizsäcker, E., Ed.; Klett-Cotta: Stuttgard, Germany, 1974; pp. 114–199. [Google Scholar]
- Kondepudi, D.; Kapcha, K. Entropy production in chiral symmetry breaking transitions. Chirality 2008, 20, 524–528. [Google Scholar] [CrossRef]
- Kondepudi, D.; Prigogine, I. Modern Thermodynamics, 2nd ed.; John Wiley & Sons: Chichester, UK, 2015. [Google Scholar]
- Plasson, R.; Bersini, H. Energetic analysis of mirror symmetry breaking processes in a recycled microreversible chemical system. J. Phys. Chem. B 2008, 113, 3477–3490. [Google Scholar] [CrossRef]
- Mauksch, M.; Tsogoeva, S.B. Spontaneous Emergence of Homochirality via Coherently Coupled Antagonistic and Reversible Reaction Cycles. ChemPhysChem 2008, 9, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Plasson, R.; Brandenburg, A.; Jullient, L.; Bersini, H. Autocatalyses. J. Phys. Chem. A 2011, 115, 8073–8085. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, D.; Bourdon García, R.D.; Ägreda Bastidas, J.A.; Ribó, J.M. Stoichiometric network analysis of spontaneous mirror symmetry breaking in chemical reactions. Phys. Chem. Chem. Phys. 2017, 19, 17618–17636. [Google Scholar] [CrossRef] [PubMed]
- Gellman, A.J.; Ernst, K.-H. Chiral autocatalysis and mirror symmetry breaking. Catal. Lett. 2018, 148, 1610. [Google Scholar] [CrossRef]
- Ribó, J.M.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Hochberg, D. Spontaneous mirror symmetry breaking in heterocatalytically coupled enantioselective replicators. Chem. Sci. 2017, 8, 763–769. [Google Scholar]
- Szathmary, E. The origin of replicators and reproducers. Philos. Trans. R. Soc. Lond. Ser. B 2006, 361, 1761–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, F.C. On spontaneous asymmetric synthesis. Biochim. Biophys. Acta 1953, 11, 459–463. [Google Scholar] [CrossRef]
- Ribó, J.M.; Hochberg, D. Competitive exclusion principle in ecology and absolute asymmetric synthesis in chemistry. Chirality 2015, 27, 722–727. [Google Scholar] [CrossRef]
- Rivera Islas, J.; Lavabre, D.; Grevy, J.-M.; Hernández Lamoneda, R.; Rojas Cabrera, K.; Micheau, J.C.; Buhse, T. Mirror-symmetry breaking in the Soai reaction: A kinetic understanding. Proc. Natl. Acad. Sci. USA 2005, 102, 13743–13748. [Google Scholar] [CrossRef] [Green Version]
- Lavabre, D.; Micheau, J.-C.; Islas, J.R.; Buhse, T. Kinetic Insight into specific features of the autocatalytic Soai reaction. In Amplification of Chirality; Springer: Berlin, Germany, 2008; Volume 284, pp. 67–96. [Google Scholar]
- Crusats, J.; Hochberg, D.; Moyano, A.; Ribó, J.M. Frank models and spontaneous emergence of chirality in closed systems. ChemPhysChem 2009, 10, 2123–2131. [Google Scholar] [CrossRef]
- Avetisov, V.; Goldanskii, V. Mirror symmetry breaking at the molecular level. Proc. Natl. Acad. Sci. USA 1996, 93, 11435–11442. [Google Scholar] [CrossRef] [PubMed]
- Ribó, J.M.; Hochberg, D. Stability of racemic and chiral steady states in open and closed chemical systems. Phys. Lett. A 2008, 373, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Blanco, C.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Hochberg, D.; Ribó, J.M. Spontaneous Emergence of Chirality in the Limited Enantioselectivity Model: Autocatalytic Cycle Driven by an External Reagent. ChemPhysChem 2013, 14, 2432–2440. [Google Scholar] [CrossRef] [PubMed]
- Ribó, J.M.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Blanco, C.; Hochberg, D. Spontaneous Mirror Symmetry Breaking in the Limited Enantioselective Autocatalysis Model: Abyssal Hydrothermal Vents as Scenario for the Emergence of Chirality in Prebiotic Chemistry. Astrobiology 2013, 13, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Blanco, C.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Veintemillas-Verdaguer, S.; Hochberg, D.; Ribó, J.M. The Viedma Deracemization of Racemic Conglomerate Mixtures as a Paradigm of Spontaneous Mirror Symmetry Breaking in Aggregation and Polymerization. ChemPhysChem 2013, 14, 3982–3993. [Google Scholar] [CrossRef]
- El-Hachemi, Z.; Crusats, Z.; Ribó, J.M.; Veintemillas-Verdaguer, S. Spontaneous transition toward chirality in the NaClO3 crystallization in boiling solutions. Cryst. Growth Des. 2009, 9, 4802–4806. [Google Scholar] [CrossRef]
- Cintas, P.; Viedma, C. Homochirality beyond grinding: Deracemizing chiral crystals by temperature gradient under boiling. Chem. Commun. 2011, 47, 12786–12788. [Google Scholar] [CrossRef]
- Crusats, J.; Veintemillas-Verdaguer, S.; Ribó, J.M. Homochirality as a Consequence of Thermodynamic Equilibrium? Chem. Eur. J. 2006, 12, 7776–7781. [Google Scholar] [CrossRef]
- El-Hachemi, Z.; Arteaga, O.; Canillas, A.; Crusats, J.; Sorrenti, A.; Veintemillas-Verdaguer, S. Achiral-to-chiral transition in benzil solidification: Analogies with racemic conglomerates systems showing deracemization. Chirality 2013, 25, 393–399. [Google Scholar] [CrossRef]
- Viedma, C. Fighting fire with fire: Racemization drives deracemization. In Proceedings of the Chiral Symmetry Breaking at Molecular Level, Solvay Workshop, Brussels, Belgium, 28–30 November 2018. [Google Scholar]
- Kondepudi, D.K.; Kauffman, R.; Singh, N. Chiral symmetry breaking in sodium chlorate crystallization. Science 1990, 250, 975–976. [Google Scholar] [CrossRef]
- Kondepudi, D.K.; Asakura, K.; Laudadio, J. Chiral symmetry breaking in stirred crystallization of 1,1′-binaphthyl melt. J. Am. Chem. Soc. 1999, 121, 1448–1451. [Google Scholar] [CrossRef]
- Ribó, J.M.; El-Hachemi, Z.; Arteaga, O.; Canillas, A.; Crusats, J. Hydrodynamic Effects in Soft-Matter Self-Assembly: The Case of J-Aggregates of Amphiphilic Porphyrins. Chem. Record. 2017, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lv, B.; Duan, P.; Liu, M.; Yin, M. Stoichiometry-controlled inversion of circulaly polarized luminiscence in co-assembly of chiral gelators with achiral tetraphenylethylene derivative. Chem. Commun. 2019, 55, 2194–2197. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, S.C.; Cafferty, B.J.; Weigert-Muñoz, A.; Schuster, G.B.; Hud, N.V. Spontaneous Symmetry Breaking in the Formation of Supramolecular Polymers: Implications for the Origin of Biological Homochirality. Angew. Chem. Int. Ed. 2019, 58, 1453–1457. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.; Stich, M.; Hochberg, D. Mechanically induced homochirality in nucleated enantioselective polymerization. J. Phys. Chem. B 2017, 121, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Saghatelian, A.; Yokobayashi, Y.; Soltaini, K.; Gadhiri, M.R. A chiroselective peptide replicator. Nature 2001, 409, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Eschenmoser, A. The search of the chemistry of life’s origin. Tetrahedron 2001, 63, 12821–12844. [Google Scholar] [CrossRef]
- Monnard, P.-A.; Walde, P. Current ideas about prebiological compartmentalization. Life 2015, 5, 1239–1262. [Google Scholar] [CrossRef]
- Kauffman, S. At Home in the Universe; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Kondepudi, D.K.; Prigogine, I.; Nelson, G. Sensitivity of branch selection in nonequilibrium systems. Phys. Lett. A 1985, 111, 29–32. [Google Scholar] [CrossRef]
- Avetisov, V.A.; Kuz’min, V.V.; Anikin, S.A. Sensitivity of chemical chiral systems to weak asymmetric factors. Chem. Phys. 1987, 112, 179–187. [Google Scholar] [CrossRef]
- Pitsch, S.; Wendeborn, S.; Jaun, B.; Eschenmoser, A. Why Pentose- and Not Hexose-Nucleic Acids? Helvetica Chim. Acta 1993, 76, 2161–2183. [Google Scholar] [CrossRef]
- Shapiro, R. Prebiotic ribose synthesis: A critical analysis. Orig. Life Evol. Biosph. 1988, 18, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Jedrzejas, M.J. Carbohydrate polymers at the center of life’s origin. Chem. Rev. 2008, 108, 50161–55085. [Google Scholar] [CrossRef] [PubMed]
- Pestunova, O.P.; Simonov, A.N.; Suytnikov, V.N.; Parmon, V.N. Prebiotic carbohydrates and their derivatives. In Biosphere Origin and Evolution; Dobretsov, N., Kolchanov, N., Rozanov, A., Zavarzin, G., Eds.; Springer: New York, NY, USA, 2008; pp. 103–118. [Google Scholar]
- Orgel, L.E. Self-organized metabolic cycles. Proc. Natl. Acad. Sci. USA 2000, 97, 12503–12507. [Google Scholar] [CrossRef] [PubMed]
- Sel’kov, E.E. Self-oscillations in glycolysis. Eur. J. Biochem. 1968, 4, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Plasson, R.; Bersini, H.; Commeyras, A. Recycling Frank: Spontaneous emergence of homochirality in noncatalytic systems. Proc. Natl. Acad. Sci. USA 2004, 101, 16733–16738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stich, M.; Blanco, C.; Hochberg, C. Chiral and chemical oscillations in a simple dimerization model. Phys. Chem. Chem. Phys. 2013, 15, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. USA 2010, 107, 2763–2768. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, R. Life’s Biological Chemistry: A Destiny or destination starting from prebiotic chemistry? Chem. Eur. J. 2018, 24, 16708–16715. [Google Scholar] [CrossRef]
- Peretó, J.; López-García, P.; Moreira, D. Ancestral lipid biosynthesis and early membrane evolution. Trends Biochem. Sci. 2004, 29, 469–477. [Google Scholar] [CrossRef]
- Davankov, V. The chirality as an inherent general property of matter. Chirality 2006, 18, 459–461. [Google Scholar] [CrossRef] [PubMed]
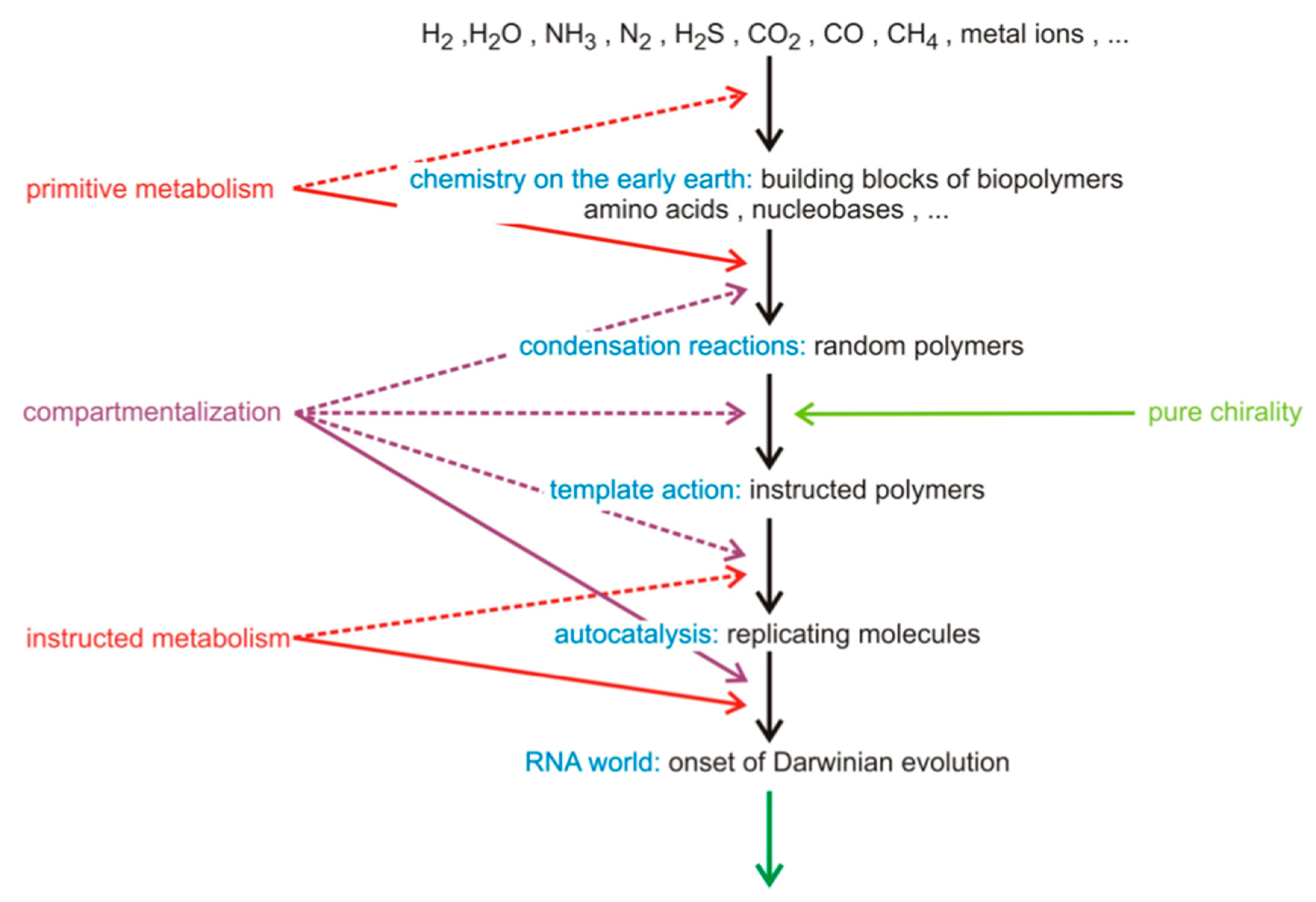
Figure 1.
Presently, most accepted stages of a scheme for a reasonable hierarchical order of the increase of complexity during chemical evolution. The scheme assumes the unreasonable existence of enantiopure pools of chemical building blocks for the formation of the chiral polymers of life. Reproduced with author’s permission from reference [34].
Figure 1.
Presently, most accepted stages of a scheme for a reasonable hierarchical order of the increase of complexity during chemical evolution. The scheme assumes the unreasonable existence of enantiopure pools of chemical building blocks for the formation of the chiral polymers of life. Reproduced with author’s permission from reference [34].

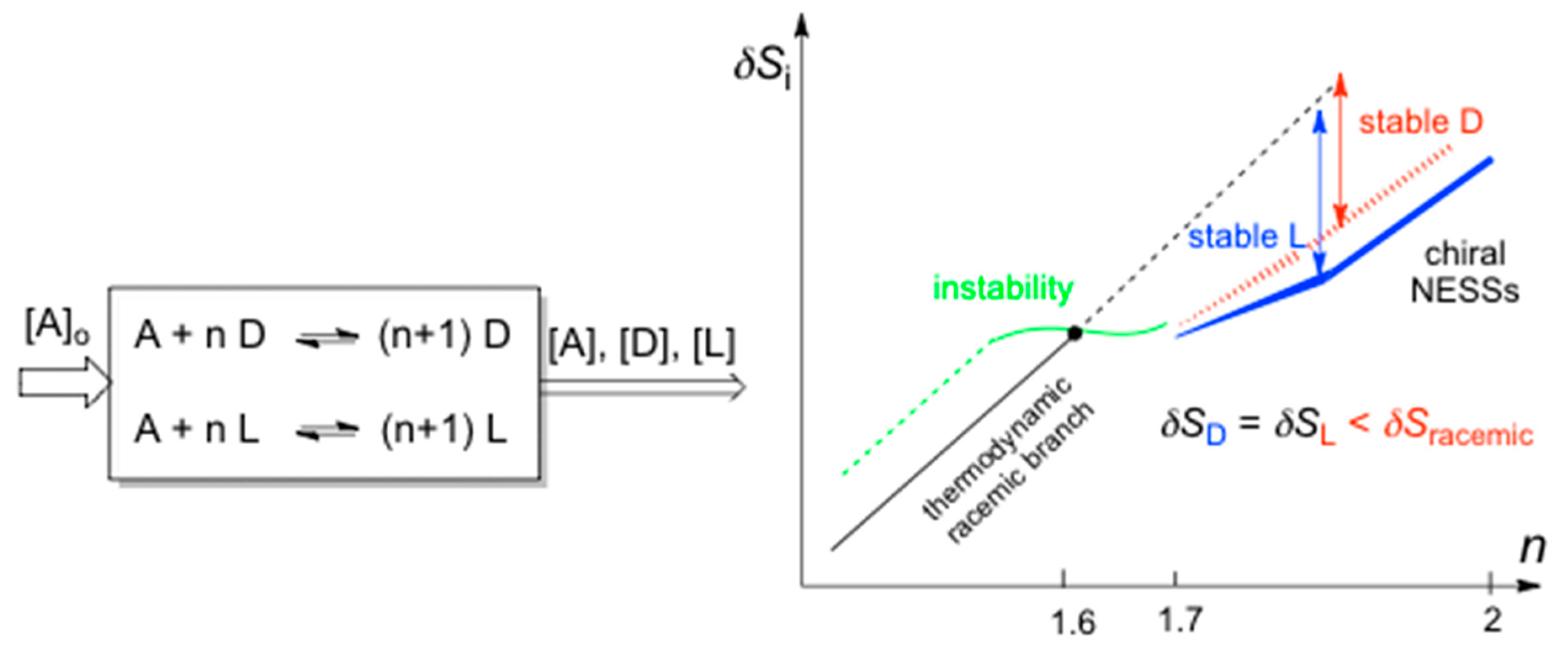
Figure 2.
Spontaneous mirror symmetry breaking (SMSB) emerging in an open system for selective enantiocatalysis, where A is the achiral resource and D and L are enantiomers (initial concentrations [D]o and [L]o ≠ 0). SMSB does not take place for n = 1. Metastability of the racemic thermodynamic branch can only occur for values of n > 1. For reasonable reaction rate constants and reaction parameters (not specified here) in this example instability of the racemic branch occurs for n > 1.6 and SMSB for n ≥ 1.7. Non-integer values of n correspond to the simplification of complex autocatalytic networks [56], which leads to the same dynamic change of the solution species.
Figure 2.
Spontaneous mirror symmetry breaking (SMSB) emerging in an open system for selective enantiocatalysis, where A is the achiral resource and D and L are enantiomers (initial concentrations [D]o and [L]o ≠ 0). SMSB does not take place for n = 1. Metastability of the racemic thermodynamic branch can only occur for values of n > 1. For reasonable reaction rate constants and reaction parameters (not specified here) in this example instability of the racemic branch occurs for n > 1.6 and SMSB for n ≥ 1.7. Non-integer values of n correspond to the simplification of complex autocatalytic networks [56], which leads to the same dynamic change of the solution species.

Figure 3.
Reaction networks (simplified) that may lead to SMSB processes, both by enantioselective transformations from achiral to chiral compounds a), b), and c), or through the deracemization of racemic mixtures d).
Figure 3.
Reaction networks (simplified) that may lead to SMSB processes, both by enantioselective transformations from achiral to chiral compounds a), b), and c), or through the deracemization of racemic mixtures d).

Figure 4.
The compartmentalization of the different worlds proposed for the origin of life, would determine that the driving force of SMSB at the interconnected nucleic acid, protein and carbohydrate domain preserves biological homochirality (BH) and determines the chiral sign of all the rest of the enantioselective reactions and chiral transfer interactions.
Figure 4.
The compartmentalization of the different worlds proposed for the origin of life, would determine that the driving force of SMSB at the interconnected nucleic acid, protein and carbohydrate domain preserves biological homochirality (BH) and determines the chiral sign of all the rest of the enantioselective reactions and chiral transfer interactions.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ribó, J.M.; Hochberg, D. Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth. Symmetry 2019, 11, 814. https://doi.org/10.3390/sym11060814
AMA Style
Ribó JM, Hochberg D. Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth. Symmetry. 2019; 11(6):814. https://doi.org/10.3390/sym11060814
Chicago/Turabian StyleRibó, Josep M., and David Hochberg. 2019. "Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth" Symmetry 11, no. 6: 814. https://doi.org/10.3390/sym11060814
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.