
Directional Bilateral Asymmetry in Fish Otolith: A Potential Tool to Evaluate Stock Boundaries?

 ,
,  , , ,
, , ,  , and
, and
Abstract
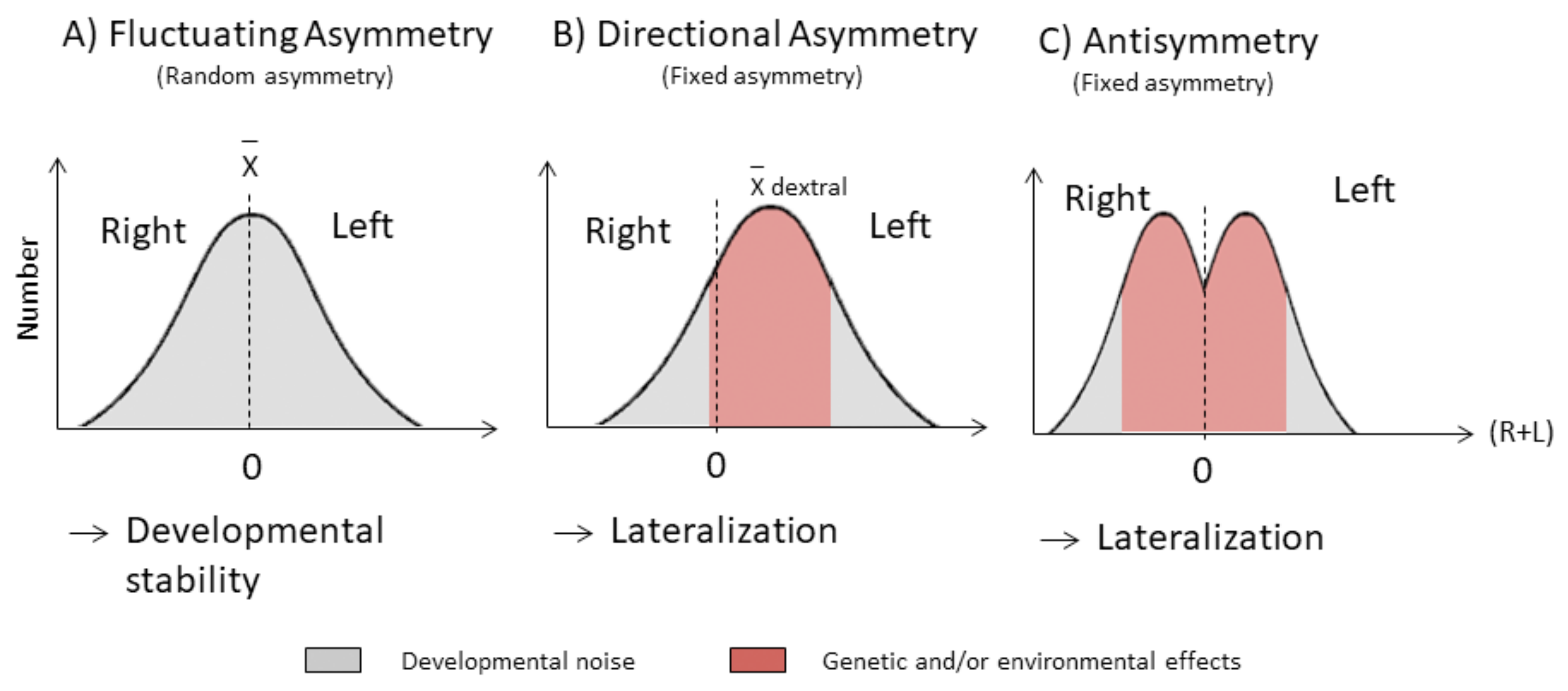
:1. Introduction
2. Materials and Methods
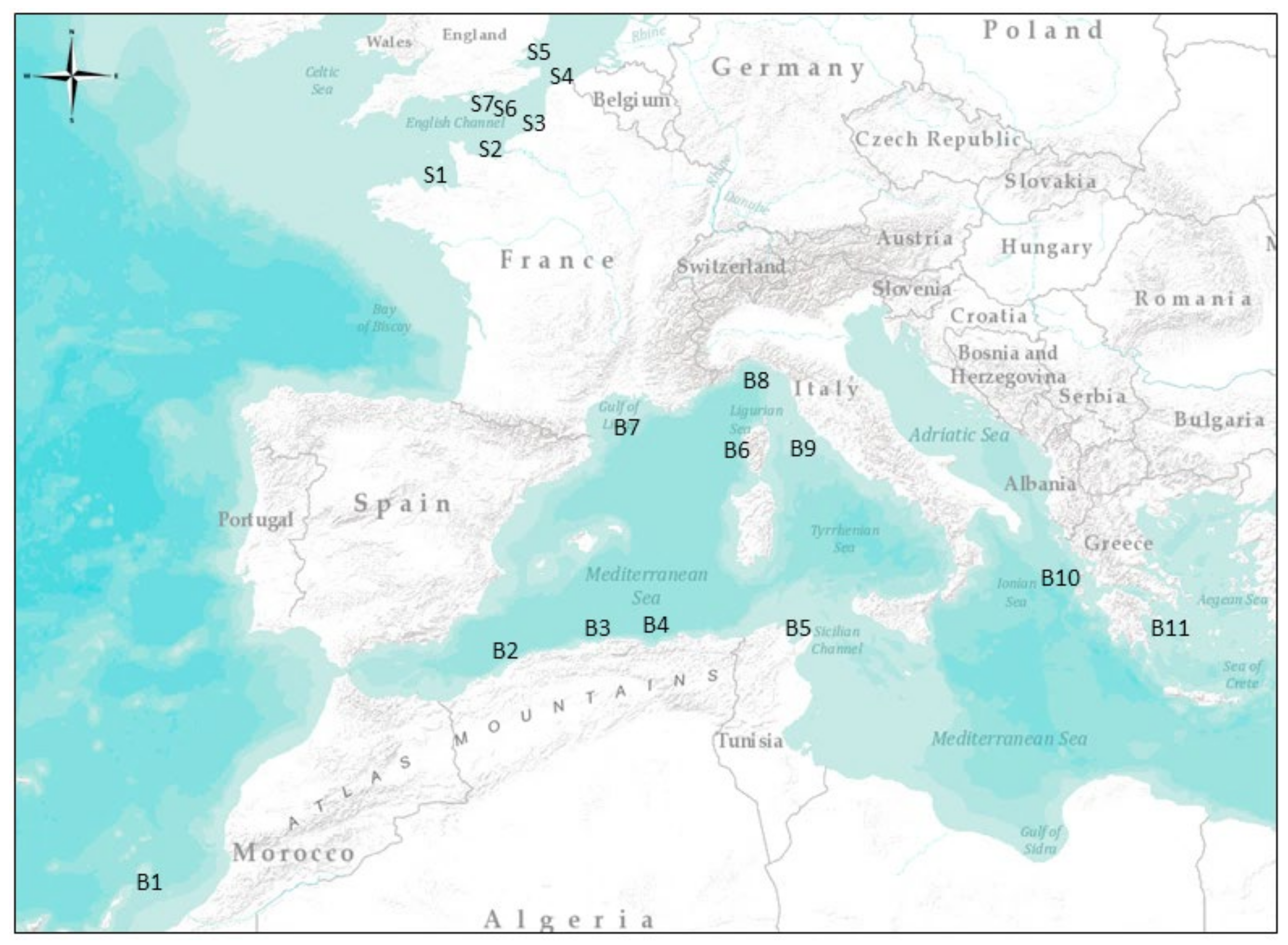
2.1. Sample Collection
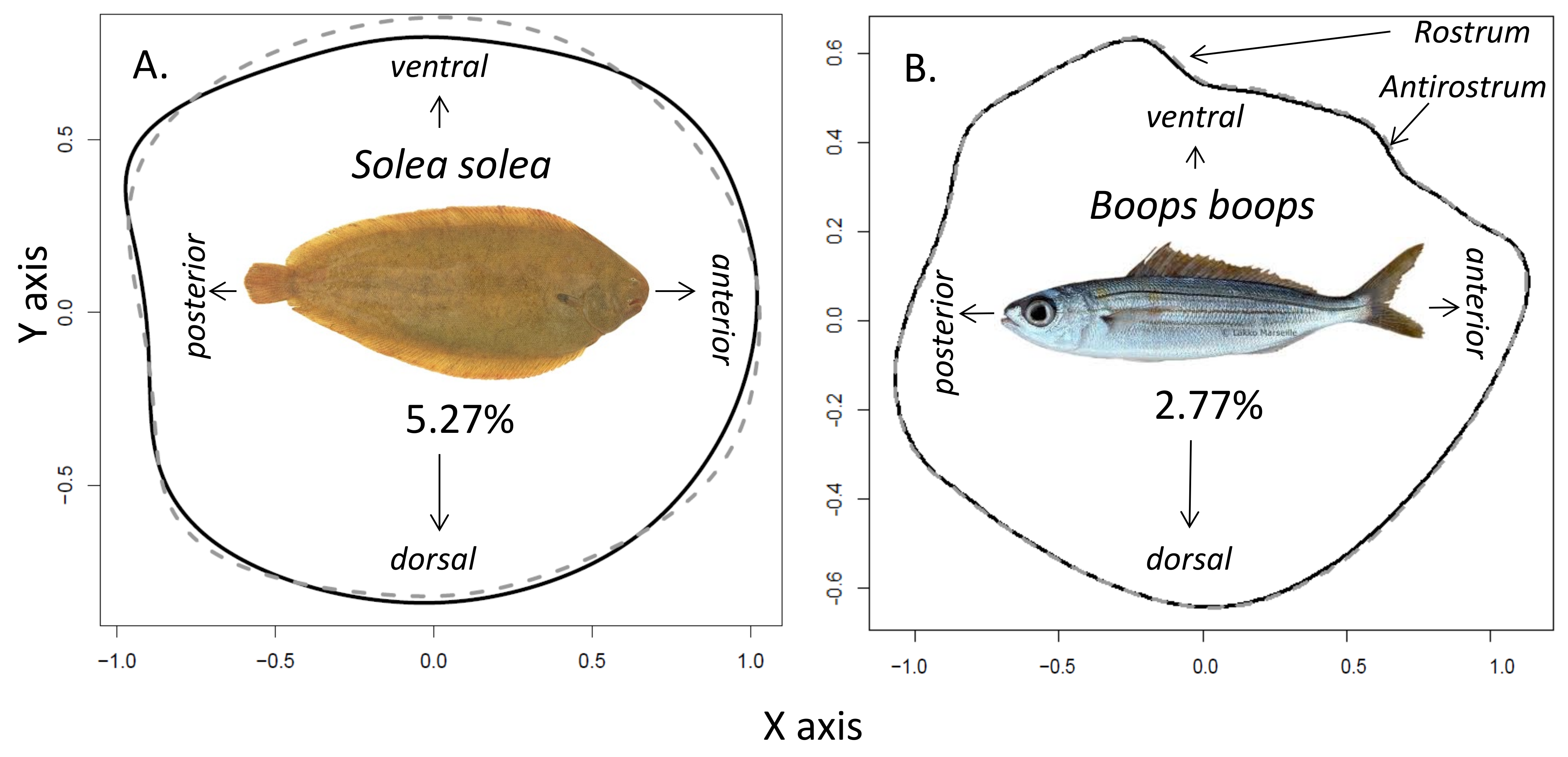
2.2. Otolith Shape Analysis
2.3. Statistical Analyses
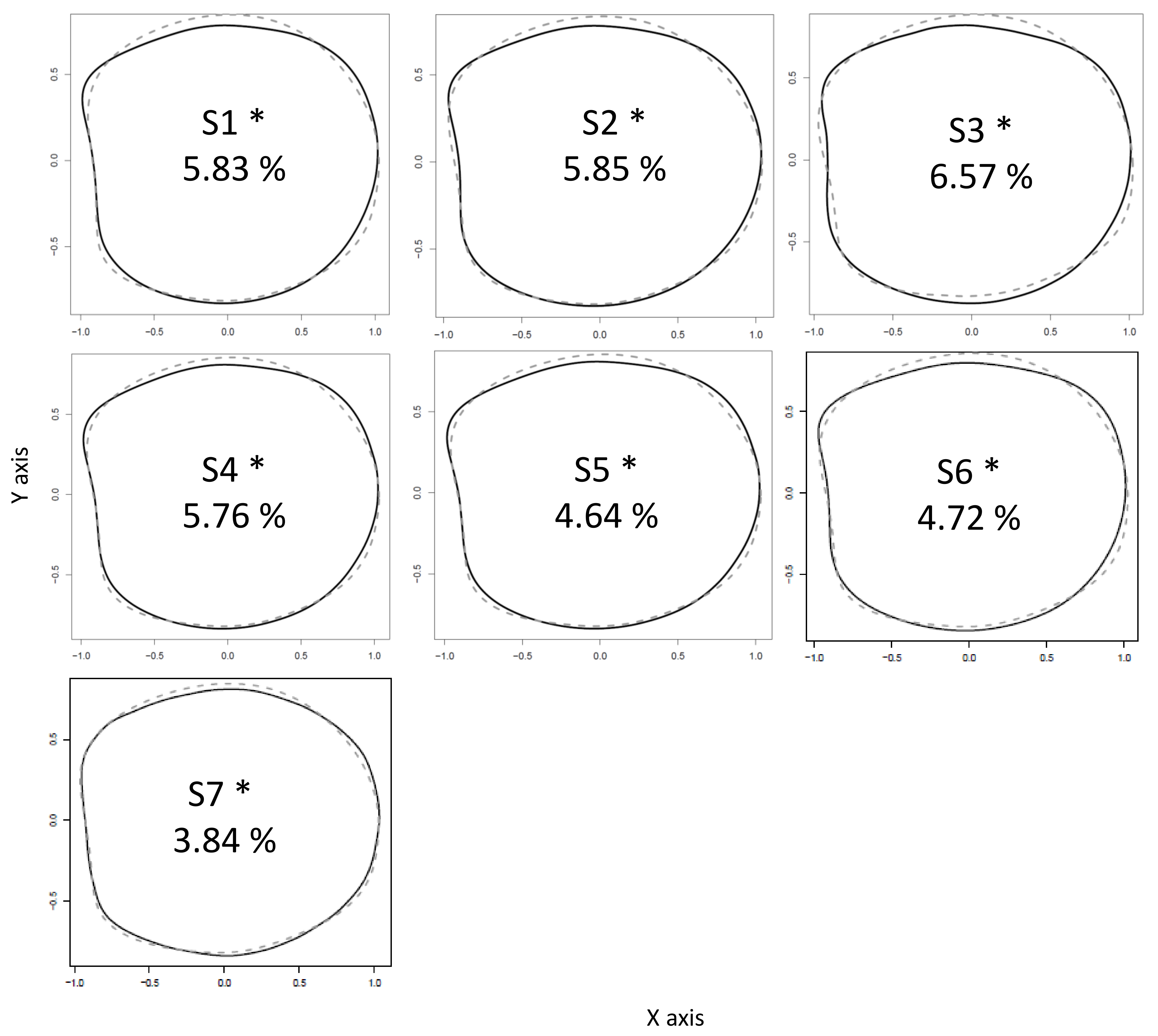
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campana, S.E. Chemistry and composition of fish otoliths: Pathways, mechanisms and applications. Mar. Ecol. Prog. Ser. 1999, 188, 263–297. [Google Scholar] [CrossRef] [Green Version]
- Campana, S.E.; Thorrold, S.R. Otoliths, increments, and elements: Keys to a comprehensive understanding of fish populations? Can. J. Fish. Aquat. Sci. 2001, 58, 30–38. [Google Scholar] [CrossRef]
- Panfili, J.; Pontual, H.de.; Troadec, H.; Wright, P.J. Manual of Fish Sclerochronology; Coédition Ifremer-IRD: Brest, France, 2002. [Google Scholar]
- Casselman, J.M. Determination of age and growth. In The Biology of Fish Growth; Weatherley, A.H., Gill, H.S., Eds.; Academic Press: New York, NY, USA, 1987; pp. 209–242. [Google Scholar]
- Campana, S.E.; Casselman, J.M. Stock discrimination using otolith shape analysis. Can. J. Fish. Aquat. Sci. 1993, 50, 1062–1083. [Google Scholar] [CrossRef]
- Reiss, H.; Hoarau, G.; Dickey-Collas, M.; Wolff, W. Genetic population structure of marine fish: Mismatch between biological and fisheries management units. Fish Fish. 2009, 10, 361–395. [Google Scholar] [CrossRef]
- Tuset, V.M. Use of otolith shape for stock identification of John’s snapper, Lutjanus johnii (Pisces: Lutjanidae), from the Persian Gulf and the Oman Sea. Fish. Res. 2014, 155, 59–63. [Google Scholar]
- Ponton, D. Is geometric morphometrics efficient for comparing otolith shape of different fish species? J. Morphol. 2006, 267, 750–757. [Google Scholar] [CrossRef]
- Ramirez-Pérez, J.S.; Quiñónez-Velázquez, C.; García-Rodríguez, F.J.; Felix-Uraga, R.; Melo-Barrera, F.N. Using the shape of sagitta otoliths in the discrimination of phenotypic stocks in Scomberomorus sierra (Jordan and Starks, 1895). Can. J. Fish. Aquat. Sci. 2010, 5, 82–93. [Google Scholar]
- Vergara-Solana, F.J.; García-Rodríguez, F.J.; De La Cruz-Agüero, J. Comparing body and otolith shape for stock discrimination of Pacific sardine, Sardinops sagax Jenyns, 1842. J. Appl. Ichthyol. 2013, 29, 1241–1246. [Google Scholar] [CrossRef]
- Sadighzadeh, Z.; Valinassab, T.; Vosugi, G.; Motallebi, A.A.; Fatemi, M.R.; Lombarte, A.; Tuset, V.M.; Lozano, I.J.; Gonzalez, J.A.; Pertusa, J.F.; et al. Shape indices to identify regional differences in otolith morphology of comber, Serranus cabrilla (L., 1758). J. Appl. Ichthyol. 2003, 19, 88–93. [Google Scholar]
- Parisi-Baradad, V.; Lombarte, A.; García-Ladona, E.; Cabestany, J.; Piera, J.; Chic, Ò. Otolith shape contour analysis using affine transformation invariant wavelet transforms and curvature scale space representation. Mar. Freshw. Res. 2005, 56, 795–804. [Google Scholar] [CrossRef]
- Benzinou, A.; Carbini, S.; Nasreddine, K.; Elleboode, R.; Mahé, K. Discriminating stocks of striped red mullet (Mullus surmuletus) in the Northwest European seas using three automatic shape classification methods. Fish. Res. 2013, 143, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Castonguay, M.; Simard, P.; Gagnon, P. Usefulness of Fourier analysis of otolith shape for Atlantic mackerel (Scomber scombrus) stock discrimination. Can. J. Fish. Aquat. Sci. 1991, 48, 296–302. [Google Scholar] [CrossRef]
- Lombarte, A.; Lleonart, J. Otolith size changes related with body growth, habitat depth and temperature. Environ. Biol. Fishes 1993, 37, 297–306. [Google Scholar] [CrossRef]
- Begg, G.A.; Brown, R.W. Stock identification of haddock Melanogrammus aeglefinus on Georges Bank based on otolith shape analysis. Trans. Am. Fish. Soc. 2000, 129, 935–945. [Google Scholar] [CrossRef]
- Cadrin, S.X.; Friedland, K.D. The utility of image processing techniques for morphometric analysis and stock identification. Fish. Res. 1999, 43, 129–139. [Google Scholar] [CrossRef]
- Capoccioni, F.; Costa, C.; Aguzzi, J.; Menesatti, P.; Lombarte, A.; Ciccotti, E. Ontogenetic and environmental effects on otolith shape variability in three Mediterranean European eel (Anguilla anguilla, L.) populations. J. Exp. Mar. Biol. Ecol. 2011, 397, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, M.; Doerin-Arjes, P.; Kastowsky, M.; Mosegaard, H. Effects of sex, stock, and environment on the shape of known-age Atlantic cod (Gadus morhua) otoliths. Can. J. Fish. Aquat. Sci. 2004, 61, 158–167. [Google Scholar] [CrossRef]
- Gagliano, M.; McCormick, M.I. Feeding history influences otolith shape in tropical fish. Mar. Ecol. Prog. Ser. 2004, 278, 291–296. [Google Scholar] [CrossRef]
- Hüssy, K. Otolith shape in juvenile cod (Gadus morhua): Ontogenetic and environmental effects. J. Exp. Mar. Biol. Ecol. 2008, 364, 35–41. [Google Scholar] [CrossRef]
- Mérigot, B.; Letourneur, Y.; Lecomte-Finiger, R. Characterization of local populations of the common sole Solea solea (Pisces, Soleidae) in the NW Mediterranean through otolith morphometrics and shape analysis. Mar. Biol. 2007, 151, 997–1008. [Google Scholar] [CrossRef]
- Mille, T.; Mahé, K.; Cachera, M.; Villanueva, C.M.; De Pontual, H.; Ernande, B. Diet is correlated with otolith shape in marine fish. Mar. Ecol. Prog. Ser. 2016, 555, 167–184. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, L.R.; Di Beneditto, A.P.M.; Guillermo, L.H.; Rivera, L.A. Allometric changes and shape differentiation of sagitta otoliths in sciaenid fishes. Fish. Res. 2005, 74, 288–299. [Google Scholar] [CrossRef]
- Swan, S.C.; Geffen, A.J.; Morales-Nin, B.; Gordon, J.D.M.; Shimmield, T.; Sawyer, T.; Massutí, E. Otolith chemistry: An aid to stock separation of Helicolenus dactylopterus (bluemouth) and Merluccius merluccius (European hake) in the Northeast Atlantic and Mediterranean. ICES J. Mar. Sci. 2006, 63, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Vignon, M.; Morat, F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish. Mar. Ecol. Prog. Ser. 2010, 411, 231–241. [Google Scholar] [CrossRef]
- Díaz-Gil, C.; Palmer, M.; Catalán, I.A.; Alós, J.; Fuiman, L.A.; García, E.; del Mar Gil, M.; Grau, A.; Kang, A.; Maneja, R.H.; et al. Otolith fluctuating asymmetry: A misconception of its biological relevance? ICES J. Mar. Sci. 2015, 72, 2079–2089. [Google Scholar] [CrossRef]
- Lemberget, T.; Mccormick, M.I. Replenishment success linked to fluctuating asymmetry in larval fish. Oecologia 2009, 159, 83–93. [Google Scholar] [CrossRef]
- Dowhower, J.F.; Blumer, L.S.; Lejeune, P.; Gaudin, P.; Marconato, A.; Bisazza, A. Otolith asymmetry in cottus bairdi and cottus gobio. Pol. Arch. Hydrobiol. 1990, 37, 209–220. [Google Scholar]
- Green, A.A.; Mosaliganti, K.R.; Swinburne, I.A.; Obholzer, N.D.; Megason, S.G. Recovery of Shape and Size in a Developing Organ Pair. Dev. Dyn. 2017, 246, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, A.; Coppin, F.; Curet, L.; Dauvin, J.C.; Delavenne, J.; Dewarumez, J.M.; Dupuis, L.; Foveau, A.; Garcia, C.; Gardel, L.; et al. Atlas des Habitats des Ressources Marines de la Manche Orientale—CHARM II/Channel Habitat Atlas for Marine Resource Management—CHARM II. PROGRAMME INTERREG 3A; IFREMER: Boulogne-sur-mer, France, 2009. [Google Scholar]
- FAO. The State of Mediterranean and Black Sea Fisheries 2016 (SoMFi 2016); Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Lestrel, P.E. Fourier Descriptors and Their Applications in Biology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Kuhl, F.; Giardina, C. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Archie, J.W. A Comparison of Fourier Methods for the Description of Wing Shape in Mosquitoes (Diptera: Culicidae). Syst. Biol. 1984, 33, 302–317. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L.F.J. Numerical Ecology, 2nd ed.; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, H.M.H.; Wagner, H. Vegan: Community Ecology Package. In R Package Version 2.0–10; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Bivand, R.S.; Pebesma, E.; Gomez-Rubio, V. Applied Spatial Data Analysis with R, 2nd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Torres, G.J.; Lombarte, A.; Morales-Nin, B. Sagittal otolith size and shape variability to identify geographical intraspecific differences in three species of genus Merluccius. J. Mar. Biol. Assoc. UK 2000, 80, 333–342. [Google Scholar] [CrossRef]
- Smith, M.K. Regional differences in otolith morphology of the deep slope red snappers Etelis carbunculus. Can. J. Fish. Aquat. Sci. 1992, 49, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Bird, J.L.; Eppler, D.T.; Checkley, D.M. Comparisons of herring otoliths using Fourier series shape analysis. Can. J. Fish. Aquat. Sci. 1986, 43, 1228–1234. [Google Scholar] [CrossRef]
- Bolles, K.L.; Begg, G.A. Distinction between silver hake (Merluccius bilineariz) stocks in U.S. waters of the northwest Atlantic using whole otolith morphometric. Fish. Bull. 2000, 98, 451–462. [Google Scholar]
- Mapp, J.; Hunter, E.; Van Der Kooijc, J.; Songer, S.; Fisher, M. Otolith shape and size: The importance of age when determining indices for fish-stock separation. Fish. Res. 2017, 190, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.R. Animal asymmetry. Curr. Biol. 2009, 19, 473–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerbach, B.M.; Ruff, C.B. Limb bone bilateral asymmetry: Variability and commonality among modern humans. J. Hum. Evol. 2006, 50, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Ambrose, S.H. Paleolithic technology and human evolution. Science 2001, 291, 1748–1753. [Google Scholar] [CrossRef]
- Lychakov, D. Behavioral lateralization and otolith asymmetry. J. Evol. Biochem. Physiol. 2013, 49, 441–456. [Google Scholar] [CrossRef]
- Lychakov, D.V.; Rebane, Y.T. Fish otolith mass asymmetry: Morphometry and influence on acoustic functionality. Hear. Res. 2005, 201, 55–69. [Google Scholar] [CrossRef]
- Lychakov, D.V.; Rebane, Y.T.; Lombarte, A.; Demestre, M.; Fuiman, L.A. Saccular otolith mass asymmetry in adult flatfishes. J. Fish Biol. 2008, 72, 2579–2594. [Google Scholar] [CrossRef] [Green Version]
- Petursdottir, G.; Begg, G.A.; Marteinsdottir, G. Discrimination between Icelandic cod (Gadus morhua L.) populations from adjacent spawning areas based on otolith growth and shape. Fish. Res. 2006, 80, 182–189. [Google Scholar] [CrossRef]
- Duarte-Neto, P.; Lessa, R.; Stosic, B.; Morize, E. The use of sagittal otoliths in discriminating stocks of common dolphinfish (Coryphaena hippurus) off northeastern brazil using multishape descriptors. ICES J. Mar. Sci. 2008, 65, 1144–1152. [Google Scholar] [CrossRef]
- Mahé, K.; Evano, H.; Mille, T.; Muths, D.; Bourjea, J. ; Otolith shape as a valuable tool to evaluate the stock structure of swordfish Xiphias gladius in the Indian Ocean. Afr. J. Mar. Sci. 2016, 38, 457–464. [Google Scholar] [CrossRef]
- Mille, T.; Mahé, K.; Villanueva, C.M.; De Pontual, H.; Ernande, B. Sagittal otolith morphogenesis asymmetry in marine fishes. J. Fish Biol. 2015, 87, 646–663. [Google Scholar] [CrossRef]
- Rebaya, M.; Ben Faleh, A.R.; Allaya, H.; Khedher, M.; Trojette, M.; Marsaoui, B.; Fatnassi, M.; Chalh, A.; Quignard, J.P.; Trabelsi, M. Otolith shape discrimination of Liza ramada (Actinopterygii: Mugiliformes: Mugilidae) from marine and estuarine populations in Tunisia. Acta Ichthyol. Piscat. 2017, 47, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Trojette, M.; Ben Faleh, A.; Fatnassi, M.; Marsaoui, B.; Mahouachi, N.H.; Chalh, A.; Quignard, J.-P.; Trabelsi, M. Stock discrimination of two insular populations of Diplodus annularis (Actinop-terygii: Perciformes: Sparidae) along the coast of Tunisia by analysis of otolith shape. Acta Ichthyol. Piscat. 2015, 45, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Ye, Z.; Li, Z.; Wan, R.; Ren, Y.; Dou, S. Population structure of Japanese Spanish mackerel Scomberomorus niphonius in the Bohai Sea, the Yellow Sea and the East China Sea: Evidence from random forests based on otolith features. Fish. Sci. 2016, 82, 251–256. [Google Scholar] [CrossRef]
- Hilbig, R.; Knie, M.; Shcherbakov, D.; Anken, R.H. Analysis of Behaviour and Habituation of Fish Exposed to Diminished Gravity in Correlation to Inner Ear Stone Formation—A Sounding Rocket Experiment (TEXUS 45). In Proceedings of the 20th ESA Symposium on European Rocket and Balloon Programmes and Related Research, Hyere, France, 22–26 May 2011. [Google Scholar]
- Anken, R.; Kappel, T.; Rahmann, H. Morphometry of fish inner ear otoliths after Development at 3g hypergravity. Acta Oto-Laryngol. 1998, 118, 534–539. [Google Scholar]
- Beier, M.; Anken, R.; Rahmann, H. Susceptibility to abnormal (kinetotic) swimming in fish correlates with inner ear carbonic anhydrase-reactivity. Neurosci. Lett. 2002, 335, 17–20. [Google Scholar] [CrossRef]
- Hilbig, R.; Anken, R.; Rahmann, H. On the origin of susceptibility to kinetotic swimming behaviour in fish: A parabolic aircraft flight study. J. Vestib. Res. 2003, 12, 185–189. [Google Scholar]
- Bao, B.; Ke, Z.; Xing, J.; Peatman, E.; Liu, Z.; Xie, C.; Xu, B.; Gai, J.; Gong, X.; Yang, G.; et al. Proliferating cells in suborbital tissue drive eye migration in flatfish. Dev. Biol. 2011, 351, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Sogard, S.M. Interpretation of otolith microstructure in juvenile winter flounder (Pseudopleuronectes americanus): Ontogenetic development, daily increment validation, and somatic growth relationships. Can. J. Fish. Aquat. Sci. 1991, 48, 1862–1871. [Google Scholar] [CrossRef]
- Helling, K.; Scherer, H.; Hausmann, S.; Clarke, A.H. Otolith mass asymmetries in the utricle and saccule of flatfish. J. Vestib. Res. 2005, 15, 59–64. [Google Scholar] [PubMed]
- Fischer, A.J.; Thompson, B.A. The age and growth of southern flounder, Paralichthys lethostigma, from Louisiana estuarine and offshorewaters. Bull. Mar. Sci. 2004, 75, 63–77. [Google Scholar]
- Sherratt, E.; Serb, J.M.; Adams, D.C. Rates of morphological evolution, asymmetry and morphological integration of shell shape in scallops. BMC Evol. Biol. 2017, 17, 248. [Google Scholar] [CrossRef] [Green Version]
- Stewart, T.; Albertson, R.C. Evolution of a unique predatory feeding apparatus: Functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids. BMC Biol. 2010, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Hata, H.; Yasugi, M.; Takeuchi, Y.; Takahashi, S.; Hori, M. Measuring and evaluating morphological asymmetry in fish: Distinct lateral dimorphism in the jaws of scale-eating cichlids. Ecol. Evol. 2013, 3, 4641–4647. [Google Scholar] [CrossRef]
- Leung, C.; Duclos, K.K.; Grünbaum, T.; Cloutier, R.; Angers, B. Asymmetry in dentition and shape of pharyngeal arches in the clonal fish Chrosomus eosneogaeus: Phenotypic plasticity and developmental instability. PLoS ONE 2017, 12, e0174235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, A.K.; Davis, E.M.; Kaplan, S.A.; Gross, J.B. Cranial asymmetry arises later in the life history of the blind Mexican cavefish, Astyanax Mex. PLoS ONE 2017, 12, e0177419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skliris, N. Past, present and future patterns of the thermohaline circulation and characteristic water masses of the Mediterranean sea. In The Mediterranean Sea—Its History and Present Challenges; Goffredo, S., Dubinky, Z.D., Eds.; Springer: New York, NY, USA, 2014; pp. 29–48. [Google Scholar]
- Ider, D.; Ramdane, Z.; Mahé, K.; Dufour, J.L.; Bacha, M.; Amara, R. Use of otolith shape analysis to discriminate stocks of Boops boops (L.) from the Algerian coast (southwestern part of the Mediterranean Sea). Afr. J. Mar. Sci. 2017, 39, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Archambault, B.; Rivot, E.; Savina, M.; Le Pape, O. Using a spatially structured life cycle model to assess the influence of multiple stressors on an exploited coastal-nursery-dependent population. Estuar. Coast. Shelf Sci. 2018, 201, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Randon, M.; Le Pape, O.; Ernande, B.; Mahé, K.; Volckaert, P.; Petit, E.J.; Lassalle, G.; LeBerre, T.; Réveillac, E. Complementarity and discriminatory power of genotype and otolith shape in describing the fine scale population structure of an exploited fish, the common sole of the Eastern English Channel. PLoS ONE 2020, 15, e0241429. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Geographical Area | Number | Total Length (cm) | Sampling Period | |||
|---|---|---|---|---|---|---|---|
| Mean±SE | Minimum | Maximum | |||||
| Boops boops | B1 | Tenerife Island | 67 | 19.00 ± 0.93 | 18 | 20 | 11/2016 |
| B2 | Gulf of Annaba | 40 | 19.12 ± 4.52 | 15 | 26 | 2/2013 to 12/2013 | |
| B3 | Gulf of Bejaia | 92 | 15.06 ± 1.70 | 11 | 19 | 1/2014 to 3/2014 | |
| B4 | Gulf of Oran | 47 | 15.50 ± 0.71 | 15 | 16 | 4/2015 | |
| B5 | Gulf of Tunis | 48 | 18.50 ± 1.02 | 17 | 21 | 7/2016 | |
| B6 | Corsica Island | 41 | 18.50 ± 1.91 | 16 | 20 | 6/2016 | |
| B7 | Gulf of Lion | 54 | 21.00 ± 4.69 | 15 | 25 | 6/2016 | |
| B8 | Ligurian Sea | 50 | 19.00 ± 1.74 | 17 | 23 | 6/2015 to 8/2015 | |
| B9 | Tyrrhenian Sea | 59 | 18.82 ± 2.07 | 16 | 21 | 6/2015 to 8/2015 | |
| B10 | Ionian Sea | 35 | 16.80 ± 2.86 | 15 | 23 | 9/2015 to 10/2015 | |
| B11 | Aegean Sea | 27 | 20.75 ± 0.71 | 17 | 22 | 9/2015 to 10/2015 | |
| Solea solea | S1 | Mont Saint Michel Bay | 476 | 29.04 ± 8.21 | 16 | 39 | 4/2017 to 5/2017 |
| S2 | Bay of Seine | 251 | 19.83 ± 7.75 | 15 | 27 | 4/2017 to 5/2017 | |
| S3 | Bay of Somme | 581 | 27.38 ± 4.60 | 24 | 34 | 4/2017 to 5/2017 | |
| S4 | French coast of the North Sea | 402 | 30.33 ± 2.50 | 26 | 34 | 4/2017 to 5/2017 | |
| S5 | Thames estuary | 297 | 28.71 ± 5.75 | 27 | 36 | 4/2017 to 5/2017 | |
| S6 | Central area of the eastern English Channel | 125 | 22.63 ± 7.04 | 18 | 26 | 4/2017 to 5/2017 | |
| S7 | English coast of the eastern English Channel | 299 | 29.04 ± 2.02 | 16 | 40 | 4/2017 to 5/2017 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahé, K.; MacKenzie, K.; Ider, D.; Massaro, A.; Hamed, O.; Jurado-Ruzafa, A.; Gonçalves, P.; Anastasopoulou, A.; Jadaud, A.; Mytilineou, C.; et al. Directional Bilateral Asymmetry in Fish Otolith: A Potential Tool to Evaluate Stock Boundaries? Symmetry 2021, 13, 987. https://doi.org/10.3390/sym13060987
Mahé K, MacKenzie K, Ider D, Massaro A, Hamed O, Jurado-Ruzafa A, Gonçalves P, Anastasopoulou A, Jadaud A, Mytilineou C, et al. Directional Bilateral Asymmetry in Fish Otolith: A Potential Tool to Evaluate Stock Boundaries? Symmetry. 2021; 13(6):987. https://doi.org/10.3390/sym13060987
Chicago/Turabian StyleMahé, Kélig, Kirsteen MacKenzie, Djamila Ider, Andrea Massaro, Oussama Hamed, Alba Jurado-Ruzafa, Patrícia Gonçalves, Aikaterini Anastasopoulou, Angelique Jadaud, Chryssi Mytilineou, and et al. 2021. "Directional Bilateral Asymmetry in Fish Otolith: A Potential Tool to Evaluate Stock Boundaries?" Symmetry 13, no. 6: 987. https://doi.org/10.3390/sym13060987
APA StyleMahé, K., MacKenzie, K., Ider, D., Massaro, A., Hamed, O., Jurado-Ruzafa, A., Gonçalves, P., Anastasopoulou, A., Jadaud, A., Mytilineou, C., Randon, M., Elleboode, R., Morell, A., Ramdane, Z., Smith, J., Bekaert, K., Amara, R., de Pontual, H., & Ernande, B. (2021). Directional Bilateral Asymmetry in Fish Otolith: A Potential Tool to Evaluate Stock Boundaries? Symmetry, 13(6), 987. https://doi.org/10.3390/sym13060987