
Enigma of Pyramidal Neurons: Chirality-Centric View on Biological Evolution. Congruence to Molecular, Cellular, Physiological, Cognitive, and Psychological Functions

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Justification of Subject-Matter
1. Introduction
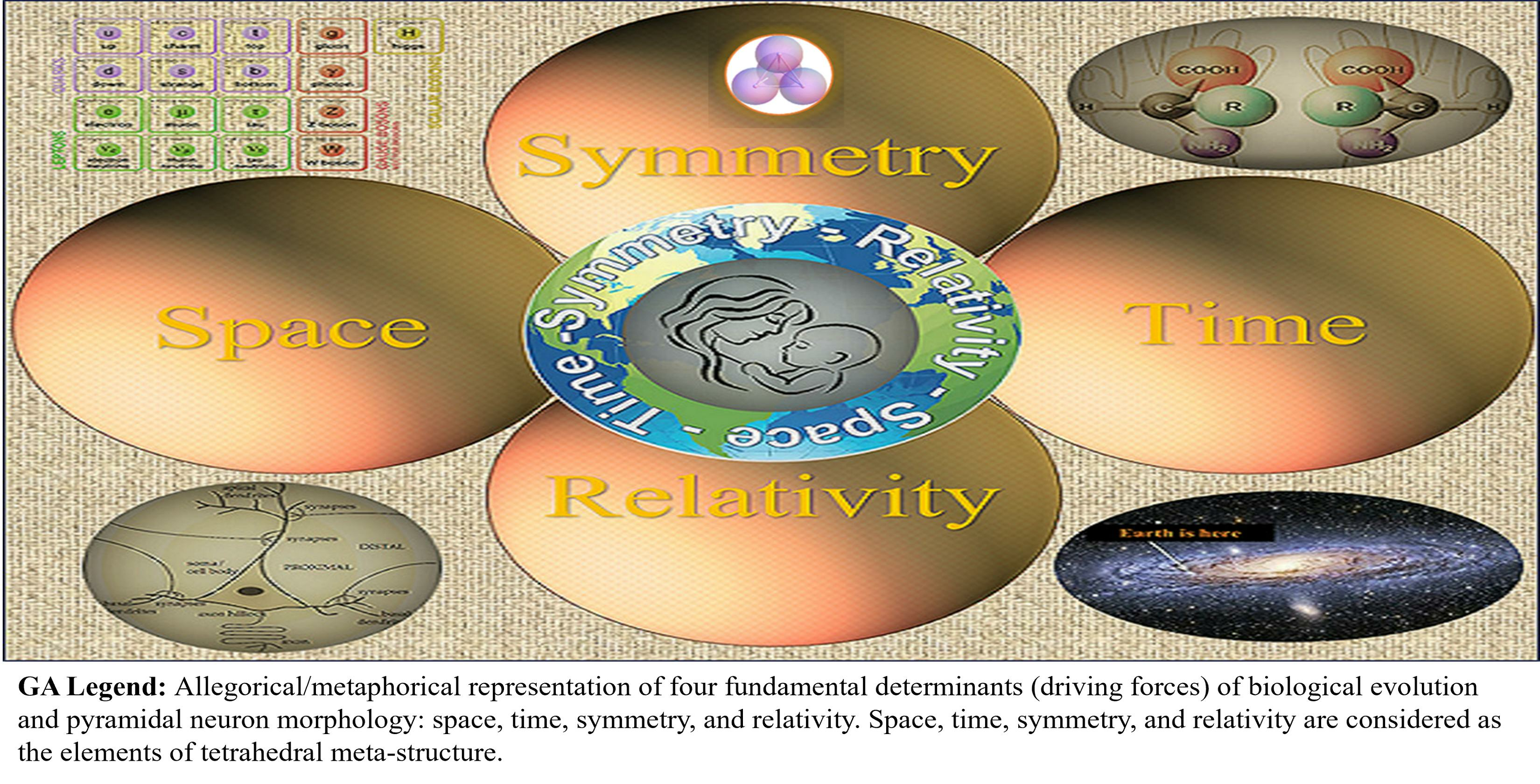
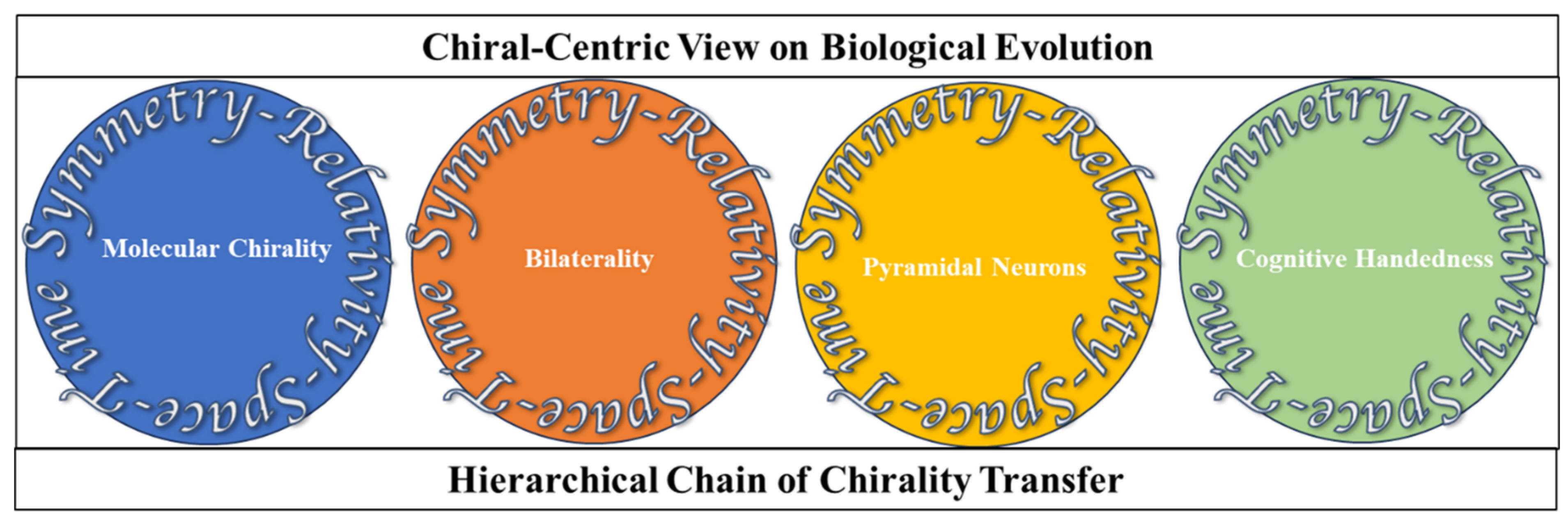
2. Hierarchical Chain of Chirality Transfer
Space, Time, Symmetry, and Relativity
3. Biological Evolution
3.1. General Trend
3.2. Molecular Chirality

3.2.1. Link of Physiological and Psychological Functions
3.2.2. Bilaterians: Symmetry–Function Interplay
3.3. Cell Chirality
3.3.1. Cell Evolution
3.3.2. Pyramidal Neurons
3.4. From Molecular to Cell Chirality
3.4.1. Self-Assembly of Biomolecules
3.4.2. Molecular–Cellular Co-Evolution
4. Conclusions
5. Closing Remark
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Michael, G.; John, S.; Witten, E. Superstring Theory. Vol. 2: Loop Amplitudes, Anomalies and Phenomenology; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Ma, Y.; Shi, L.; Yue, H.; Gao, G. Recognition at chiral interfaces: From molecules to cells. Colloids Surf. B Biointerfaces 2020, 195, 111268. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Bowerman, B. Symmetry Breaking in Biology. Cold Spring Harb. Perspect. Biol. 2010, 2, a003475. [Google Scholar] [CrossRef]
- Bormashenko, E. Fibonacci Sequences, Symmetry and Order in Biological Patterns, Their Sources, Information Origin and the Landauer Principle. Biophysica 2022, 2, 292–307. [Google Scholar] [CrossRef]
- Hollo, G. A new paradigm for animal symmetry. Interface Focus 2015, 5, 20150032. [Google Scholar] [CrossRef]
- Lee, C.; Weber, J.M.; Rodriguez, L.E.; Sheppard, R.Y.; Barge, L.M.; Berger, E.L.; Burton, A. Chirality in Organic and Mineral Systems: A Review of Reactivity and Alteration Processes Relevant to Prebiotic Chemistry and Life Detection Missions. Symmetry 2022, 14, 460. [Google Scholar] [CrossRef]
- Xin, Z.; Cai, Y.; Dang, L.T.; Burke, H.M.S.; Revote, J.; Charitakis, N.; Bienroth, D.; Nim, H.T.; Li, Y.-F.; Ramialison, M. MonaGO: A novel gene ontology enrichment analysis visualisation system. BMC Bioinform. 2022, 23, 69. [Google Scholar] [CrossRef] [PubMed]
- Dyakin, V.V. Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review. Symmetry 2023, 15, 79. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, M.; Kuang, X.; Li, Y.; Hu, S. A simplified morphological classification scheme for pyramidal cells in six layers of primary somatosensory cortex of juvenile rats. IBRO Rep. 2018, 5, 74–90. [Google Scholar] [CrossRef]
- Luine, V.; Frankfurt, M. Interactions between estradiol, BDNF and dendritic spines in promoting memory. Neuroscience 2013, 239, 34–45. [Google Scholar] [CrossRef]
- Vigouroux, R.J.; Duroure, K.; Vougny, J.; Albadri, S.; Kozulin, P.; Herrera, E.; Nguyen-Ba-Charvet, K.; Braasch, I.; Suárez, R.; Del Bene, F.; et al. Bilateral visual projections exist in non-teleost bony fish and predate the emergence of tetrapods. Science 2021, 372, 150–156. [Google Scholar] [CrossRef]
- Palladino, P. Stereochemistry and the Nature of Life: Mechanist, Vitalist, and Evolutionary Perspectives. JSTOR 1990, 81, 44–67. [Google Scholar] [CrossRef]
- Kushchayev, S.V.; Moskalenko, V.F.; Wiener, P.C.; Tsymbaliuk, V.I.; Cherkasov, V.G.; Dzyavulska, I.V.; Kovalchuk, O.I.; Sonntag, V.K.H.; Spetzler, R.F.; Preul, M.C. The discovery of the pyramidal neurons: Vladimir Betz and a new era of neuroscience. Brain J. Neurol. 2012, 135, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Radler, M.R.; Liu, X.; Peng, M.; Doyle, B.; Toyo-Oka, K.; Spiliotis, E.T. Pyramidal neuron morphogenesis requires a septin network that stabilizes filopodia and suppresses lamellipodia during neurite initiation. Curr. Biol. 2023, 33, 434–448.e8. [Google Scholar] [CrossRef]
- Sousa, A.M.M.; Meyer, K.A.; Santpere, G.; Gulden, F.O.; Sestan, N. Evolution of the Human Nervous System Function, Structure, and Development. Cell 2017, 170, 226–247. [Google Scholar] [CrossRef] [PubMed]
- Dahanayake, J.N.; Mitchell-Koch, K.R. Entropy connects water structure and dynamics in protein hydration layer. Phys. Chem. Chem. Phys. 2018, 20, 14765. [Google Scholar] [CrossRef] [PubMed]
- Marvan, T.; Polák, M.; Bachmann, T.; Phillips, W.A. Apical amplification—A cellular mechanism of conscious perception? Neurosci. Conscious. 2021, 2021, niab036. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Buldyrev, S.V.; Eugene, H. A tetrahedral entropy for water. Proc. Natl. Aacd. Sci. USA 2009, 106, 22130–22134. [Google Scholar] [CrossRef]
- Mason, S.F. The development of concepts of chiral discrimination. Chirality 1989, 1, 183–191. [Google Scholar] [CrossRef]
- Bastings, J.J.A.J.; van Eijk, H.M.; Damink, S.W.O.; Sander, S.; Rensen, S.S. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11, 2205. [Google Scholar] [CrossRef]
- Kondepudi, D. Chiral Asymmetry in Nature. In Chiral Analysis, 2nd ed.; Advances in Spectroscopy, Chromatography and Emerging Methods; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 1; pp. 3–28. [Google Scholar] [CrossRef]
- Cristadoro, G.; Degli Esposti, M.; Altmann, E.G. The common origin of symmetry and structure in genetic sequences. Sci. Rep. 2018, 8, 15817. [Google Scholar] [CrossRef]
- Book by Ibanez, L.E.; Uranga, A.M. String Theory and Particle Physics: An Introduction to String Phenomenology; Cambridge University Press: Cambridge, UK, 2012; 688p. [Google Scholar]
- Inaki, M.; Liu, J.; Matsuno, K. Cell chirality: Its origin and roles in left–right asymmetric development. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150403. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.M. Creating chirality. Nat. Chem. Biol. 2008, 4, 330. [Google Scholar] [CrossRef]
- Takaoka, K.; Yamamoto, M.; Hamada, H. Origin of body axes in the mouse embryo. Curr. Opin. Genet. Dev. 2007, 17, 44–350. [Google Scholar] [CrossRef] [PubMed]
- Dyakin, V.V.; Lucas, J.; Dyakina-Fagnano, N.V.; Posner, E.P.; Vadasz, C. The Chain of Chirality Transfer as Determinant of Brain Functional Laterality. Breaking the Chirality Silence: Search for New Generation of Biomarkers; Relevance to Neurodegenerative Diseases, Cognitive Psychology, and Nutrition Science. Neurol. Neurosci. Res. 2017, 1, 2. [Google Scholar] [CrossRef]
- Duan, P.; Cao, H.; Zhang, L.; Liu, M. Gelation induced supramolecular chirality: Chirality transfer, amplification and application. Soft Matter 2014, 10, 5428. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G. The evolutionary psychology of left and right: Costs and benefits of lateralization. Dev. Psychobiol. 2006, 48, 418–427. [Google Scholar] [CrossRef]
- Francks, C. Exploring human brain lateralization with molecular genetics and genomics. Ann. N. Y. Acad. Sci. 2015, 1359, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stacho, M.; Manahan-Vaughan, D. Mechanistic flexibility of the retrosplenial cortex enables its contribution to spatial cognition. Trends Neurosci. 2022, 45, P284–P296. [Google Scholar] [CrossRef]
- Lee, T.-W.; Dolan, R.J.; Critchley, H.D. Controlling Emotional Expression: Behavioral and Neural Correlates of Nonimitative Emotional Responses. Cereb. Cortex 2008, 18, 104–113. [Google Scholar] [CrossRef]
- Yuan, J.; Lu, X.; Zhang, S.; Zheng, F.; Deng, Q.; Han, L.; Lu, Q. Molecular Chirality and Morphological Structural Chirality of Exogenous Chirality-Induced Liquid Crystalline Block Copolymers. Macromolecules 2022, 55, 1566–1575. [Google Scholar] [CrossRef]
- Jung, C.G. Psychological Types (The Collected Works of C. G. Jung, Vol. 6) (Bollingen Series XX); Part of the Jung’s Collected Works (#6) Series and Dzieła (#2) Series; Princeton University Press: Princeton, NJ, USA, 1976. [Google Scholar]
- Assagioli, A. Dynamic Psychology and Psychosynthesis; Psychosynthesis Research Foundation, Inc.: New York, NY, USA, 1958. [Google Scholar]
- Myers, S. The five functions of psychological type. Anal. Psychol. 2016, 61, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Pillai, A.S.; Jirsa, V.K. Perspective. Symmetry Breaking in Space-Time Hierarchies Shapes Brain Dynamics and Behavior. Neuron 2017, 94, 1010–1026. [Google Scholar] [CrossRef] [PubMed]
- Jirsa, V.; Sheheitli, H. Entropy, free energy, symmetry and dynamics in the brain. J. Phys. Complex. 2022, 3, 015007. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Uversky, V.N. Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. Int. J. Mol. Sci. 2022, 23, 3687. [Google Scholar] [CrossRef] [PubMed]
- Iohnston, I.G.; Dingle, K.; Greenbury, S.F.; Camargo, C.Q.; Doye, J.P.K.; Ahnert, S.E.; Louis, A.A. Symmetry and simplicity spontaneously emerge from the algorithmic nature of evolution. Proc. Natl. Acad. Sci. USA 2022, 119, e2113883119. [Google Scholar] [CrossRef] [PubMed]
- Sharman, R.J.; Gheorghiu, E. Speed tuning properties of mirror symmetry detection mechanisms. Sci. Rep. 2019, 9, 3431. [Google Scholar] [CrossRef] [PubMed]
- Jammer, M. Concepts of Space: The history of Theories of Space in Physics, 3rd ed.; Dover: New York, NY, USA, 1993. [Google Scholar]
- Capecchi, D. Development of the Concept of Space up to Newton. Encyclopedia 2022, 2, 1528–1544. [Google Scholar] [CrossRef]
- Nederbr, H. Hierarchical Organization of Biological Systems and the Structure of Adaptation in Evolution and Tumorigenesis. J. Theor. Biol. 1997, 184, 149–156. [Google Scholar] [CrossRef]
- Eronen, M.I. Levels of Organization in Biology; Stanford Encyclopedia of Philosophy: Stanford, CA, USA, 2023; Available online: https://plato.stanford.edu/entries/levels-org-biology/ (accessed on 22 January 2023).
- Blackmond, D.G. The Origin of Biological Homochirality. Cold Spring Harb. Perspect. Biol. 2010, 2, a002147. [Google Scholar] [CrossRef]
- Ocklenburg, M.; Ocklenburg, S.; Mundorf, A. Symmetry and asymmetry in biological structures. Proc. Natl. Acad. Sci. USA 2022, 119, e2204881119. [Google Scholar] [CrossRef]
- Tasson, J.D. What Do We Know About Lorentz Invariance? Rep. Prog. Phys. 2014, 77, 062901. [Google Scholar] [CrossRef] [PubMed]
- Comte, C. Symmetry, relativity and quantum mechanics. Nuov Cim. B 1996, 111, 937–956. [Google Scholar] [CrossRef]
- Field, J.N. Space-Time Exchange Invariance: Special Relativity as a Symmetry Principle. Am. J. Phys. 2001, 69, 569. [Google Scholar] [CrossRef]
- Ajaltouni, Z.J. Symmetry and relativity: From classical mechanics to modern particle physics. Nat. Sci. 2014, 6, 43343. [Google Scholar] [CrossRef]
- Auffray, C.; Nottale, L. Scale relativity theory and integrative systems biology: 1: Founding principles and scale laws. Progress Biophys. Mol. Biol. 2008, 97, 79–114. [Google Scholar] [CrossRef]
- Noble, R.; Tasaki, K.; Noble, P.J.; Noble, D. Biological Relativity Requires Circular Causality but Not Symmetry of Causation: So, Where, What and When Are the Boundaries? Front. Physiol. Sec. Integr. Physiol. 2019, 10, 827. [Google Scholar] [CrossRef] [PubMed]
- de Kovel, C.G.F.; Lisgo, S.N.; Fisher, S.E.; Francks, C. Subtle left-right asymmetry of gene expression profiles in embryonic and foetal human brains. Sci. Rep. 2018, 8, 12606. [Google Scholar] [CrossRef]
- Bennett, M.S. Five Breakthroughs: A First Approximation of Brain Evolution from Early Bilaterians to Humans. Front. Neuroanat. 2021, 15, 693346. [Google Scholar] [CrossRef]
- Corballis, M.C. Bilaterally Symmetrical: To Be or Not to Be? Symmetry 2020, 12, 326. [Google Scholar] [CrossRef]
- Muskhelishvili, G.; Travers, A. Integration of syntactic and semantic properties of the DNA code reveals chromosomes as thermodynamic machines converting energy into information. Cell. Mol. Life Sci. 2013, 70, 4555–4567. [Google Scholar] [CrossRef]
- Chatterjee, S.; Yadav, S. The Coevolution of Biomolecules and Prebiotic Information Systems in the Origin of Life: A Visualization Model for Assembling the First Gene. Life 2022, 12, 834. [Google Scholar] [CrossRef] [PubMed]
- Devínsky, F. Chirality and the Origin of Life. Symmetry 2021, 13, 2277. [Google Scholar] [CrossRef]
- Piñeros, W.D.; Tlusty, T. Spontaneous chiral symmetry breaking in a random driven chemical system. Nat. Commun. 2022, 13, 2244. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.J.L.M.; Rowan, A.E.; Nolte, R.J.M.; Sommerdijk, N.A.J.M. Chiral Architectures from Macromolecular Building Blocks. Chem. Rev. 2001, 101, 4039–4070. [Google Scholar] [CrossRef] [PubMed]
- Todoroff, N.; Kunze, J.; Schreuder, H.; Hessler, G.; Baringhaus, K.-H.; Schneider, G. Fractal Dimensions of Macromolecular Structures. Mol. Inform. 2014, 33, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Panagiotou, E. The protein folding rate and the geometry and topology of the native state. Sci. Rep. 2022, 12, 6384. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Mandal, S.; Danielsen, M.B.; Brown, A.; Hu, C.; Christensen, N.J.; Kulakova, A.V.; Song, S.; Brown, T.; Jensen, K.J.; et al. Chirality transmission in macromolecular domains. Nat. Common. 2022, 13, 76. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Choi, H.; Shahzad, Z.M.; Ki, H.; Lee, J.; Chae, H.; Kim, Y.H. Supramolecular assembly of protein building blocks: From folding to function. Nano Converg. 2022, 9, 4. [Google Scholar] [CrossRef]
- MacKenzie, L.E.; Stachelek, P. The twists and turns of chiral chemistry. Nat. Chem. 2021, 13, 521–522. [Google Scholar] [CrossRef]
- Dhanavade, M.J.; Sonawane, K.D. Amyloid beta peptide-degrading microbial enzymes and its implication in drug design. 3 Biotech 2020, 10, 247. [Google Scholar] [CrossRef]
- Reetz, M.T.; Garcia-Borràs, M. The Unexplored Importance of Fleeting Chiral Intermediates in Enzyme-Catalyzed Reactions. J. Am. Chem. Soc. 2021, 143, 14939–14950. [Google Scholar] [CrossRef] [PubMed]
- Dyakin, V.V.; Dyakina-Fagnano, N.V.; Mcintire, L.B.; Uversky, V.N. Fundamental Clock of Biological Aging: Convergence of Molecular, Neurodegenerative, Cognitive and Psychiatric Pathways: Non-Equilibrium Thermodynamics Meet Psychology. Int. J. Mol. Sci. 2022, 23, 285. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Wisniewski, T.M.; Lajtha, A. Chiral Interface of Amyloid Beta (Aβ): Relevance to Protein Aging, Aggregation and Neurodegeneration. Symmetry 2020, 12, 585. [Google Scholar] [CrossRef] [PubMed]
- Stansley, B.L.; Yamamoto, B.K. L-dopa-induced dopamine synthesis and oxidative stress in serotonergic cells. Neuropharmacology 2013, 67, 243–251. [Google Scholar] [CrossRef] [PubMed]
- van Hooff, J.J.E. Towards unraveling the origins of eukaryotic nuclear genome organization. Trends Cell Biol. 2023, 33, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Caforio, A.; Driessen, A.J.M. Archaeal phospholipids: Structural properties and biosynthesis. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 1325–1339. [Google Scholar] [CrossRef] [PubMed]
- Fujii, N.; Takata, T.; Fujii, N.; Aki, K.; Sakaue, H. D-Amino Acid Residues in Proteins Related to Aging and Age-Related Diseases and a New Analysis of the Isomers in Proteins. In D-Amino Acid; Yoshimura, T., Nishikawa, T., Homma, H., Eds.; D-Amino Acids Springer: Tokyo, Japan, 2016; pp. 241–245. [Google Scholar] [CrossRef]
- Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Omar Cauli, Academic Editor and Soraya L. Valles, Academic Editor D-Amino Acids as a Biomarker in Schizophrenia. Diseases 2022, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Endicott, M.; Jones, M.; Hull, J. Amino acid metabolism as a therapeutic target in cancer: A review. Amino Acids. 2021, 53, 1169–1179. [Google Scholar] [CrossRef]
- Murtas, G.; Pollegioni, L. D-Amino Acids and Cancer: Friends or Foes? Int. J. Mol. Sci. 2023, 24, 3274. [Google Scholar] [CrossRef]
- Wolosker, H.; Balu, D.T. D-Serine as the gatekeeper of NMDA receptor activity: Implications for the pharmacologic management of anxiety disorders. Transl. Psychiatry 2020, 10, 184. [Google Scholar] [CrossRef]
- Li, Y.; Han, H.; Yin, J.; Li, T.; Yina, Y. Role of D-aspartate on biosynthesis, racemization, and potential functions: A mini-review. Anim. Nutr. 2018, 4, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, Y.; Katane, M.; Miyamoto, T.; Sekine, M.; Sakai-Kato, K.; Homma, H. D-Serine and D-Alanine Regulate Adaptive Foraging Behavior in Caenorhabditis elegans via the NMDA Receptor. J. Neurosci. 2020, 40, 7531–7544. [Google Scholar] [CrossRef] [PubMed]
- Kera, Y.; Aoyama, H.; Matsumura, H.; Hisae Nagasaki, H.; Yamada, R. Presence of free D-glutamate and D-aspartate in rat tissues. Biochim. Biophys. Acta (BBA) 1995, 1243, 282–286. [Google Scholar] [CrossRef]
- Mangas, A.; Coveñas, R.; Bodet, D.; Geffard, M.; Aguilar, L.A.; Yajeya, J. Immunocytochemical visualization of d-glutamate in the rat brain. Neuroscience 2007, 144, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Yang, H.T.; Lane, H.Y. d-glutamate, D-serine, and d-alanine differ in their roles in cognitive decline in patients with Alzheimer’s disease or mild cognitive impairment. Pharmacol. Biochem. Behav. 2019, 185, 172760. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Hou, C.; Bai, Y.; Wang, R.; Liu, J. Protein Assembly: Versatile Approaches to Construct Highly Ordered Nanostructures. Chem. Rev. (Am. Chem. Soc.) 2016, 116, 13571–13632. [Google Scholar] [CrossRef] [PubMed]
- Riccio, A.; Vitagliano, L.; di Prisco, G.; Zagari, A.; Mazzarella, L. The crystal structure of a tetrameric hemoglobin in a partial hemichrome state. Proc. Natl. Acad. Sci. USA 2002, 99, 9801–9806. [Google Scholar] [CrossRef]
- Ha, C.-E.; Bhagavan, N.V. Hemoglobin Chapter in Essentials of Medical Biochemistry, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2015; ISBN 9780124166974. [Google Scholar]
- Goldstein, E.B. Crosstalk between psychophysics and physiology in the study of perception. In Blackwell Handbook of Perception; Goldstein, E.B., Ed.; Blackwell Publishing: Oxford, UK, 2001; pp. 1–23. [Google Scholar]
- Xu, X.; Hanganu-Opatz, I.L.; Bieler, M. Cross-Talk of Low-Level Sensory and High-Level Cognitive Processing: Development, Mechanisms, and Relevance for Cross-Modal Abilities of the Brain. Front. Neurorobot. 2020, 14, 7. [Google Scholar] [CrossRef]
- Logothetis, N.K.; Pauls, J. Psychophysical and Physiological Evidence for Viewer-centered Object Representations in the Primate. Cereb. Cortex 1995, 3, 270–288. [Google Scholar] [CrossRef]
- Mascalzoni, E.; Osorio, D.; Regolinm, L.; Vallortigara, G. Symmetry perception by poultry chicks and its implications for three-dimensional object recognition. Proc. Biol. Sci. 2012, 279, 841–846. [Google Scholar] [CrossRef]
- Pizlo, Z.; de Barros, J.A. The Concept of Symmetry and the Theory of Perception. Front. Comput. Neurosci. 2021, 15, 681162. [Google Scholar] [CrossRef] [PubMed]
- Fabien, C. Design of a Bio-Inspired 3D Orientation Coordinate System and Application in Robotised Tele-Sonography. In Robot Arms; Goto, S., Ed.; 5 Princes Gate Court; IntecOpen: London, UK, 2011. [Google Scholar] [CrossRef]
- Craighero, L. The Role of the Sensorimotor System in Cognitive Functions. Brain Sci. 2022, 12, 604. [Google Scholar] [CrossRef] [PubMed]
- Paillard, J. Cognitive versus sensorimotor encoding of spatial information. In Cognitive Processes and Spatial Orientation in Animal and Man, Vol. II, Neurophysiology and Developmental Aspects; Ellen, P., Thinus-Blanc, C., Eds.; Martinus Nijhoff: Dordrecht, The Netherlands, 1987; Volume 37, ISBN 9-02473-448-7. [Google Scholar] [CrossRef]
- Fernandez-Baizan, C.; Nuñez, P.; Arias, J.L.; Mendez, M. Egocentric and allocentric spatial memory in typically developed children: Is spatial memory associated with visuospatial skills, behavior, and cortisol? Brain Behav. 2020, 10, e01532. [Google Scholar] [CrossRef] [PubMed]
- Nakade, Y.; Iwata, Y.; Furuichi, K.; Mita, M.; Hamase, K.; Konno, R.; Miyake, T.; Sakai, N.; Kitajima, S.; Toyama, T.; et al. Gut microbiota-derived D-serine protects against acute kidney injury. J. Clin. Investig. Insight 2018, 3, e97957. [Google Scholar] [CrossRef] [PubMed]
- Freud, S. An Outline of Psychoanalysis, 45th ed.; Vintage: London, UK, 1940; Volume 23. [Google Scholar]
- Wada, K.; Yamamoto, M.; Nakashima, K. Psychological function in aging. Nihon Rinsho 2013, 71, 1713–1719. [Google Scholar]
- Bottaccioli, A.G.; Bologna, M.; Bottaccioli, F. Psychic Life-Biological Molecule Bidirectional Relationship: Pathways, Mechanisms, and Consequences for Medical and Psychological Sciences—A Narrative Review. Int. J. Mol. Sci. 2022, 23, 3932. [Google Scholar] [CrossRef] [PubMed]
- Corballis, M.C.; Beale, I.L. Bilateral symmetry and behavior. Psychol. Rev. 1970, 77, 451–464. [Google Scholar] [CrossRef]
- Savin, D.N.; Tseng, S.-C.; Morton, S.M. Bilateral Adaptation During Locomotion Following a Unilaterally Applied Resistance to Swing in Nondisabled Adult. JPN J. Neirophysiol. 2010, 104I, 3600–3611. [Google Scholar] [CrossRef]
- Holló, G.; Novák, M. The manoeuvrability hypothesis to explain the maintenance of bilateral symmetry in animal evolution. Biol. Direct. 2012, 7, 22. [Google Scholar] [CrossRef]
- Alqadah, A.; Hsieh, Y.-W.; Morrissey, Z.D.; Chuang, C.-F. Asymmetric development of the nervous system. Dev. Dyn. 2018, 247, 124–137. [Google Scholar] [CrossRef]
- Heger, P.; Zheng, W.; Rottmann, A.; Panfilio, K.A.; Wiehe, T. The genetic factors of bilaterian evolution. eLife 2020, 9, e45530. [Google Scholar] [CrossRef] [PubMed]
- Tamada, A.; Igarashi, M. Revealing chiral cell motility by 3D Riesz transform-differential interference contrast microscopy and computational kinematic analysis. Nat. Commun. 2017, 8, 2194. [Google Scholar] [CrossRef] [PubMed]
- Delvenne, J.-F.; Castronovo, J.; Demeyere, N.; Umphreys, G.W. Bilateral Field Advantage in Visual Enumeration. PLoS ONE 2011, 6, e17743. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Toichi, M. The Role of Alternating Bilateral Stimulation in Establishing Positive Cognition in EMDR. Therapy: A Multi-Channel Near-Infrared Spectroscopy Study. PLoS ONE 2016, 11, e0162735. [Google Scholar] [CrossRef] [PubMed]
- Boukezzi, S.; Silva, C.; Nazarian, B.; Rousseau, P.-F.; Guedj, E.; Valenzuela-Moguillansky, C. Bilateral Alternating Auditory Stimulations Facilitate Fear Extinction and Retrieval. Front. Psychol. Sec. Psychol. Clin. Set. 2017, 8, 990. [Google Scholar] [CrossRef] [PubMed]
- Kasprian, G.; Langs, G.; Brugger, P.C.; Bittner, M.; Weber, M.; Arantes, M.; Prayer, D. The prenatal origin of hemispheric asymmetry: An in-utero neuroimaging study. Cereb. Cortex 2011, 21, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Mitter, C.; Prayer, D.; Brugger, P.C.; Weber, M.; Kaspria, G. In Vivo Tractography of Fetal Association Fibers. PLoS ONE 2015, 10, e011953. [Google Scholar] [CrossRef]
- Nakamura, M.; Hashimoto, T. Mechanistic Insights into Plant Chiral Growth. Symmetry 2020, 12, 2056. [Google Scholar] [CrossRef]
- Zagórska-Marek, B.; Sokołowska, K.; Turzańska, M. Chiral events in developing gametophores of Physcomitrella patens and other moss species are driven by an unknown, universal direction-sensing mechanism. Am. J. Bot. 2018, 105, 1986–1994. [Google Scholar] [CrossRef]
- Than, K. Symmetry in Nature: Fundamental Fact or Human Bias. Live Science. 2005. Available online: https://www.livescience.com/4002-symmetry-nature-fundamental-fact-human-bias.html (accessed on 22 January 2023).
- Gunturkun, O.; Ocklenburg, S. Ontogenesis of Lateralization. Neuron 2017, 94, 249–263. [Google Scholar] [CrossRef]
- Mehler, M.F. Epigenetic Principles and Mechanisms Underlying Nervous System Functions in Health and Disease. Prog. Neurobiol. 2008, 86, 305–341. [Google Scholar] [CrossRef] [PubMed]
- Zion, E.; Chen, X. Breaking Symmetry: The Asymmetries in Epigenetic Inheritance. Biochem 2021, 43, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Q.; Chin, A.S.; Worley, K.E.; Ray, P. Cell chirality: Emergence of asymmetry from cell culture. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150413. [Google Scholar] [CrossRef]
- Sun, N.; Dou, X.; Tang, Z.; Zhang, D.; Ni, N.; Wang, J.; Gao, H.; Ju, Y.; Dai, X.; Zhao, C.; et al. Bio-inspired chiral self-assemblies promoted neuronal differentiation of retinal progenitor cells through activation of metabolic pathway. Bioact. Mater. 2021, 6, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Madar, A.D.; Sheffield, M.E.J. Distinct place cell dynamics in CA1 and CA3 encode experience in new environments. Nat. Commun. 2021, 12, 2977. [Google Scholar] [CrossRef]
- Graham, H.K.; Duan, X. Molecular mechanisms regulating synaptic specificity and retinal circuit formation. Wiley Interdiscip. Rev. Dev. Biol. 2021, 10, e379. [Google Scholar] [CrossRef]
- Striedter, G.F. Special Issue: Cortical Evolution. Evolution of the hippocampus in reptiles and birds. JCN (Res. Syst. Neurosci.) 2016, 524, 496–517. [Google Scholar] [CrossRef]
- Matho, K.S.; Huilgol, D.; Galbavy, W.; He, M.; Kim, G.; An, X.; Lu, J.; Wu, P.; Di Bella, D.J.; Shetty, A.S.; et al. Genetic dissection of the glutamatergic neuron system in cerebral cortex. Nature 2021, 598, 182–187. [Google Scholar] [CrossRef]
- Zhang, H.; Wan, L.Q. Cell Chirality as A Novel Measure for Cytotoxicity. Adv. Biol. 2022, 6, e2101088. [Google Scholar] [CrossRef]
- Bekkers, J.M. Pyramidal neurons. Curr. Biol. Curr. Biol. 2011, 21, R975. [Google Scholar] [CrossRef]
- Banovac, I.; Sedmak, D.; Džaja, D.; Jalšovec, D.; Milošević, N.J.; Rašin, M.R.; Petanjek, Z. Somato-dendritic morphology and axon origin site specify von Economo neurons as a subclass of modified pyramidal neurons in the human anterior cingulate cortex. J. Anat. 2019, 235, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Johns, P. Neurons and glial cells. In Clinical Neuroscience; Elsevier: St George’s University of London, London, UK, 2014; Chapter 5; pp. 61–69. [Google Scholar] [CrossRef]
- Barger, N.; Sheley, M.F.; Schumann, C.M. Stereological study of pyramidal neurons in the human superior temporal gyrus from childhood to adulthood. J. Comp. Neurol. 2015, 523, 1054–1072. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.; Schijven, D.; Carrion-Castillo, A.; Joliot, M.; Mazoyer, B.; Fisher, S.E.; Crivello, F.; Francks, C. The genetic architecture of structural left-right asymmetry of the human brain. Nat. Hum. Behav. 2021, 5, 1226–1239. [Google Scholar] [CrossRef] [PubMed]
- Ottersen, O.P.; Zhang, N.; Walberg, F. Metabolic compartmentation of glutamate and glutamine: Morphological evidence obtained by quantitative immunocytochemistry in rat cerebellum. Neuroscience 1992, 46, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Sanghai, N.; Tranmer, G.K. Biochemical and Molecular Pathways in Neurodegenerative Diseases: An Integrated View. Cell 2023, 12, 2318. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.D.; Enerbäck, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.; Li, L.; Moss, J.; Petrelli, F.; Cassé, F.; Gebara, E.; Lopatar, J.; Pfrieger, F.W.; Bezzi, P.; Bischofberger, J.; et al. Synaptic Integration of Adult-Born Hippocampal Neurons Is Locally Controlled by Astrocytes. Neuron 2015, 88, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Krupic, J.; Bauza, M.; Burton, S.; Barry, C.; O’Keefe, J. Grid cell symmetry is shaped by environmental geometry. Nature 2015, 518, 232–235. [Google Scholar] [CrossRef]
- Oh, M.M.; Simkin, D.; Disterhoft, J.F. Intrinsic Hippocampal Excitability Changes of Opposite Signs and Different Origins in CA1 and CA3 Pyramidal Neurons Underlie Aging-Related Cognitive Deficits. Front. Syst. Neurosci. 2016, 10, 52. [Google Scholar] [CrossRef]
- Constant, M.; Mellet, E. The Impact of Handedness, Sex, and Cognitive Abilities on Left–Right Discrimination: A Behavioral Study. Front. Psychol. 2018, 9, 405. [Google Scholar] [CrossRef]
- Merrick, C.M.; Dixon, T.C.; Breska, A.; Lin, J.; Chang, E.F.; King-Stephens, D.; Laxer, K.D.; Weber, P.B.; Carmena, J.; Knight, R.T.; et al. Left hemisphere dominance for bilateral kinematic encoding in the human brain. eLife 2022, 11, e69977. [Google Scholar] [CrossRef]
- Jiang, S.; Guan, Y.; Chen, S.; Jia, X.; Ni, H.; Zhang, Y.; Han, Y.; Peng, X.; Zhou, C.; Li, A.; et al. Anatomically revealed morphological patterns of pyramidal neurons in layer 5 of the motor cortex. Sci. Rep. 2020, 10, 7916. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, R.; Shinohara, Y.; Kato, Y.; Sugiyama, H.; Shigemoto, R.; Ito, I. Asymmetrical allocation of NMDA receptor ε2 subunits in hippocampal sircuitry. Science 2003, 300, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Hafting, T.; Fyhn, M.; Molden, S.; Moser, M.-B.; Moser, E.I. Microstructure of a spatial map in the entorhinal cortex. Nature 2005, 436, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Mauro, M.; Monsalve-Mercado, C.L. The effect of boundaries on grid cell patterns. Phys. Rev. Res. 2020, 2, 043137. [Google Scholar] [CrossRef]
- Galakhova, A.A.; Hunt, A.S.; Wilbers, R.; de Kock, C.P.J.; Mansvelder, H.D.; Goriounova, N.A.; Goriounova, N.A. Evolution of cortical neurons supporting human cognition. Trends Cognit. Sci. 2022, 26, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, Y.; Hirase, H.; Watanabe, M.; Itakura, M.; Takahashi, M.; Shigemoto, R. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. Proc. Natl. Aacd. Sci. USA 2008, 105, 19498–19503. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.F.; Mascagni, F.; McDonald, A.J. Pyramidal Cells of the Rat Basolateral Amygdala. Synaptology and Innervation by Parvalbumin-immunoreactive Interneurons. J. Comp. Neurol. 2006, 494, 635–650. [Google Scholar] [CrossRef]
- Lorente de Nó, R.L. Cerebral cortex: Architecture, intracortical connections and motor projections. In Physiology of the Nervous System, 3rd ed.; Fulton, J.F., Ed.; Oxford University Press: Oxford, UK, 1949; pp. 288–330. [Google Scholar]
- Chan, C.H.; Godinho, L.N.; Thomaidou, D.; Tan, S.S.; Gulisano, M.; Parnavelas, J.G. Emx1 is a Marker for Pyramidal Neurons of the Cerebral Cortex. Cereb. Cortex 2001, 11, 1191–1198. [Google Scholar] [CrossRef]
- Hawkins, J.; Ahmad, S. Hypothesis and theory article. Why Neurons Have Thousands of Synapses, a Theory of Sequence Memory in Neocortex. Front. Neural Circuits 2016, 10, 23. [Google Scholar] [CrossRef]
- Jia, W.; Hu, C.; Wang, Y.; Liu, Y.; Wang, L.; Zhang, S.; Zhu, Q.; Gu, Y.; Zhang, P.; Ma, J.; et al. Identification of Single-Molecule Catecholamine Enantiomers Using a Programmable Nanopore. ACS Nano 2022, 16, 6615–6624. [Google Scholar] [CrossRef] [PubMed]
- Parnavelas, J.G.; Dinopoulos, A.; Davies, S.W. The central visual pathways. In Handbook of Chemical Neuroanatomy, Vol. 7. Integrated Systems of the CNS, Part II; Björklund, A., Hökfelt, T., Swanson, L.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 1–164. [Google Scholar]
- Dallérac, G.; Li, X.; Lecouflet, P.; Morisot, N.; Sacchi, S.; Asselot, R.; Pham, T.H.; Potier, B.; Watson, D.J.G.; Schmidt, S.; et al. Dopaminergic neuromodulation of prefrontal cortex activity requires the NMDA receptor coagonist D-serine. Proc. Natl. Acad. Sci. USA 2021, 118, e2023750118. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Krupic, J. Do hippocampal pyramidal cells respond to nonspatial stimuli? Physiol. Rev. 2021, 101, 1427–1456. [Google Scholar] [CrossRef] [PubMed]
- Purpura, D.P.; Suzuki, K. Distortion of neuronal geometry and formation of aberrant synapses in neuronal storage disease. Brain Res. 1976, 116, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Adámek, P.; Langová, V.; Horáček, J. Early-stage visual perception impairment in schizophrenia, bottom-up and back again. Schizophrenia 2022, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Granato, A.; De Giorgio, A. Alterations of neocortical pyramidal neurons: Turning points in the genesis of mental retardation. Front. Pediatr. 2014, 2, 86. [Google Scholar] [CrossRef] [PubMed]
- Rennó-Costa, C.; Adriano Tort, A.B.L. Place and Grid Cells in a Loop: Implications for Memory Function and Spatial Coding. J. Neurosci. 2017, 37, 8062–8076. [Google Scholar] [CrossRef] [PubMed]
- Kitanishi, T.; Ito, H.T.; Hayashi, Y.; Shinohara, Y.; Mizuseki, K. Network mechanisms of hippocampal laterality, place coding, and goal-directed navigation. J. Physiol. Sci. 2017, 67, 247–258. [Google Scholar] [CrossRef]
- Zhong, S.; He, Y.; Shu, H.; Gaolang Gong, G. Developmental Changes in Topological Asymmetry Between Hemispheric Brain White Matter Networks from Adolescence to Young Adulthood. Cereb. Cortex 2017, 7, 2560–2570. [Google Scholar] [CrossRef]
- Goriounova, N.A.; Heyer, D.B.; Wilbers, R.; Verhoog, M.B.; Giugliano, M.; Christophe Verbist, C.; Obermayer, J.; Kerkhofs, A.; Smeding, H.; Verberne, M.; et al. Large and fast human pyramidal neurons associated with intelligence. Elife 2018, 18, e41714. [Google Scholar] [CrossRef]
- Shipton, O.A.; El-Gaby, M.; Apergis-Schoute, J.; Deisseroth, K.; Bannerman, D.M.; Paulsen, O.; Kohl, M.M. Left–right dissociation of hippocampal memory processes in mice. Proc. Natl. Aacd. Sci. USA 2014, 111, 15238–15243. [Google Scholar] [CrossRef] [PubMed]
- Sá, M.J.; Ruela, C.; Madeira, M.D. Dendritic right/left asymmetries in the neurons of the human hippocampal formation: A quantitative Golgi study. Arq. Neuropsiquiatr. 2007, 65, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Spruston, N. Pyramidal neurons: Dendritic structure and synaptic integration. Nat. Rev. Neurosci. 2008, 9, 206–221. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Kurashima, R.; Gokan, H.; Inoue, N.; Ito, I.; Watanabe, S. Left−Right Asymmetry Defect in the Hippocampal Circuitry Impairs Spatial Learning and Working Memory in iv Mice. PLoS ONE 2010, 5, e15468. [Google Scholar] [CrossRef] [PubMed]
- Ukai, H.; Kawahara, A.; Hirayama, K.; Isao, I. ItPirB regulates asymmetries in hippocampal circuitry. PLoS ONE 2017, 12, e179377. [Google Scholar] [CrossRef] [PubMed]
- Luengo-Sanchez, S.; Bielza, C.; Benavides-Piccione, R.; Fernaud-Espinosa, I.; DeFelipe, J.; Larrañaga, P. A univocal definition of the neuronal soma morphology using Gaussian mixture models. Front. Neuroanat. 2015, 9, 137. [Google Scholar] [CrossRef]
- Rockland, K.S. Pyramidal Neurons: Looking for the origins of axons. Evol. Biol. Neurosci. 2022, 11, e79839. [Google Scholar] [CrossRef]
- Abrous, D.N.; Koehl, M.; Lemoine, M.A. Baldwin interpretation of adult hippocampal neurogenesis: From functional relevance to physiopathology. Mol. Psychiatry 2022, 27, 383–402. [Google Scholar] [CrossRef]
- Leguey, I.; Benavides-Piccione, R.; Rojo, C.; Larrañaga, P.; Bielza, C.; DeFelipe, J. Patterns of Dendritic Basal Field Orientation of Pyramidal Neurons in the Rat Somatosensory Cortex. eNeuro 2018, 5, ENEURO.0142-18.2018. [Google Scholar] [CrossRef]
- Weiler, S.; Nilo, D.G.; Bonhoeffer, T.; Hübener, M.; Rose, T.; Scheuss, V. Orientation and direction tuning align with dendritic morphology and spatial connectivity in mouse visual cortex. Curr. Biol. 2020, 32, 1743–1753.e7. [Google Scholar] [CrossRef]
- Wahle, P.; Sobierajski, E.; Gasterstädt, I.; Lehmann, N.; Weber, S.; Lübke, H.R.; Engelhardt, M.; Distler, C.; Meyer, G. Neocortical pyramidal neurons with axons emerging from dendrites are frequent in non-primates, but rare in monkey and human. Elife 2022, 11, e76101. [Google Scholar] [CrossRef] [PubMed]
- Musall, S.; Sun, X.R.; Mohan, H.; An, X.; Gluf, S.; Li, S.-J.; Drewes, R.; Cravo, E.; Lenzi, I.; Yin, C.; et al. Pyramidal cell types drive functionally distinct cortical activity patterns during decision-making. Nat. Neurosci. 2023, 26, 495–505. [Google Scholar] [CrossRef]
- Park, J.; Papoutsi, A.; Ash, R.T.; Poirazi, P.; Smirnakis, S.M.; Ghani, N. Contribution of apical and basal dendrites to orientation encoding in mouse V1 L2/3 pyramidal neurons. Nat. Commun. 2019, 10, 5372. [Google Scholar] [CrossRef] [PubMed]
- Elaine, M.; Pinheiro, E.M.; Xie, Z.; Norovich, A.L.; Vidaki, M.; Tsai, L.-H.; Gertler, F.B. Lpd depletion reveals that SRF specifies radial versus tangential migration of pyramidal neurons. Nat. Cell Biol. 2011, 13, 989–995. [Google Scholar] [CrossRef]
- Hobert, O. Development of left/right asymmetry in the Caenorhabditis elegans nervous system: From zygote to postmitotic neuron. Genesis 2014, 52, 528–543. [Google Scholar] [CrossRef]
- Tee, Y.; Shemesh, T.; Thiagarajan, V.; Hariadi, R.F.; Anderson, K.L.; Page, C.; Volkmann, N.; Hanein, D.; Sivaramakrishnan, S.; Kozlov, M.M.; et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat. Cell Biol. 2015, 17, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Falk, J.; Boubakar, L.; Valérie Castellani, V. Septin functions during neuro-development, a yeast perspective. Curr. Opin. Neurobiol. 2019, 57, 102–109. [Google Scholar] [CrossRef]
- Inaki, M.; Sasamura, T.; Matsuno, K. Cell Chirality Drives Left-Right Asymmetric Morphogenesis. Front. Cell Dev. Biol. 2018, 6, 34. [Google Scholar] [CrossRef]
- Konietzny, A.; Bär, J.; Mikhaylova, M. Dendritic Actin Cytoskeleton: Structure, Functions, and Regulations. Front. Cell. Neurosci. Sec. Cell. Neurophysiol. 2017, 11, 147. [Google Scholar] [CrossRef]
- Machnicka, B.; Czogalla, A.; Hryniewicz-Jankowska, A.; Bogusławska, D.M.; Grochowalska, R.; Heger, E.; Sikorski, A.F. Spectrins: A structural platform for stabilization and activation of membrane channels, receptors and transporters. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 620–634. [Google Scholar] [CrossRef]
- Lorenzo, D.N.; Edwards, R.J.; Slavutsky, A.L. Spectrins: Molecular organizers and targets of neurological disorders. Nat. Rev. Neurosci. 2023, 24, 195–212. [Google Scholar] [CrossRef]
- Satir, P. Chirality of the cytoskeleton in the origins of cellular asymmetry. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150408. [Google Scholar] [CrossRef] [PubMed]
- Parato, J.; Bartolini, F. The microtubule cytoskeleton at the synapse. Neurosci. Lett. 2021, 753, 135850. [Google Scholar] [CrossRef]
- Lamprecht, R. The actin cytoskeleton in memory formation. Progress. Neurobiol. 2014, 117, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ponce, D.; Blázquez-Llorca, L.; De Felipe, J.; Garrido, J.J.; Muñoz, A. Colocalization of α-actinin and Synaptopodin in the Pyramidal Cell Axon Initial Segment. Cereb. Cortex 2012, 22, 1648–1661. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.T.; Gupton, S.L. Cytoskeleton: Septin wreaths regulate actin in neuritogenesis. Curr. Biol. 2023, 33, R98–R100. [Google Scholar] [CrossRef] [PubMed]
- Bocquet, A.; Berges, R.; Frank, R.; Robert, P.; Peterson, A.C.; Eyer, J. Neurofilaments Bind Tubulin and Modulate Its Polymerization. J. Neurosci. 2009, 29, 11043–11054. [Google Scholar] [CrossRef]
- Uylings, H.B.; Jacobsen, A.M.; Zilles, K.; Amunts, K. Left-right asymmetry in volume and number of neurons in adult Broca’s area. Cortex 2006, 42, 652–658. [Google Scholar] [CrossRef]
- El-Gaby, M.; Reeve, H.M.; Lopes-dos-Santos, V.; Campo-Urriza, N.; Perestenko, P.V.; Morley, A.; Strickland, L.A.M.; Lukács, I.P.; Paulsen, O.; Dupret, D. An Emergent Neural Coactivity Code for Dynamic Memory. Nat. Neurosci. 2021, 24, 694–704. [Google Scholar] [CrossRef]
- Cullen, T.J.; Walker, M.A.; Eastwood, S.L.; Esiri, M.M.; Harrison, P.J.; Crow, T.J. Anomalies of asymmetry of pyramidal cell density and structure in dorsolateral prefrontal cortex in schizophrenia. Br. J. Psychiatry 2006, 188, 26–31. [Google Scholar] [CrossRef]
- Souta, R. An Introductory Perspective on the Emerging Application of qEEG. In Introduction to Quantitative EEG and Neurofeedback. Advsanced Theory and Applications, 2nd ed.; Budzynski, T.H., Washington, P., Budzynski, H.K., Washington, P., Evans, J.R., Abarbanel, A., Eds.; Academic Press: New York, NY, USA, 2009. [Google Scholar]
- Beniaguev, D.; Segev, I.; London, M. Single cortical neurons as deep artificial neural networks. Neuron 2021, 109, 2727–2739.e3. [Google Scholar] [CrossRef] [PubMed]
- Toxvaerd, S. The emergence of the bilateral symmetry in animals: A review and a new hypothesis. Symmetry 2021, 13, 261. [Google Scholar] [CrossRef]
- Matsuo, K.; Tamura, R.; Hotta, K.; Okada, M.; Takeuchi, A.; Wu, Y.; Hashimoto, K.; Takano, H.; Momose, A.; Nishino, A. Bilaterally Asymmetric Helical Myofibrils in Ascidian Tadpole Larvae. Front. Cell Dev. Biol. 2021, 9, 800455. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.A.; Giomi, L. Theory of cellular homochirality and trait evolution in flocking systems. arXiv 2023, arXiv:2307.14360. [Google Scholar] [CrossRef]
- Zhu, D.Y.; Cao, T.T.; Fan, H.W.; Zhang, M.-Z.; Duan, H.-K.; Li, J.; Zhang, X.-J.; Li, Q.; Wang, P.; Chen, T. The increased in vivo firing of pyramidal cells but not interneurons in the anterior cingulate cortex after neuropathic pain. Mol. Brain. 2022, 15, 12. [Google Scholar] [CrossRef]
- Koga, K. Early evolution of membrane lipids: How did the lipid divide occur? J. Mol. Evol. 2011, 72, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.-X.; Jain, N.; Preuss, T.M.; Kaas, J.K. Inverted pyramidal neurons in chimpanzee sensorimotor cortex are revealed by immunostaining with monoclonal antibody SMI-32. Somatosens. Mot. Res. 1999, 16, 49–56. [Google Scholar] [CrossRef]
- Steger, R.; Ramos, R.L.; Cao, R.; Yang, Q.; Chen, C.-C.; Dominici, J.; Brumberg, J.C. Physiology and morphology of inverted pyramidal neurons in the rodent neocortex. Neuroscience 2013, 17, 165–179. [Google Scholar] [CrossRef]
- Martin, H.S.; Podolsky, K.A.; Devaraj, N.K. Probing the role of chirality in phospholipid membranes. ChemBioChem 2021, 22, 3148–3315. [Google Scholar] [CrossRef]
- Kong, X.-Z.; Postema, M.C.; Guadalupe, T.; de Kovel, C.; Boedhoe, P.C.W.; Hoogman, M.; Mathias, S.R.; van Rooij, D.; Schijven, D.; Glahn, D.C.; et al. Mapping brain asymmety in health and disease through the ENIGMA consortium. Hum. Brain Map. 2022, 43, 167–181. [Google Scholar] [CrossRef]
- Kay, J.W.; Phillips, W.A. Contextual Modulation in Mammalian Neocortex is Asymmetric. Symmetry 2020, 12, 815. [Google Scholar] [CrossRef]
- Tripathi, S.; Dey, A.; Shanmugam, M.; Narayanan, R.S.; Chandrsekhar, V. Cobalt (II) Complexes as Single-Ion Magnets. In Organometallic Magnets; Topics in Organometallic Chemistry Series; Springer: Cham, Switzerland, 2019; Volume 64. [Google Scholar] [CrossRef]
- Kagamiyama, H.; Hayashi, H. Crystallographic Structures. Branched-Chain Amino Acids, Part B. In Methods in Enzymoligy; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Housecroft, C.E.; Sharpe, A.G. Inorganic Chemistry, 2nd ed.; Prentice Hall: Hoboken, NJ, USA, 2004; ISBN 978-0-13-039913-7. [Google Scholar]
- Gradišar, H.; Božič, S.; Doles, T.; Vengust, D.; Hafner-Bratkovič, I.; Mertelj, A.; Webb, B.; Šali, A.; Klavžar, S.; Jerala, R. Design of a single-chain polypeptide tetrahedron assembled from coiled-coil segments. Nat. Chem. Biol. 2013, 9, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Le Bel, J.A. Sur les relations qui existent entre les formules atomiques des corps organiques et le pouvoir rotatoire de leurs dissolutions. Bull. Soc. Chim. Fr. 1874, 22, 337. [Google Scholar]
- Mun, J.; Kim, M.; Yang, Y.; Badloe, T.; Ni, J.; Chen, Y.; Qiu, C.; Rho, J. Electromagnetic chirality: From fundamentals to nontraditional chiroptical phenomena. Light. Sci. Appl. 2020, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R.J. Use of chiral cell shape to ensure highly directional swimming in trypanosomes. PLoS Comput. Biol. 2017, 13, e1005353. [Google Scholar] [CrossRef] [PubMed]
- Van‘t Hoff, J.H. A suggestion looking to the extension into space of the structural formulas at present used in chemistry and a note upon the relation between the optical activity and the chemical constitution of organic compounds. Arch. Neérl. Sci. Exactes. Nat. 1874, 9, 445–454. Available online: https://www.chemteam.info/Chem-History/Van’t-Hoff-1874.html (accessed on 12 July 2023).
- Xie, N.; Liu, S.; Yang, X.; He, X.; Huang, J.; Wang, K. DNA tetrahedron nanostructures for biological applications: Biosensors and drug delivery. Analyst 2017, 18, 3322–3332. [Google Scholar] [CrossRef]
- Mathur, D.; Rogers, K.E.; Díaz, S.A.; Muroski, M.E.; Klein, W.P.; Nag, O.K.; Lee, K.; Field, L.D.; Delehanty, J.B.; Medin, I.L. Determining the Cytosolic Stability of Small DNA Nanostructures in Cellula. Nano Lett. 2022, 22, 5037–5045. [Google Scholar] [CrossRef]
- Aumiller, W.A., Jr.; Uchida, M.; Douglas, T. Protein cage assembly across multiple length scales. Chem. Soc. Rev. 2018, 47, 3433–3469. [Google Scholar] [CrossRef]
- Tenchov, R.; Bird, R.; Curtze, A.E.; Zhou, Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef]
- Hong, S.; Jiang, W.; Ding, Q.; Lin, K.; Zhao, C.; Wang, X. The Current Progress of Tetrahedral DNA Nanostructure for Antibacterial Application and Bone Tissue Regeneration. Int. J. Nanomed. 2023, 18, 3761–3780. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Mao, K.; Chen, S.; Zhu, H. Chirality Effects in Peptide Assembly Structures. Front. Bioeng. Biothechnol. 2021, 9, 703004. [Google Scholar] [CrossRef] [PubMed]
- Hassabis, D.; Kumaran, D.; Summerfield, C.; Botvinick, M. Neuroscience-Inspired Artificial Intelligence. Neuron 2017, 95, P245–P258. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, M.; Zhou, L.; Wang, X.; Jia, J. The Application of Artificial Intelligence in Brain-Computer Interface and Neural System Rehabilitation. Front. Neurosci. 2022, 16, 1024316. [Google Scholar] [CrossRef] [PubMed]
- Surianarayanan, C.; Lawrence, J.J.; Chelliah, P.R.; Prakash, E.; Hewage, C. Convergence of Artificial Intelligence and Neuroscience towards the Diagnosis of Neurological Disorders—A Scoping Review. Sensors 2023, 23, 3062. [Google Scholar] [CrossRef] [PubMed]
- Strychalski, W. 3D Computational Modeling of Bleb Initiation Dynamics. Front. Phys. Sec. Biophys. 2021, 9, 775465. [Google Scholar] [CrossRef]
- Johnson, E.; Cascio, D.; Sawaya, M.R.; Gingery, M.; Schröder, I. Crystal Structures of a Tetrahedral Open Pore Ferritin from the Hyperthermophilic Archaeon Archaeoglobus fulgidus. Structure 2005, 13, 637–648. [Google Scholar] [CrossRef]
- Zhang, K.; Ezemaduka, A.; Wang, Z.; Hu, H.; Shi, X.; Liu, C.; Lu, X.; Fu, X.; Chang, Z.; Yin, C. A Novel Mechanism for Small Heat Shock Proteins to Function as Molecular Chaperones. Sci. Rep. 2015, 5, 8811. [Google Scholar] [CrossRef]
- Yoo, S.H.; Lee, H.-S. Foldectures: 3D Molecular Architectures from Self-Assembly of Peptide Foldamers. Acc. Chem. Res. 2017, 50, 832–841. [Google Scholar] [CrossRef]
- Sasaki, E.; Böhringer, D.; van de Waterbeemd, M.; Leibundgut, M.; Zschoche, R. Structure and assembly of scalable porous protein cages. Nat. Commun. 2017, 8, 14663. [Google Scholar] [CrossRef]
- Quaye, I.K. Extracellular hemoglobin: The case of a friend turned foe. Front. Physiol. 2015, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Eaton, A.W. Retrospective on statistical mechanical models for hemoglobin allostery editors-pick. J. Chem. Phys. 2022, 157, 184104. [Google Scholar] [CrossRef]
- Richter, F.; Meurers, B.H.; Zhu, C.; Medvedeva, V.P.; Chesselet, M.-F. Neurons Express Hemoglobin α- and β-Chains in Rat and Human Brains. J. Comp. Neurol. 2009, 15, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ronson, T.K.; Mosquera, J.; Martinez, A.; Guy, L.; Nitschke, J.R. Anion Binding in Water Drives Structural Adaptation in an Azaphosphatrane-Functionalized FeII4L4 Tetrahedron. J. Am. Chem. Soc. 2017, 139, 6574–6577. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Li, Q.; Mogilner, A.; Bershadsky, A.D.; Shivashankar, G.V. Mechanical stimulation induces formin-dependent assembly of a perinuclear actin rim. Proc. Natl. Acad. Sci. USA 2015, 112, E2595–E2601. [Google Scholar] [CrossRef] [PubMed]
- Kruppa, A.J.; Kishi-Itakura, C.; Masters, T.A.; Rorbach, J.E.; Grice, G.L.; Kendrick-Jones, J.; James, A.; Nathan, J.A.; Minczuk, M. Myosin VI-Dependent Actin Cages Encapsulate Parkin-Positive Damaged Mitochondria. Dev. Cell. 2018, 26, 484–499.e6. [Google Scholar] [CrossRef]
- Mostowy, S.; Bonazzi, M.; Hamon, M.A.; Tham, T.N.; Mallet, A.; Lelek, M.; Gouin, E.; Demangel, C.; Brosch, R.; Zimmer, C.; et al. Entrapment of intracytosolic bacteria by septin cage-like structures. Cell Host Microbe 2010, 8, 433–444. [Google Scholar] [CrossRef]
- Heid, H.; Rickelt, S.; Zimbelmann, R.; Winter, S.; Schumacher, H.; Dörflinger, Y.; Kuhn, C.; Franke, W.W. On the formation of lipid droplets in human adipocytes: The organization of the perilipin-vimentin cortex. PLoS ONE 2014, 9, e90386. [Google Scholar] [CrossRef]
- Tessarz, P.; Schwarz, M.; Mogk, A.; Bukau, B. The yeast AAA+ chaperone Hsp104 is part of a network that links the actin cytoskeleton with the inheritance of damaged proteins. Mol. Cell Biol. 2009, 29, 3738–3745. [Google Scholar] [CrossRef]
- Franke, W.W.; Hergt, M.; Grund, C. Rearrangement of the vimentin cytoskeleton during adipose conversion: Formation of an intermediate filament cage around lipid globules. Cell 1987, 49, 131–141. [Google Scholar] [CrossRef]
- Biliński, S.M.; Jaglarz, M.K. Organization and possible functions of microtubule cytoskeleton in hymenopteran nurse cells. Cell Motil. Cytoskeleton. 1999, 43, 213–220. [Google Scholar] [CrossRef]
- Xia, Z.; Wang, P.; Liu, X.; Liu, T.; Yan, Y.; Yan, J.; Zhong, J.; Sun, G.; He, D. Tumor-Penetrating Peptide-Modified DNA Tetrahedron for Targeting Drug Delivery. Biochemistry 2016, 55, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xu, T.; Yan, Y.; Zhou, Y.-R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Wu, S.; Chu, L.T.; Kwong, H.K.; Hartanto, H.; Huang, Y.; Lam, M.L.; Lam, R.H.W.; Chen, T.-H. Early Committed Clockwise Cell Chirality Upregulates Adipogenic Differentiation of Mesenchymal Stem Cells. Adv. Biosyst. 2020, 10, e2000161. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Hu, Y.; Cao, B.; Peng, R.; Ding, J. Effects of surface molecular chirality on adhesion and differentiation of stem cells. Biomaterials 2013, 34, 9001–9009. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yoshitomi, T.; Yoshimoto, K. Analysis of Chirality Effects on Stem Cell Fate Using Three-dimensional Fibrous Peptide Hydrogels. ACS Appl. Bio. Mater. 2018, 1, 538–543. [Google Scholar] [CrossRef]
- Walser, M.; Svensson, J.; Karlsson, M.R.; Åberg, M.; Kuhn, H.G.; Isgaard, J.; Åberg, N.D. Growth Hormone and Neuronal Hemoglobin in the Brain—Roles in Neuroprotection and Neurodegenerative Diseases. Front. Endocrinol. 2021, 11, 606089. [Google Scholar] [CrossRef]
- Zheng, R.; Yan, Y.; Pu, J.; Zhang, B. Physiological and Pathological Functions of Neuronal Hemoglobin: A Key Underappreciated Protein in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 9088. [Google Scholar] [CrossRef]
- Flynn, K.S. The cytoskeleton and neurite initiation. Bioarchitecture 2013, 3, 86–109. [Google Scholar] [CrossRef]
- Indelicato, G.; Wahome, N.; Müller, P.R.S.A.; Nieh, M.-P.; Burkhard, P.; Twarock, R. Principles Governing the Self-Assembly of Coiled-Coil Protein Nanoparticles. Biophys. J. 2016, 110, P646–P660. [Google Scholar] [CrossRef]
- Heberle, F.A.; Feigenson, G.W. Phase Separation in Lipid Membranes. Cold Spring Harb. Perspect. Biol. 2011, 3, a004630. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, Q.; Feng, Z.; Zhang, M. Liquid-Liquid Phase Separation in Neuronal Development and Synaptic Signaling. Dev. Cell 2020, 55, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Wonka, P.; Gervautz, M. Ray Tracing of Nonlinear Fractals. J. WSCG 1998, 6, 424–431. Available online: https://wscg.zcu.cz/wscg1998/wscg98.htm (accessed on 13 January 2022).
- Nieuwenhuys, R. The neocortex. Anat. Embryol. 1994, 190, 307–337. [Google Scholar] [CrossRef] [PubMed]
- Pabic, P.L.; Dranow, D.B.; Hoyle, D.J.; Schilling, T.F. Zebrafish endochondral growth zones as they relate to human bone size, shape and disease. Front. Endocrinol. 2022, 13, 1060187. [Google Scholar] [CrossRef] [PubMed]
- Hall, E. The amygdala of the cat: A Golgi study. Z. Zellforsch. Mikrosk. Anat. 1972, 134, 439–458. [Google Scholar] [CrossRef]
- DeMarco, E.; Tesmer, A.L.; Hech, B.; Kawakami, K.; Robles, E. Pyramidal Neurons of the Zebrafish Tectum Receive Highly Convergent Input From Torus Longitudinalis. Front. Neuroanat. 2021, 15, 636683. [Google Scholar] [CrossRef]
- Suryanarayana, S.M. On the Evolutionary Origin of the Vertebrate Cortex; Department of Neuroscience Karolinska Institutet: Srickholm, Sweden, 2019. [Google Scholar]
- Taylor, L.L.K.; Riddell, I.A.; Smulders, M.M.J. Self-Assembly of Functional Discrete Three-Dimensional Architectures in Water. Angew. Chem. 2019, 58, 1280–1307. [Google Scholar] [CrossRef]
- Chirikjian, G.S. Group theory and biomolecular conformation: I. Mathematical and computational models. J. Phys. Condens. Matter. 2010, 22, 323103. [Google Scholar] [CrossRef]
- Dyakin, V.V.; Posner, E.V. Molecular Chirality as an Integral Biomarker of Lactation, Nutrition and Cognitive Development Express-Review. Biomed. Sci. Res. 2020, 8, AJBSR.2020.08.001337. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Z.; Armstrong, D.W.; Wolosker, H.; Zheng, Y. Detection and analysis of chiral molecules as disease biomarkers. Nat. Rev. Chem. 2023, 7, 355–373. [Google Scholar] [CrossRef] [PubMed]
- Ariga, K.; Richards, G.J.; Ishihara, S.; Izawa, H.; Jonathan, P.; Hill, J.P. Intelligent Chiral Sensing Based on Supramolecular and Interfacial Concepts. Sensors 2010, 10, 6796–6820. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, J.; Liu, R.; Tu, Y.; Li, Y.; Cheng, J.; He, T.; Zhu, X. Autonomous discovery of optically active chiral inorganic perovskite nanocrystals through an intelligent cloud lab. Nat. Commun. 2020, 11, 2046. [Google Scholar] [CrossRef] [PubMed]
- Rainey, P.B. Major evolutionary transitions in individuality between humans and AI. Philos. Trans. R. Soc. B 2023, 378. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, N.; Amar, M.; Baux, G.; Fossier, P. Homeostatic control of the excitation–inhibition balance in cortical layer 5 pyramidal neurons. Eur. J. Neurosci. 2006, 24, 3507–3518. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.C.; Horwood, N.; Isaacs, S.L.; Mann, D.M.A. The effect of age and Alzheimer’s disease on pyramidal neuron density in the individual fields of the hippocampal formation. Acta Neuropathol. 1992, 83, 510–517. [Google Scholar] [CrossRef]
- El-Gaby, M.; Shipton, O.A.; Paulsen, O. Synaptic Plasticity and Memory: New Insights from Hippocampal Left–Right Asymmetries. Neuroscientist 2014, 21, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Shimbo, A.; Kosaki, Y.; Ito, I.; Watanabe, S. Mice lacking hippocampal left-right asymmetry show non-spatial learning deficits. Behav. Brain Res. 2018, 336, 156–165. [Google Scholar] [CrossRef]
- Bussière, T.; Giannakopoulos, P.; Bouras, C.; Perl, D.P.; Morrison, J.H.; Patrick, R.; Hof, P.R. Progressive degeneration of nonphosphorylated neurofilament protein-enriched pyramidal neurons predicts cognitive impairment in Alzheimer’s disease: Stereologic analysis of prefrontal cortex area 9. J. Compap. Neurol. 2003, 463, 281–302. [Google Scholar] [CrossRef]
- Cholvin, T.; Bartos, M. Hemisphere-specific spatial representation by hippocampal granule cells. Nat. Commun. 2022, 13, 6227. [Google Scholar] [CrossRef]
- Wang, J.; Ma, S.; Yu, P.; He, X. Evolution of Human Brain Left–Right Asymmetry: Old Genes with New Functions. Mol. Biol. Evol. 2023, 40, msad181. [Google Scholar] [CrossRef] [PubMed]
- Cutia, C.A.; Leverton, L.K.; Christian-Hinman, C.A. Sex and Estrous Cycle Stage Shape Left-Right Asymmetry in Chronic Hippocampal Seizures in Mice. eNeuro 2023, 10, ENEURO.0041-23.2023. [Google Scholar] [CrossRef]
- Shiptona, O.A.; Tanga, C.S.; Paulsena, O.; Vargas-Caballero, M. Differential vulnerability of hippocampal CA3-CA1 synapses to Ab. Acta Neuropathol. Commun. 2022, 10, 45. [Google Scholar] [CrossRef]
- Hanane, U.; Ben-Moshe, A.; Diamant, H.; Markovich, G. Edited by Lia Addadi, Weizmann Institute of Science, Rehovot, Israel. Spontaneous and directed symmetry breaking in the formation of chiral nanocrystals. Proc. Natl. Aacd. Sci. USA 2019, 116, 11159–11164. [Google Scholar] [CrossRef]
- Higginbotham, H.R.; Gleeson, J.G. The centrosome in neuronal development. Trends Neurosci. 2007, 30, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, A.; Fornasiero, E.F.; Dotti, C.G. Cadherins as regulators of neuronal polarity. Cell Adh. Migr. 2015, 9, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Lubben, N.; Ensink, E.; Coetzee, G.A.; Labrie, V. The enigma and implications of brain hemispheric asymmetry in neurodegenerative diseases. Brain Commun. 2021, 3, fcab211. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dyakin, V.V.; Dyakina-Fagnano, N.V. Enigma of Pyramidal Neurons: Chirality-Centric View on Biological Evolution. Congruence to Molecular, Cellular, Physiological, Cognitive, and Psychological Functions. Symmetry 2024, 16, 355. https://doi.org/10.3390/sym16030355
Dyakin VV, Dyakina-Fagnano NV. Enigma of Pyramidal Neurons: Chirality-Centric View on Biological Evolution. Congruence to Molecular, Cellular, Physiological, Cognitive, and Psychological Functions. Symmetry. 2024; 16(3):355. https://doi.org/10.3390/sym16030355
Chicago/Turabian StyleDyakin, Victor Vasilyevich, and Nika Viktorovna Dyakina-Fagnano. 2024. "Enigma of Pyramidal Neurons: Chirality-Centric View on Biological Evolution. Congruence to Molecular, Cellular, Physiological, Cognitive, and Psychological Functions" Symmetry 16, no. 3: 355. https://doi.org/10.3390/sym16030355