4.1. Behavioural Data
We have applied the classical analytical pipeline normally applied in morphometric studies where σp2 and FA are utilized for estimating DI for behavioural data. We have chosen completely inbred lines because σp2 can be used as an estimator of DI if the influence of σe2 is minimal or equal to zero. However, the FA indices can be considered reliable estimators of DI even if σg2 ≠ 0, which is the most common situation when dealing with sexually reproducing organisms. We have converted locomotor behaviour into a deviation from an expected value of 0; however, there are many other possible conversions of behavioural data, as, for example, the number of times in a given interval of time or the time spent holding the head on the right or on the left compared to the body of the axis of a bilateral animal. Another possibility is to estimate the number of times in a given interval of time or the time spent by a grazer in holding the head down and graze compared to when the grazer holds the head up. The concept of behavioural instability can also be applied to the behaviour of animals, which can be tracked with GPS or with a radar system counting the number of times that the individual is turning to the right or to the left in different environmental conditions.
4.2. Phenotypic Variance
Several significant differences between the medians of the total activity were found (
Table 1), and these differences can be attributed to genetic differences among the different lines as the σ
e2 is expected to be negligible. However, we have shown that the σ
e2 is not negligible, as in five (lines 385, 392, 491, 508, 584 and 703) of the investigated lines, significant negative correlations between clockwise (CW) and counter-clockwise (CCW) movements were found producing a bias of both the medians of the total activity but also on the σ
p2 and FA estimates.
Therefore, these lines should be excluded from the statistical analyses, or, if possible, the experiments should be repeated on these lines until the correlation between CW and CCW disappear.
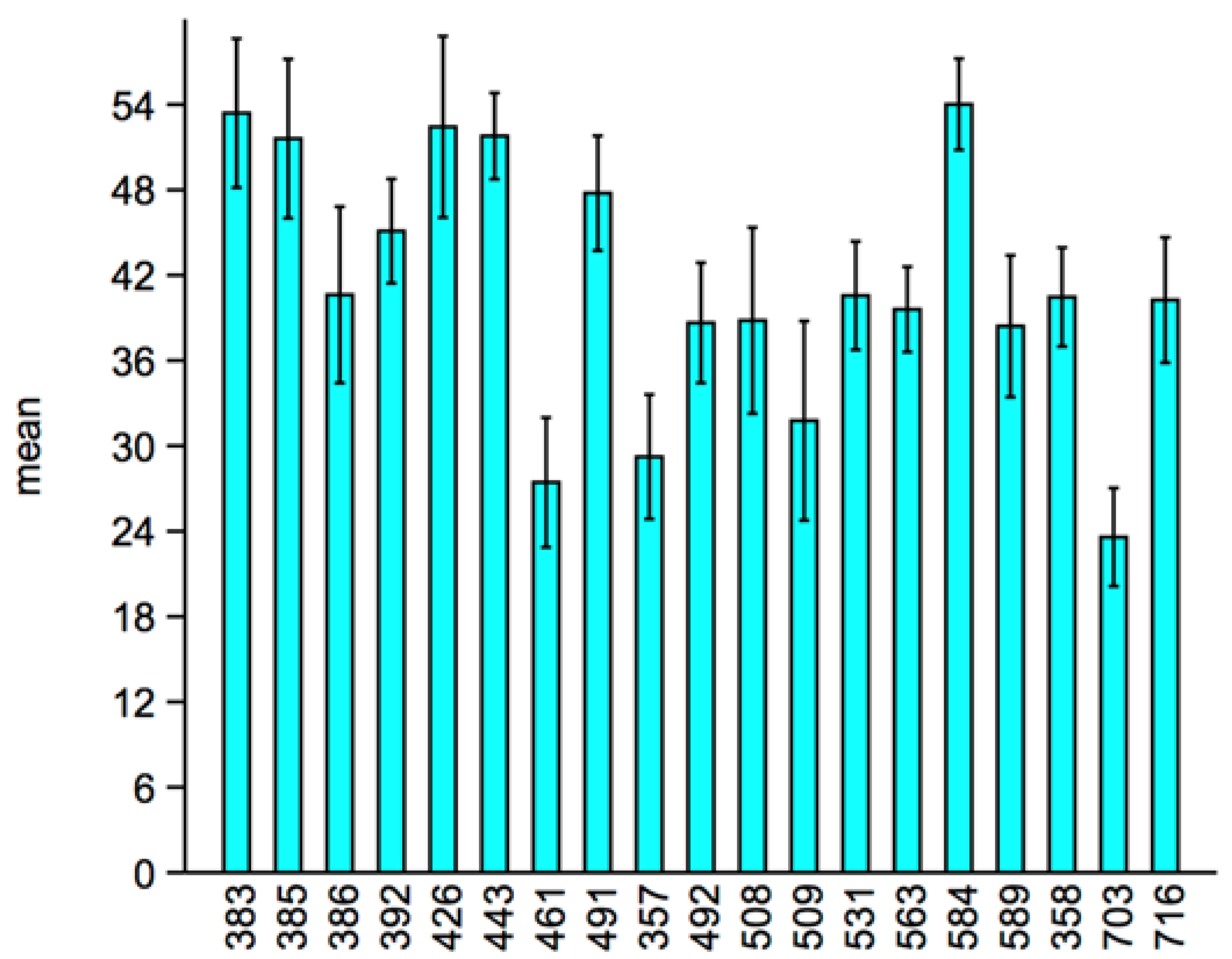
The
F-tests conducted on the log-transformed values of the variance of the total activity of the 19 lines revealed several significant differences of σ
p2 as shown in
Table 3 and
Figure 1. The heterogeneity of the 95% confidence intervals indicate large differences of σ
p2, and several of these differences were also found between lines in which σ
e2 was negligible. Hence, we can consider these differences as differences in the degree of DI, and the lines in which the presence of σ
e2 will produce biased estimates of σ
p2 will be overestimated in the case of positive covariance between the CW and CCW or underestimated in the case of negative correlation between the CW and CCW (which is our case).
Some deviations from normal distributions of the distributions of the total activity were found. However, after the correction for multiple comparisons, none of the 19 lines showed significant deviations. These results should be interpreted with caution as the Bonferroni correction can be quite conservative when several tests are conducted, and, therefore, we should consider experiments where hypotheses requiring pairwise comparisons are utilized in order to reduce the chance of committing error type II [
28]. The presence of σ
e2 should, in fact, reduce the kurtosis making the distributions platykurtic, and the fact that we have not been able to detect significant deviations from normality does not mean that a deviation will be found if we have increased the number of replicates per line. However, the fact that we were able to detect the presence of σ
e2 by testing the correlations between the CW and CCW is clearly showing that the correlational analysis is more sensible even with small sample size (see [
12], for details of the method).
4.3. Fluctuating Asymmetry
Significant differences among lines were found both for the FA1 and FA3 indices, and
Table 5 and
Table 6 are showing which of the pairwise comparisons are significant. Notably, the matrices shown in
Table 5 and
Table 6 of the pairwise comparisons between lines of the FA1 and FA3 indices are not always concordant and sometimes different significant comparisons were found. This is not surprising, as the FA3 index is a scaled index, which is correcting for the mean. However, this index should only be used when a significant correlation between FA and the mean is found (see below). What is also notable is that the pairwise comparisons between the medians (
Table 1) and the σ
p2 (
Table 3) of the total activity are also not always concordant with the results found in
Table 5 and
Table 6. The medians of the total activity are not an estimator of DI, but σ
p2 should hypothetically find the same differences found by the FA estimators especially in the absence of a scaling effect. The reasons for the discrepancy could be due to the sampling error of the variance [
29].
4.4. Directional Asymmetry and Antisymmetry
No sign of directional asymmetry (DA) has been found in this investigation, and a debate continues concerning the reliability of DA as a measure of DI [
14]. We should consider the possibility that directional asymmetry could appear also when looking at behavioural data. Directional asymmetry could potentially be present in experiments in very stressful environments, such as [
16] regarding transitions from FA to DA as responses to heavy stress. In any case, as DA may have some heritable component [
2], it cannot be excluded that a genetic pool could produce such a phenomenon even when looking at behavioural traits [
2].
The fact that no significant antisymmetry has been detected could once again be due to some of the reasons mentioned above.
A negative correlation between
r and
l (which is our case) can, in fact, generate a platykurtic distribution, and, at the same time, it overestimates the FA1 index [
1]. For this reason, the five lines (lines 385, 392, 491, 508, 584 and 703) of the investigated lines indicate a significant negative correlation between clockwise (CW) and counter-clockwise (CCW) movement, which, as we previously argued (or the experiment should be repeated), should also be eliminated when estimating the FA1 and FA3 indices.
4.6. Perspectives
Potentially, there is a very promising application of FA and σ
p2 as an indicator of behavioural instability if we follow the correct procedures described in this paper. The concept of behavioural instability could have several possible applications as a biomonitoring tool if we expect that changes in environmental conditions can produce changes in the behavioural traits. For example, behavioural traits have been used in toxicological studies as a sensitive indicator of stress using traits such as swimming activity, locomotor activity or flight [
30,
31,
32,
33]. Behavioural instability provides another way of analysing data compared to traditional measures such as total distance traveled, activity per hour or swimming distance. Likewise, behavioural traits have been used in evolutionary studies to evaluate plasticity and genotypic difference or in psychological studies [
34]. Similarly, several genome-wide association studies (GWAS) studies are trying to associate genetic variation with variation in behavioral traits [
35]. These studies will clearly be beneficial if it is possible to associate the concept of behavioural instability with genetic variation. In addition, the procedure described in this paper could also be useful for excluding samples where σ
e2 is present, as it will bias the GWA analysis due to the fact that σ
e2 will, most of the time, increase the sizes of the traits and therefore invalidate the analysis.




{kind=link}