1. Introduction
Anthropogenic activities, such as the production of municipal wastes, burning of fossil fuels, mining activities, use of pesticides, and fertilizers in agricultural practice, etc., [
1], are a source of contamination by harmful elements that can disperse into the atmospheric, terrestrial, and aquatic ecosystems [
2,
3,
4]. This environmental threat has resulted in the institution of different legislation controls and in the development of several types of remediation actions that include chemical, physical, and biological methods [
5,
6,
7,
8]. Among the bio-techniques, phytoremediation exploits the ability of plants to concentrate harmful elements in their tissues (phytoextraction) or to stabilize them into new mineral forms in the soil and root tissues (phytostabilization) [
1,
9].
The cost-effective application of phytoremediation methods requires an accurate understanding of the processes ruling metal distribution and their chemical speciation in the soil-plant system. The rhizosphere is the narrow soil zone acting as the interface between soil, plant roots, microbes, water, and air [
10]. Processes occurring at the rhizosphere depend on several factors, both biotic and abiotic, and can deeply affect metal mobility. The mechanisms by which plants can tolerate excess of metals have been investigated with an increasing interest in the last decades, and the research is often focused on pioneer plants that grow on low-function (lack of nutrients and organic matter) and heavily polluted soils. To support their resilient behavior, pioneer plants could be able to: (i) sequester metals in organs or subcellular compartments (i.e., vacuoles) with little or no sensitive metabolic activity; (ii) chelate metals with exudates, phytochelatins, and peptides; (iii) mediate biomineral formation [
11,
12,
13,
14].
Several types of biominerals related to physiological needs and environmental stresses, i.e., phytoliths, cystoliths, and mineralized trichomes [
15], can form in plant tissues [
11,
16]. Biominerals in plants belong to many different classes; the most common are Ca oxalate, Ca carbonate, and silica ([
11] and references therein). Calcium [
17], Sr and Ba [
18] sulfates, Ca phosphate [
19], and Mg and Sr oxalates [
20] have been observed in some plant species. Other biominerals found in plants grown on polluted substrates are jarosite (KFe
3(OH)
6(SO
4)
2) [
21], Fe oxides [
22], hemimorphite-like phase [
23], Zn-rich phyllomanganate [
24], Fe plaques composed mainly of Fe, S, and K [
25], Zn and Cd incorporated in calcite [
26], or Cd incorporated in vaterite [
27].
Besides the chemical composition, plant biominerals respond to specific physiological functions changing their shape, size, and localization [
11]. For example, several studies have demonstrated the role of Ca oxalate in Al and metal (e.g., Sr, Cd, and Pb) detoxification [
28]. However, in leaves of common bean (
Phaseolus vulgaris L.), addition of metals such as Zn may decrease the number of Ca oxalate crystals, without showing incorporation of metals in crystals, suggesting that deposition of metals in Ca oxalate is both metal- and plant species-specific [
11]. Coprecipitation of Al and other metals with Si may be responsible for the alleviation of their toxicity [
11] via: (i) the complexation or co-precipitation of metals with Si; (ii) the immobilization of metals in growth media; (iii) the compartmentation of metals within plants; (iv) uptake processes [
29,
30,
31,
32]. Beyond Ca-oxalate and silica, organic molecules (e.g., cysteine, histidine, organic acids, etc.), Ca carbonate and sulfate biominerals may play a role in detoxifying metals [
11,
18,
20].
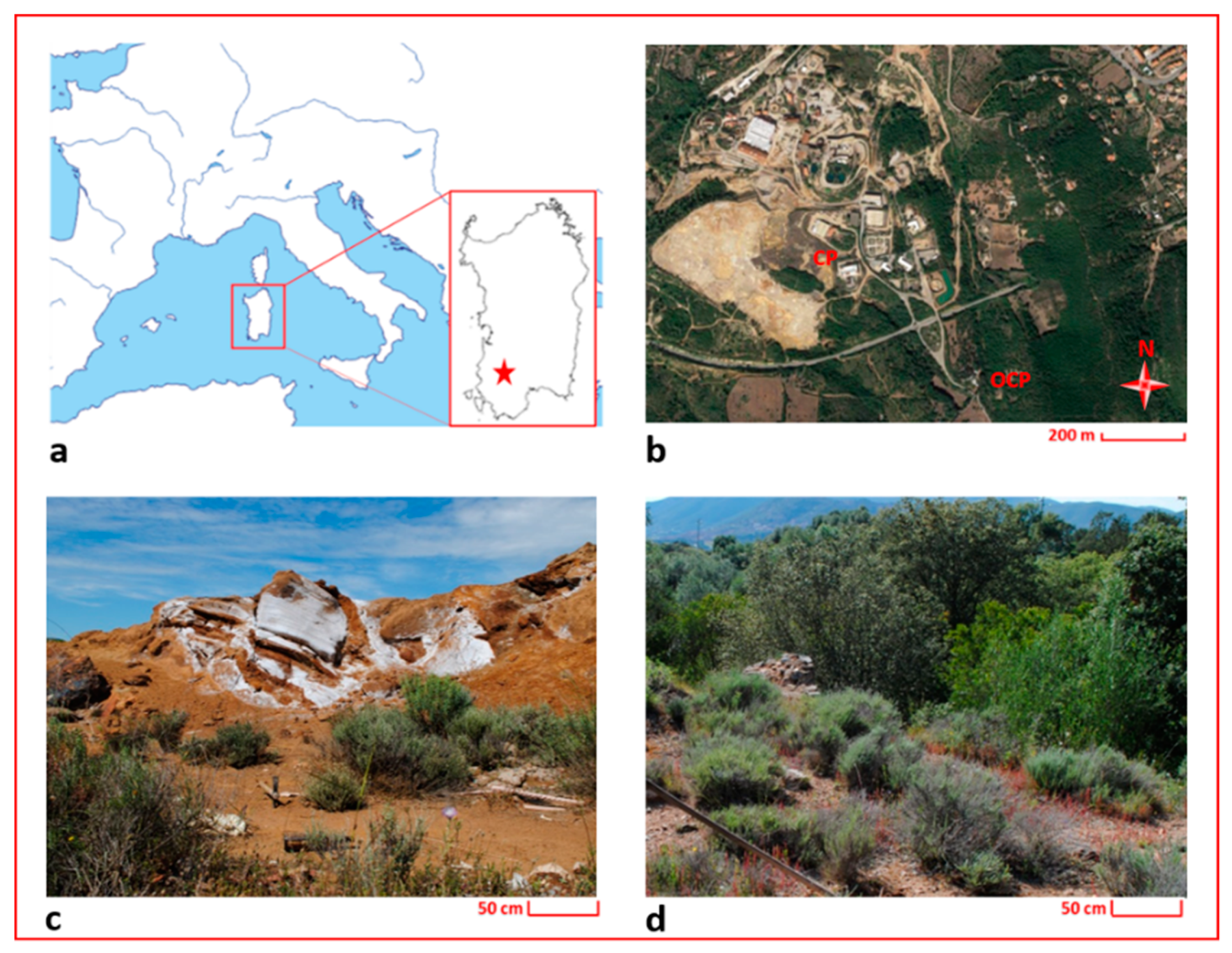
During the 19th and the 20th centuries, Sardinia (Italy) was one of the most prominent mining poles in Europe [
33], and after the closure of mines, only few remediation actions were applied. The largest amount of mine wastes in Sardinia occurs in the Rio San Giorgio catchment basin (Sulcis-Iglesiente district, SW Sardinia), where about 17 million m
3 of open-pit excavations and 13 million m
3 of dumps and tailings have been stored in dams characterized by inadequate containment control or dispersed into the rivers and lagoons [
4,
34,
35,
36,
37]. These wastes have a highly contamination potential due to the high concentrations of metals [
38] and the predominantly thin size of their particles that facilitate the aeolian dispersion [
39].
In Sardinia, several autochthonous vascular plants are able to spontaneously colonize mine wastes and tolerate high concentrations of metals, such as
Pistacia lentiscus L. [
40],
Euphorbia pithyusa subsp.
cupanii (Guss. ex Bertol.) Radcl.-Sm. [
9],
Phragmites australis (Cav.) Trin. ex Steud.,
Helichrysum microphyllum Cambess. subsp.
tyrrhenicum Bacch., Brullo, and Giusso [
41] (hereafter referred to
H. tyrrhenicum), and
Epipactis helleborine (L.) Crantz subsp.
tremolsii (Pau) E. Klein [
42].
Helichrysum tyrrhenicum is an endemic perennial shrub of Sardinia and Corsica. It grows on different ecological conditions and substrates, especially muddy and sandy soils. This plant species is able to grow on metal contaminated substrates, and it is a pioneer plant in mine tailings [
43,
44], forming many plant assemblages typical of mine environments [
45], and making it a potential candidate for phytostabilization techniques [
39,
46,
47,
48].
Despite the development of our researches, the microscopic biomineralization processes ruling Zn bioavailability at the soil-root interface and in the tissues of
H. tyrrhenicum are still poorly known. The goal of this paper is to study the mineralogical evolution from soil to
H. tyrrhenicum tissues harvested from the mine dump of Campo Pisano (SW, Sardinia, Italy). Synchrotron radiation-based techniques represent the state of the art tools used to investigate the microscopic processes occurring in plant-soil systems [
49,
50,
51,
52]. The most commonly applied techniques in environmental sciences include X-ray diffraction (XRD), X-ray imaging, and X-ray absorption spectroscopy (XAS), providing the finest complementary details about the atomic and crystallographic structure, distribution of elements, their chemical speciation, and their valence state [
53]. Specifically, our investigation was carried out using a multi-method approach exploiting laboratory (chemical analysis, XRD, and scanning electron microscopy) and synchrotron radiation-based techniques (XAS, Infrared microspectroscopy, and soft X-ray microscopy combined with low energy X-ray fluorescence mapping) in order to achieve insight into the understanding of composition and structure of biomineralizations in
H. tyrrhenicum.
3. Results
3.1. Mineralogical Composition and Metal Contents
Table 1 shows mineral phases detected in some selected samples (CP1 and OCP1) that can be considered as representative, because no significant variations were observed in mineral composition. Quartz and dolomite were observed both in the bulk soil and rhizosphere samples from CP (Campo Pisano mine dump) and OCP (outside the mine dump of Campo Pisano), and they represent the gangue minerals of the ore deposits. Pyrite (FeS
2) was detected in soils and rhizospheres from CP and not in OCP samples, whereas gypsum (CaSO
4) and jarosite (KFe
3+3(SO
4)
2(OH)
6) were found in soil samples collected in the mine dump (CP). Smithsonite was found only in the soils collected outside the mine dump (OCP samples), probably resulting from the oxidation of the primary Zn sulfides [
57]. In plant samples, quartz, dolomite, weddellite (Ca(C
2O
4)·2(H
2O)), and amorphous cellulose were detected.
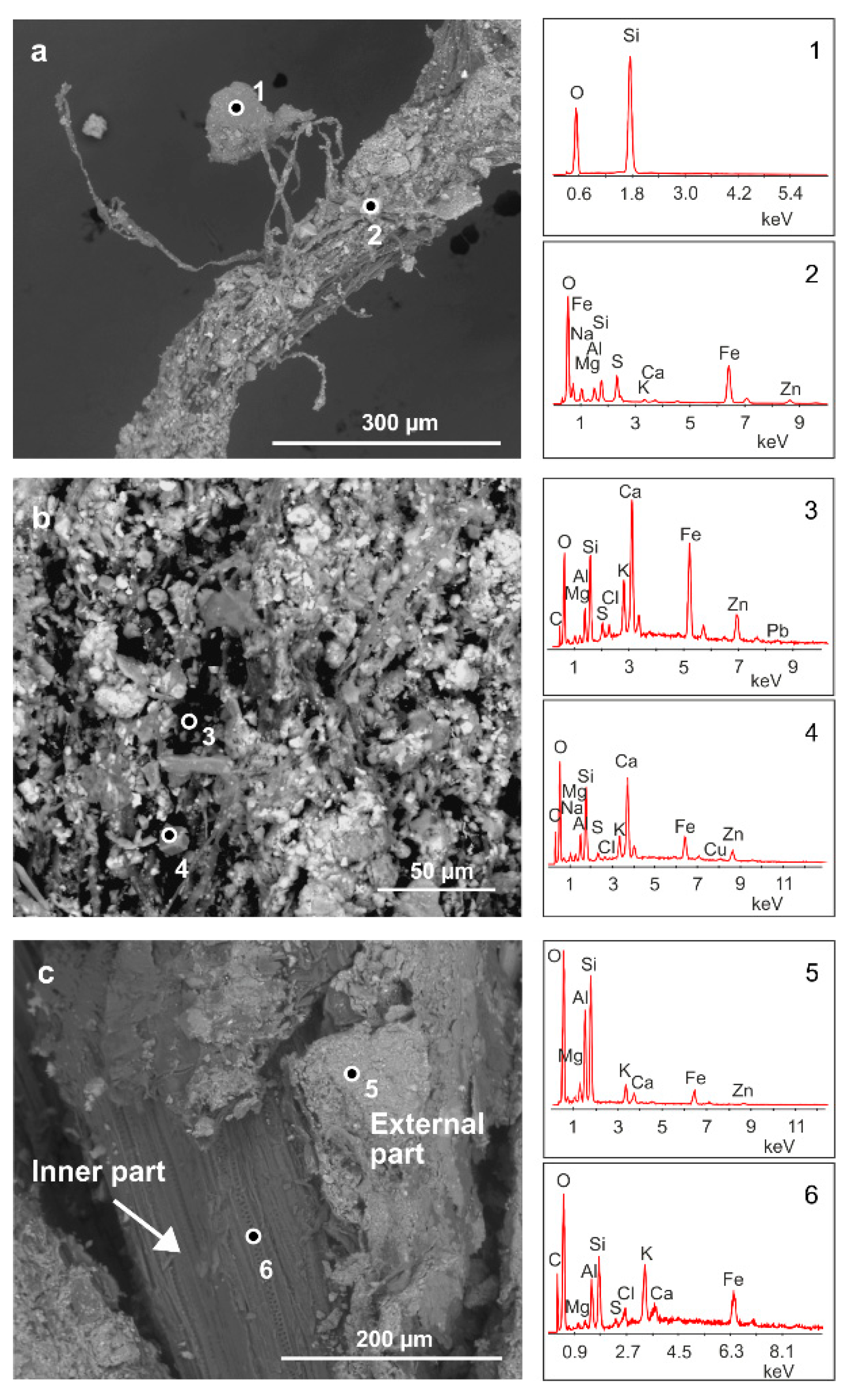
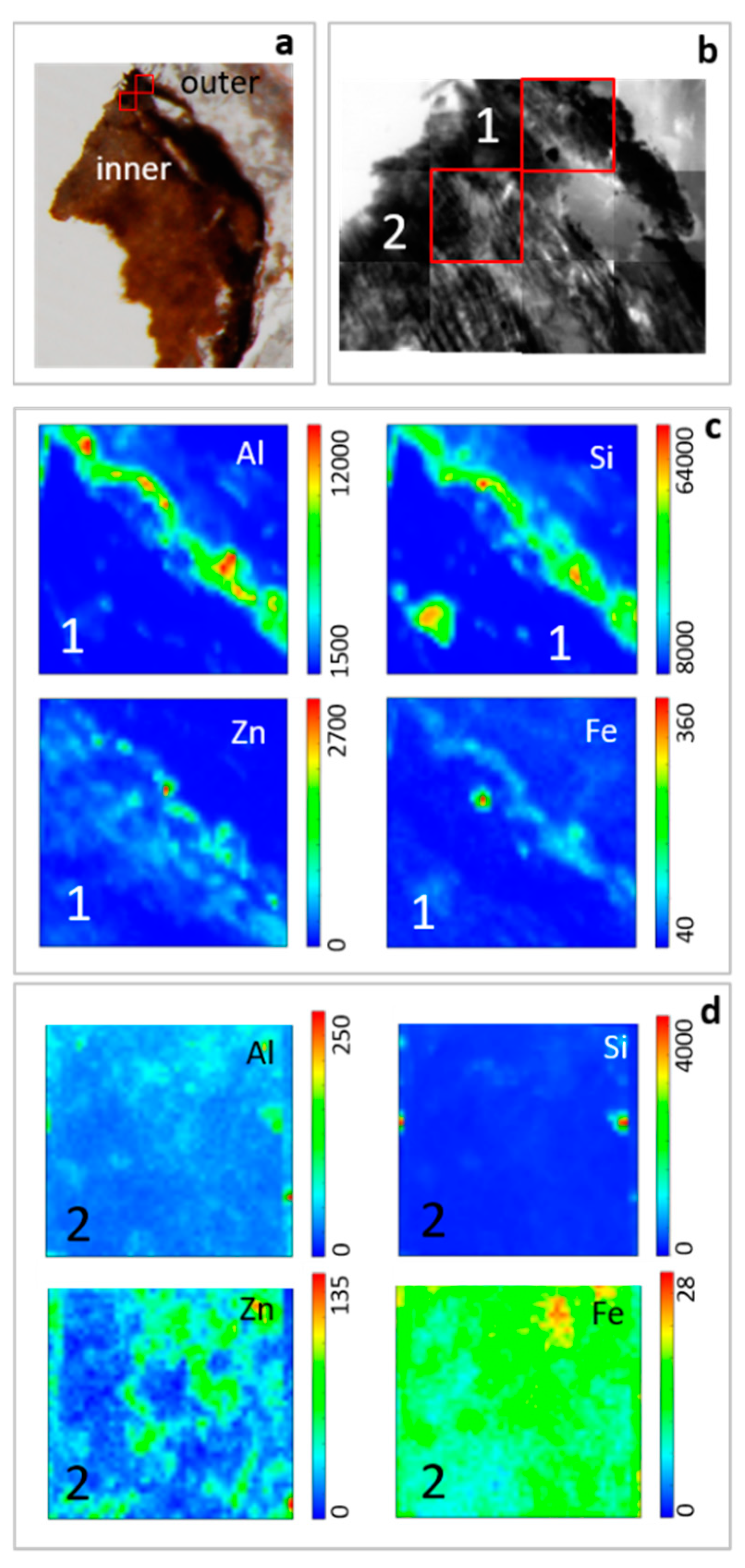
Figure 2 shows SEM analysis performed on some selected samples of the roots of
H. tyrrhenicum grown on the mine dump (CP). Plant roots embed mineral particles (
Figure 2a) that strongly adhere to their surface. We mainly recognized quartz and Al-silicates (
Figure 2a, points 1 and 2); also, Zn, Pb, and Fe were detected by EDS analysis in the rhizosphere grains located on the surface of the roots (
Figure 2b, points 3 and 4). In detail,
Figure 2c shows a longitudinal section of a plant root, where we can observe a mineral rim mainly made up of Si, Al, and O (
Figure 2c, point 5). In the inner part of the roots, we mainly detected Si, Al, O, and K (
Figure 2c, point 6).
Table 2 reports the mean values of Zn, Pb, and Cd concentrations measured in three specimens of
H. tyrrhenicum (roots and epigean organs), in the respective bulk soils and in the related rhizosphere materials. For comparison purposes, the (i) threshold contamination levels established for an industrial use of soil by the Italian laws (D.lgs 152/2006) [
90], and (ii) background values for the area are also reported. Zinc is the most abundant metal in the bulk soils (24,900
CP–32,700
OCP mg/kg), rhizospheres (26,300
CP–27,300
OCP mg/kg) and in the plant tissues (890
OCP–3290
CP mg/kg), followed by Pb (1240
OCP–5000
CP mg/kg in the soils, 1600
OCP–5030
CP mg/kg in the rhizospheres, and 50
OCP–1020
CP mg/kg in the plant tissues) and Cd (100
CP–340
OCP mg/kg in the soils, 170
CP–280
OCP mg/kg in the rhizospheres, and 13
OCP–31
OCP mg/kg in the plant tissues). It should be noted that metal contents detected in this work are of the same order of magnitude as in previous studies [
39,
41,
46,
91], in which relevant local variations due to heterogeneities of the mine tailings can be highlighted.
BCF, BAC, and TF (
Table 3) were calculated to investigate the transfer of metals from geosphere to plant tissues and their translocation from the roots to the epigean organs. BCF may be an indicator of the accumulation and phytostabilization potentialities of
H. tyrrhenicum. BCF
soil and BCF
rhizo values calculated in relation to Zn (Zn-BCF
soil-CP 0.13, Zn-BCF
rhizo-CP 0.13) and Pb (Pb-BCF
soil-CP 0.14, Pb-BCF
rhizo-CP 0.13) for samples collected inside the mining areas are higher than those calculated for the samples collected outside the mining site (Zn-BCF
soil-OCP 0.04, Zn-BCF
rhizo-OCP 0.05, Pb-BCF
soil-OCP 0.05, Pb-BCF
rhizo-OCP 0.04). Cadmium is characterized by very similar values of BCF
rhizo (Cd-BCF
rhizo-CP 0.14, Cd-BCF
rhizo-OCP 0.11) for the two sampling areas, whereas BCF
soil is higher in the CP samples (Cd-BCF
soil-CP 0.26) than in the OCP samples (Cd-BCF
soil-OCP 0.09). Considering the standard errors reported in
Table 3, the calculated values show no significant differences between BCF
soil and BCF
rhizo.
BAC values show that H. tyrrhenicum has a low capability (BAC ≤ 0.2) to accumulate the investigated metals in epigean organs. At the Campo Pisano mine, the highest value of BAC was observed for Pb (Pb-BACsoil-CP 0.2, Pb-BACrhizo-CP 0.2), followed by Zn (Zn-BACsoil-CP 0.12, Zn-BACrhizo-CP 0.12) and Cd (Cd-BACsoil-CP 0.16, Cd-BACrhizo-CP 0.09). OCP samples have lower BAC than CP samples, and they are characterized by very similar values for Zn, Pb, and Cd (Zn-BACOCP 0.03, Pb-BACOCP 0.03–0.04, Cd-BACOCP 0.04–0.05). As for the BCF values, Cd-BACsoil-CP (0.16) is higher than Cd-BACsoil-OCP (0.04), and no significant differences were observed between BACsoil and BACrhizo for the investigated areas.
TF values decrease in the order Pb-TF > Zn-TF > Cd-TF both for CP (Pb-TFCP 1.5, Zn-TFCP 0.93, Cd-TFCP 0.64) and OCP samples (Pb-TFOCP 0.76, Zn-TFOCP 0.63, Cd-TFOCP 0.42). H. tyrrhenicum specimens harvested inside the mining area show the highest translocation for Pb (Pb-TFCP 1.5).
Measurement of plant biomass (
Table 4) indicates that the root biomass is always lower than epigean biomass, in particular for OCP, where specimens show an important epigean development. From Zn concentrations in plant tissues (
Table 2), and assuming that 5 specimens of
H. tyrrhenicum can fit in 1 m
2, we estimated that this species could accumulate from 6 to 11 kg/ha of Zn (
Table 5).
3.2. Chemical Characterization of the Plant Tissues
Several sections of each
H. tyrrhenicum tissue were imaged and results of selected samples are shown in
Figure 3 and
Figure 4 (a larger view of the slices is presented in
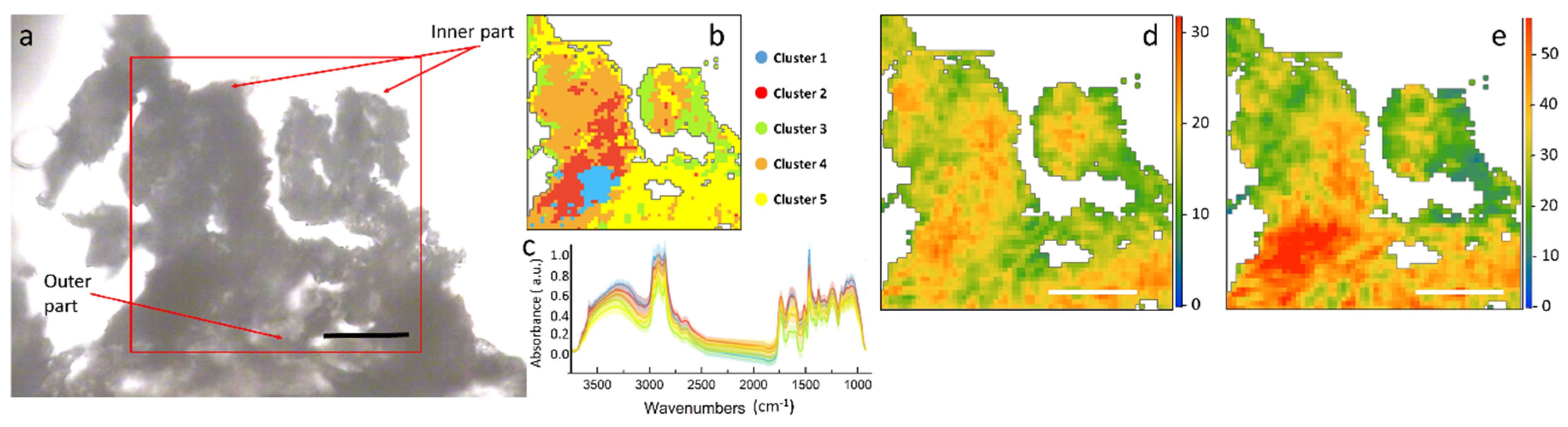
Figure S2 in Supplementary Materials). At first, the different areas of the plant were roughly identified through a visual inspection. In
Figure 3a, the optical image of a stem section is shown. This section was one of the best preserved, though some areas of the outer endothelium broke and folded on top of the other. Since the image was not clear, a cluster analysis was used to support the identification of the different areas of the plant section and to remove the background pixels as well as those containing mainly paraffin.
The results of the HCA are presented in
Figure 3b,c. From the color map, we can identify three main areas within the inspected sample: one represented by the yellow cluster, closer to the outer part of the section, could be the epidermis/cortex; one, orange, could be assigned to the endodermis; the red and blue clusters, almost in the inner part, could be the phloem and xylem, since they result also in aromatic polysaccharides.
The heat maps obtained by integrating the peaks of the main components of the plant cell wall, such as hemicellulose and ring in plane vibrations of aromatics, are presented in
Figure 3d,e. These maps represent the spatial distribution of the different macromolecules within the stem section. Assuming a constant thickness of the thin section, the intensity of the colors in the maps is proportional to the chemical concentration. From the chemical profiles in
Figure 3, it can be seen that aromatics have a hot spot in the center of the xylem. In almost all of the stem sections of this set of samples the external layer of epidermis was heavily affected by the paraffin.
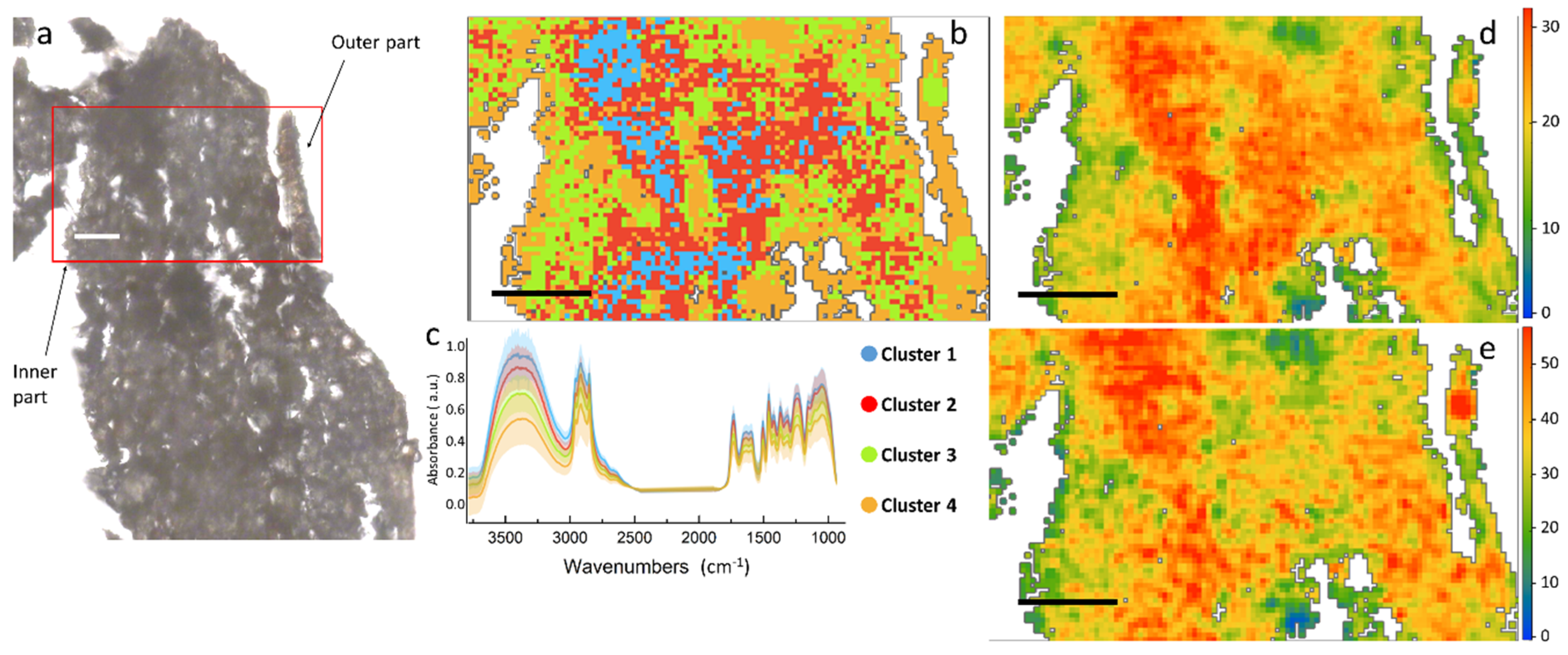
Similar information on the chemical distribution of the main cell wall components can be obtained by the analysis of
Figure 4, which represents a cross section of the root. Differently from stem, aromatics and C=O signals for root in panels 5d and 5e tend to co-localize, presenting a similar spatial distribution in the left part of the section. From the HCA map it is possible to identify two macro areas, one represented by cluster 1 and 2, mostly present in the center of the map, and a second district characterized by the presence of clusters 3 and 4. Tentatively, we can assign it to the outer right part of the epidermis (clusters 3 and 4), and the central part more rich in proteins (clusters 1 and 2).
Comparing the centroid plots in
Figure 3c (for stem) and
Figure 4c (for root), it is possible to appreciate some spectral differences. The root is richer than the stem in carbohydrates, since both the –OH stretching at 3500 cm
−1 and the C–O–C stretching at ~1180–1000 cm
−1 are more intense that the C=O at 1750 cm
−1 in the root. On the contrary, if we compare the same signals in the stem, the C=O results almost as high as the OH and C–O–C. From the maps in
Figure 3d and
Figure 4d it can be seen that even though the maximum value of the C=O band is comparable between the two regions of the plant, this chemical moiety is more diffused into the overall root section, whereas in the stem it is limited only to a small area in the inner part. This can be a symptom of higher oxidative stress within the root in respect to the stem [
93,
94].
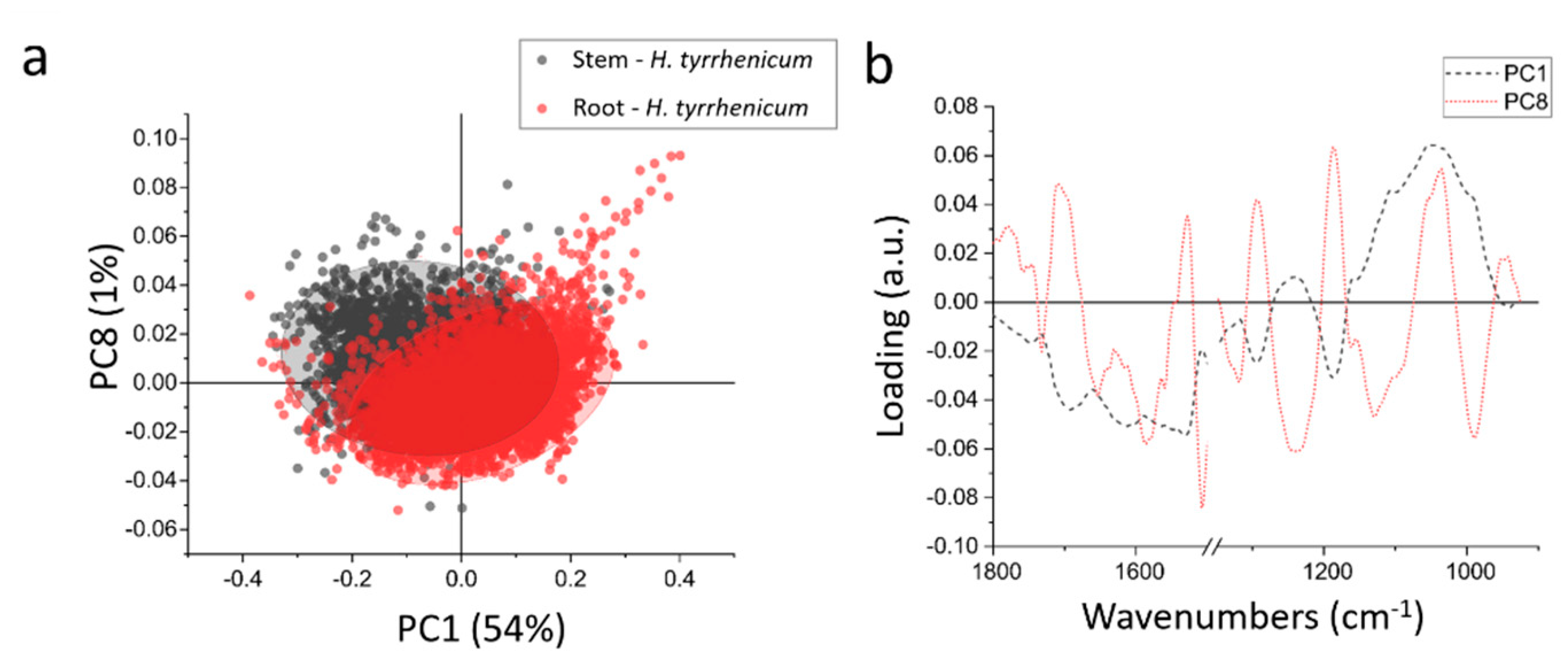
In order to better highlight the chemical differences between root and stem, the data were normalized and then subjected to PCA.
Figure 5a shows the scatter plot resulting from the PCA; the root and stem spectra do not differ much from the chemical point of view and only a partial separation can be achieved along PC1 (which accounts for ~54% of the variance) and PC8 (~1% of the variance). In
Figure 5b the loadings vectors of PC1 and PC8 are presented. PC1 mainly takes into account the variation in carbohydrates that was discussed in the previous paragraph, when comparing the centroids of the HCA. Summarizing, the roots are richer in carbohydrates than the stems.
PC8 accounts only for 1% of the variance, thus the spectral variations that it represents can be barely appreciated in the average spectra from the samples. It is a quite complex loading, with 5 main positive peaks and 6 main negative peaks: 1710 cm−1, 1525 cm−1, 1290 cm−1, 1185 cm−1, and 1036 cm−1 are the positive peaks, whereas 1652 cm−1, 1580 cm−1, 1508 cm−1, 1239 cm−1, 1129 cm−1, and 968 cm−1 are negative ones. The negative peaks like 1652 and 1508 cm−1, can be interpreted as a minor content of alpha helix proteins in the root in respect of the stem. The same can be seen in the signals at 1239–1129 cm−1 that can be due to a decrease in phosphates. The positive peaks, instead, represent a higher amount of those specific chemical moieties in the roots in respect to the stem. Like the band at 1710 cm−1, that represents some more oxidized species, that are more widespread in the roots, or the peaks at 1290 cm−1, 1185 cm−1, and 1036 cm−1, that can be assigned to cellulose.
3.3. Element Distribution and Zn Chemical Speciation
Thin cross sections of roots were analyzed by STXM coupled with LEXRF to investigate the distribution of Si, Al, Zn and Fe (
Figure 6). Silicon and Al are mainly concentrated on the root surface, forming a rim on the epidermis according to SEM observations. Generally, their content decreases from the external part toward the inner zone, and also, they can occur as concentrated spots in the internal part of the roots. Zinc and Fe are mainly located in the external part of the root but their variation in distribution is less marked than Al and Si.
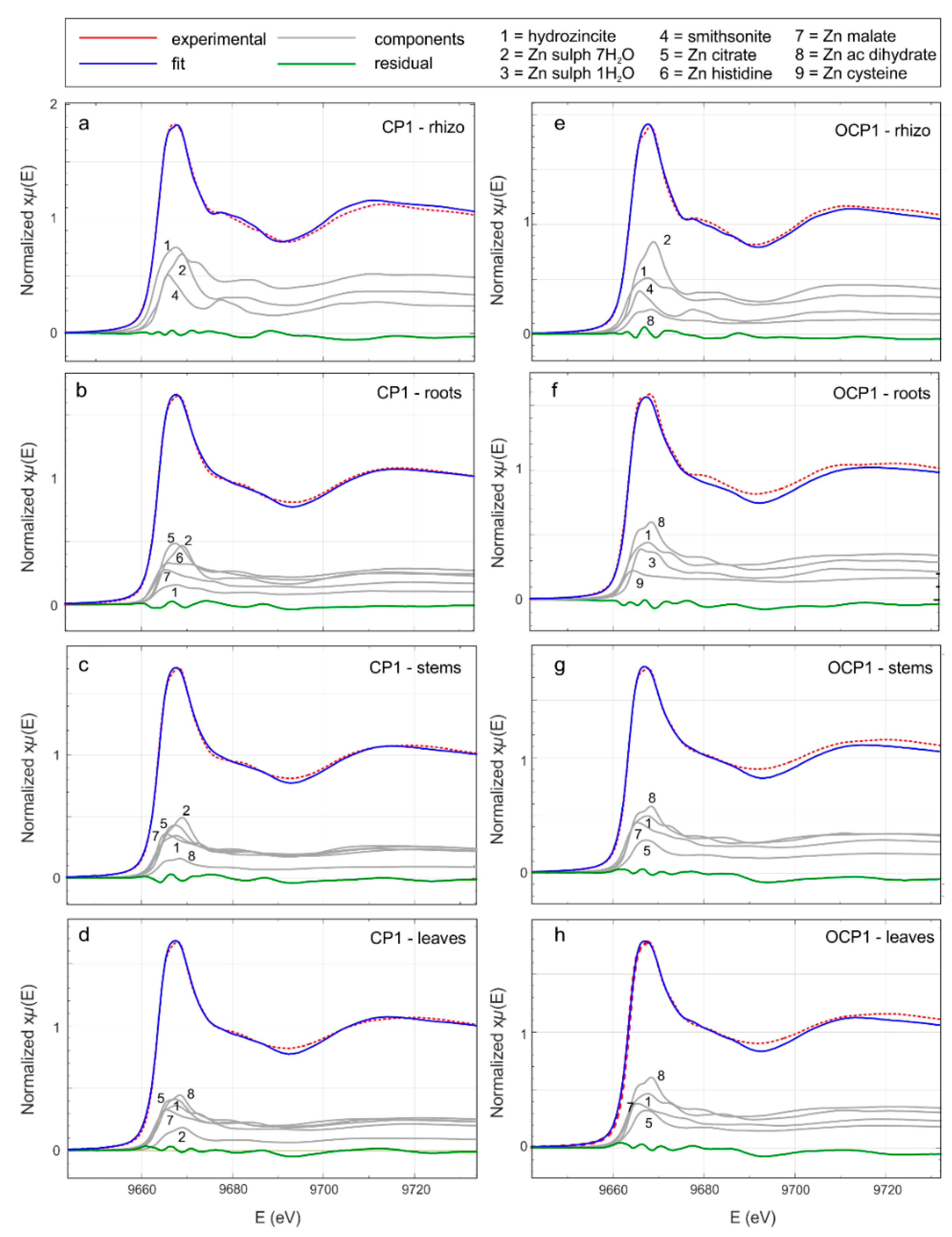
Zn K-edge XANES analysis was performed to investigate the average chemical environment of Zn. The quantitative analysis of the XANES region of biological samples is a complex task and the comparison of experimental spectra with reference compounds can help to shed light on their average mineralogical environment.
Figure S1 and
Figure 7 show the Zn-K edge absorption spectra in the XANES region of the reference compounds and selected rhizosphere and plant samples, respectively. XANES spectra of the rhizospheres and plant tissues are smoother and broader than the reference compounds, suggesting that in the investigated samples Zn occurs in a more disordered chemical/coordinative environment. Also, the XANES spectral features are more pronounced in the rhizosphere spectra than in the plant samples, indicating that the local structure around Zn becomes progressively more disordered during its transfer from the geosphere to the biosphere.
Qualitative comparison of the XANES spectra (
Figure S1 and
Figure 7) suggests that in rhizosphere and plant samples, Zn probably occurs in different chemical environments. To obtain information about the average Zn coordination chemistry, we performed a LCA, using the reference compounds from
Figure S1. A trial and error procedure allowed the selection of a minimal subset of reference compounds; the results are shown in
Figure 7 and
Table 6. In all the rhizosphere samples (
Figure 7a,e), the main contribution comes from smithsonite, hydrozincite, and Zn sulfate heptahydrate. In the fitting procedure, the contribution of Zn acetate dihydrate proved to be relevant in the rhizosphere solid materials from the OCP site. Moving toward the roots (
Figure 7b,f), stems (
Figure 7c,g), and leaves (
Figure 7d,h), the contribution of smithsonite disappears and the addition of Zn organic compounds (Zn citrate, Zn histidine, Zn acetate dihydrate, Zn malate, and Zn cysteine) becomes necessary to achieve a good fit.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}