
Cyanobacterial Communities of Carbonate Sediments and Biomineralization in Peterhof Fountains’ Water Supply System, Russia

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
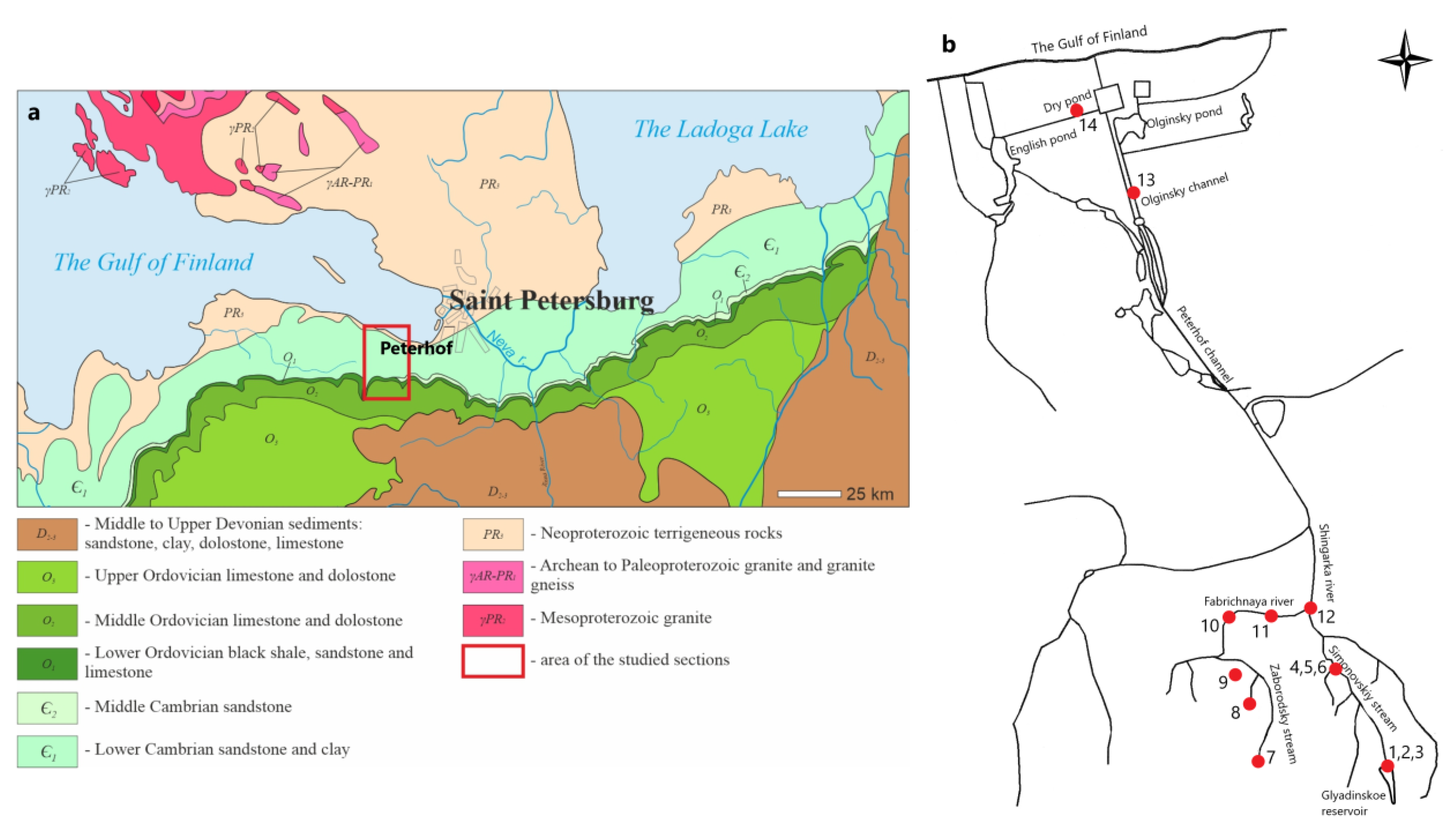
2.1. Study Area and Sample Collection
2.2. Identification of Cyanobacteria
2.3. Laboratory Experiments on Biomineral Precipitation
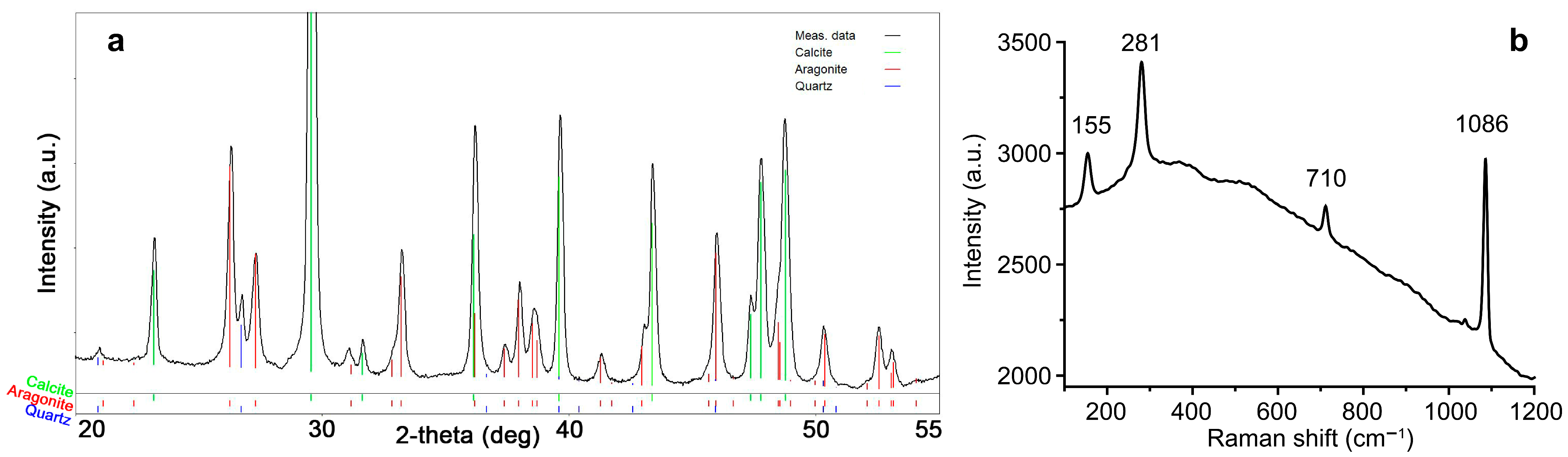
2.4. Mineral Identification
3. Results and Discussion
3.1. Mineral Composition of Sediments in Peterhof Fountains’ Water Supply System
3.2. Composition of Microbial Communities in Peterhof Fountains’ Water Supply System
3.3. Carbonate Precipitation in Fresh Water with the Participation of Cyanobacterial Species In Situ and In Vitro
- Cultures with Pleurocapsa sp., Scytonema ocellatum, Phormidium sp. (CALU 1550);
- In experiments without marble pieces with all cyanobacterial strains;
- In control (Gromov-6 medium with marble piece).
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Janssen, A.; Swennen, R.; Podoor, N.; Keppens, E. Biological and Diagenetic Influence in Recent and Fossil Tufa Deposits from Belgium. Sediment. Geol. 1999, 126, 75–95. [Google Scholar] [CrossRef]
- Pedley, M. Freshwater (Phytoherm) Reefs: The Role of Biofilms and Their Bearing on Marine Reef Cementation. Sediment. Geol. 1992, 79, 255–274. [Google Scholar] [CrossRef]
- Pentcost, A. Travertine; Springer: Berlin/Heidelberg, Germany, 2005; 445p, ISBN 978-1-4020-3523-4. [Google Scholar]
- Irion, G.; Müller, G. Mineralogy, Petrology and Chemical Composition of Some Calcareous Tufa from the Schwäbische Alb, Germany. In Recent Developments in Carbonate Sedimentology in Central Europe; Müller, G., Friedman, G.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1968; pp. 157–171. ISBN 978-3-642-88054-4. [Google Scholar]
- Pedley, M.; Rogerson, M.; Middleton, R. Freshwater Calcite Precipitates from in Vitro Mesocosm Flume Experiments: A Case for Biomediation of Tufas. Sedimentology 2009, 56, 511–527. [Google Scholar] [CrossRef]
- Kleinteich, J.; Golubic, S.; Pessi, I.S.; Velázquez, D.; Storme, J.-Y.; Darchambeau, F.; Borges, A.V.; Compère, P.; Radtke, G.; Lee, S.-J.; et al. Cyanobacterial Contribution to Travertine Deposition in the Hoyoux River System, Belgium. Microb. Ecol. 2017, 74, 33–53. [Google Scholar] [CrossRef]
- Roche, A.; Vennin, E.; Bundeleva, I.; Bouton, A.; Payandi-Rolland, D.; Amiotte-Suchet, P.; Gaucher, E.C.; Courvoisier, H.; Visscher, P.T. The Role of the Substrate on the Mineralization Potential of Microbial Mats in A Modern Freshwater River (Paris Basin, France). Minerals 2019, 9, 359. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, P.; Casamatta, D.A.; Hašler, P.; Jahodářová, E.; Norwich, A.R.; Poulíčková, A. Diversity of the Cyanobacteria. In Modern Topics in the Phototrophic Prokaryotes; Hallenbeck, P.C., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–46. ISBN 978-3-319-46259-2. [Google Scholar]
- Kamennaya, N.; Ajo-Franklin, C.; Northen, T.; Jansson, C. Cyanobacteria as Biocatalysts for Carbonate Mineralization. Minerals 2012, 2, 338–364. [Google Scholar] [CrossRef]
- Vidal, L.; Ballot, A.; Azevedo, S.M.F.O.; Padisák, J.; Welker, M. Introduction to cyanobacteria. In Toxic Cyanobacteria in Water, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: Boca Rataon, FL, USA, 2021; pp. 163–211. ISBN 978-1-00-308144-9. [Google Scholar]
- Katkova, V.I.; Mityusheva, T.P.; Filippov, B.N.; Simakova, Y.S. Mineral inclusions in cyanobacretia from water objects of southern Komi Republic. Vestn. Kolskogo Nauchnogo Cent. RAN 2017, 9, 64–70. (In Russian) [Google Scholar]
- Golubić, S.; Violante, C.; Plenković-Moraj, A.; Grgasović, T. Travertines and Calcareous Tufa Deposits: An Insight into Diagenesis. Geol. Croat. 2008, 61, 363–378. [Google Scholar] [CrossRef]
- Kolokoltsev, V.G.; Nikitin, M.Y.; Kovalevskaya, E.O. Modern travertines in the St. Petersburg area. Priroda 2014, 7, 17–29. (In Russian) [Google Scholar]
- Li, X.; Luo, K.; Ren, J.; Wang, X.; Mu, Q.; Fan, W. Characterisation of Extracellular Polymeric Substances from Different Cyanobacterial Species and Their Influence on Biocalcification Processes. Environ. Chem. 2017, 14, 254. [Google Scholar] [CrossRef]
- Vereshchagin, O.S.; Frank-Kamenetskaya, O.V.; Kuz’mina, M.A.; Chernyshova, I.A.; Shilovskikh, V.V. Effect of Magnesium on Monohydrocalcite Formation and Unit-Cell Parameters. Am. Mineral. 2021, 106, 1294–1305. [Google Scholar] [CrossRef]
- Ryabova, V.N.; Vasilieva, V.A. Vegetation restoration in recultivated ponds of the Lugovoy park (Petergof ’s fountains water supply system). Biol. Commun. 2014, 4, 49–66. (In Russian) [Google Scholar]
- Potravnov, A.L.; Khmelnik, T.Y. From the Kovashi River to the Samson Bowl: The History of the Water Supply System of the Peterhof Fountains; Avrora: St. Petersburg, Russia, 2019; ISBN 978-5-7300-0984-4. (In Russian) [Google Scholar]
- Yu, N.M. Travertinogenesis of the Izhora Plateau in the Holocene: Dissertation of the Candidate of Geographical Sciences: 25.00.25/Nikitin Mikhail Yurievich; St. Petersburg State University: St. Petersburg, Russia, 2015; 197p. (In Russian) [Google Scholar]
- Vereshchagin, O.S.; Frank-Kamenetskaya, O.V.; Shumilova, K.V.; Khadeeva, N.Y. Carbonate Sediments on Decorative Fountains in Peterhof, Russia. Environ. Earth Sci. 2018, 77, 56. [Google Scholar] [CrossRef]
- Payandi-Rolland, D.; Roche, A.; Vennin, E.; Visscher, P.T.; Amiotte-Suchet, P.; Thomas, C.; Bundeleva, I.A. Carbonate Precipitation in Mixed Cyanobacterial Biofilms Forming Freshwater Microbial Tufa. Minerals 2019, 9, 409. [Google Scholar] [CrossRef] [Green Version]
- Climate of Orzhitsy Village, Lomonosov District, Leningrad Region. Available online: https://goodmeteo.ru/pogoda-orzhitsy-lomonosovskiy-leningradskaya/god/ (accessed on 21 September 2021).
- Popov, L.E.; Khazanovich, K.K.; Borovko, N.G.; Sergeeva, S.P.; Sobolevskaya, R.F. Reference Sections and Stratigraphy of the Cambro-Ordovician Phosphorite-Bearing Shell in the Northwest of the Russian Platform; Nauka: Leningrad, Russia, 1989; 222p, ISBN 978-5-02-024551-8. (In Russian) [Google Scholar]
- Selivanova, A.V. Ordovician system. In Geology of the USSR. Vol. 1: Leningrad, Pskov and Novgorod Regions. Geological Description; Sidorenko, A.V., Ed.; Nedra: Moscow, Russia, 1971; pp. 127–173. (In Russian) [Google Scholar]
- Verbitsky, V.R.; Verbitsky, I.V.; Vasilyeva, O.; Savanin, V.V. Hydr. State Geological Map of the Russian Federation; Scale 1:1,000,000 (Third Generation); Central European Series; Sheets O-35—Pskov, (N-35), O-36—St. Petersburg. Explanatory Letter; Cartographic Factory VSEGEI: St. Petersburg, Russia, 2012; 510p. (In Russian) [Google Scholar]
- Kolokoltsev, V.G.; Auslender, V.G.; Kovalevskaya, E.O. The modern formation of calcareous tuffs in Leningrad region. Reg. Geol. Metallog. 2005, 23, 82–93. (In Russian) [Google Scholar]
- Kolokoltsev, V.G.; Zhuravlev, A.V.; Kovalevskaya, E.O. Abiotic factors of modern fossilization of living plants in the vicinity of St. Petersburg. In Proceedings of the III International Symposium “Bioinert interactions: Life and Stone”, St. Petersburg, Russia, 26–29 June 2007; pp. 66–69. (In Russian). [Google Scholar]
- Komárek, J.; Anagnostidis, K.; Komárek, J. Chroococcales; Cyanoprokaryota/Komárek, Jiří; Unaltered Repr.; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008; ISBN 978-3-8274-2111-1. [Google Scholar]
- Komárek, J.; Anagnostidis, K.; Komárek, J. Oscillatoriales; Cyanoprokaryota/Komárek, Jiří; Unaltered Repr.; 2. Print; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008; ISBN 978-3-8274-1914-9. [Google Scholar]
- Komárek, J.; Komárek, J. Heterocytous Genera; Cyanoprokaryota/Komárek, Jiří; Springer-Spektrum: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-8274-0932-4. [Google Scholar]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae. Nor. Inst. Water Res. 1972, 11, 5. [Google Scholar]
- Rippka, R. Isolation and purification of cyanobacteria. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1988; Volume 167, pp. 3–27. ISBN 978-0-12-182068-8. [Google Scholar]
- Waterbury, J.B. The Cyanobacteria—Isolation, Purification and Identification. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 1053–1073. ISBN 978-0-387-25494-4. [Google Scholar]
- Melechin, A.; Davydov, D.; Shalygin, S.; Borovichev, E. Open information system on biodiversity cyanoprokaryotes and lichens CRIS (CRYPTOGAMIC RUSSIAN INFORMATION SYSTEM). Bull. Mosc. Soc. Nat. Biol. Ser. 2013, 118, 51–56. (In Russian) [Google Scholar]
- Melekhin, A.V.; Davydov, D.A.; Borovichev, E.A.; Shalygin, S.S.; Konstantinova, N.A. CRIS—Service for Input, Storage and Analysis of the Biodiversity Data of the Cryptogams. Folia Cryptogam. Est. 2019, 56, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Davydov, D. Peculiarity (property) geographical distribution and analysis Cyanoprokaryota (Cyanobacteria) by the example of biota Murmansk region. Bull. Mosc. Soc. Nat. Biol. Otd. Biol. 2010, 115, 43–54. (In Russian) [Google Scholar]
- Vladimirov, I.A.; Matveeva, T.V.; Lutova, L.A. Real-Time PCR to Study the Spread of Agrobacteria; Galanika: St. Petersburg, Russia, 2014; p. 67. ISBN 978-5-906555-86-1. (In Russian) [Google Scholar]
- 16S Metagenomic Sequencing Library Preparation. Available online: https://www.illumina.com/content/dam/illumina-support/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 11 August 2021).
- Blum, J.L. The Ecology of River Algae. Bot. Rev. 1956, 22, 291–341. [Google Scholar] [CrossRef]
- Fritsch, F.E. The encrusting algal communities of certain fast-flowing streams1. New Phytol. 1929, 28, 165–196. [Google Scholar] [CrossRef]
- Wu, J.; Xi, C. Evaluation of Different Methods for Extracting Extracellular DNA from the Biofilm Matrix. Appl. Environ. Microbiol. 2009, 75, 5390–5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wade, B.D.; Garcia-Pichel, F. Evaluation of DNA Extraction Methods for Molecular Analyses of Microbial Communities in Modern Calcareous Microbialites. Geomicrobiol. J. 2003, 20, 549–561. [Google Scholar] [CrossRef]
- Gómez-Acata, E.S.; Centeno, C.M.; Falcón, L.I. Methods for Extracting ’omes from Microbialites. J. Microbiol. Methods 2019, 160, 1–10. [Google Scholar] [CrossRef]
- Águila, B.; Alcántara-Hernández, R.J.; Montejano, G.; López-Martínez, R.; Falcón, L.I.; Becerra-Absalón, I. Cyanobacteria in Microbialites of Alchichica Crater Lake: A Polyphasic Characterization. Eur. J. Phycol. 2021, 1–16. [Google Scholar] [CrossRef]
- Davydov, D.A.; Patova, E. The Diversity of Cyanoprokaryota from Freshwater and Terrestrial Habitats in the Eurasian Arctic and Subarctic. In Proceedings of the 20th IAC Cyanophyte/Cyanobacteria Research Symposium 2016 Ordinal: 20th Affiliation, Innsburck, Austria, 28 August–2 September 2016. [Google Scholar] [CrossRef]
- Uher, B. Cyanophytes/Cyanobacteria in the Gorges of the National Park Slovak Paradise. Master’s Thesis, Comenius University, Bratislava, Slovakia, 2001. [Google Scholar] [CrossRef]
- Stanislavskaja, E.V.; Gorchenko, A.S. Variety of periphyton algae in tributaries of Lake Ladoga. Nov. Sist. Nizshih Rastenij 2005, 39, 89–108. (In Russian) [Google Scholar]
- García-Del-Cura, M.Á.; Sanz-Montero, M.E.; De-los-Ríos, M.A.; Ascaso, C. Microbial Dolomite in Fresh Water Carbonate Deposits. Sedimentology 2014, 61, 41–55. [Google Scholar] [CrossRef]
- Pentecost, A. Taxonomic Identity, Ecology and Distribution of the Calcite-Depositing Cyanobacterium Phormidium Incrustatum (Oscillatoriaceae). Cryptogam. Algol. 2003, 24, 307–321. [Google Scholar]
- Sharp, J.H. Blue-green algae and carbonates Schizithrix calcicola and algal stromatolites from Bermuda1: Schizithrix calcicola and algal stromatolites. Limnol. Oceanogr. 1969, 14, 568–578. [Google Scholar] [CrossRef]
- Ivanova, Z.D.; Tsyrenova, D.D.; Barkhutova, D.D. Role of cyanobacteria in mineral formation in the hot source Alla (Buryatia). Bull. Buryat State Univ. Biol. Geogr. 2018, 88, 34–41. (In Russian) [Google Scholar] [CrossRef]
- Zaitseva, L.V.; Orleanskii, V.K.; Alekseev, A.O.; Ushatinskaya, G.T.; Gerasimenko, L.M. Transformation of Carbonate Minerals in a Cyano-Bacterial Mat in the Course of Laboratory Modeling. Microbiology 2007, 76, 342–356. [Google Scholar] [CrossRef]
- Ariño, X.; Hernandez-Marine, M.; Saiz-Jimenez, C. Colonization of Roman Tombs by Calcifying Cyanobacteria. Phycologia 1997, 36, 366–373. [Google Scholar] [CrossRef]
- Samylina, O.S.; Zaitseva, L.V.; Sinetova, M.A. Participation of the algo-bacterial community in the formation of modern stromatolites in the Petukhovskoe soda lake (Altai Territory). Paleont. J. 2016, 2016, 92–101. (In Russian) [Google Scholar] [CrossRef]
- Della Porta, G.; Hoppert, M.; Hallmann, C.; Schneider, D.; Reitner, J. The Influence of Microbial Mats on Travertine Precipitation in Active Hydrothermal Systems (Central Italy). Depositional Rec. 2021, dep2.147. [Google Scholar] [CrossRef]
- De Boever, E.; Brasier, A.T.; Foubert, A.; Kele, S. What Do We Really Know about Early Diagenesis of Non-Marine Carbonates? Sediment. Geol. 2017, 361, 25–51. [Google Scholar] [CrossRef] [Green Version]
- Cleaning the Olginsky Canal of the Water Supply System of the Fountains of Peterhof in Order to Preserve the Cultural Heritage Site; Technical Report on the Results of Environmental Engineering Surveys for the Preparation of Project Documentation; IEI: St. Petersburg, Russia, 2016; Volume 1.4, pp. 307–316, Unpublished Work. (In Russian)
- Gomez, F.J.; Mlewski, C.; Boidi, F.J.; Farías, M.E.; Gérard, E. Calcium Carbonate Precipitation in Diatom-Rich Microbial Mats: The Laguna Negra Hypersaline Lake, Catamarca, Argentina. J. Sediment. Res. 2018, 88, 727–742. [Google Scholar] [CrossRef]

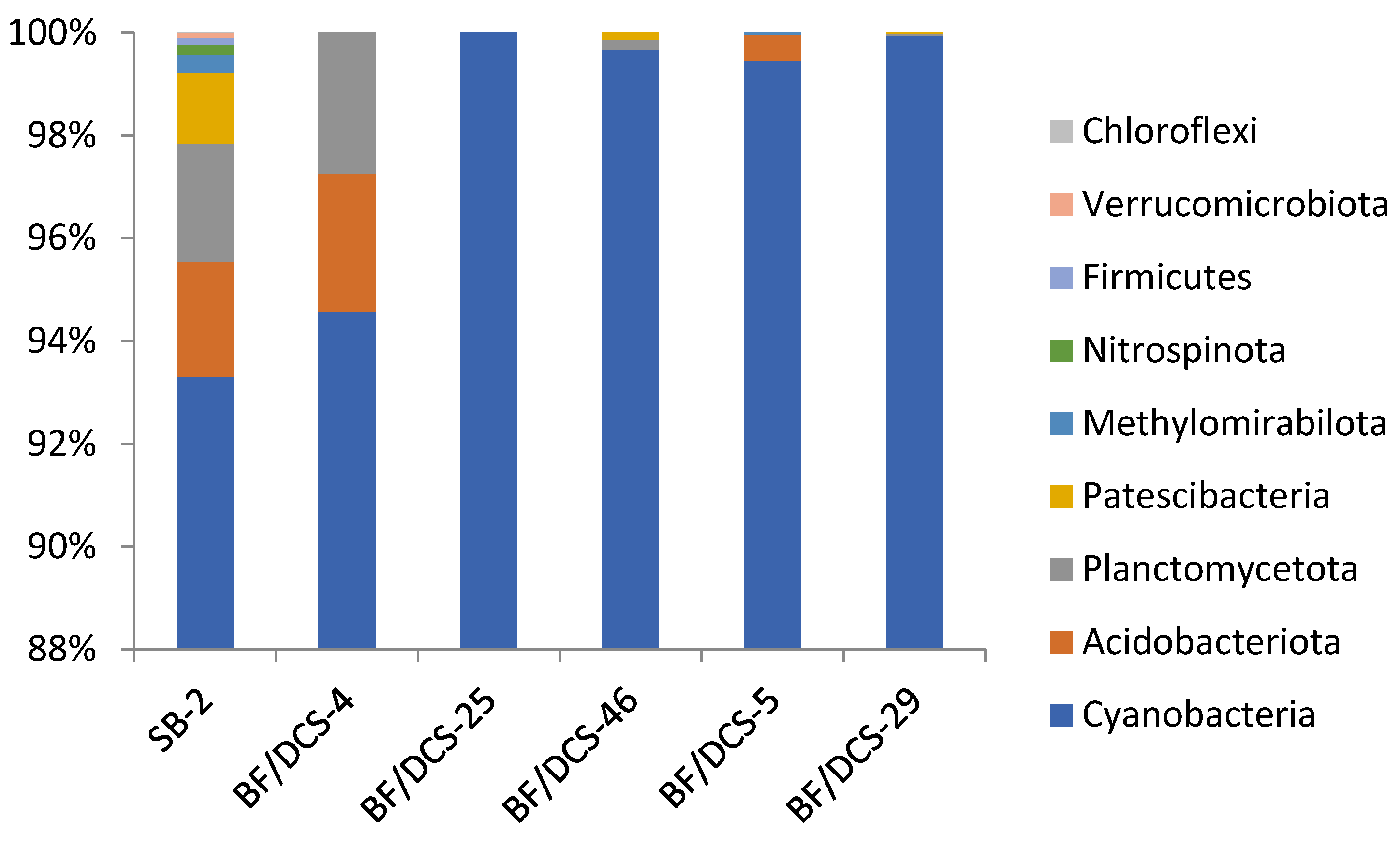






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Point | Water Bodies | Coordinates | Sample Name | Habitat Types (Water Flow) | |
|---|---|---|---|---|---|
| N | E | ||||
| 1 | Glyadinskoe reservoir | 59°42′52″ | 29°46′31″ | SB-1, SB-2 * | I(A) |
| 2 | Water discharge from the reservoir | 59°42′52″ | 29°46′31″ | SB-3, BF/DCS-4 * | III(C) |
| 3 | Splash area near water discharge | 59°42′52″ | 29°46′31″ | BF/DCS-5 * | IV(E) |
| 4 | Simonovskiy stream | 59°44′26″ | 29°45′53″ | BF-6, BF-7, BF-8, BF-9, BF-10, DCS-11, DCS-12, DCS-13, DCS-14, LCS-15, DCS-16, DCS-17, BF-18, DCS-19 | II(B) |
| 5 | Waterfall on Simonovskiy stream | 59°44′23″ | 29°45′30″ | DCS-20, DCS-21, DCS-22, DCS-23, BF/DCS-24, BF/DCS-25 * | III(C) |
| 6 | Splash area near waterfall | 59°44′23″ | 29°45′30″ | BF-26, SB/DCS-27, BF/DCS-28, BF/DCS-29 * | IV(E) |
| 7 | Bolshoye Zaborodie village (spring, stream and fire pond) | 59°44′20″ | 29°45′6″ | SB-30, SB-31, SB-32, BF-33 | V(A) |
| SB-34, DCS-35, DCS-36, DCS-37 | II(B) | ||||
| LCS-38 | VI(D) | ||||
| 8 | Spring 1 in the Maloye Zaborodie village | 59°44′41″ | 29°44′19″ | BF-39, BF-40 | V(A) |
| 9 | Spring 2 in the Maloye Zaborodie village | 59°45′11″ | 29°44′44″ | BF/DCS-41 | V(A) |
| 10 | Riverbed bend of the Fabrichnaya river | 59°44′55″ | 29°45′34″ | BF/DCS-42 | II(B) |
| 11 | Fabrichnaya river | 59°42′58″ | 29°44′29″ | BF-43, BF-44, BF/DCS-45, BF/DCS-46 * | II(B) |
| 12 | Confluence of Fabrichnaya river and Simonovskiy stream | 59°44′56″ | 29°46′27″ | BF/DCS-47 | II(B) |
| 13 | Olginsky channel | 59°52′30″ | 29°54′14″ | LCS-48, LCS-48 | VI(D) |
| 14 | Dry pond | 59°53′0″ | 29°54′3″ | LCS-50 | VI(D) |
| Samples | Carbonates | Silicates | pH |
|---|---|---|---|
| BF-6, BF-7, BF-8, BF-9, BF-10, BF/DCS-24, DCS-17, BF-18, | Calcite | Quartz | 8.0–8.86 |
| DCS-11, DCS-13, LCS-15 | Magnesian calcite | Quartz, feldspar | 8.0–8.5 |
| DCS-12, DCS-14, DCS-16 | Calcite | Quartz, feldspar | 8.0–8.5 |
| DCS-19 | Magnesian calcite | - | 8.0–8.5 |
| BF-26, SB/DCS-27, BF/DCS-28, DCS-20, DCS-22 | Calcite | Quartz | 8.0–8.5 |
| DCS-21, DCS-23 | Magnesian calcite | Quartz | 8.0–8.5 |
| SB-1 | Calcite, dolomite | Feldspar, mica, quartz | 8.0 |
| SB-3 | Calcite | Quartz | 8.0 |
| BF-43, BF-44, BF/DCS-45, BF/DCS-42 | Calcite | Quartz ± mica | 8.92 |
| DCS-41 | Calcite | - | 8.67 |
| BF/DCS-47 | Calcite | - | 8.95 |
| SB-30, SB-31, SB-32 | Calcite, dolomite | Quartz, feldspar, mica | 7.33 |
| SB-34 | Calcite | - | 8.66 |
| DCS-35, DCS-36 | Calcite | Quartz | 8.0–8.5 |
| DCS-37 | Magnesian calcite | Quartz | 8.0–8.5 |
| LCS-38 | Magnesian calcite, aragonite | Quartz | 8.0–8.5 |
| LCS-48 | Magnesian calcite, aragonite | Quartz | 8.0 |
| LCS-49 | Magnesian calcite, aragonite | Quartz, feldspar | 9.0 |
| LCS-50 | Magnesian calcite, aragonite | Quartz | 8.5 |
| SB-2, BF/DCS-4, BF/DCS-5 | Calcite | Quartz, feldspar, mica | 8.0 |
| BF/DCS-25, BF/DCS-29 | Calcite | Quartz, feldspar, mica | 8.1 |
| BF/DCS-46 | Calcite | Quartz, feldspar, mica | 8.2 |
| Species | Number of Sample, % | |||||
|---|---|---|---|---|---|---|
| SB-2 | BF/T-4 | BF/T-25 | BF/T-46 | BF/T-5 | BF/T-29 | |
| Unclassified | 51.9 | 50.1 | 51.4 | 57.3 | 26.4 | 27.4 |
| Phormidium_CYN64 | 3.7 | 1.3 | 3.1 | 8.4 | 47.6 | 44.9 |
| Tychonema_CCAP_1459-11B | 22.8 | 27.6 | 11.4 | |||
| Chamaesiphon_PCC-7430 | 16.5 | 2.8 | 7.0 | 4.1 | 7.0 | 6.8 |
| Leptolyngbya_ANT.L52.2 | 15.3 | 0.3 | 0.4 | 11.0 | 12.4 | |
| Calothrix_KVSF5 | 20.5 | 5.7 | 3.2 | 6.2 | ||
| Synechococcus_PCC-7502 | 3.4 | ≤0.01 | 0.5 | 5.0 | 4.9 | 5.5 |
| Leptolyngbya_FYG | ≤0.1 | 1.3 | 4.8 | 3.7 | 0.5 | 0.6 |
| Calothrix_PCC-6303 | ≤0.1 | 1.4 | 2.0 | 1.7 | 1.5 | |
| Pseudanabaena_PCC-7429 | 2.4 | ≤0.1 | 0.0 | |||
| Phormidesmis_ANT.LACV5.1 | ≤0.01 | 0.4 | 1.3 | ≤0.1 | 0.2 | |
| Leptolyngbya_ANT.L67.1 | ≤0.1 | 0.6 | 0.6 | |||
| Geitlerinema_LD9 | ≤0.01 | 0.7 | ≤0.1 | |||
| Cyanobium_PCC-6307 | 0.6 | ≤0.1 | ||||
| Pseudanabaena_PCC-6802 | 0.3 | ≤0.01 | ||||
| Nodosilinea_PCC-7104 | ≤0.1 | |||||
| Oscillatoria_SAG_1459-8 | ≤0.1 | |||||
| Snowella_0TU37S04 | ≤0.01 | |||||
| Order | Families | Genus | Species |
|---|---|---|---|
| Chroococcales | Chroococcaceae | Chroococcus | Chroococcus sp. |
| Gloeocapsopsis | Gloeocapsopsis crepidinum (Thuret) Geitler ex Komárek | ||
| Gloeocapsa | Gloeocapsa calcarea Tilden | ||
| Gloeocapsa punctata Näg. ampl. Hollerb. | |||
| Gloeocapsa sp. | |||
| Gloeocapsa atrata Kützing, nom. illeg. | |||
| Microcystaceae | Microcystis | Microcystis pulverea (H.C.Wood) Forti | |
| Nostocales | Nostocaceae | Nostoc | Nostoc punctiforme Hariot |
| Nostoc sp. | |||
| Microchaetaceae | Leptobasis | Leptobasis sp. | |
| Oscillatoriales | Coleofasciculaceae | Geitlerinema * | Geitlerinema sp. |
| Geitlerinema splendidum (Greville ex Gomont) Anagnostidis | |||
| Microcoleaceae | Kamptonema | Kamptonema jasorvense (Vouk) Strunecký, Komárek & J.Smarda | |
| Microcoleus | Microcoleus autumnalis (Gomont) Strunecky, Komárek & J.R.Johansen | ||
| Microcoleus vaginatus Gomont ex Gomont | |||
| Oscillatoriaceae | Oscillatoria * | Oscillatoria tenuis C.Agardh ex Gomont | |
| Lyngbya * | Lyngbya spp. | ||
| Phormidium * | Phormidium animale (C.Agardh ex Gomont) Anagnostidis & Komárek | ||
| Phormidium breve (Kützing ex Gomont) Anagnostidis & Komárek | |||
| Phormidium spp. | |||
| Synechococcales | Schizotrichaceae | Schizothrix | Schizothrix sp. |
| Leptolyngbyaceae | Leptolyngbya * | Leptolyngbya laminosa (Gomont ex Gomont) Anagnostidis & Komárek | |
| Leptolyngbya spp. | |||
| Merismopediaceae | Eucapsis | Eucapsis minor (Skuja) Elenkin | |
| Pseudanabaenaceae | Pseudanabaena * | Pseudanabaena sp. |
| Species | Studied Habitats, Frequency | Carbonate Sediments | ||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | Mg-Cc, Ara | Cc | |
| Chroococcus sp. | 1 | + | ||||||
| Eucapsis minor | 1 | + | ||||||
| Geitlelinema sp. | 2 | 1 | + | |||||
| Geitlelinema splendidum | 1 | + | ||||||
| Gloeocapsa atrata | 1 | + | ||||||
| Gloeocapsa calcarea | 1 | + | ||||||
| Gloeocapsa punctata | 1 | + | ||||||
| Gloeocapsa sp. | 1 | 1 | + | |||||
| Gloeocapsopsis crepidinum | 1 | + | ||||||
| Kamptonema jasorvense | 1 | + | ||||||
| Leptobasis sp. | 1 | + | ||||||
| Leptolyngbya laminosa | 1 | + | ||||||
| Leptolyngbya spp. | 1 | 7 | 1 | 2 | 3 | 1 | + | + |
| Lyngbya spp. | 2 | 1 | + | |||||
| Microcoleus autumnalis | 1 | + | ||||||
| Microcoleus vaginatus | 1 | + | ||||||
| Microcystis pulverea | 1 | + | ||||||
| Nostoc punctiforme | 1 | + | ||||||
| Nostoc sp. | 2 | 1 | + | |||||
| Oscillatoria tenuis | 1 | + | ||||||
| Phormidium animale | 1 | + | ||||||
| Phormidium breve | 1 | + | ||||||
| Phormidium spp. | 1 | 1 | 3 | 1 | + | + | ||
| Pseudanabaena sp. | 1 | 1 | 1 | + | ||||
| Schizothrix sp. | 2 | + | ||||||
| Number of species | 2 | 10 | 5 | 2 | 5 | 15 | 15 | 12 |
| Cyanobacteria | Environmental Conditions | Mineral | Ref. |
|---|---|---|---|
| In vitro | |||
| Anabaena, Calothrix, Phormidium, Gloeocapsa | Z8 + MgSO4 and CaCl2 | Cc | [50] |
| Microcoleus chthonoplastes | nutrient medium “S” + carbonate sediment 65% MgCO3 + 35% CaCO3 with monohydrocalcite | Ara, Mg-Cc, Dol, Dyp | [51] |
| Scytonema julianum | BG11+Agar+powedered calcium carbonate | Cc, Acc | [52] |
| Biofilm with domination of the cyanobacteria Phormidium spp., Geitlelinema sp., Pseudanabaena sp. | Nature water in experiment | Cc | This study |
| 13 strains of cyanobacteria from CALU | Gromov-6 mineral medium + piece of marble (source of calcium ions) | Cc | This study |
| Biofilm with domination of the cyanobacteria Lyngbya sp., Phormidium sp., Leptolyngbya sp. With diatoms | Nature water in experiment | Cc | This study |
| In situ | |||
| Geitlerinema sp. and Nodosilinea sp. | Soda lake | CC, Ara, Mg-Cc, Dol, | [53] |
| Gloeotrichia sp. | Freshwater lakes | Wh, Cc | [11] |
| Rivularia sp. | A brackish technogenically altered watercourse (Bogodelsky stream) | CC | [11] |
| Phormidium incrustatum | Freshwater rivers Hoyoux and Triffoy, Belgium | Cc | [6] |
| Phormidiaceae and Oscillatoriaceae filaments (inc. Tychonema) | The stream (tributary of the Mérriver) to the Southwest of Paris | Cc | [7,20] |
| Spirulina, Phormidium | active hydrothermal systems (Central Italy) | Cc | [54] |
| Shizothrix sp. | Freshwater pond (Dry pond) | Mag-Cc, Ara | This study |
| Cyanobacteria community (inc. Shizothrix sp.) with Phormidiaceae domination | Olginsky channel | Mag-Cc, Ara | This study |
| Community with Phormidium sp., Leptolyngbya sp., and other identified cyanobacteria With diatoms and green algae | Freshwater springs | Cc | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodina, O.A.; Vereshchagin, O.S.; Vlasov, D.Y.; Zelenskaya, M.S.; Pankin, D.V.; Mitrofanov, N.V.; Nikitin, M.Y.; Vasileva, K.Y.; Frank-Kamenetskaya, O.V. Cyanobacterial Communities of Carbonate Sediments and Biomineralization in Peterhof Fountains’ Water Supply System, Russia. Minerals 2021, 11, 1199. https://doi.org/10.3390/min11111199
Rodina OA, Vereshchagin OS, Vlasov DY, Zelenskaya MS, Pankin DV, Mitrofanov NV, Nikitin MY, Vasileva KY, Frank-Kamenetskaya OV. Cyanobacterial Communities of Carbonate Sediments and Biomineralization in Peterhof Fountains’ Water Supply System, Russia. Minerals. 2021; 11(11):1199. https://doi.org/10.3390/min11111199
Chicago/Turabian StyleRodina, Oksana A., Oleg S. Vereshchagin, Dmitry Yu. Vlasov, Marina S. Zelenskaya, Dmitrii V. Pankin, Nikita V. Mitrofanov, Michael Yu. Nikitin, Kseniia Yu. Vasileva, and Olga V. Frank-Kamenetskaya. 2021. "Cyanobacterial Communities of Carbonate Sediments and Biomineralization in Peterhof Fountains’ Water Supply System, Russia" Minerals 11, no. 11: 1199. https://doi.org/10.3390/min11111199