Microbial Reducibility of Fe(III) Phases Associated with the Genesis of Iron Ore Caves in the Iron Quadrangle, Minas Gerais, Brazil

Abstract
:1. Introduction

2. Results and Discussion
2.1. Characterization of Fe(III)-Containing Phases

{kind=link}
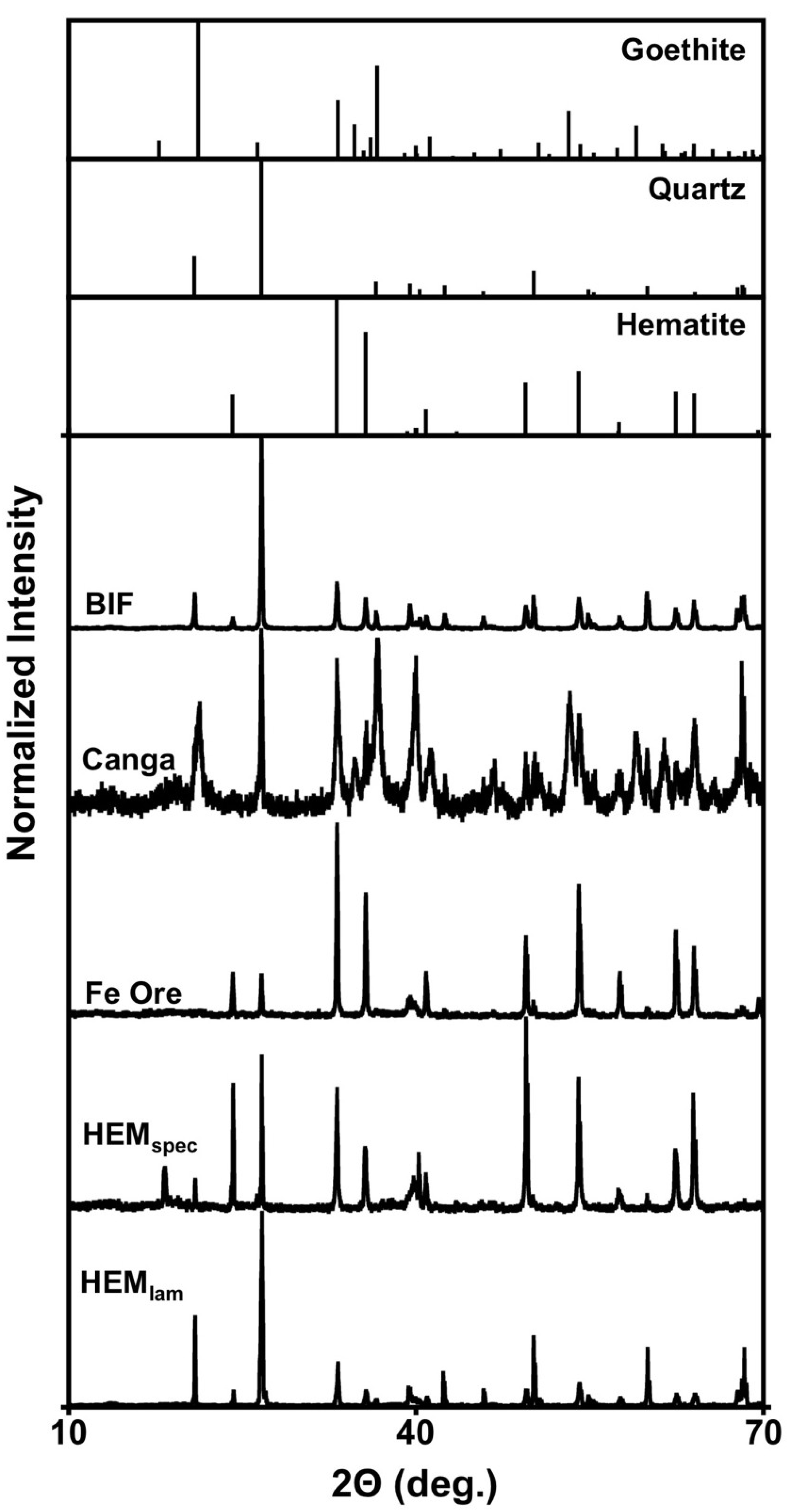{kind=link}
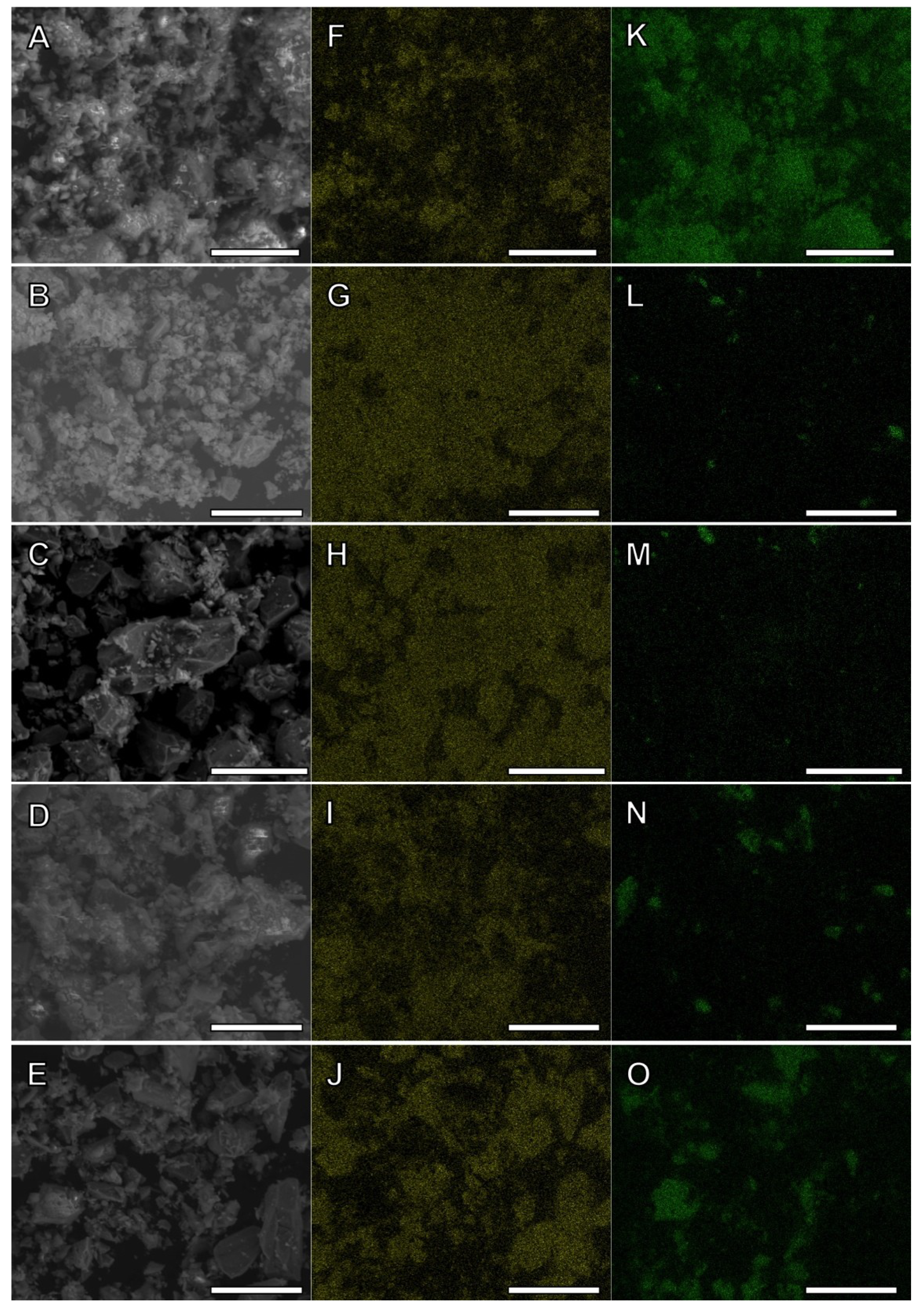{kind=link}
{kind=link}
{kind=link}
| Sample | Fe(II) (μmol/g) | Poorly-crystalline Fe(III) (μmol/g) | Total Fe (mmol/g) |
|---|---|---|---|
| BIF | 1.37 ± 0.10 | 3.22 ± 1.18 | 1.79 ± 0.17 |
| Canga | 3.30 ± 0.05 | 12.54 ± 5.32 | 6.35 ± 0.33 |
| Iron Ore | 1.77 ± 0.25 | 24.37 ± 5.28 | 6.32 ± 0.52 |
| HEMspec | 1.77 ± 1.02 | 8.14 ± 0.71 | 2.19 ± 0.61 |
| HEMlam | 1.65 ± 0.14 | 3.65 ± 1.14 | 1.69 ± 0.82 |
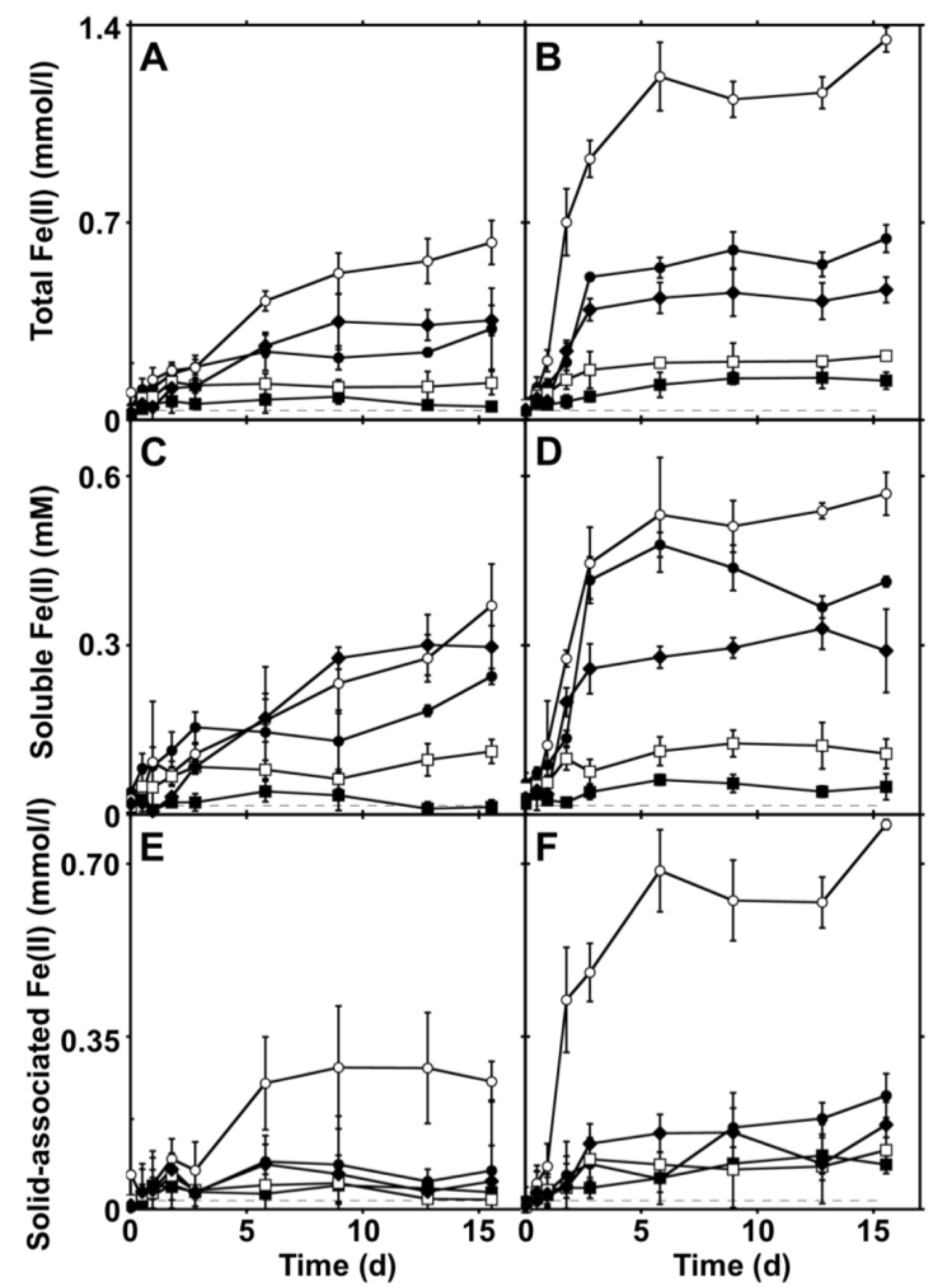
2.2. Extents and Rates of Fe(III) Bioreduction



| Sample | Fe(III) reduction k (day−1) | Total Fe(III) reduced (%) | Hydroxylamine-HCl-extractable Fe(III) reduced (%) | |||
|---|---|---|---|---|---|---|
| (−) AQDS | (+) AQDS | (−) AQDS | (+) AQDS | (−) AQDS | (+) AQDS | |
| BIF | 0.0012 | 0.0039 | 0.81 | 1.61 | 450 | 894 |
| Canga | 0.0013 | 0.0089 | 1.57 | 3.37 | 797 | 1707 |
| Iron Ore | 0.0007 | 0.0004 | 0.21 | 0.37 | 27 | 49 |
| HEMspec | 0.0008 | 0.0011 | 0.34 | 0.57 | 91 | 153 |
| HEMlam | 0.0009 | 0.0034 | 0.88 | 1.15 | 409 | 534 |
2.3. Enhancement of Fe(III) Bioreduction by an Electron Shuttling Compound
2.4. Controls on Fe(III) Bioreduction
2.5. Implications for the Role of FeRB in IOC Formation
3. Experimental Section
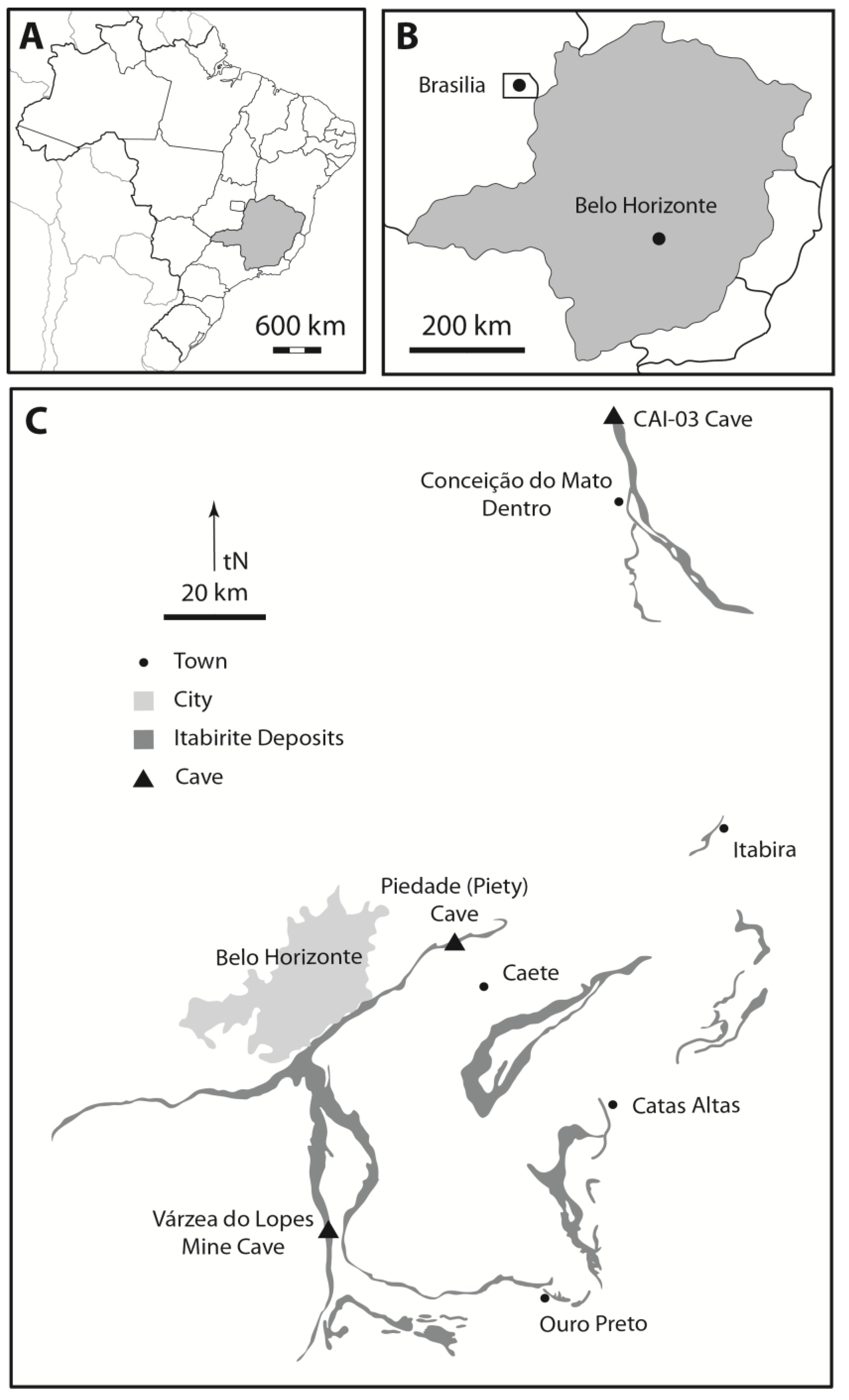
3.1. Sample Collection
3.2. Characterization of Fe Phases

3.3. Bioreduction Incubations
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Simmons, G.C. Canga caves in the Quadrilátero Ferrífero, Minas Gerais, Brazil. Bull. Nat. Speleol. Soc. 1963, 25, 66–72. [Google Scholar]
- Auler, A.; Farrant, A.R. A brief introduction to karst and caves in Brazil. Proc. Univ. Bristol Spelaeol. Soc. 1996, 20, 187–200. [Google Scholar]
- Bekker, A.; Slack, J.F.; Planavsky, N.; Krapež, B.; Hoffmann, A.; Konhauser, K.O.; Rouxel, O.J. Iron formation: The sedimentary product of a complex interplay among mantle, tectonic, oceanic, and biospheric processes. Econ. Geol. 2010, 105, 467–508. [Google Scholar] [CrossRef]
- Morris, R.C.; Thornber, M.R.; Ewer, W.E. Deep-seated iron ores from banded-iron formation. Nature 1980, 288, 250–252. [Google Scholar] [CrossRef]
- Morris, R.C. A textural and mineralogical study of the relationship between iron ore to banded-iron formation in the Hamersley iron province of Western Australia. Econ. Geol. 1980, 75, 184–209. [Google Scholar] [CrossRef]
- Varajão, C.A.C.; Bruand, A.; Ramanaidou, E.R.; Gilkes, R.J. Microporosity of BIF hosted hematite ore, Iron Quadrangle, Brazil. An. Acad. Bras. Ciênc. 2002, 74, 113–126. [Google Scholar] [CrossRef]
- Harder, E.C. The “itabirite” iron ores of Brazil. Econ. Geol. 1914, 9, 101–111. [Google Scholar] [CrossRef]
- Schuster, D.L.; Farley, K.A.; Vasconcelos, P.M.; Balco, G.; Monteiro, H.S.; Waltenberg, K.; Stone, J.O. Cosmogenic 3He in hematite and goethite from Brazilian “canga” duricrust demonstrates the extreme stability of these surfaces. Earth Planet. Sci. Lett. 2012, 329–330, 41–50. [Google Scholar] [CrossRef]
- Rosière, C.A.; Siemes, H.; Quade, H.; Brokmeier, H.-G.; Jansen, E.M. Microstructures, textures, and deformation mechanisms in hematite. J. Struct. Geol. 2001, 23, 1429–1440. [Google Scholar] [CrossRef]
- Guedes, S.C.; Rosière, C.A.; Barley, M.E. The association of carbonate alteration of banded iron formation with the Carajás high-grade hematite deposits. Appl. Earth Sci. 2003, 112, 26–30. [Google Scholar] [CrossRef]
- Salgado, A.A.R.; Braucher, R.; Colin, F.; Nalini, H.A.; Varajão, A.F.D.C.; Varajão, C.A.C. Denudation rates of the Quadrilátero Ferrífero (Minas Gerais, Brazil): Preliminary results from measurements of solute fluxes in rivers and in situ-produced cosmogenic 10Be. J. Geochem. Explor. 2006, 88, 313–317. [Google Scholar] [CrossRef]
- Salgado, A.A.R.; Braucher, R.; Varajão, A.C.; Colin, F.; Varajão, A.F.D.C.; Nalini, H.A. Relief evolution of the Quadrilátero Ferrífero (Minas Gerais, Brazil) by means of (10Be) cosmogenic nuclei. Z. Geomorphol. 2008, 52, 317–323. [Google Scholar] [CrossRef]
- Parker, C.W.; Auler, A.S.; Senko, J.M.; Sasowsky, I.D.; Piló, L.B.; Smith, M.; Johnston, M.; Barton, H.A. Microbial Iron Cycling and Biospeleogenesis: Cave Development in the Carajás Formation, Brazil. In Proceedings of the 16th International Congress of Speleology, Brno, Czech Republic, 21–27 July 2013; Filippi, M., Bosák, P., Eds.; Czech Speleological Society: Praha, Czech Republic, 2013; pp. 442–446. [Google Scholar]
- Lovley, D.R.; Holmes, D.E.; Nevin, K.P. Dissimilatory Fe(III) and Mn(IV) reduction. Adv. Microb. Physiol. 2004, 49, 219–286. [Google Scholar] [CrossRef]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752–764. [Google Scholar] [CrossRef]
- McFarlane, M.J.; Twidale, C.R. Karstic features associated with tropical weathering profiles. Z. Geomorphol. Suppl. 1987, 64, 73–95. [Google Scholar]
- Barton, H.A.; Luiszer, F. Microbial metabolic structure in a sulfidic cave hot spring: Potential mechanisms of biospeleogenesis. J. Cave Karst Stud. 2005, 67, 28–38. [Google Scholar]
- Engel, A.S. Microbial Diversity of Cave Ecosystems. In Geomicrobiology: Molecular and Environmental Perspectives; Barton, L.L., Mandl, M., Loy, A., Eds.; Springer: New York, NY, USA, 2010; pp. 219–238. [Google Scholar]
- Barton, H.A. Biospeleogenesis. In Treatise on Geomorphology; Shroder, J.F., Ed.; Academic Press: San Diego, CA, USA, 2013; Volume 6, pp. 39–55. [Google Scholar]
- Spier, C.A.; Barros de Oliveira, S.M.; Rosière, C.A.; Ardisson, J.D. Mineralogy and trace-element geochemistry of the high-grade iron ores of the Águas Claras Mine and comparison with the Capão Xavier and Tamaduá iron ore deposits, Quadrilátero Ferrífero, Brazil. Miner. Deposita 2008, 43, 229–254. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Rapid assay for microbially reducible ferric iron in aquatic sediments. Appl. Environ. Microbiol. 1987, 53, 1536–1540. [Google Scholar]
- Downs, R.T.; Hall-Wallace, M. The American Mineralogist crystal structure database. Am. Mineral. 2003, 88, 247–250. [Google Scholar]
- Burgos, W.D.; Royer, R.A.; Fang, Y.; Yeh, G.-T.; Fisher, A.S.; Jeon, B.-H.; Dempsey, B.A. Theoretical and experimental considerations related to reaction-based modeling: A case study using iron(III) oxide bioreduction. Geomicrobiol. J. 2002, 19, 253–287. [Google Scholar] [CrossRef]
- Neal, A.L.; Rosso, K.M.; Geesy, G.G.; Gorby, Y.A.; Little, B.J. Surface structure effects on direct reduction of iron oxides by Shewanella oneidensis. Geochim. Cosmochim. Acta. 2003, 67, 4489–4503. [Google Scholar] [CrossRef]
- Bose, S.; Hochella, M.F., Jr.; Gorby, Y.A.; Kennedy, D.W.; McCready, D.E.; Madden, A.S.; Lower, B.H. Bioreduction of hematite nanoparticles by the dissimilatory iron reducing bacterium Shewanella oneidensis MR-1. Geochim. Cosmochim. Acta. 2009, 73, 962–976. [Google Scholar] [CrossRef]
- Zachara, J.M.; Fredrickson, J.K.; Li, S.-M.; Kennedy, D.W.; Smith, S.C.; Gassman, P.L. Bacterial reduction of crystalline Fe3+ oxides in single phase suspensions and subsurface materials. Am. Mineral. 1998, 83, 1426–1443. [Google Scholar]
- Royer, R.A.; Burgos, W.D.; Fisher, A.S.; Unz, R.F.; Dempsey, B.A. Enhancement of biological reduction of hematite by electron shuttling and Fe(II) complexation. Environ. Sci. Technol. 2002, 36, 1939–1946. [Google Scholar] [CrossRef]
- Royer, R.A.; Burgos, W.D.; Fisher, A.S.; Jeon, B.H.; Unz, R.F.; Dempsey, B.A. Enhancement of hematite bioreduction by natural organic matter. Environ. Sci. Technol. 2002, 36, 2897–2904. [Google Scholar] [CrossRef]
- Royer, R.A.; Dempsey, B.A.; Jeon, B.-H.; Burgos, W.D. Inhibition of biological reductive dissolution of hematite by ferrous iron. Environ. Sci. Technol. 2004, 38, 187–193. [Google Scholar] [CrossRef]
- Stone, J.J.; Royer, R.A.; Dempsey, B.A.; Burgos, W.D. Effect of natural organic matter on zinc inhibition of hematite bioreduction by Shewanella putrefaciens CN32. Environ. Sci. Technol. 2007, 41, 5284–5290. [Google Scholar] [CrossRef]
- Luan, F.; Burgos, W.D.; Xie, L.; Zhou, Q. Bioreduction of nitrobenzene, natural organic matter, and hematite by Shewanella putrefaciens CN3. Environ. Sci. Technol. 2010, 44, 184–190. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Resck, D.V.S.; da Silva, J.E. Soil organic carbon as affected by afforestation with Eucalyptus and Pinus in the Cerrado region of Brazil. For. Ecol. Manage. 2002, 166, 285–294. [Google Scholar] [CrossRef]
- Lovley, D.R.; Coates, J.D.; Blunt-Harris, E.L.; Phillips, E.J.P.; Woodward, J.C. Humic substances as electron acceptors for microbial respiration. Nature 1996, 382, 445–448. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Zachara, J.M.; Kennedy, D.W.; Dong, H.; Onstott, T.C.; Hinman, N.W.; Li, S. Biogenic iron mineralization accompanying the dissimilatory reduction of hydrous ferric oxide by a groundwater bacterium. Geochim. Cosmoshim. Acta. 1998, 62, 3239–3257. [Google Scholar] [CrossRef]
- Scott, D.T.; McKnight, D.M.; Blunt-Harris, E.L.; Kolesar, S.E.; Lovley, D.R. Quinone moieties act as electron acceptors in the reduction of humic substances by humics-reducing microorganisms. Environ. Sci. Technol. 1998, 32, 2984–2989. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, C.; Shi, Z. Micromodel investigation of transport effects on the kinetics of reductive dissolution of hematite. Environ. Sci. Technol. 2013, 47, 4131–4139. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Availability of ferric iron for microbial reduction in bottom sediments of the freshwater tidal Potomac River. Appl. Environ. Microbiol. 1986, 52, 751–757. [Google Scholar]
- Lovley, D.R.; Phillips, E.J.P. Novel mode of microbial energy metabolism: organic carbon oxidation coupled to dissimilatory reduction of iron or manganese. Appl. Environ. Microbiol. 1988, 54, 1472–1480. [Google Scholar]
- Dominik, P.; Kaupenjohann, M. Reduction of Fe(III) (hydr)oxides with known thermodynamic stability by Geobacter metallireducens. Geomicrobiol. J. 2004, 21, 287–295. [Google Scholar] [CrossRef]
- Roden, E.E.; Zachara, J.M. Microbial reduction of crystalline iron(III) oxides: Influence of oxide surface area and potential for cell growth. Environ. Sci. Technol. 1996, 30, 1618–1628. [Google Scholar] [CrossRef]
- Kukkadapu, R.K.; Zachara, J.M.; Smith, S.C.; Fredrickson, J.K.; Liu, C. Dissimilatory bacterial reduction of Al-substituted goethite in subsurface sediments. Geochim. Cosmochim. Acta. 2001, 65, 2913–2924. [Google Scholar] [CrossRef]
- Dominik, P.; Pohl, H.N.; Bousserrhine, N.; Berthelin, J.; Kauperjohann, M. Limitations to the reductive dissolution of Al-substituted goethites by Clostridium butyricum. Soil Biol. Biochem. 2002, 34, 1147–1155. [Google Scholar] [CrossRef]
- Ekstrom, E.B.; Learman, D.R.; Madden, A.S.; Hansel, C.M. Contrasting effects of Al substitutions on microbial reduction of Fe(III) (hydr)oxides. Geochim. Cosmochim. Acta. 2010, 74, 7086–7099. [Google Scholar] [CrossRef]
- Urrutia, M.M.; Roden, E.E.; Zachara, J.M. Influence of aqueous and solid-phase Fe(II) complexants on microbial reduction of crystalline iron(III) oxides. Environ. Sci. Technol. 1999, 33, 4022–4028. [Google Scholar] [CrossRef]
- Sasowsky, I.D.; Foos, A.; Miller, C.M. Lithic controls on the removal of iron and remediation of acidic mine drainage. Water Res. 2000, 34, 2742–2746. [Google Scholar] [CrossRef]
- Piló, L.B.; Auler, A. Geoespeleologia das Cavernas em Rochas Ferríferas da Região de Carajás, Pa. Sociedade Brasileira de Espeleologia (in Portuguese). In Proceedings of the Anais do 30th Congresso Brasileiro de Espeleologia, Montes Claros, MG, Brazil, 9–12 July 2009.
- Minyard, M.L.; Burgos, W.D. Hydrologic flow controls on biologic iron(III) reduction in natural sediments. Environ. Sci. Technol. 2007, 41, 1218–1224. [Google Scholar] [CrossRef]
- Roden, E.E.; Urrutia, M.M. Ferrous iron removal promotes microbial reduction of crystalline iron(III) oxides. Environ. Sci. Technnol. 1999, 33, 1847–1853. [Google Scholar] [CrossRef]
- Gonzalez-Gil, G.; Amonette, J.E.; Romine, M.F.; Gorby, Y.A.; Geesy, G.G. Bioreduction of natural specular hematite under flow conditions. Geochim. Cosmochim. Acta. 2005, 69, 1145–1155. [Google Scholar] [CrossRef]
- Roden, E.E.; Urrutia, M.M.; Mann, C.J. Bacterial reductive dissolution of crystalline Fe(III) oxide in continuous-flow column reactors. Appl. Environ. Microbiol. 2000, 66, 1062–1065. [Google Scholar] [CrossRef]
- Stookey, L.L. Ferrozine—A new spectrophotometric reagent for iron. Anal. Chem. 1970, 42, 779–781. [Google Scholar] [CrossRef]
- Tanner, R.S. Cultivation of Bacteria and Fungi. In Manual of Environmental Microbiology; Hurst, C.J., Knudsen, G.R., McInerney, M.J., Stetzenbach, L.D., Walter, M.V., Eds.; ASM Press: Washington, DC, USA, 1997; pp. 52–60. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Parker, C.W.; Wolf, J.A.; Auler, A.S.; Barton, H.A.; Senko, J.M. Microbial Reducibility of Fe(III) Phases Associated with the Genesis of Iron Ore Caves in the Iron Quadrangle, Minas Gerais, Brazil. Minerals 2013, 3, 395-411. https://doi.org/10.3390/min3040395
Parker CW, Wolf JA, Auler AS, Barton HA, Senko JM. Microbial Reducibility of Fe(III) Phases Associated with the Genesis of Iron Ore Caves in the Iron Quadrangle, Minas Gerais, Brazil. Minerals. 2013; 3(4):395-411. https://doi.org/10.3390/min3040395
Chicago/Turabian StyleParker, Ceth W., Julie A. Wolf, Augusto S. Auler, Hazel A. Barton, and John M. Senko. 2013. "Microbial Reducibility of Fe(III) Phases Associated with the Genesis of Iron Ore Caves in the Iron Quadrangle, Minas Gerais, Brazil" Minerals 3, no. 4: 395-411. https://doi.org/10.3390/min3040395